
Do Pomegranate Hydrolyzable Tannins and Their Derived Metabolites Provide Relief in Osteoarthritis? Findings from a Scoping Review



Abstract
:
1. Introduction
2. Results and Discussion
2.1. Randomized Clinical Trials
2.2. In Vivo OA Studies
2.2.1. Expression of Chondrogenic Genes
2.2.2. Protein Expression and Inflammatory Biomarkers Level
2.2.3. OA Scoring Systems
2.3. In Vitro and Ex Vivo Studies
3. Materials and Methods
3.1. Data Sources and Search Strategy
3.2. Eligibility Criteria
3.3. Study Selection and Data Extraction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buckwalter, J.A.; Martin, J.A. Osteoarthritis. Adv. Drug Deliv. Rev. 2006, 58, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.D.; Dieppe, P.; Radin, E. Etiopathogenesis of osteoarthritis. Med. Clin. N. Am. 2009, 93, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Bloom, D.E.; Luca, D.L. The global demography of aging. In Handbook of the Economics of Population Aging; Piggott, J., Woodland, A., Eds.; North-Holland: Amsterdam, The Netherlands, 2016; Volume 1, pp. 3–56. [Google Scholar]
- Kloppenburg, M.; Berenbaum, F. Osteoarthritis year in review 2019: Epidemiology and therapy. Osteoarthr. Cartil. 2020, 28, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Cui, A.; Li, H.; Wang, D.; Zhong, J.; Chen, Y.; Lu, H. Global, regional prevalence, incidence and risk factors of knee osteoarthritis in population-based studies. EClinicalMedicine 2020, 29-30, 100587. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Cibere, J.; Goldsmith, C.H.; Anis, A.H.; Kopec, J.A. Osteoarthritis incidence and trends in administrative health records from British Columbia, Canada. J. Rheumatol. 2014, 41, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Jordan, K.P.; Bedson, J.; Englund, M.; Blyth, F.; Turkiewicz, A.; Prieto-Alhambra, D.; Peat, G. Population trends in the incidence and initial management of osteoarthritis: Age-period-cohort analysis of the Clinical Practice Research Datalink, 1992–2013. Rheumatology 2017, 56, 1902–1917. [Google Scholar] [CrossRef] [Green Version]
- Grässel, S.; Muschter, D. Recent advances in the treatment of osteoarthritis [version 1; peer review: 3 approved]. F1000Res 2020, 9, 325. [Google Scholar] [CrossRef]
- Zhang, W.; Ouyang, H.; Dass, C.R.; Xu, J. Current research on pharmacologic and regenerative therapies for osteoarthritis. Bone Res. 2016, 4, 15040. [Google Scholar] [CrossRef]
- Altman, R.D. Practical considerations for the pharmacologic management of osteoarthritis. Am. J. Manag. Care 2009, 15, S236–S243. [Google Scholar]
- Huang, W.N.; Tso, T.K. Etoricoxib improves osteoarthritis pain relief, joint function, and quality of life in the extreme elderly. Bosn. J. Basic Med. Sci. 2018, 18, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.F.; Jobanputra, P.; Barton, P.; Bryan, S.; Fry-Smith, A.; Harris, G.; Taylor, R.S. Cyclooxygenase-2 selective non-steroidal anti-inflammatory drugs (etodolac, meloxicam, celecoxib, rofecoxib, etoricoxib, valdecoxib and lumiracoxib) for osteoarthritis and rheumatoid arthritis: A systematic review and economic evaluation. Health Technol. Assess. 2008, 12, 1–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.E.; Allen, K.D.; Golightly, Y.M.; Goode, A.P.; Jordan, J.M. A systematic review of recommendations and guidelines for the management of osteoarthritis: The chronic osteoarthritis management initiative of the U.S. bone and joint initiative. Semin. Arthritis Rheum. 2014, 43, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Lefèvre-Colau, M.M.; Poiraudeau, S.; Rannou, F. Evidence and recommendations for use of intra-articular injections for knee osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.A.; Togashi, R.; Wilson, M.L.; Heckmann, N.; Vangsness, C.T., Jr. Intra-articular treatment options for knee osteoarthritis. Nat. Rev. Rheumatol. 2019, 15, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, N.; Campbell, J.; Robinson, V.; Gee, T.; Bourne, R.; Wells, G. Viscosupplementation for the treatment of osteoarthritis of the knee. Cochrane Database Syst. Rev. 2006, 2, CD005321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gennero, L.; Denysenko, T.; Calisti, G.F.; Vercelli, A.; Vercelli, C.M.; Amedeo, S.; Mioletti, S.; Parino, E.; Montanaro, M.; Melcarne, A.; et al. Protective effects of polydeoxyribonucleotides on cartilage degradation in experimental cultures. Cell Biochem. Funct. 2013, 31, 214–227. [Google Scholar] [CrossRef]
