
AHNAK Contributes to Hepatocellular Carcinoma Growth by Interacting with IGF-1R

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
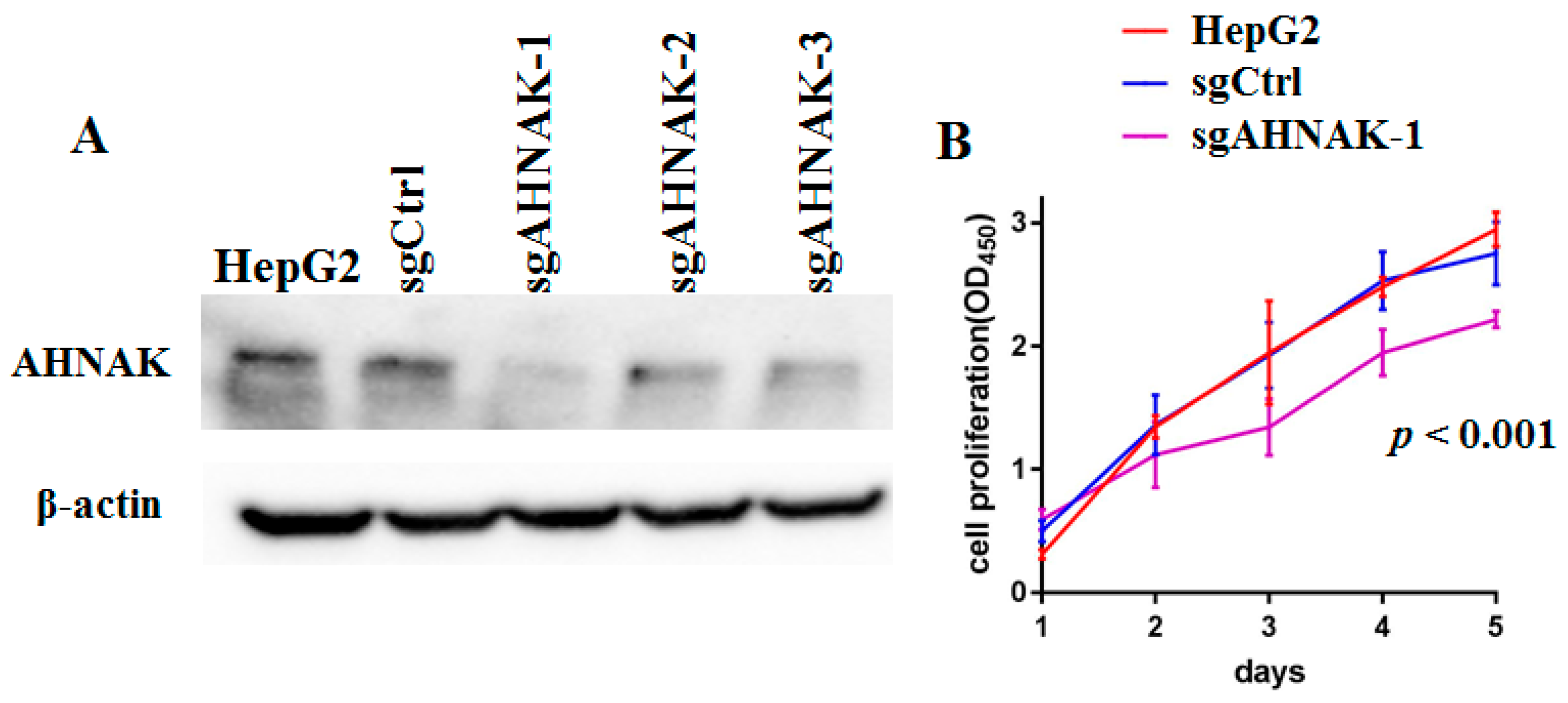
2.1. Specific Knockout of AHNAK Protein Can Inhibit the Proliferation of HepG2 Cells
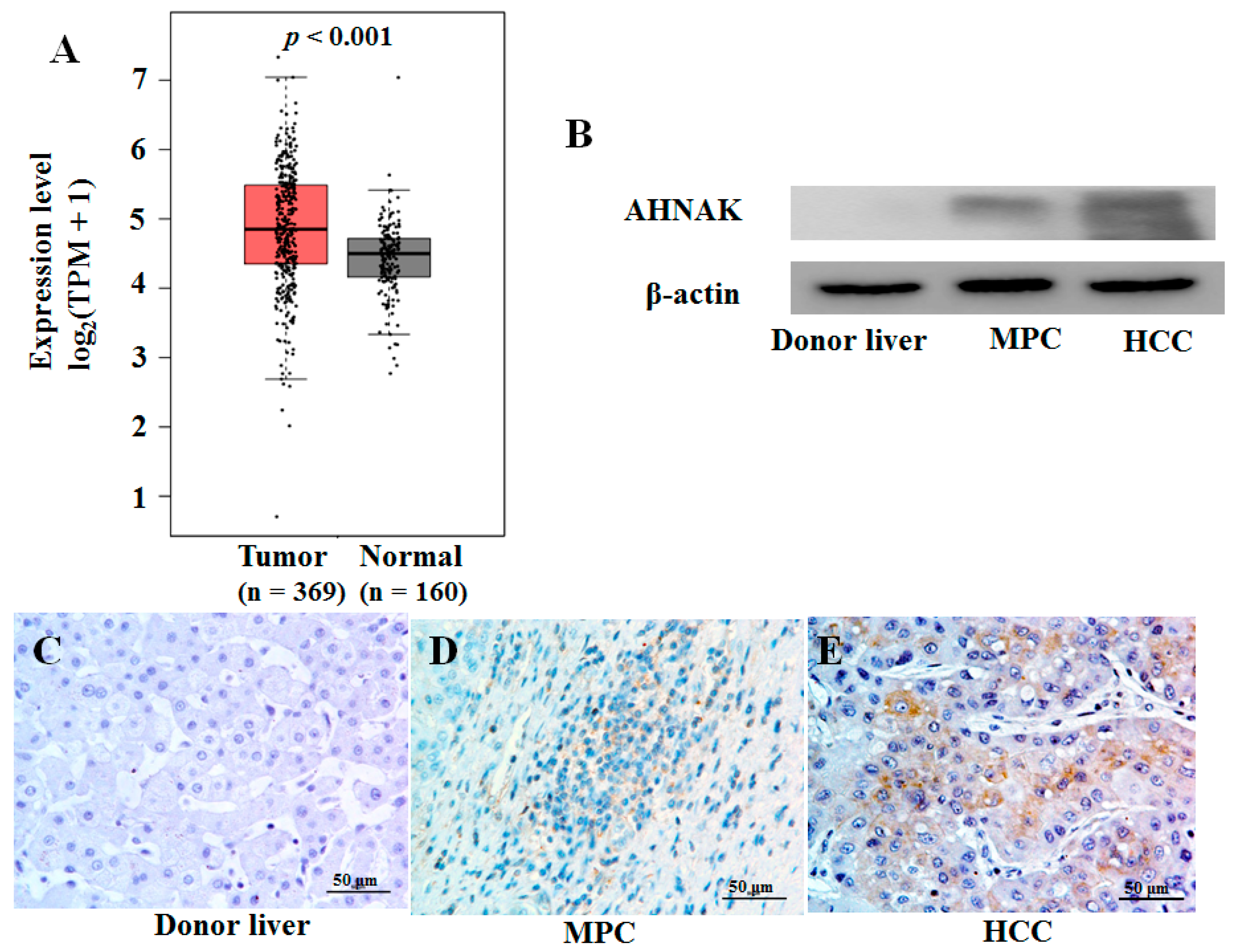
2.2. Expression Analysis of AHNAK Protein in Liver Cancer Tissues
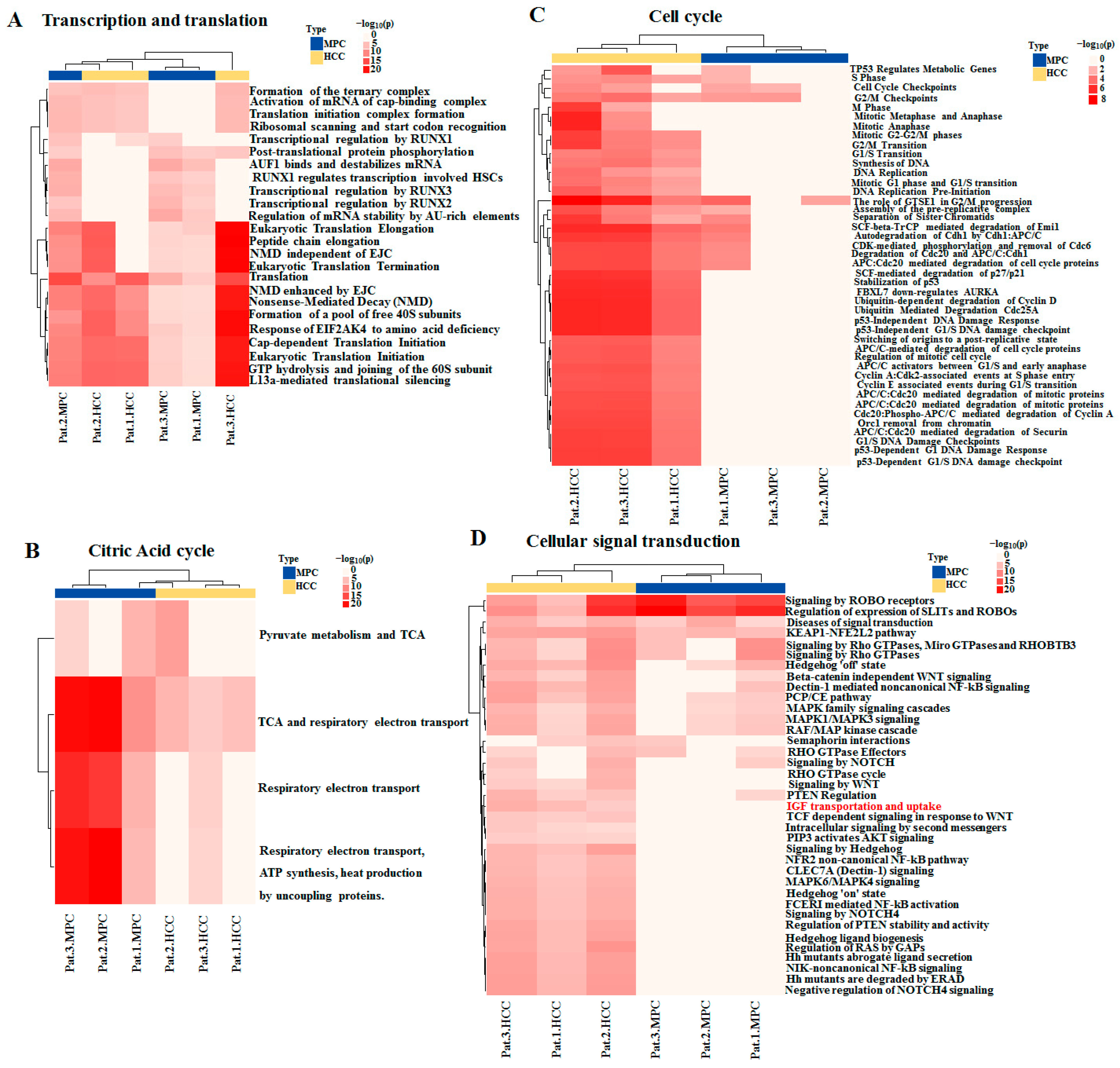
2.3. Identification of AHNAK as a Candidate Involved in IGF-1 Signaling Transduction
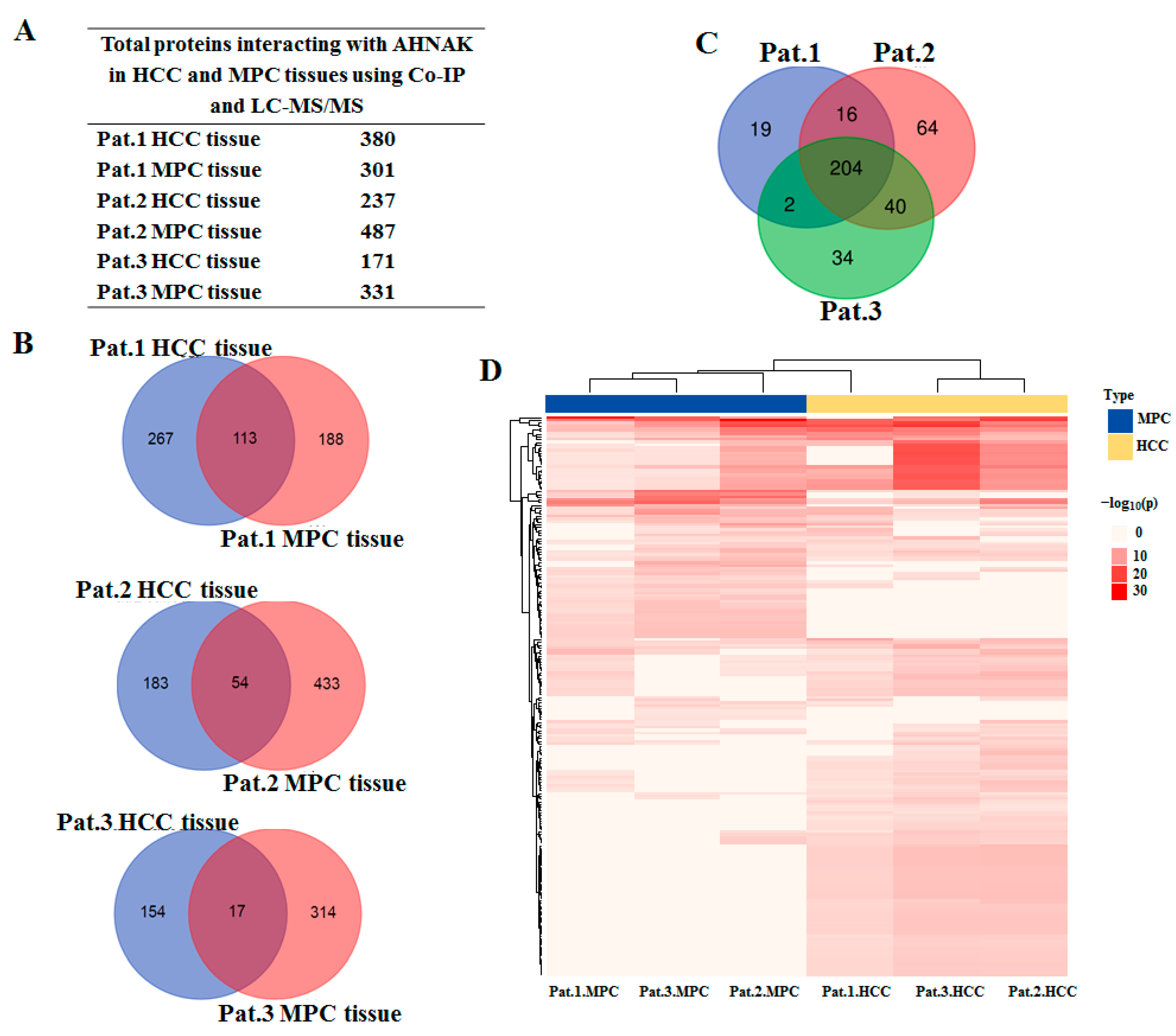
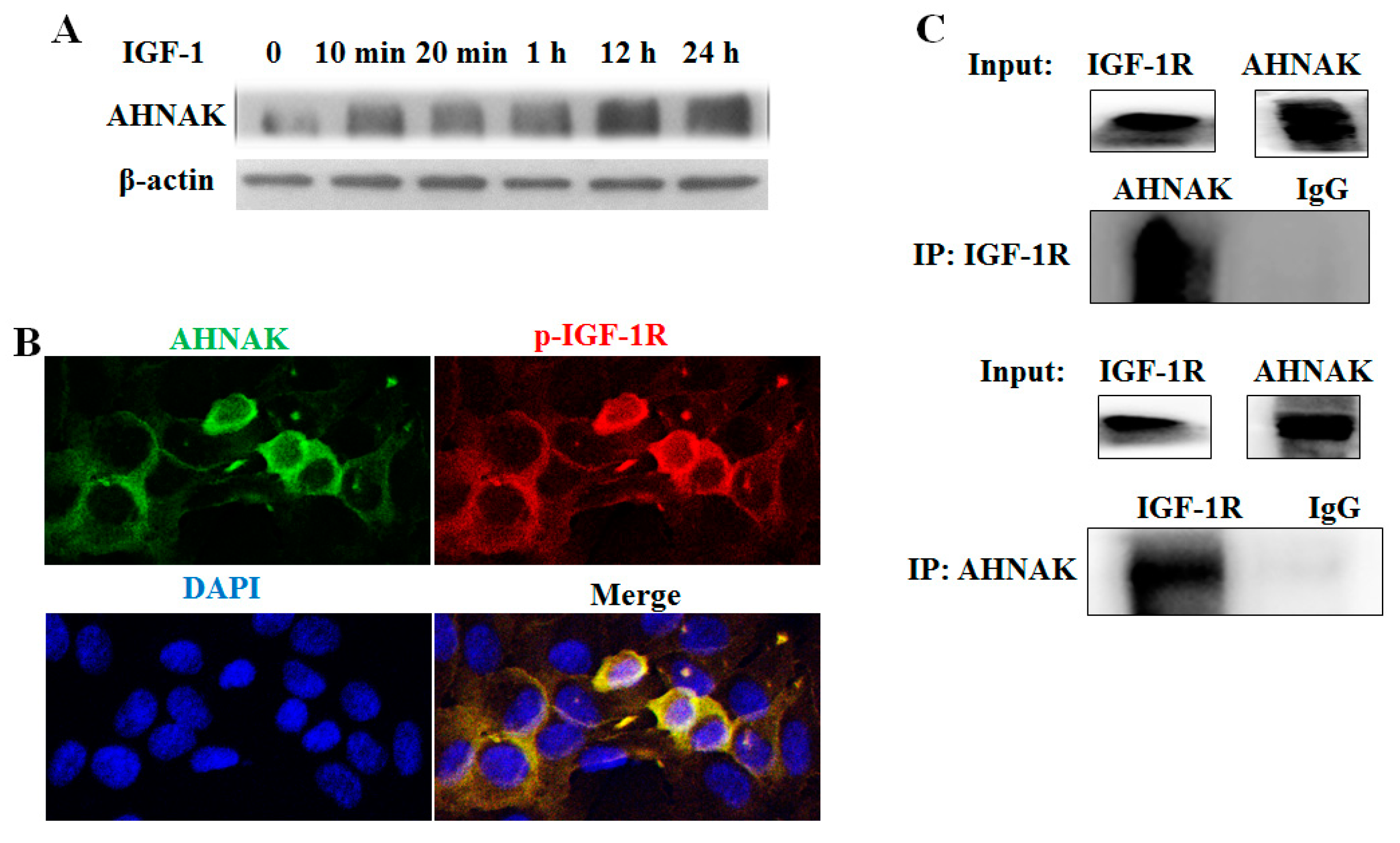
2.4. Identification of AHNAK as a Novel IGF-1R-Interacting Protein
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Tissue Samples
4.3. CoIP for Nano-LC-MS/MS Analysis
4.4. Trypsin Digestion
4.5. Liquid Chromatography-Tandem Mass Analysis
4.6. Protein Identification and Annotation
4.7. Immunoprecipitation (IP) and Western Blotting Analysis
4.8. Immunohistochemistry Array
4.9. SgRNA Plasmid Constructs and Generation of a Stable AHNAK-Knockout HepG2 Cell Line
4.10. Cell Viability Assays
4.11. Confocal Immunofluorescence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive analysis of normal adjacent to tumor transcriptomes. Nat. Commun. 2017, 8, 017–01027. