
Neocinnamomum caudatum Essential Oil Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Cells by Inhibiting NF-κB Activation and ROS Production

 ,
,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
2.1. Volatile Profiling of N. caudatum Essential Oil
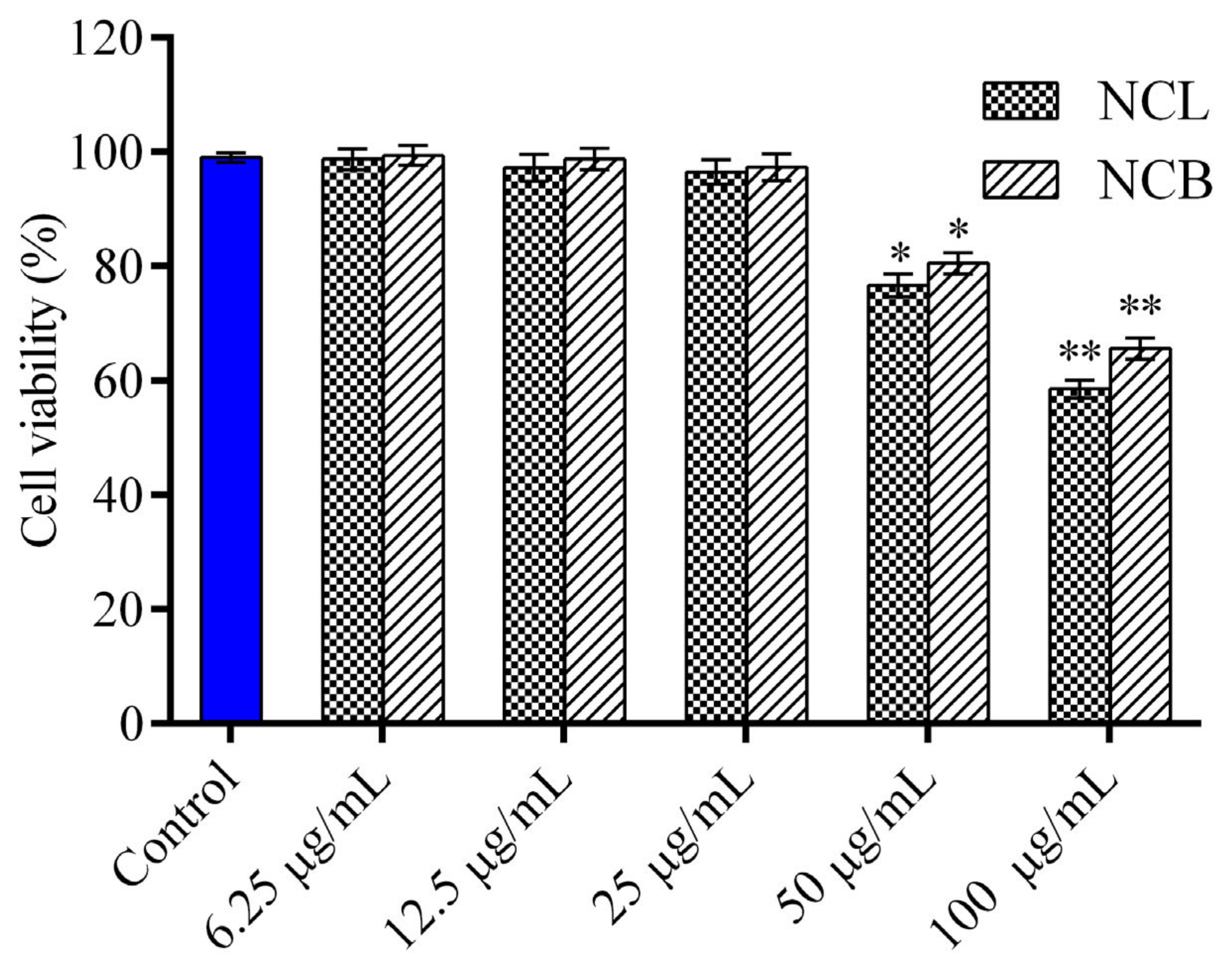
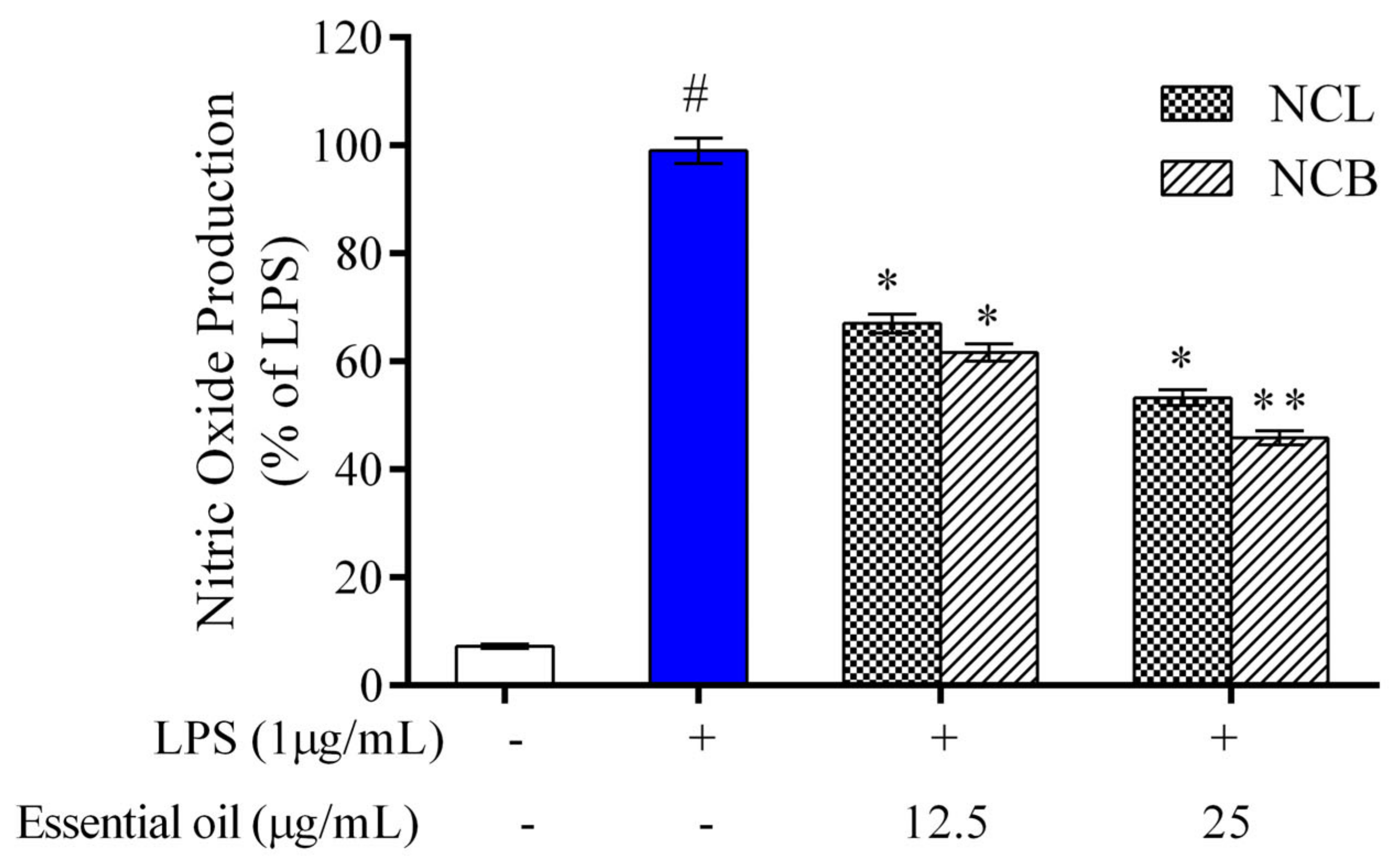
2.2. Effect of N. caudatum Essential Oil on Cell Viability and NO Production of RAW 264.7 Cells
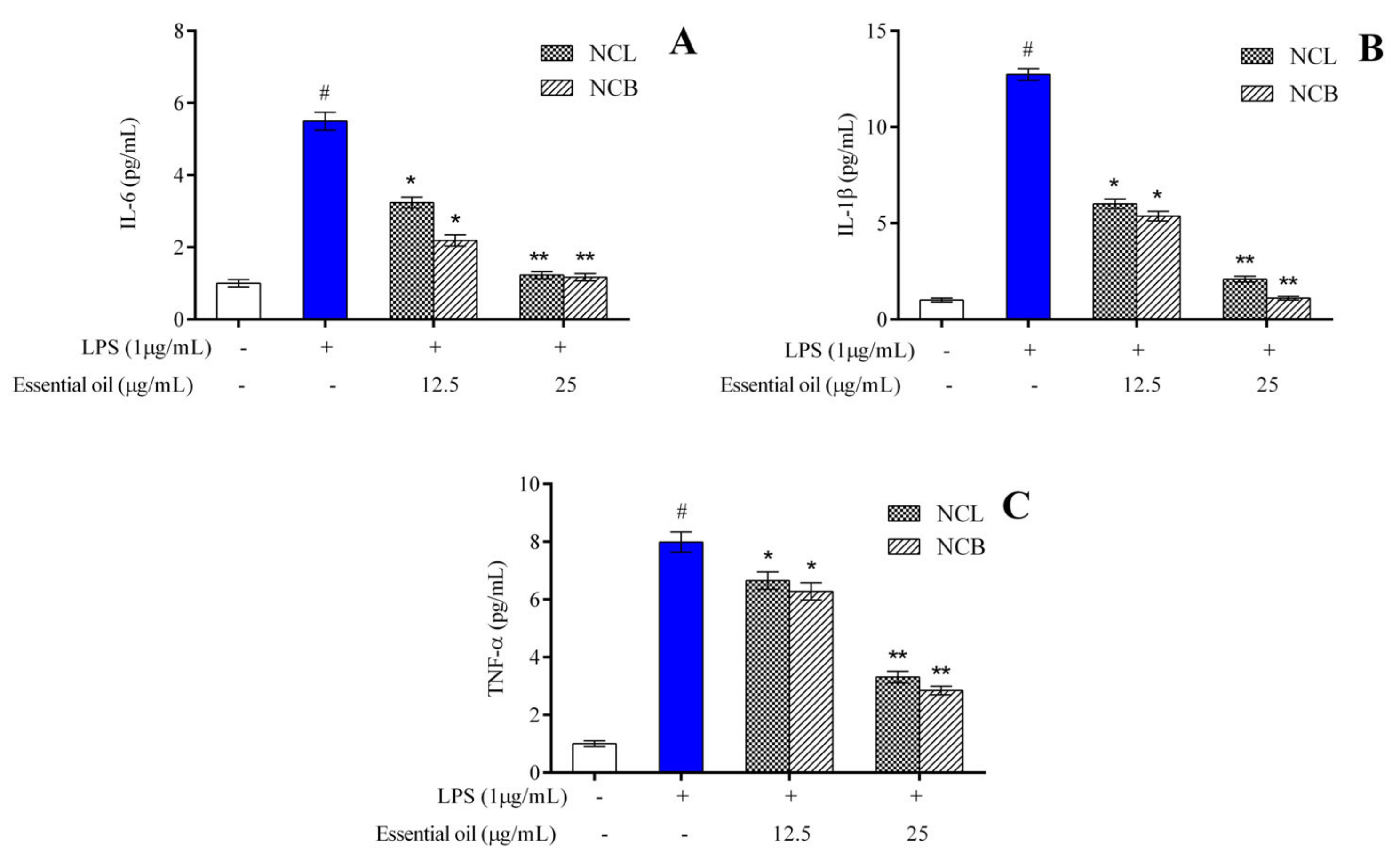