- Dallari, D.; Sabbioni, G.; Del Piccolo, N.; Carubbi, C.; Veronesi, F.; Torricelli, P.; Fini, M. Efficacy of intra-articular polynucleotides associated with hyaluronic acid versus hyaluronic acid alone in the treatment of knee osteoarthritis: A randomized, double-blind, controlled clinical trial. Clin. J. Sport Med. 2020, 30, 1–7. [Google Scholar] [CrossRef]
- Stagni, C.; Rocchi, M.; Mazzotta, A.; Del Piccolo, N.; Rani, N.; Govoni, M.; Vivarelli, L.; Veronesi, F.; Fini, M.; Dallari, D. Randomised, double-blind comparison of a fixed co-formulation of intra-articular polynucleotides and hyaluronic acid versus hyaluronic acid alone in the treatment of knee osteoarthritis: Two-year follow-up. BMC Musculoskelet Disord. 2021, 22, 773. [Google Scholar] [CrossRef]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed. Pharm. 2020, 129, 110452. [Google Scholar] [CrossRef]
- Rübenhagen, R.; Schüttrumpf, J.P.; Stürmer, K.M.; Frosch, K.H. Interleukin-7 levels in synovial fluid increase with age and MMP-1 levels decrease with progression of osteoarthritis. Acta Orthop. 2012, 83, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Greene, M.A.; Loeser, R.F. Aging-related inflammation in osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1966–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, T.M.; Scanzello, C.R. Innate inflammation and synovial macrophages in osteoarthritis pathophysiology. Clin. Exp. Rheumatol. 2019, 37, S57–S63. [Google Scholar]
- Khan, N.M.; Haseeb, A.; Ansari, M.Y.; Devarapalli, P.; Haynie, S.; Haqqi, T.M. Wogonin, a plant derived small molecule, exerts potent anti-inflammatory and chondroprotective effects through the activation of ROS/ERK/Nrf2 signaling pathways in human osteoarthritis chondrocytes. Free Radic. Biol. Med. 2017, 106, 288–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Hebert, J.R.; Shivappa, N.; Guo, J.; Tao, K.; Zeng, C.; Lei, G.; Lin, J.; Zhang, Y. Inflammatory potential of diet and risk of incident knee osteoarthritis: A prospective cohort study. Arthritis Res. 2020, 22, 209. [Google Scholar] [CrossRef]
- Mena, P.; Calani, L.; Bruni, R.; Del Rio, D. Bioactivation of high-molecular-weight polyphenols by the gut microbiome. In Diet-Microbe Interactions in the Gut; Tuohy, K., Del Rio, D., Eds.; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]
- Yoshida, T.; Hatano, T.; Ito, H. High molecular weight plant poplyphenols (tannins): Prospective functions. Recent Adv. Phytochem. 2005, 39, 163–190. [Google Scholar]
- Bar-Ya’akov, I.; Tian, L.; Amir, R.; Holland, D. Primary metabolites, anthocyanins, and hydrolyzable tannins in the pomegranate fruit. Front. Plant. Sci. 2019, 10, 620. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A.K.; Piwowarski, J.P. Ellagitannins, gallotannins and their metabolites- the contribution to the anti-inflammatory effect of food products and medicinal plants. Curr. Med. Chem. 2018, 25, 4946–4967. [Google Scholar] [CrossRef]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharm. 2017, 174, 1244–1262. [Google Scholar] [CrossRef] [Green Version]
- Clifford, M.N.; Scalbert, A. Ellagitannins—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1118–1125. [Google Scholar] [CrossRef]
- Danesi, F.; Ferguson, L.R. Could pomegranate juice help in the control of inflammatory diseases? Nutrients 2017, 9, 958. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Banerjee, N.; Ivanov, I.; Pfent, C.M.; Prudhomme, K.R.; Bisson, W.H.; Dashwood, R.H.; Talcott, S.T.; Mertens-Talcott, S.U. Comparison of anti-inflammatory mechanisms of mango (Mangifera Indica L.) and pomegranate (Punica Granatum L.) in a preclinical model of colitis. Mol. Nutr. Food Res. 2016, 60, 1912–1923. [Google Scholar] [CrossRef] [Green Version]
- Battino, M.; Giampieri, F.; Cianciosi, D.; Ansary, J.; Chen, X.; Zhang, D.; Gil, E.; Forbes-Hernández, T. The roles of strawberry and honey phytochemicals on human health: A possible clue on the molecular mechanisms involved in the prevention of oxidative stress and inflammation. Phytomedicine 2021, 86, 153170. [Google Scholar] [CrossRef]
- López Mantecón, A.M.; Garrido, G.; Delgado-Hernández, R.; Garrido-Suárez, B.B. Combination of Mangifera indica L. extract supplementation plus methotrexate in rheumatoid arthritis patients: A pilot study. Phytother. Res. 2014, 28, 1163–1172. [Google Scholar] [CrossRef]
- Kim, H.; Venancio, V.P.; Fang, C.; Dupont, A.W.; Talcott, S.T.; Mertens-Talcott, S.U. Mango (Mangifera indica L.) polyphenols reduce IL-8, GRO, and GM-SCF plasma levels and increase Lactobacillus species in a pilot study in patients with inflammatory bowel disease. Nutr. Res. 2020, 75, 85–94. [Google Scholar] [CrossRef]