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.F.; Ribeiro-Dos-Santos, A.M.; Vinasco-Sandoval, T.; Magalhães, L.; Pinto, P.; Anaissi, A.K.M.; Demachki, S.; de Assumpção, P.P.; Dos Santos, S.E.B.; Ribeiro-Dos-Santos, Â. The comprehensive expression analysis of circular RNAs in gastric cancer and its association with field cancerization. Sci. Rep. 2017, 7, 15061. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, L.; Ribeiro-dos-Santos, A.M.; Cruz, R.L.; Nakamura, K.D.d.M.; Brianese, R.; Burbano, R.; Ferreira, S.P.; Oliveira, E.L.F.d.; Anaissi, A.K.M.; Nahúm, M.C.d.S.; et al. Triple-Negative Breast Cancer circRNAome Reveals Hsa_circ_0072309 as a Potential Risk Biomarker. Cancers 2022, 14, 3280. [Google Scholar] [CrossRef]
- Amagai, M. A mystery of AHNAK/desmoyokin still goes on. J. Investig. Dermatol. 2004, 123, xiv–xv. [Google Scholar] [CrossRef]
- Davis, T.A.; Loos, B.; Engelbrecht, A.M. Corrigendum to AHNAK: The giant jack of all trades. Cell Signal. 2014, 26, 2683–2693, Erratum in Cell Signal. 2015, 27, 187–188. [Google Scholar] [CrossRef]
- Chen, B.; Wang, J.; Dai, D.; Zhou, Q.; Guo, X.; Tian, Z.; Huang, X.; Yang, L.; Tang, H.; Xie, X. AHNAK suppresses tumour proliferation and invasion by targeting multiple pathways in triple-negative breast cancer. J. Exp. Clin. Cancer Res. 2017, 36, 0522. [Google Scholar] [CrossRef]
