
β-Myrcene Mitigates Colon Inflammation by Inhibiting MAP Kinase and NF-κB Signaling Pathways

 , and
, and
Abstract
1. Introduction
2. Results
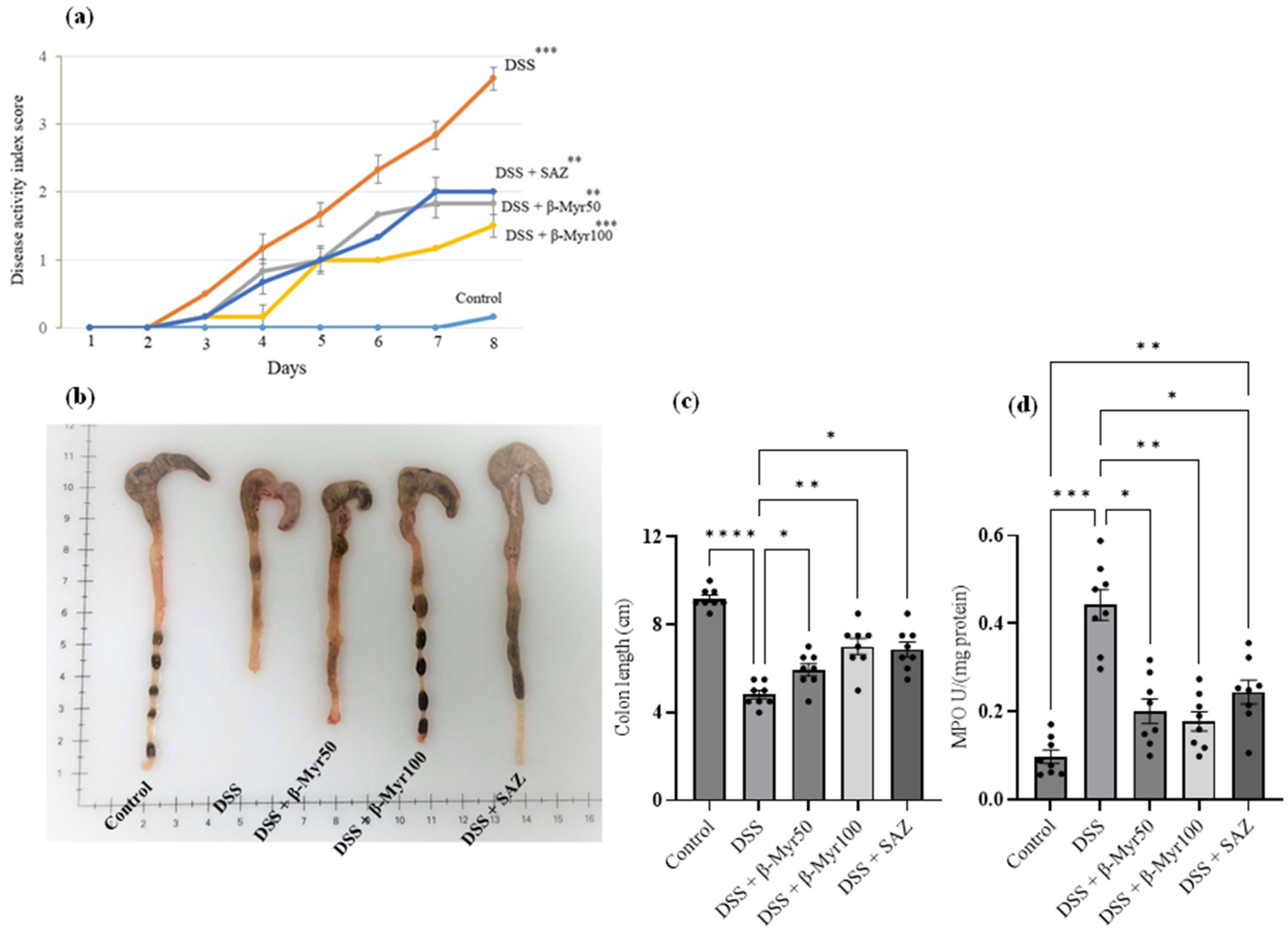
2.1. Effect of β-Myrcene on Disease Activity Index (DAI), Colon Length, and Myeloperoxidase (MPO) Enzyme Activity
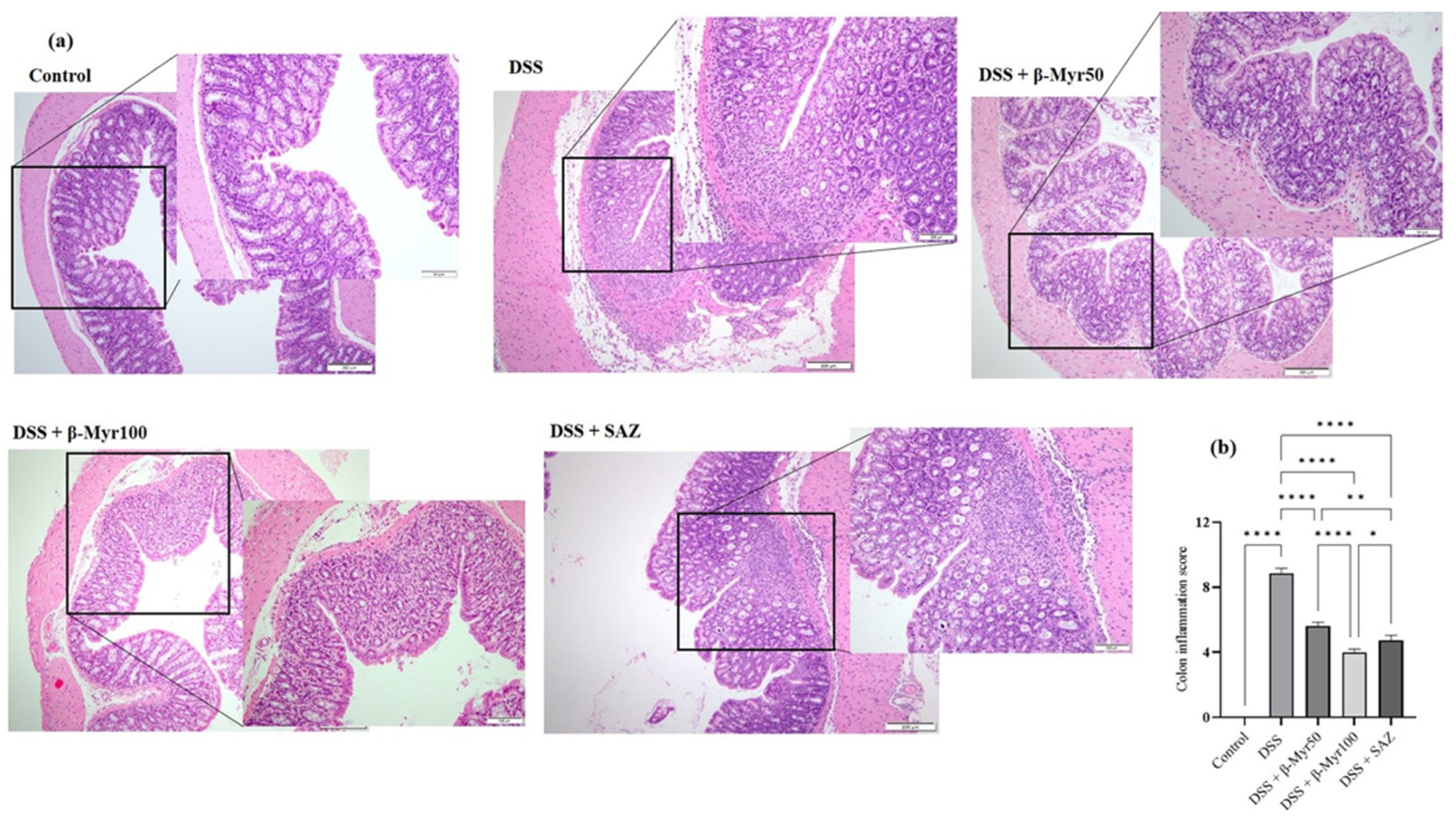
2.2. Effect of β-Myrcene on Colon Histology and Colon Inflammation Scores
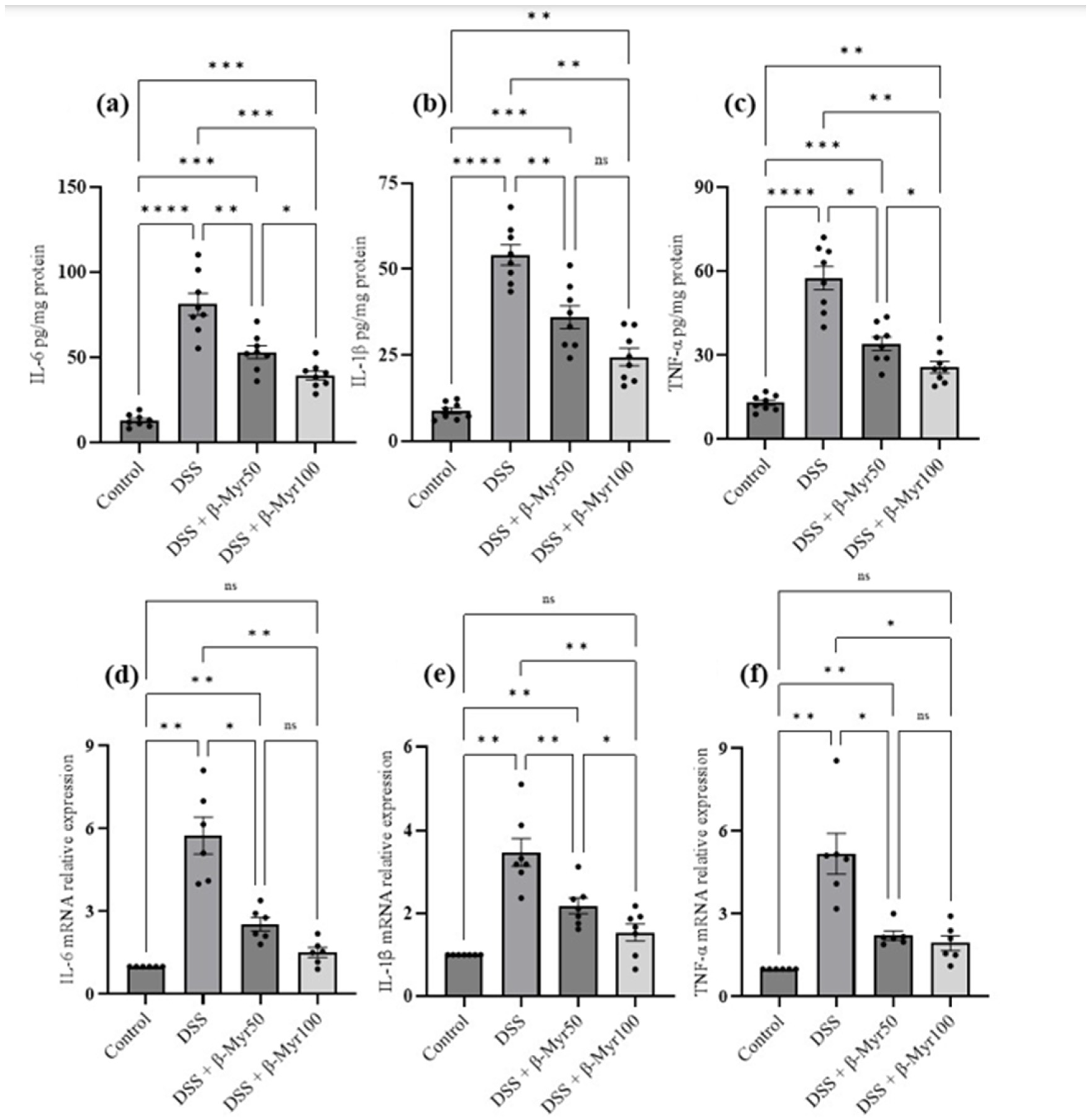
2.3. Effect of β-Myrcene on Proinflammatory Cytokines and Proinflammatory Mediators
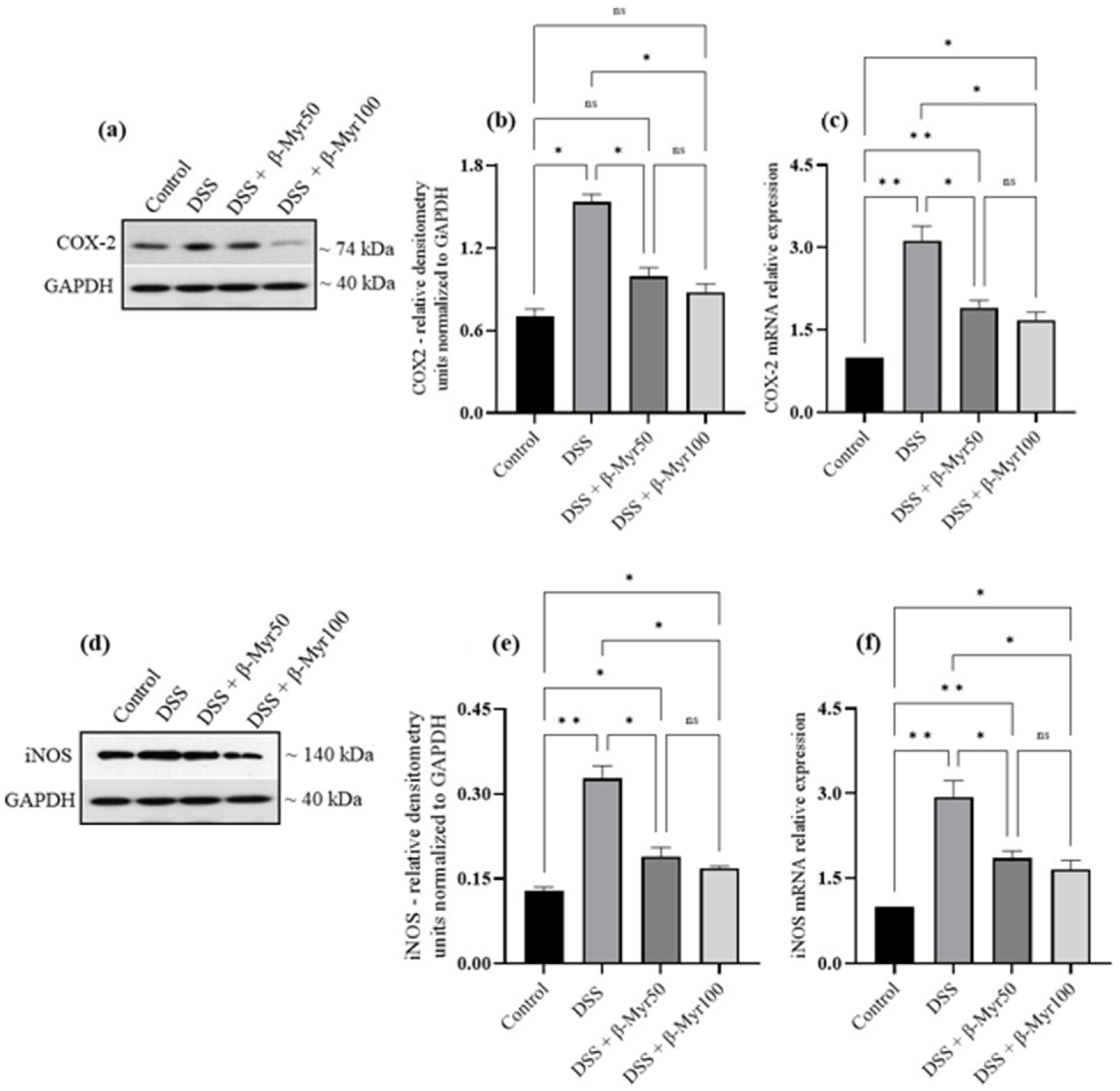
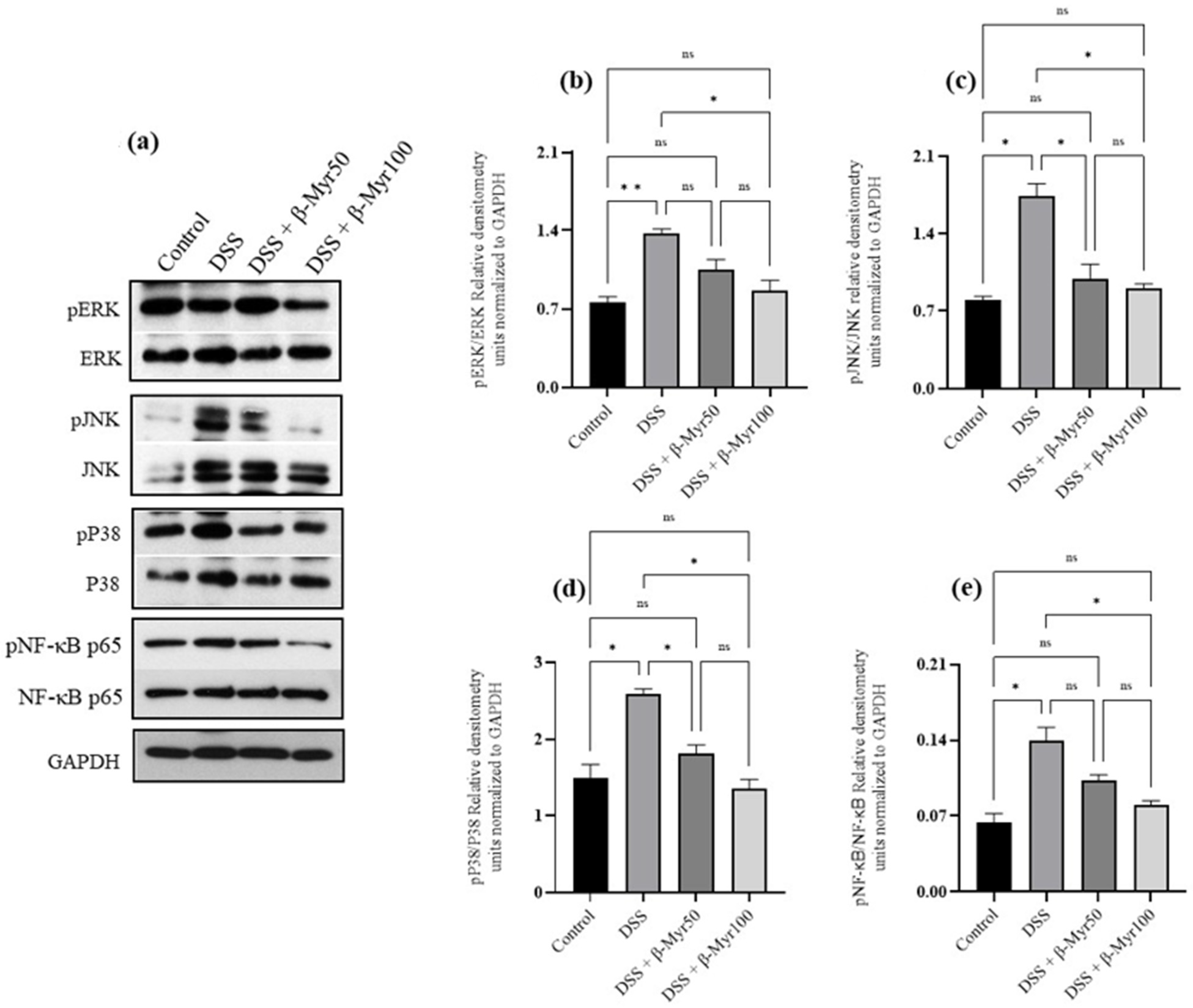
2.4. Effect of β-Myrcene on MAPK and Nf-kB Signaling Pathways
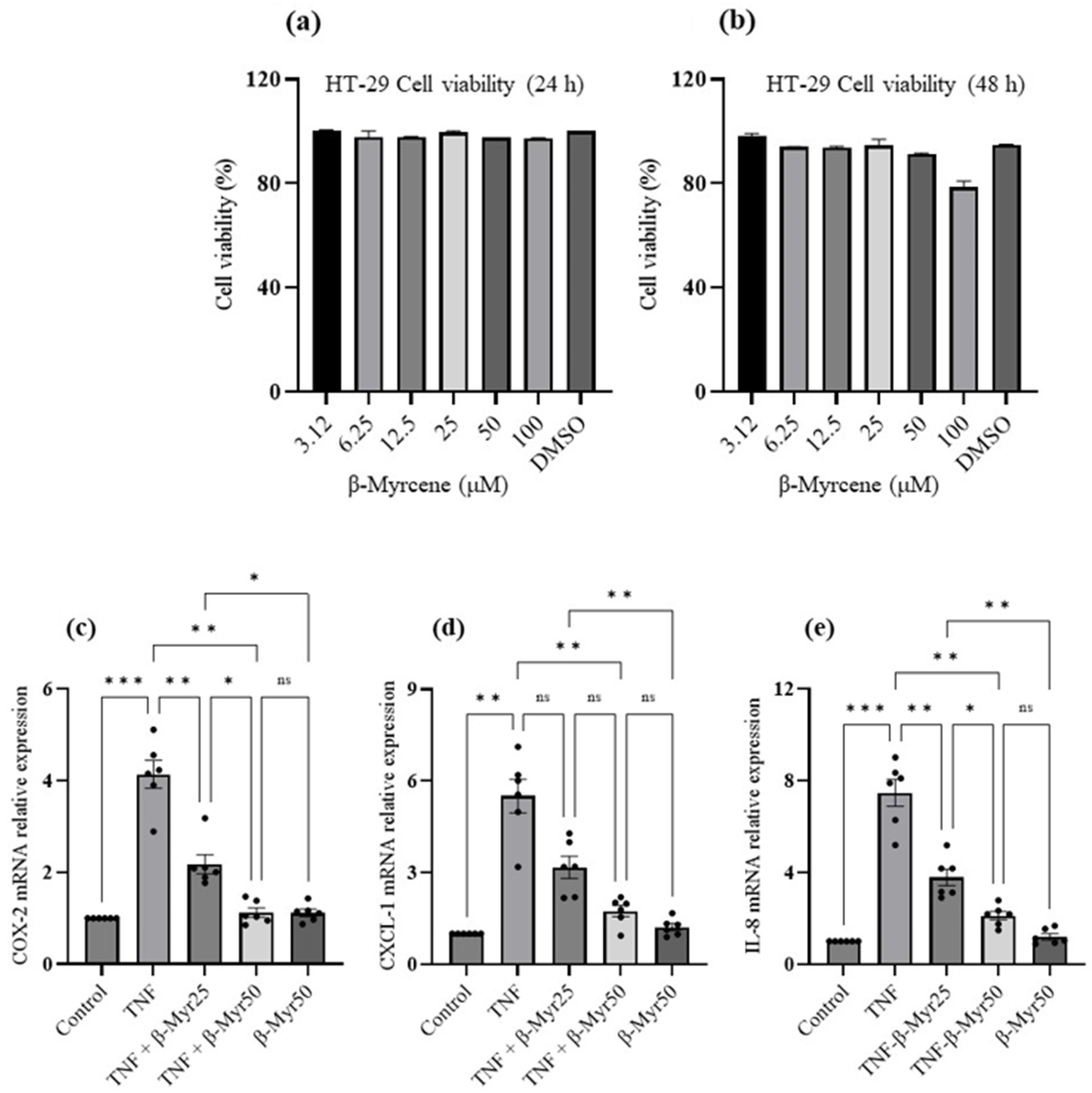
2.5. Effect of β-Myrcene on TNF-α Challenged HT-29 Colonic Adenocarcinoma Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Experimental Design
4.4. Evaluation of the Clinical Score for Colitis
4.5. Histopathological Evaluation
4.6. Myeloperoxidase Assay and ELISA
4.7. RNA Extraction and Real-Time PCR
4.8. Western Blot