- Shahrzad, S.; Bitsch, I. Determination of gallic acid and its metabolites in human plasma and urine by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1998, 705, 87–95. [Google Scholar] [CrossRef]
- Shahrzad, S.; Aoyagi, K.; Winter, A.; Koyama, A.; Bitsch, I. Pharmacokinetics of gallic acid and its relative bioavailability from tea in healthy humans. J. Nutr. 2001, 131, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Schell, J.; Scofield, R.H. Dietary fruits and arthritis. Food Funct. 2018, 9, 70–77. [Google Scholar] [CrossRef]
- Green, J.A.; Hirst-Jones, K.L.; Davidson, R.K.; Jupp, O.; Bao, Y.; MacGregor, A.J.; Donell, S.T.; Cassidy, A.; Clark, I.M. The potential for dietary factors to prevent or treat osteoarthritis. Proc. Nutr. Soc. 2014, 73, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Ghoochani, N.; Karandish, M.; Mowla, K.; Haghighizadeh, M.H.; Jalali, M.T. The effect of pomegranate juice on clinical signs, matrix metalloproteinases and antioxidant status in patients with knee osteoarthritis. J. Sci. Food Agric. 2016, 96, 4377–4381. [Google Scholar] [CrossRef]
- Rafraf, M.; Hemmati, S.; Jafarabadi, M.A.; Moghaddam, A.; Haghighian, M.K. Pomegranate (Punica Granatum L.) peel hydroalcoholic extract supplementation reduces pain and improves clinical symptoms of knee osteoarthritis: A randomized double-blind placebo controlled study. Iran. Red Crescent Med. J. 2017, 19, e38577. [Google Scholar] [CrossRef]
- Haghighian, M.K.; Rafraf, M.; Hemmati, S.; Haghravan, S.; Asghari-Jafarabadi, M. Effects of pomegranate (Punica granatum L.) peel extract supplementation on serum lipid profile and oxidative stress in obese women with knee osteoarthritis: A double blind, randomized, placebo controlled study. Adv. Integr. Med. 2021, 8, 107–113. [Google Scholar] [CrossRef]
- Schell, J.; Scofield, R.H.; Barrett, J.R.; Kurien, B.T.; Betts, N.; Lyons, T.J.; Zhao, Y.D.; Basu, A. Strawberries improve pain and inflammation in obese adults with radiographic evidence of knee osteoarthritis. Nutrients 2017, 9, 949. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Kurien, B.T.; Tran, H.; Byrd, B.; Maher, J.; Schell, J.; Masek, E.; Barrett, J.R.; Lyons, T.J.; Betts, N.M.; et al. Strawberries decrease circulating levels of tumor necrosis factor and lipid peroxides in obese adults with knee osteoarthritis. Food Funct. 2018, 9, 6218–6226. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Smith, A.; Avalos, M.; South, S.; Crabtree, K.; Wang, W.; Kwon, Y.H.; Vijayagopal, P.; Juma, S. Blueberries improve pain, gait performance, and inflammation in individuals with symptomatic knee osteoarthritis. Nutrients 2019, 11, 290. [Google Scholar] [CrossRef] [Green Version]
- Diaconeasa, Z.; Ranga, F.; Rugină, D.; Leopold, L.; Pop, O.; Vodnar, D.; Cuibus, L.; Socaciu, C. Phenolic content and their antioxidant activity in various berries cultivated in Romania. Bull. Univ. Agric. Sci. Vet. Med. 2015, 72, 99–103. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [Green Version]
- Long, J.; Guo, Y.; Yang, J.; Henning, S.M.; Lee, R.P.; Rasmussen, A.; Zhang, L.; Lu, Q.Y.; Heber, D.; Li, Z. Bioavailability and bioactivity of free ellagic acid compared to pomegranate juice. Food Funct. 2019, 10, 6582–6588. [Google Scholar] [CrossRef]
- CENTRAL Cochrane Central Register of Controlled Trials. Available online: https://www.cochranelibrary.com/central (accessed on 27 August 2021).
- ClinicalTrials.gov Identifier NCT03703024. An Intervention Study Evaluating the Effects of a Raspberry Leaf Extract in an Osteoarthritic Population. Available online: https://clinicaltrials.gov/ct2/show/NCT03703024 (accessed on 27 August 2021).
- IRCT Registry Identifier IRCT2014031517017N1. Effects of Pomegranate Juice on Osteoarthritis of the Knee. Available online: https://trialsearch.who.int/Trial2.aspx?TrialID=IRCT2014031517017N1 (accessed on 27 August 2021).
- IRCT Registry Identifier IRCT201405183664N11. Effect of Dried Hydro Alcoholic Extract of Pomegranate Pericarp on Oxidattive and Inflammation State, Metaloproteinase 1, Metabolic Markers and Clinical Sign in Women with Knee Osteoarthritis. Available online: https://www.who.int/trialsearch/Trial2.aspx?TrialID=IRCT201405183664N11 (accessed on 27 August 2021).
- ClinicalTrials.gov Identifier NCT02518347. Strawberries and Knee Osteoarthritis (OA). Available online: https://clinicaltrials.gov/ct2/show/NCT02518347 (accessed on 27 August 2021).