2.3. Effect of N. caudatum Essential Oil on Proinflammatory Cytokines
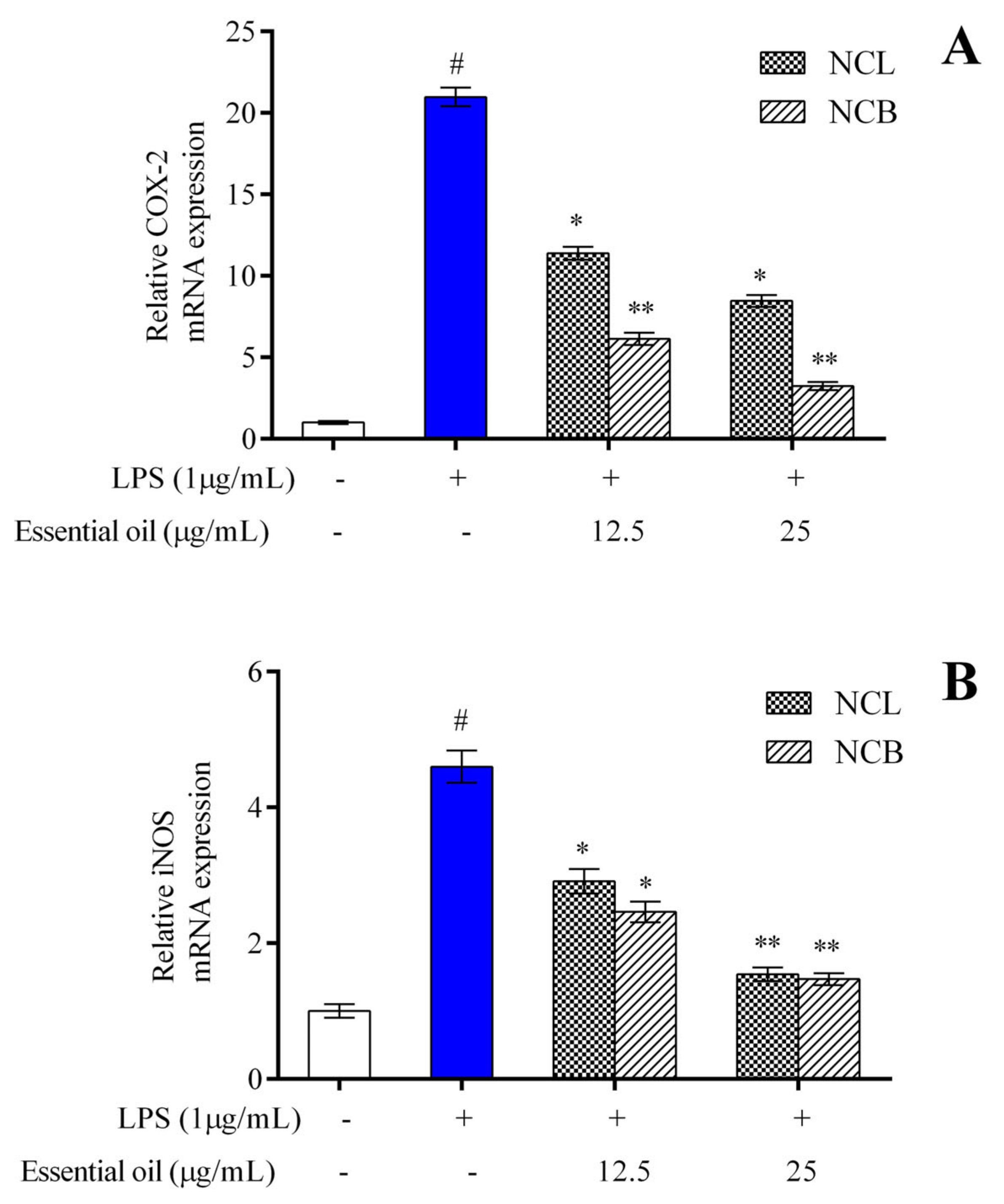
2.4. Effect of N. caudatum Essential Oil on LPS-Induced iNOS and COX-2 mRNA Expression
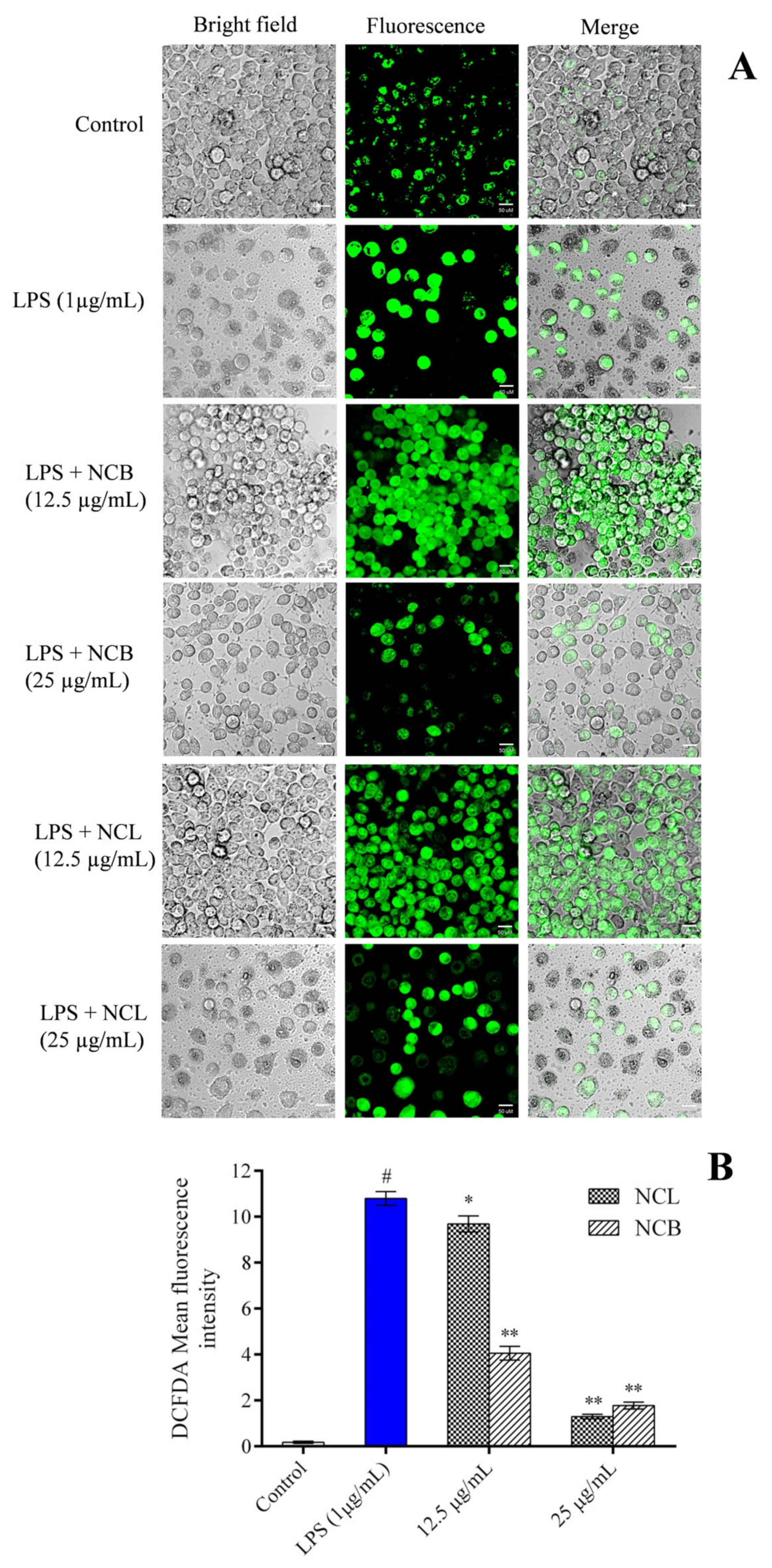
2.5. Effect of N. caudatum Essential Oil on LPS-Induced Reactive Oxygen Species (ROS) Levels
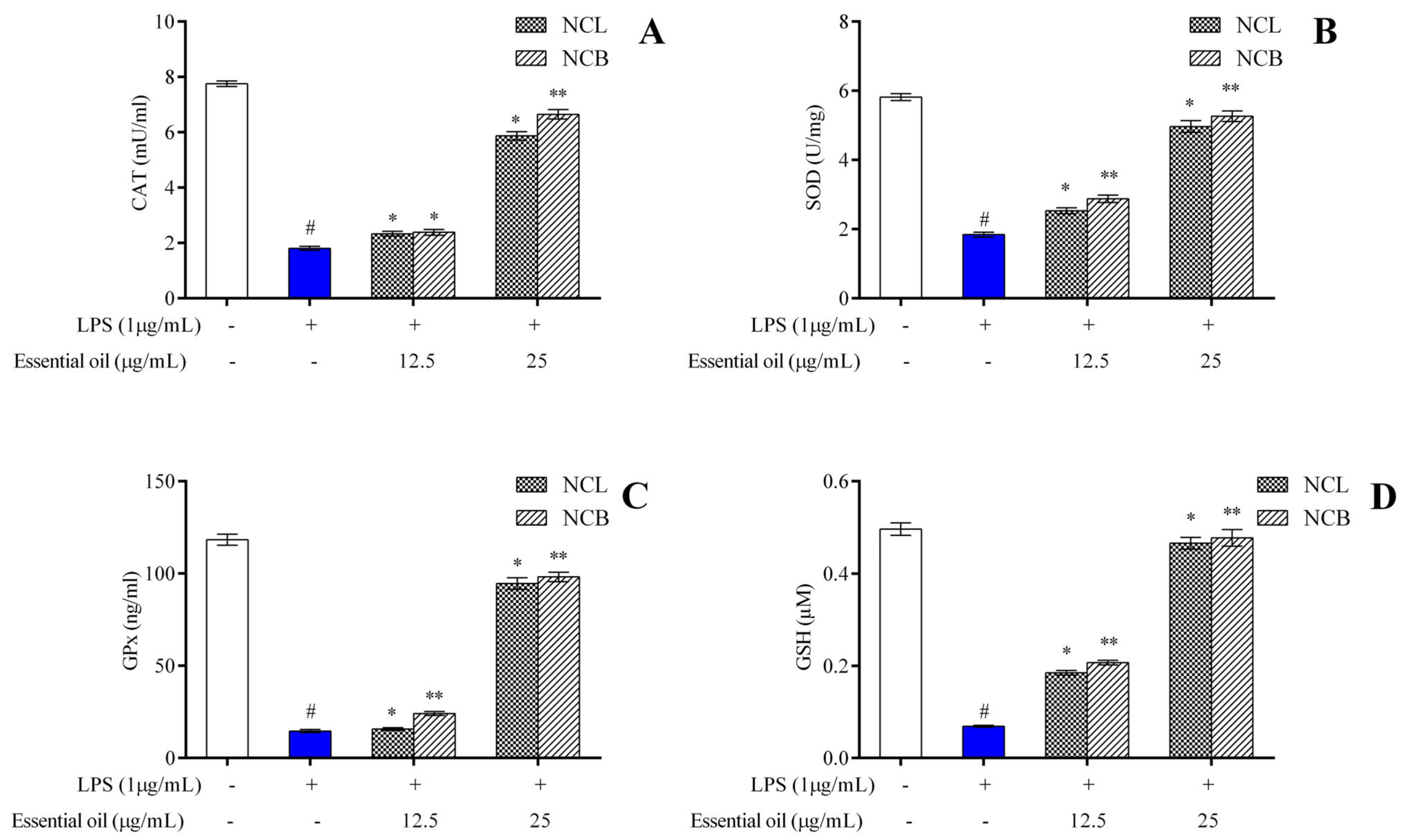
2.6. Effect of N. caudatum Essential Oil on LPS-Induced Antioxidant Enzymes in RAW 264.7 Cells
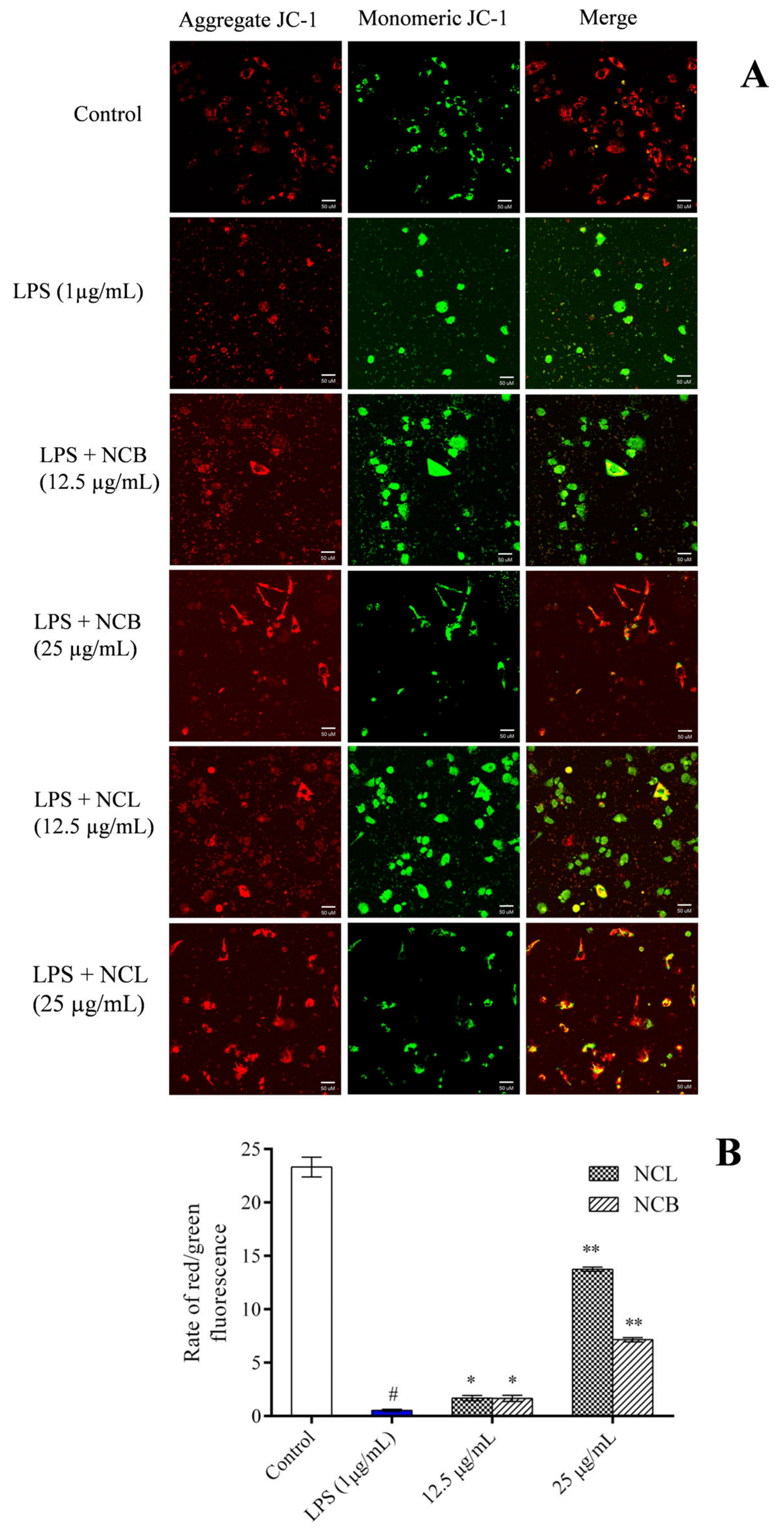