- Cai, Y.; Hu, Y.; Yu, F.; Tong, W.; Wang, S.; Sheng, S.; Zhu, J. AHNAK suppresses ovarian cancer progression through the Wnt/β-catenin signaling pathway. Aging 2021, 13, 23579–23587. [Google Scholar] [CrossRef]
- Cho, W.C.; Jang, J.E.; Kim, K.H.; Yoo, B.C.; Ku, J.L. SORBS1 serves a metastatic role via suppression of AHNAK in colorectal cancer cell lines. Int. J. Oncol. 2020, 56, 1140–1151. [Google Scholar] [CrossRef]
- Sheppard, H.M.; Feisst, V.; Chen, J.; Print, C.; Dunbar, P.R. AHNAK is downregulated in melanoma, predicts poor outcome, and may be required for the expression of functional cadherin-1. Melanoma Res. 2016, 26, 108–116. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Huang, R.; Liang, Z.; Liu, T. Upregulation of nucleoprotein AHNAK is associated with poor outcome of pancreatic ductal adenocarcinoma prognosis via mediating epithelial-mesenchymal transition. J. Cancer 2019, 10, 3860–3870. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, C.A.; Bankfalvi, A.; Gu, X.; Zeidler, R.; Brandau, S.; Lang, S. AHNAK and inflammatory markers predict poor survival in laryngeal carcinoma. PLoS ONE 2013, 8, e56420. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.; Shin, S.; Yoo, J.-Y.; Goh, Y.; Lee, I.H.; Bae, Y.S. Ahnak promotes tumor metastasis through transforming growth factor-β-mediated epithelial-mesenchymal transition. Sci. Rep. 2018, 8, 14379. [Google Scholar] [CrossRef] [PubMed]