- Kuyinu, E.L.; Narayanan, G.; Nair, L.S.; Laurencin, C.T. Animal models of osteoarthritis: Classification, update, and measurement of outcomes. J. Orthop. Surg. Res. 2016, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadipour-Jahromy, M.; Mozaffari-Kermani, R. Chondroprotective effects of pomegranate juice on monoiodoacetate-induced osteoarthritis of the knee joint of mice. Phytother. Res. 2010, 24, 182–185. [Google Scholar] [CrossRef]
- Kong, J.; Wang, J.; Gong, X.; Zheng, X.; Chen, T. Punicalagin inhibits tert-butyl hydroperoxide-induced apoptosis and extracellular matrix degradation in chondrocytes by activating autophagy and ameliorates murine osteoarthritis. Drug Des. Devel. 2020, 14, 5521–5533. [Google Scholar] [CrossRef]
- Yang, L.; Fan, C.; Shu, T.; Wang, S. Punicalin alleviates TNF-α- and IL-1β-induced chondrocyte dysfunction and cartilage metabolism via mediating FOXO3 signaling axis. J. Food Biochem. 2021, 45, e13755. [Google Scholar] [CrossRef]
- Lin, Z.; Lin, C.; Fu, C.; Lu, H.; Jin, H.; Chen, Q.; Pan, J. The protective effect of ellagic acid (EA) in osteoarthritis: An in vitro and in vivo study. Biomed. Pharm. 2020, 125, 109845. [Google Scholar] [CrossRef]
- Fu, X.; Gong, L.F.; Wu, Y.F.; Lin, Z.; Jiang, B.J.; Wu, L.; Yu, K.H. Urolithin A targets the PI3K/Akt/NF-κB pathways and prevents IL-1β-induced inflammatory response in human osteoarthritis: In vitro and in vivo studies. Food Funct. 2019, 10, 6135–6146. [Google Scholar] [CrossRef] [PubMed]
- Shivnath, N.; Rawat, V.; Siddiqui, S.; Verma, S.; Gupta, P.; Rais, J.; Khan, M.S.; Arshad, M. Antiosteoarthritic effect of Punica granatum L. peel extract on collagenase induced osteoarthritis rat by modulation of COL-2, MMP-3, and COX-2 expression. Environ. Toxicol. 2021, 36, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.J.; Chen, L.G.; Liang, W.L.; Hsieh, M.S.; Wang, C.C. Inhibitory effects of punicalagin from Punica granatum against type II collagenase-induced osteoarthritis. J. Funct. Foods 2018, 41, 216–222. [Google Scholar] [CrossRef]
- Liu, F.; Yang, H.; Li, D.; Wu, X.; Han, Q. Punicalagin attenuates osteoarthritis progression via regulating Foxo1/Prg4/HIF3α axis. Bone 2021, 152, 116070. [Google Scholar] [CrossRef]
- Elder, S.H.; Mosher, M.L.; Jarquin, P.; Smith, P.; Chironis, A. Effects of short-duration treatment of cartilage with punicalagin and genipin and the implications for treatment of osteoarthritis. J. Biomed. Mater. Res. B Appl. Biomater. 2021, 109, 818–828. [Google Scholar] [CrossRef]
- Shruthi, S.D.; Sujan Ganapathy, P.S.; Kumar, R.; Shivakumara; Dharshan, J.C.; Ramachandra, Y.L. In vivo, in vitro anti-arthritic studies of ellagic acid from Kirganelia reticulata baill and its molecular docking. J Appl. Pharm. Sci. 2014, 4, 24–31. [Google Scholar]
- Tanideh, N.; Nazhvani, S.D.; Bahrami, A.; Mehrabani, D.; Akbarzadeh, A.; Azarpira, N.; Lotfi, M.; Bagheri, M.H.; Koohi-Hosseinabadi, O. The effect of hydroalcoholic extract of Mangifera indica on induced osteoarthritis of knee in male guinea pigs. Comp. Clin. Path. 2016, 25, 973–979. [Google Scholar] [CrossRef]
- Akhtar, N.; Khan, N.M.; Ashruf, O.S.; Haqqi, T.M. Inhibition of cartilage degradation and suppression of PGE2 and MMPs expression by pomegranate fruit extract in a model of posttraumatic osteoarthritis. Nutrition 2017, 33, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Qu, T.B.; Zhai, K.; Ding, J.; Hai, Y.; Zhou, J.L. Gallic acid can play a chondroprotective role against AGE-induced osteoarthritis progression. J. Orthop. Sci. 2015, 20, 734–741. [Google Scholar] [CrossRef]
- Demoor, M.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Fabre, H.; Lafont, J.; Denoix, J.M.; Audigié, F.; Mallein-Gerin, F.; et al. Cartilage tissue engineering: Molecular control of chondrocyte differentiation for proper cartilage matrix reconstruction. Biochim. Biophys. Acta 2014, 1840, 2414–2440. [Google Scholar] [CrossRef]
- Mehana, E.E.; Khafaga, A.F.; El-Blehi, S.S. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef] [PubMed]
- Carames, B.; Taniguchi, N.; Otsuki, S.; Blanco, F.J.; Lotz, M. Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis Rheum. 2010, 62, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, M.F.; Onnerfjord, P.; Kraus, V.B. Biomarkers and proteomic analysis of osteoarthritis. Matrix Biol. 2014, 39, 56–66. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Pelletier, J.P.; Fahmi, H. Cyclooxygenase-2 and prostaglandins in articular tissues. Semin. Arthritis Rheum. 2003, 33, 155–167. [Google Scholar] [CrossRef]