4.9. HT-29 Cell Culture
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Park, S.C.; Jeen, Y.T. Genetic Studies of Inflammatory Bowel Disease-Focusing on Asian Patients. Cells 2019, 8, 404. [Google Scholar] [CrossRef]
- Larabi, A.; Barnich, N.; Nguyen, H.T.T. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy 2020, 16, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Lee, M.; Chang, E.B. The Gut Microbiome and Inflammatory Bowel Diseases. Annu. Rev. Med. 2022, 73, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.J.; Dhawan, A.; Saeed, S.A. Inflammatory Bowel Disease in Children and Adolescents. JAMA Pediatr. 2015, 169, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Verstockt, B.; Ferrante, M.; Vermeire, S.; Van Assche, G. New treatment options for inflammatory bowel diseases. J. Gastroenterol. 2018, 53, 585–590. [Google Scholar] [CrossRef]
- Kennedy, N.A.; Heap, G.A.; Green, H.D.; Hamilton, B.; Bewshea, C.; Walker, G.J.; Thomas, A.; Nice, R.; Perry, M.H.; Bouri, S.; et al. Predictors of anti-TNF treatment failure in anti-TNF-naive patients with active luminal Crohn’s disease: A prospective, multicentre, cohort study. Lancet Gastroenterol. Hepatol. 