2.7. Effect of N. caudatum Essential Oil on Mitochondrial Membrane Potential
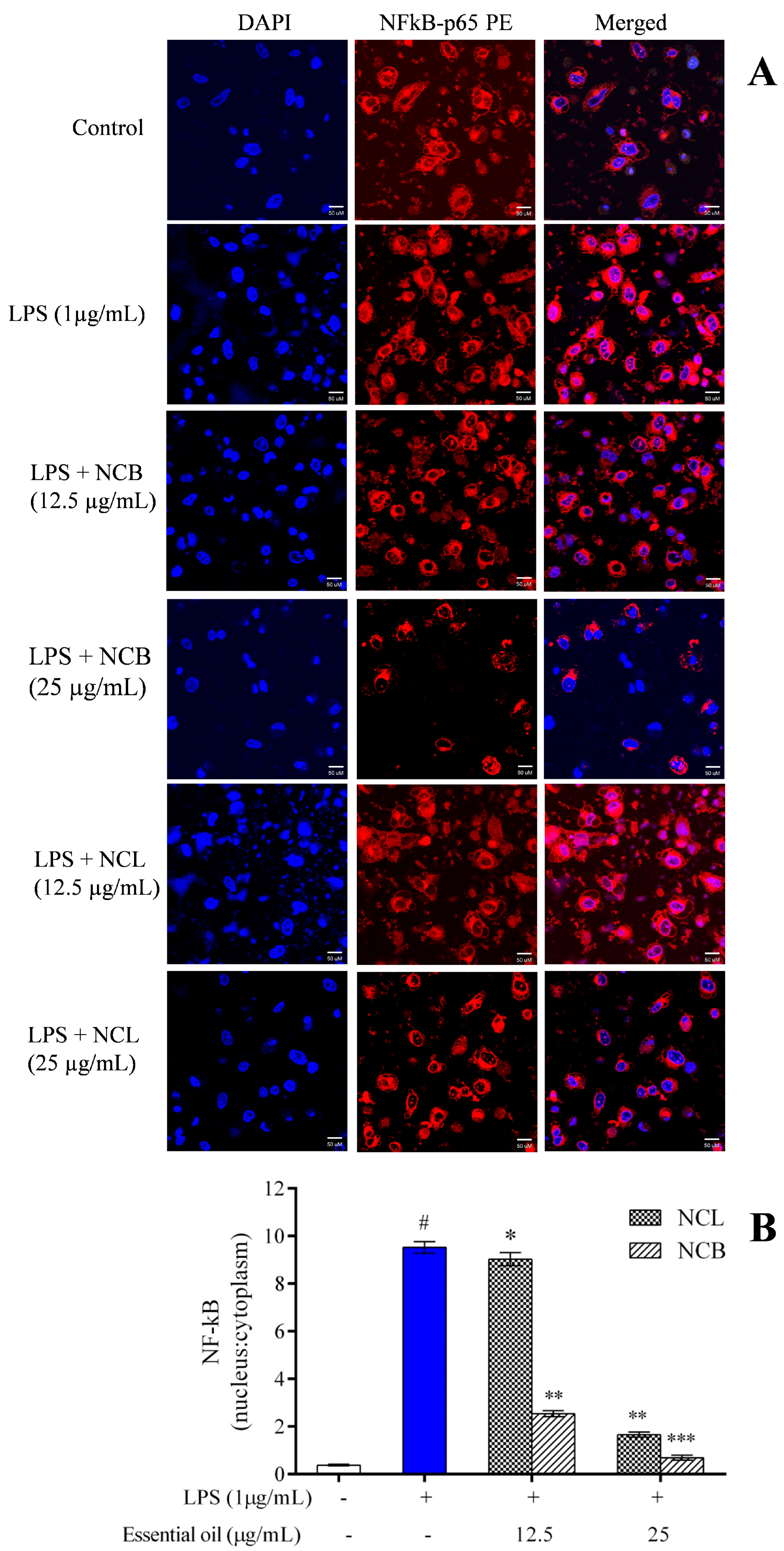
2.8. Effect of N. caudatum Essential Oil on LPS-Induced NF-κB Activation
2.9. Differential Anti-Inflammatory Activity of Bark and Leaf Essential Oil of N. caudatum
3. Materials and Methods
3.1. Essential Oil Extraction
3.2. Chemical Characterization of Essential Oil
3.3. Cytotoxicity Assay
3.4. Determination of Nitric Oxide Production
3.5. Estimation of Proinflammatory Cytokines
3.6. mRNA Expression of iNOS and COX-2