- Shukla, M.; Gupta, K.; Rasheed, Z.; Khan, K.A.; Haqqi, T.M. Bioavailable constituents/metabolites of pomegranate (Punica granatum L) preferentially inhibit COX2 activity ex vivo and IL-1β-induced PGE2 production in human chondrocytes in vitro. J. Inflamm. 2008, 5, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, T.; Alvarez-Garcia, O.; Mokuda, S.; Nagira, K.; Olmer, M.; Gamini, R.; Miyata, K.; Akasaki, Y.; Su, A.I.; Asahara, H.; et al. FoxO transcription factors modulate autophagy and proteoglycan 4 in cartilage homeostasis and osteoarthritis. Sci. Transl. Med. 2018, 10, eaan0746. [Google Scholar] [CrossRef] [Green Version]
- Freise, N.; Burghard, A.; Ortkras, T.; Daber, N.; Imam Chasan, A.; Jauch, S.L.; Fehler, O.; Hillebrand, J.; Schakaki, M.; Rojas, J.; et al. Signaling mechanisms inducing hyporesponsiveness of phagocytes during systemic inflammation. Blood 2019, 134, 134–146. [Google Scholar] [CrossRef]
- He, L.; Pan, Y.; Yu, J.; Wang, B.; Dai, G.; Ying, X. Decursin alleviates the aggravation of osteoarthritis via inhibiting PI3K-Akt and NF-κB signal pathway. Int. Immunopharmacol. 2021, 97, 107657. [Google Scholar] [CrossRef] [PubMed]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18, S17–S23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerwin, N.; Bendele, A.M.; Glasson, S.; Carlson, C.S. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the rat. Osteoarthr. Cartil. 2010, 18, S24–S34. [Google Scholar] [CrossRef] [Green Version]
- Kraus, V.B.; Huebner, J.L.; DeGroot, J.; Bendele, A. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the guinea pig. Osteoarthr. Cartil. 2010, 18, S35–S52. [Google Scholar] [CrossRef] [Green Version]
- Laverty, S.; Girard, C.A.; Williams, J.M.; Hunziker, E.B.; Pritzker, K.P. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the rabbit. Osteoarthr. Cartil. 2010, 18, S53–S65. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.L.; Pang, Z.Y.; Chen, X.M.; Li, Z.; Liu, X.X.; Zhai, Q.L.; Huang, J.M.; Ruan, Z.Y. Urolithin A attenuates IL-1β-induced inflammatory responses and cartilage degradation via inhibiting the MAPK/NF-κB signaling pathways in rat articular chondrocytes. J. Inflamm. 2020, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Wang, N.; Hafeez, B.B.; Cheruvu, V.K.; Haqqi, T.M. Punica granatum L. extracts inhibits IL-1β-induced expression of matrix metalloproteinases by inhibiting the activation of MAP kinases and NF-κB in human chondrocytes in vitro. J. Nutr. 2005, 135, 2096–2102. [Google Scholar] [CrossRef] [Green Version]
- Haseeb, A.; Khan, N.M.; Ashruf, O.S.; Haqqi, T.M. A polyphenol-rich pomegranate fruit extract suppresses NF-κB and IL-6 expression by blocking the activation of IKKβ and NIK in primary human chondrocytes. Phytother. Res. 2017, 31, 778–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasheed, Z.; Akhtar, N.; Haqqi, T.M. Pomegranate extract inhibits the interleukin-1β-induced activation of MKK-3, p38α-MAPK and transcription factor RUNX-2 in human osteoarthritis chondrocytes. Arthritis Res. 2010, 12, R195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, N.; Bank, R.A.; TeKoppele, J.M.; DeGroot, J. AGEing and osteoarthritis: A different perspective. Curr. Opin. Rheumatol. 2003, 15, 616–622. [Google Scholar] [CrossRef] [PubMed]
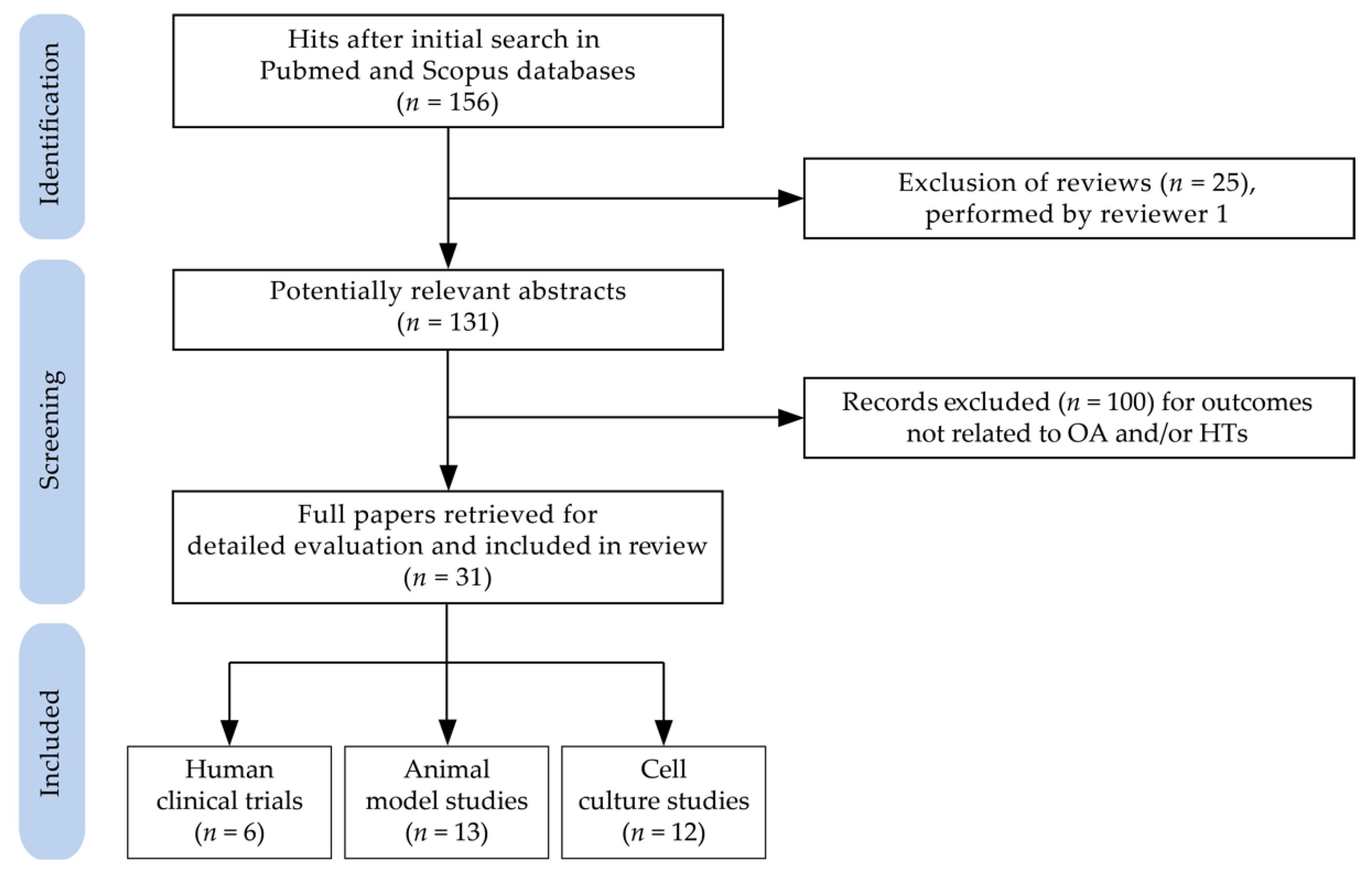
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA extension for scoping reviews (PRISMA-ScR): Checklist and explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, K.D.; Thoma, L.M.; Golightly, Y.M. Epidemiology of osteoarthritis. Osteoarthr. Cartil. 2022, 30, 184–195. [Google Scholar] [CrossRef]
- Kim, H.; Castellon-Chicas, M.J.; Arbizu, S.; Talcott, S.T.; Drury, N.L.; Smith, S.; Mertens-Talcott, S.U. Mango (Mangifera indica L.) polyphenols: Anti-inflammatory intestinal microbial health benefits, and associated mechanisms of actions. Molecules 2021, 26, 2732. [Google Scholar] [CrossRef]
- Danesi, F.; Kroon, P.A.; Saha, S.; de Biase, D.; D’Antuono, L.F.; Bordoni, A. Mixed pro- and anti-oxidative effects of pomegranate polyphenols in cultured cells. Int. J. Mol. Sci. 2014, 15, 19458–19471. [Google Scholar] [CrossRef]
- Aragonès, G.; Danesi, F.; Del Rio, D.; Mena, P. The importance of studying cell metabolism when testing the bioactivity of phenolic compounds. Trends Food Sci. Technol. 2017, 69, 230–242. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.A.; González-Sarrías, A.; Espín, J.C. In vitro research on dietary polyphenols and health: A call of caution and a guide on how to proceed. J. Agric. Food Chem. 2018, 66, 7857–7858. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Study Design | Population | Subjects (Gender, No., Age, BMI, Physical Activity) | Intervention | Control/Comparator | Duration | Main OA-Related Outcomes | Reference |