2019, 4, 341–353. [Google Scholar] [CrossRef]
- Danese, S.; Vermeire, S.; Hellstern, P.; Panaccione, R.; Rogler, G.; Fraser, G.; Kohn, A.; Desreumaux, P.; Leong, R.W.; Comer, G.M.; et al. Randomised trial and open-label extension study of an anti-interleukin-6 antibody in Crohn’s disease (ANDANTE I and II). Gut 2019, 68, 40–48. [Google Scholar] [CrossRef]
- Hwang, E.; Ngo HT, T.; Park, B.; Seo, S.A.; Yang, J.E.; Yi, T.H. Myrcene, an Aromatic Volatile Compound, Ameliorates Human Skin Extrinsic Aging via Regulation of MMPs Production. Am. J. Chin. Med. 2017, 45, 1113–1124. [Google Scholar] [CrossRef]
- National Toxicology, P. NTP technical report on the toxicology and carcinogenesis studies of beta-myrcene (CAS No. 123-35-3) in F344/N rats and B6C3F1 mice (Gavage studies). Natl. Toxicol. Program. Tech. Rep. Ser. 2010, 557, 1–163. [Google Scholar]
- Fabbri, J.; Maggiore, M.A.; Pensel, P.E.; Albani, C.M.; Denegri, G.M.; Elissondo, M.C. Could beta-myrcene be an alternative to albendazole for the treatment of experimental cystic echinococcosis? Acta Trop. 2018, 187, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, O.; Tanyildizi, S.; Godekmerdan, A. Curcumin, myrecen and cineol modulate the percentage of lymphocyte subsets altered by 2,3,7, 8-tetracholorodibenzo-p-dioxins (TCDD) in rats. Hum. Exp. Toxicol. 2011, 30, 1986–1994. [Google Scholar] [CrossRef] [PubMed]
- Rufino, A.T.; Ribeiro, M.; Sousa, C.; Judas, F.; Salgueiro, L.; Cavaleiro, C.; Mendes, A.F. Evaluation of the anti-inflammatory, anti-catabolic and pro-anabolic effects of E-caryophyllene, myrcene and limonene in a cell model of osteoarthritis. Eur. J. Pharmacol. 2015, 750, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Bonamin, F.; Moraes, T.M.; Dos Santos, R.C.; Kushima, H.; Faria, F.M.; Silva, M.A.; Junior, I.V.; Nogueira, L.; Bauab, T.M.; Souza Brito, A.R.; et al. The effect of a minor constituent of essential oil from Citrus aurantium: The role of beta-myrcene in preventing peptic ulcer disease. Chem. Biol. Interact. 