- Sudo, H.; Tsuji, A.B.; Sugyo, A.; Abe, M.; Hino, O.; Saga, T. AHNAK is highly expressed and plays a key role in cell migration and invasion in mesothelioma. Int. J. Oncol. 2014, 44, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, K.; Liu, G.; Xu, Y.; Zhang, A.; Lin, D.; Zhang, H.; Zhao, X.; Jin, B.; Li, N.; et al. Distinctive pattern of AHNAK methylation level in peripheral blood mononuclear cells and the association with HBV-related liver diseases. Cancer Med. 2018, 7, 5178–5186. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; He, M.; Chan, A.W.H.; Song, Q.X.; Sze, S.C.; Chen, H.; Man, M.K.H.; Man, K.; Chan, S.L.; Lai, P.B.S.; et al. Genomic and Epigenomic Features of Primary and Recurrent Hepatocellular Carcinomas. Gastroenterology 2019, 157, 1630–1645.e6. [Google Scholar] [CrossRef]
- Shimizu, M.; Shirakami, Y.; Imai, K.; Takai, K.; Moriwaki, H. Acyclic retinoid in chemoprevention of hepatocellular carcinoma: Targeting phosphorylated retinoid X receptor-α for prevention of liver carcinogenesis. J. Carcinog. 2012, 11, 1477–3163. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.-Y.; Suzuki, H.; Honda, M.; Okada, H.; Kaneko, S.; Inoue, I.; Ebisui, E.; Hashimoto, K.; Carninci, P.; Kanki, K.; et al. Prevention of hepatocellular carcinoma by targeting MYCN-positive liver cancer stem cells with acyclic retinoid. Proc. Natl. Acad. Sci. USA 2018, 115, 4969–4974. [Google Scholar] [CrossRef]