|---|---|---|---|---|---|---|---|
| Randomized, pre- and post-test, parallel-arm | Adults with knee OA | F: 34, M: 4; 41–66 years; BMI: 23–36; I: 17, LA: 17, A: 4 | Pomegranate juice 200 mL/day | – | 6 weeks | Clinical indexes: ↔ WOMAC pain, ↓ WOMAC stiffness, physical function, and total score. Biochemical outcomes: ↔ serum MMP-1, MMP-13 and ↑ GPx serum levels | Ghoochani et al. 2016 [43] |
| Randomized, double-blind, placebo-controlled, parallel-arm | Adult obese women with knee OA | F: 60; 40–58 years; BMI: 30–34; I: 0, LA: 46, A: 14 | Encapsulated pomegranate peel hydroalcoholic extract 500 mg b.i.d. | Placebo 1 capsule b.i.d. | 8 weeks | Biochemical outcomes: ↑ KOOS, ↓ VAS | Rafraf et al. 2017 [44] |
| Randomized, double-blind, placebo-controlled, parallel-arm | Adult obese women with knee OA | F: 60; 40–58 years; BMI: 30–34; I: 0, LA: 46, A: 14 | Encapsulated pomegranate peel hydroalcoholic extract 500 mg b.i.d. | Placebo 1 capsule b.i.d. | 8 weeks | Biochemical outcomes: ↓ MDA, ↑ SOD, GPx, and TAC activity | Haghighian et al. 2021 [45] |
| Randomized, double-blind, placebo-controlled, crossover | Obese adults with knee OA | F: 13, M: 4; 50–64 years; BMI: 38–41 | Freeze-dried strawberry powder 50 g b.i.d. (equivalent to ≈ 500 g of fresh strawberries) | Placebo powder 50 g b.i.d. | 12 weeks | Clinical indexes: ↓ HAQ-DI, ↔ pain VAS, ↓ ICOAP constant pain, intermittent pain, and total pain. Biochemical outcomes: ↔ CRP, MMP-8, and nitrite levels, ↓ IL-1β, IL-6, and MMP-3 levels | Schell et al. 2017 [46] |
| Randomized, double-blind, placebo-controlled, crossover | Obese adults with knee OA | F: 13, M: 4; 54–60 years; BMI: 38–41 | Freeze-dried strawberry powder 50 g b.i.d. (equivalent to ≈ 500 g of fresh strawberries) | Placebo powder 50 g b.i.d. | 12 weeks | Biochemical outcomes: ↓ TNF-α, TNF-R2, 4-HNE, and CD levels, ↔ IL-19, CD163, TNF-R1, and PTX3 levels | Basu et al. 2018 [47] |
| Randomized, double-blind placebo-controlled, parallel-arm | Obese adults with knee OA | F: 47, M: 16; 55–57years; BMI: 31–33 | Freeze-dried blueberry powder 40 g/day | Placebo powder 40 g/day | 4 months | Clinical indexes: ↓ WOMAC pain, stiffness, physical function, and total score. Biochemical outcomes: ↔ TNF-α, IL-1, IL-6, IL-10, IL-13, MMP-3, MMP-13, and MCP-1 levels | Du et al. 2019 [48] |
| CENTRAL Identifier | Study Focus | Intervention/ Treatment | Study Design, Duration | Sponsor | Estimated Enrolment | Study Start Date | Completion Date | Reference |
|---|---|---|---|---|---|---|---|---|
| IRCT2014031517017N1 | Knee OA | Pomegranate juice | Double-blind, placebo-controlled, parallel-arm, randomized, 6 weeks | Ahvaz Jundishapur University of Medical Sciences (Ahvaz, Iran) | 50 | March 2014 | March 2015 | [54] |
| IRCT201405183664N11 | Knee OA | Pomegranate pericarp hydroalcoholic extract | Double-blind, placebo-controlled, parallel-arm, randomized, 2 months | Tabriz University of Medical Sciences (Tabriz, Iran) | 30 | August 2014 | December 2014 | [55] |
| NCT02518347 | Knee OA | Freeze-dried strawberries | Triple-blind, placebo-controlled, crossover, randomized, 12 weeks | Oklahoma State University (Stillwater, OK, USA) | 20 | March 2015 | May 2017 | [56] |
| NCT03703024 | Knee OA | Raspberry leaf extract | Double-blind, placebo-controlled, parallel-arm, randomized, 12–13 weeks | Atlantia Food Clinical Trials (Blackpool, Munster, Ireland) | 195 | June 2017 | December 2018 | [53] |
| Species | Animal Model Characteristics (Sex, Age, Weight) | Tested Product(s), Vehicle, Route of Administration, Duration | OA Induction | Effects | Reference |
|---|---|---|---|---|---|
| Mouse | BALB/c, male, 20–25 g | Pomegranate juice (4, 10, 20 mL/kg b.w. b.i.d. in divided doses) via oral gavage for 2 weeks | MIA intra-articular injection | ↓ MIA effects (especially with the 20 mL/kg dose) | Hadipour-Jahromy and Mozaffari-Kermani 2010 [58] |
| C57BL/6, male, 7–8-week-old, 20–25 g | PUNI (20 mg/kg b.w. per day) via oral gavage for 8 weeks | DMM | ↓ cellular apoptosis, ↓ OARSI score | Kong et al. 2020 [59] | |
| C57BL/6J, male, 10-week-old | Punicalin (100 mg/kg b.w. per day, twice a week) in PBS via gastric gavage for 4 weeks | 10 ng IL-1β and 50 ng TNF-α in 5-μL PBS (twice a week for 4 weeks) | ↓ FOXO3 phosphorylation, ↑ Sox9, Col2a1, and Foxo3 gene expression, ↓ Mmp-9, Mmp-13, Col10a1, Runx2, Ihh, and Pthlh gene expression | Yang et al. 2021 [60] | |
| C57BL/6 wild type, male, 10-week-old | EA (40 mg/kg b.w. every 2 days) in saline via gastric gavage for 8 weeks | DMM | ↓ OARSI score, ↓ synovitis score | Lin et al. 2020 [61] | |