2014, 212, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyama, K.; Suzuki, A.; Tomiyasu, N.; Tsuruta, O.; Kitazaki, S.; Takeda, T.; Satoh, Y.; Bennett, B.L.; Toyonaga, A.; Sata, M. Pro-inflammatory signaling by Jun-N-terminal kinase in inflammatory bowel disease. Int. J. Mol. Med. 2006, 17, 449–455. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Waetzig, G.H.; Seegert, D.; Rosenstiel, P.; Nikolaus, S.; Schreiber, S. p38 mitogen-activated protein kinase is activated and linked to TNF-alpha signaling in inflammatory bowel disease. J. Immunol. 2002, 168, 5342–5351. [Google Scholar] [CrossRef]
- Cardia GF, E.; Silva-Comar FM, S.; Bonetti, C.I.; da Rocha EM, T.; Zagoto, M.; do Amaral, V.; Bracht, L.; Silva-Filho, S.E.; Bersani-Amado, C.A.; Cuman, R.K.N. Hepatoprotective effect of beta-myrcene pretreatment against acetaminophen-induced liver injury. Avicenna J. Phytomed. 2022, 12, 388–400. [Google Scholar]
- Jiawei, T.; Rui, Z.; Qiaoji, Q.; Xuezhi, Z.; Aiguo, L.; Nan, L. Myrcene enhances the cardioprotective effect through matrix remodeling in an experimental model of heart failure. Arch. Med. Sci. 2020, 16, 1–12. [Google Scholar]
- Broom, O.J.; Widjaya, B.; Troelsen, J.; Olsen, J.; Nielsen, O.H. Mitogen activated protein kinases: A role in inflammatory bowel disease? Clin. Exp. Immunol. 2009, 158, 272–280. [Google Scholar] [CrossRef]
- Rao, V.S.; Menezes, A.M.; Viana, G.S. Effect of myrcene on nociception in mice. J. Pharm. Pharmacol. 1990, 42, 877–878. [Google Scholar] [CrossRef]
- do Vale, T.G.; Furtado, E.C.; Santos, J.G., Jr.; Viana, G.S. Central effects of citral, myrcene and limonene, constituents of essential oil chemotypes from Lippia alba (Mill.) n.e. Brown. Phytomedicine 2002, 9, 709–714. [Google Scholar] [CrossRef]
- Ojeda-Sana, M.A.; Catalina, M.V.B.; Miguel, A.E.; Miguel, A.J.; Silvia, M. New insights into antibacterial and antioxidant activities of rosemary essential oils and their main components. Food Control 2012, 13, 189–195. [Google Scholar] [CrossRef]