- Han, H.; Kursula, P. Periaxin and AHNAK Nucleoprotein 2 Form Intertwined Homodimers through Domain Swapping. J. Biol. Chem. 2014, 289, 14121–14131. [Google Scholar] [CrossRef]
- Ligensa, T.; Krauss, S.; Demuth, D.; Schumacher, R.; Camonis, J.; Jaques, G.; Weidner, K.M. A PDZ Domain Protein Interacts with the C-terminal Tail of the Insulin-like Growth Factor-1 Receptor but Not with the Insulin Receptor. J. Biol. Chem. 2001, 276, 33419–33427. [Google Scholar] [CrossRef]
- Lee, H.; Kim, K.; Woo, J.; Park, J.; Kim, H.; Lee, K.E.; Kim, Y.; Moon, K.C.; Kim, J.Y.; Park, I.A.; et al. Quantitative Proteomic Analysis Identifies AHNAK (Neuroblast Differentiation-associated Protein AHNAK) as a Novel Candidate Biomarker for Bladder Urothelial Carcinoma Diagnosis by Liquid-based Cytology. Mol. Cell. Proteoms. 2018, 17, 1788–1802. [Google Scholar] [CrossRef]
- Shtivelman, E.; Cohen, F.E.; Bishop, J.M. A human gene (AHNAK) encoding an unusually large protein with a 1.2-microns polyionic rod structure. Proc. Natl. Acad. Sci. USA 1992, 89, 5472–5476. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. Field cancerization in oral stratified squamous epithelium; clinical implications of multicentric origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, E.; Thorn, G.J.; Ross-Adams, H.; Jones, L.J.; Chelala, C. Field cancerization in breast cancer. J. Pathol. 