| C57BL/6 wild type, male, 1-week-old | Uro-A (20 mg/kg b.w. per day) in CMC-Na (0.5%) via gastric gavage for 8 weeks | DMM | ↓ OARSI scores, ↓ p-PI3K and p-AKT-positive chondrocytes, ↓ p65-positive nuclei | Fu et al. 2019 [62] | |
| Rat | Sprague Dawley, female, 100–150 g | Pomegranate peel hydroalcoholic extract (250 or 500 mg/kg b.w. per day) via oral administration for 1 month | Collagenase II | ↓ Mankin score, ↓ serum ALP, ↓ Mmp-3 and Cox-2 gene expression, ↑ Col2 gene expression, ↑ collagen and glycosaminoglycan content | Shivnath et al. 2021 [63] |
| Wistar, ≈300 g | Pomegranate peel acetone extract (15 and 150 mg/kg b.w.) via oral administration for 28 days | Collagenase II | ↓ weight-bearing ratio | Lee et al. 2018 [64] | |
| Sprague Dawley, male, 8-week-old, 300–350 g | PUNI (10 mg/kg b.w. per day) in saline via oral gavage for 12 weeks | ACLT-MCLT-DMM | ↓ OARSI score, ↓ apoptosis rate, ↑ protein expression of FOXO1, PRG4, HIF3α, ACAN, COL2, p-ULK1, p-Beclin1, LC3II/I ratio, ↓ protein expression of ADAMTS5, MMP-13, and p62 | Liu et al. 2021 [65] | |
| Sprague-Dawley, male, 200–250 g | PUNI (30 μL of 9.2 mM, twice a week, in saline, via intravenous injection for 5 weeks | MIA intra-articular injection | ↓ OARSI cartilage matrix width loss measurements | Elder et al. 2021 [66] | |
| Wistar albino rats, male, 150–200 g | EA (250 mg/mL) in saline via intra-articular injection for 20 days | Formaldehyde | ↓ paw edema volume, ↑ movement ability, ↑ b.w. | Shruthi et al. 2014 [67] | |
| Guinea pig | Short-haired England, male, 700–800 g | Mango fruit hydroalcoholic extract (500 mg/kg b.w. per day) via oral gavage for 8 weeks (oral treatment) or intra-articular injection 2 times with a time interval of 4 weeks (injection treatment) | ACLT | ↑ radiological and histopathological assessments (intra-articular injection only) | Tanideh et al. 2016 [68] |
| Rabbit | New Zealand White rabbits, male, 8-month-old | Pomegranate fruit hydroalcoholic extract (34 mg/kg b.w. per day) via drinking water, ad libitum, for 8 weeks | ACLT | ↓ OARSI score, ↓ Mmp-3, Mmp-9, and Mmp-13 gene expression, ↑ Acan and Col2a1 gene expression, ↓ IL-1β, IL-6, and PGE2 levels in plasma and synovial fluid | Akhtar et al. 2017 [69] |
| New Zealand rabbits, male, 3-week-old, 400–600 g | GA (0.5 mL of 80 μM) in PBS via intra-articular injection every 72 h for 8 consecutive weeks | Collagenase II | ↓ Mankin score | Wen et al. 2015 [70] |
| Cell Model | Primary Cell/Cell Line/Tissue | Tested Compound(s), Dose, Duration | Pro-Inflammatory Treatment | Biological Effects | Reference |
|---|---|---|---|---|---|
| Mouse chondrocytes | Primary chondrocytes from immature C57BL/6 mice | PUNI (25 or 50 μg/mL) for 24 h | 100 μM TBHP | ↑ protein expression of ATG12-5, LC3 II/I, p-ULK, Beclin1, Bcl-2, COL2, HO-1, SOD1, and NQO1, ↓ protein expression of BAX, cleaved caspase 3, ADAMTS5, MMP-3 and MMP-13, ↓ p62 protein expression (dose-dependent) | Kong et al. 2020 [59] |
| Primary chondrocytes C57BL/6J mice | Punicalin (80 or 100 μg/mL) for 24 h | 10 ng/mL IL-1β and 50 ng/mL TNF-α | ↓ p-FOXO3 protein expression, ↑ Sox9 and Col2a1 mRNA levels, ↓ Col10a1, Mmp-9, Ihh, Pthlh, and Runx2 mRNA levels, ↔ Mmp-13, Col9a1, and Runx3 mRNA levels (dose-dependent) | Yang et al. 2021 [60] | |
| Rat chondrocytes | Primary chondrocytes from Wistar rats | Pomegranate peel acetone extract (12.5–100 μg/mL) or PUNI (25–50 μg/mL) for 16 h as co-treatment with pro-inflammatory stimulus | 10 ng/mL IL-1β | ↓ iNOS, MMP-13, and COX-2 protein expression, ↓ PGE2 release (dose-dependent) | Lee et al. 2018 [64] |
| Primary chondrocytes from 5-day-old Sprague Dawley rats | PUNI (25, 50, and 100 μM of) for 8 h prior to pro-inflammatory stimulus | 10 ng/mL of LPS | ↑ mRNA levels and protein expression of FOXO1, PRG4, HIF3α, p-ULK1, p-Beclin1, and LC3 II/I ratio, ↓ p62 protein expression | Liu et al. 2021 [65] | |
| Primary chondrocytes from 2- (in vitro study) and 4-week-old (ex vivo study) Sprague Dawley rats | Uro-A (1–15 μM) for 2 or 3 days, or 2 h prior to pro-inflammatory stimulus | 20 ng/mL IL-1β for 2 days (in vitro) or 30 ng/mL IL-1β for 3 days (ex vivo) | in vitro: ↓ protein expression of MMP-3, MMP-9, MMP-13, ADAMTS4, COX-2 and iNOS, ↑ gene and protein expression of COL2 and SOX9, ↓ p-NF-κB, p65, p-ERK1/2, p-JNK, and p-p38 MAPK protein expression (dose-dependent); ex vivo: ↓ OARSI score, ↑ COL2 and ACAN protein expression (dose-dependent) | Ding et al. 2020 [84] | |