- Souza, M.C.; Siani, A.C.; Ramos, M.F.; Menezes-de-Lima, O.J.; Henriques, M.G. Evaluation of anti-inflammatory activity of essential oils from two Asteraceae species. Pharmazie 2003, 58, 582–586. [Google Scholar] [PubMed]
- Inoue, Y.; Shiraishi, A.; Hada, T.; Hirose, K.; Hamashima, H.; Shimada, J. The antibacterial effects of terpene alcohols on Staphylococcus aureus and their mode of action. FEMS Microbiol. Lett. 2004, 237, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Chaouki, W.; Leger, D.Y.; Liagre, B.; Beneytout, J.L.; Hmamouchi, M. Citral inhibits cell proliferation and induces apoptosis and cell cycle arrest in MCF-7 cells. Fundam. Clin. Pharmacol. 2009, 23, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Sawamura, M.; Sun, S.H.; Ozaki, K.; Ishikawa, J.; Ukeda, H. Inhibitory effects of citrus essential oils and their components on the formation of N-nitrosodimethylamine. J. Agric. Food Chem. 1999, 47, 4868–4872. [Google Scholar] [CrossRef] [PubMed]
- FDA. Food Additive Regulations; Synthetic Flavoring Agents and Adjuvants; FDA: Silver Spring, MD, USA.