2022, 257, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lu, W.; Garcia-Prieto, C.; Huang, P. The Warburg effect and its cancer therapeutic implications. J. Bioenerg. Biomembr. 2007, 39, 267–274. [Google Scholar] [CrossRef]
- Ghodke, I.; Remisova, M.; Furst, A.; Kilic, S.; Reina-San-Martin, B.; Poetsch, A.R.; Altmeyer, M.; Soutoglou, E. AHNAK controls 53BP1-mediated p53 response by restraining 53BP1 oligomerization and phase separation. Mol. Cell 2021, 81, 2596–2610.e7. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Mao, W.; Ren, W.; Xu, F.; Zhu, Q.; Lu, C.; Lin, Z.; Zhang, Z.; Chu, Y.; Liu, R.; et al. Ubiquitin-protein ligase E3C maintains non-small-cell lung cancer stemness by targeting AHNAK-p53 complex. Cancer Lett. 2019, 443, 125–134. [Google Scholar] [CrossRef]
- Cao, Z.; Liu, L.Z.; Dixon, D.A.; Zheng, J.Z.; Chandran, B.; Jiang, B.H. Insulin-like growth factor-I induces cyclooxygenase-2 expression via PI3K, MAPK and PKC signaling pathways in human ovarian cancer cells. Cell. Signal. 2007, 19, 1542–1553. [Google Scholar] [CrossRef]
- Na, L.; Bai, Y.; Sun, Y.; Wang, Z.; Wang, W.; Yuan, L.; Zhao, C. Identification of 9-Core Immune-Related Genes in Bladder Urothelial Carcinoma Prognosis. Front. Oncol. 2020, 10, 1142. [Google Scholar] [CrossRef]