| Rabbit chondrocytes | Primary chondrocytes from 1-year-old New Zealand white male rabbits | Blood plasma from rabbits given pomegranate fruit extract (10 mL, 34 mg/kg b.w., via oral gavage for 48 h) for 2 h prior to pro-inflammatory stimulus | 5 ng/mL IL-1β for 24 | ↓ PGE2 and NO production | Shukla et al. 2008 [76] |
| Primary chondrocytes from 3-week-old New Zealand male rabbits | GA (10–80 μg/mL) for 48 h prior to pro-inflammatory stimulus | 100 μg/mL AGEs | ↑ Col2a1 gene expression, ↑ COL2 and ACAN protein expression, ↓ iNOS and COX-2 protein expression, ↓ NO and PGE2 release, ↑ SOD activity and GSH content, ↓ ROS release (dose-dependent) | Wen et al. 2015 [70] | |
| Human chondrocytes | Primary chondrocytes (in vitro) and articular cartilage slides (ex vivo) | Pomegranate fruit extract (6.25–50 mg/L) for 24 h (in vitro), pomegranate fruit extract (25 or 50 mg/L) for 72 h (ex vivo) | 5 μg/L IL-1β (in vitro), 10 μg/L IL-1β (ex vivo) | in vitro: ↓ MMP-1, MMP-3, MMP-13 gene and protein expression, ↓ protein expression of p-ERK, p-JNK, p-p38-MAPK, c-JUN, ATF-2, NF-κB, and p-IκBα (dose-dependent); ex vivo: ↓ glycosaminoglycan release (dose-dependent) | Ahmed et al. 2005 [85] |
| Primary chondrocytes | Pomegranate fruit extract (10 or 50 μg/mL) for 2 h after an overnight serum starvation | 10 ng/mL IL-1β | ↓ IL-6 gene and protein expression, ↓ ROS level, ↓ NF-κB/p65, ↑ IκBα, ↓ p-IKKα/β, ↓ p-IKKβ gene and protein expression (dose-dependent) | Haseeb et al. 2017 [86] | |
| Primary chondrocytes from 19 patients (58- 77 years, 14 women and 5 men) | Pomegranate fruit extract (6.25 to 100 μg/mL) for 1 or 2 h after overnight serum starvation, prior to pro-inflammatory stimulus | 10 ng/mL IL-1β | ↓ protein expression of p-MKK3, p-MKK6, p-p38 MAPKα, and RUNX2 (dose-dependent) | Rasheed et al. 2010 [87] | |
| Primary chondrocytes from 8 patients (52–73 years, 4 women and 4 men) | EA (12.5, 25, and 50 μM) for 24 or 48 h prior to pro-inflammatory stimulus | 10 ng/mL IL-1β | ↓ NO, iNOS, PGE2, COX-2, IL-6, TNF-α, ADAMTS5, and MMP-13 levels, ↓ iNOS and COX-2 gene expression, ↑ COL2 and ACAN protein expression, ↓ NF-κB protein expression (dose-dependent) | Lin et al. 2020 [61] | |
| Primary chondrocytes from 6 patients (65–70 years, 3 women and 3 men) | Uro-A (3, 10, and 30 μM) for 24 h prior to pro-inflammatory stimulus | 10 ng/mL IL-1β | ↓ iNOS, COX-2, IL-6, and TNF-α mRNA levels, ↓ PGE2, NO, iNOS, TNF-α, IL-6, COX-2, ADAMTS5, and MMP-13 levels, ↑ COL2 and ACAN protein expression, ↑ IκBα, ↓ p65, ↓ p-PI3K, and p-AKT protein expression (dose-dependent) | Fu et al. 2019 [62] |
| Research Study Type | Studies Where Bioavailability and Metabolism of HTs Have Been Considered † | Studies Where Bioavailability and Metabolism of HTs Have Not Been Considered ‡ |
|---|---|---|
| Animal model studies | Akhtar et al., 2017 [69], Fu et al., 2019 [62], Hadipour-Jahromy and Mozaffari-Kermani 2010 [58], Kong et al., 2020 [59], Lee et al., 2018 [64], Lin et al., 2020 [61], Liu et al., 2021 [65], Shivnath et al., 2021 [63], Shruthi et al., 2014 [67], Tanideh et al., 2016 (oral treatment) [68], Wen et al., 2015 [70], Yang et al. 2021 [60] | Ding et al., 2020 [84], Fu et al., 2019 [62], Lin et al., 2020 [61], Shukla et al., 2008 [76], Wen et al., 2015 [70] |
| Cell culture studies | Ding et al., 2020 [84], Fu et al., 2019 [62], Lin et al., 2020 [61], Shukla et al., 2008 [76], Wen et al., 2015 [70] | Ahmed et al., 2005 [85], Haseeb et al., 2017 [86], Kong et al., 2020 [59], Lee et al., 2018 [64], Liu et al., 2021 [65], Rasheed et al., 2010 [87], Yang et al., 2021 [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govoni, M.; Danesi, F. Do Pomegranate Hydrolyzable Tannins and Their Derived Metabolites Provide Relief in Osteoarthritis? Findings from a Scoping Review. Molecules 2022, 27, 1033. https://doi.org/10.3390/molecules27031033
Govoni M, Danesi F. Do Pomegranate Hydrolyzable Tannins and Their Derived Metabolites Provide Relief in Osteoarthritis? Findings from a Scoping Review. Molecules. 2022; 27(3):1033. https://doi.org/10.3390/molecules27031033
Chicago/Turabian StyleGovoni, Marco, and Francesca Danesi. 2022. "Do Pomegranate Hydrolyzable Tannins and Their Derived Metabolites Provide Relief in Osteoarthritis? Findings from a Scoping Review" Molecules 27, no. 3: 1033. https://doi.org/10.3390/molecules27031033
APA StyleGovoni, M., & Danesi, F. (2022). Do Pomegranate Hydrolyzable Tannins and Their Derived Metabolites Provide Relief in Osteoarthritis? Findings from a Scoping Review. Molecules, 27(3), 1033. https://doi.org/10.3390/molecules27031033