- Joint, F.A.O.; WHO Expert Committee on Food Additives. Safety Evaluation of Certain Food Additives. In Proceedings of the Sixty-Third Meeting of the Joint FAO/WHO Expert Committee on Food Additives (JEFCA), Rome, Italy, 22–28 February 2006. [Google Scholar]
- Venkataraman, B.; Almarzooqi, S.; Raj, V.; Dudeja, P.K.; Bhongade, B.A.; Patil, R.B.; Ojha, S.K.; Attoub, S.; Adrian, T.E.; Subramanya, S.B. alpha-Bisabolol Mitigates Colon Inflammation by Stimulating Colon PPAR-gamma Transcription Factor: In Vivo and In Vitro Study. PPAR Res. 2022, 2022, 5498115. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | PMID |
|---|---|---|---|
| Mouse IL-6 | 5′-TGTGTCGTGCTGTTCAGAACC-3′ | 5′-AGGAATCCCGCAATGATGG-3′ | 22326488 |
| Mouse Il-1β | 5′-TCGCTCAGGGTCACAAGAAA-3′ | 5′-CATCAGAGGCAAGGAGGAAAC-3′ | 21735552 |
| Mouse TNF-α | 5′-AGGCTGCCCCGACTACGT-3′ | 5′-GACTTTCTCCTGGTATGAGATAGCAAA-3′ | 21705622 |
| Mouse COX2 | 5′-AACCGCATTGCCTCTGAAT-3′ | 5′-CATGTTCCAGGAGGATGGAG-3′ | 22158945 |
| Mouse iNOS | 5′-CGAAACGCTTCACTTCCAA-3′ | 5′-TGAGCCTATATTGCTGTGGCT-3′ | 22158945 |
| Human COX2 | 5′- CAAATCCTTGCTGTTCCCACCCAT-3′ | 5′-GTGCACTGTGTTTGGAGTGGGTTT-3′ | 25810745 |
| Human CXXL-1 | 5′-GCGGAAAGCTTGCCTCAATC-3′ | 5′-GGTCAGTTGGATTTGTCACTGT-3′ | 25938459 |
| Human IL-8 | 5′-ACTGAGAGTGATTGAGAGTGGAC-3′ | 5′-AACCCTCTGCACCCAGTTTTC-3′ | 31273598 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almarzooqi, S.; Venkataraman, B.; Raj, V.; Alkuwaiti, S.A.A.; Das, K.M.; Collin, P.D.; Adrian, T.E.; Subramanya, S.B. β-Myrcene Mitigates Colon Inflammation by Inhibiting MAP Kinase and NF-κB Signaling Pathways. Molecules 2022, 27, 8744. https://doi.org/10.3390/molecules27248744
Almarzooqi S, Venkataraman B, Raj V, Alkuwaiti SAA, Das KM, Collin PD, Adrian TE, Subramanya SB. β-Myrcene Mitigates Colon Inflammation by Inhibiting MAP Kinase and NF-κB Signaling Pathways. Molecules. 2022; 27(24):8744. https://doi.org/10.3390/molecules27248744
Chicago/Turabian StyleAlmarzooqi, Saeeda, Balaji Venkataraman, Vishnu Raj, Sultan Ali Abdulla Alkuwaiti, Karuna M. Das, Peter D. Collin, Thomas E. Adrian, and Sandeep B. Subramanya. 2022. "β-Myrcene Mitigates Colon Inflammation by Inhibiting MAP Kinase and NF-κB Signaling Pathways" Molecules 27, no. 24: 8744. https://doi.org/10.3390/molecules27248744
APA StyleAlmarzooqi, S., Venkataraman, B., Raj, V., Alkuwaiti, S. A. A., Das, K. M., Collin, P. D., Adrian, T. E., & Subramanya, S. B. (2022). β-Myrcene Mitigates Colon Inflammation by Inhibiting MAP Kinase and NF-κB Signaling Pathways. Molecules, 27(24), 8744. https://doi.org/10.3390/molecules27248744