- Chang, T.S.; Wu, Y.C.; Chi, C.C.; Su, W.C.; Chang, P.J.; Lee, K.F.; Tung, T.H.; Wang, J.; Liu, J.J.; Tung, S.Y.; et al. Activation of IL6/IGFIR confers poor prognosis of HBV-related hepatocellular carcinoma through induction of OCT4/NANOG expression. Clin. Cancer Res. 2015, 21, 201–210. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Song, K.; Hou, Y.; Tian, Y.; Wang, H.; Sun, L.; Li, A.; Zhang, Y. AHNAK Contributes to Hepatocellular Carcinoma Growth by Interacting with IGF-1R. Molecules 2022, 27, 8680. https://doi.org/10.3390/molecules27248680
Li K, Song K, Hou Y, Tian Y, Wang H, Sun L, Li A, Zhang Y. AHNAK Contributes to Hepatocellular Carcinoma Growth by Interacting with IGF-1R. Molecules. 2022; 27(24):8680. https://doi.org/10.3390/molecules27248680
Chicago/Turabian StyleLi, Kang, Ke Song, Yuli Hou, Yuan Tian, Huijuan Wang, Libo Sun, Ang Li, and Yonghong Zhang. 2022. "AHNAK Contributes to Hepatocellular Carcinoma Growth by Interacting with IGF-1R" Molecules 27, no. 24: 8680. https://doi.org/10.3390/molecules27248680
APA StyleLi, K., Song, K., Hou, Y., Tian, Y., Wang, H., Sun, L., Li, A., & Zhang, Y. (2022). AHNAK Contributes to Hepatocellular Carcinoma Growth by Interacting with IGF-1R. Molecules, 27(24), 8680. https://doi.org/10.3390/molecules27248680