3.7. Estimation of Reactive Oxygen Species (ROS) Levels
3.8. Estimation of Endogenous Antioxidant Enzymes
3.9. Estimation of Mitochondrial Membrane Potential
3.10. Assessment of NF-κB Nuclear Translocation
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Fürst, R.; Zündorf, I. Plant-derived anti-inflammatory compounds: Hopes and disappointments regarding the translation of preclinical knowledge into clinical progress. Mediat. Inflamm. 2014, 2014, 146832. [Google Scholar] [CrossRef]
- Wan, P.; Xie, M.; Chen, G.; Dai, Z.; Hu, B.; Zeng, X.; Sun, Y. Anti-inflammatory effects of dicaffeoylquinic acids from Ilex kudingcha on lipopolysaccharide-treated RAW264.7 macrophages and potential mechanisms. Food Chem. Toxicol. 2019, 126, 332–342. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Neuman, M.G. Immune dysfunction in inflammatory bowel disease. Transl. Res. 2007, 149, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Zou, Y.; Peng, J. Oregano essential oil attenuates RAW264.7 cells from lipopolysaccharide-induced inflammatory response through regulating NADPH oxidase activation-driven oxidative stress. Molecules 2018, 23, 1857. [Google Scholar] [CrossRef]
- Yoon, H.J.; Moon, M.E.; Park, H.S.; Im, S.Y.; Kim, Y.H. Chitosan oligosaccharide (COS) inhibits LPS-induced inflammatory effects in RAW 264.7 macrophage cells. Biochem. Biophys. Res. Commun. 2007, 358, 954–959. [Google Scholar] [CrossRef] [PubMed]
- De Cássia da Silveira e Sá, R.; Andrade, L.N.; De Sousa, D.P. Sesquiterpenes from essential oils and anti-inflammatory activity. Nat. Prod. Commun. 2015, 10, 1767–1774. [Google Scholar] [CrossRef]
- Cagiola, M.; Giulio, S.; Miriam, M.; Katia, F.; Paola, P.; Macrì, A.; Pasquali, P. In vitro down regulation of proinflammatory cytokines induced by LPS tolerance in pig CD14+ cells. Vet. Immunol. Immunopathol. 2006, 112, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Boscá, L.; Zeini, M.; Través, P.G.; Hortelano, S. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Ko, Y.J.; Yang, H.M.; Ham, Y.M.; Roh, S.W.; Jeon, Y.J.; Ahn, G.; Kang, M.C.; Yoon, W.J.; Kim, D.; et al. Anti-inflammatory effect of essential oil and its constituents from fingered citron (Citrus medica L. var. sarcodactylis) through blocking JNK, ERK and NF-κB signaling pathways in LPS-activated RAW 264.7 cells. Food Chem. Toxicol. 2013, 57, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Triadafilopoulos, G. Epidemiology of NSAID induced gastrointestinal complications. J. Rheumatol. 1999, 56, 18–24. [Google Scholar]
- Gambaro, G.; Perazella, M.A. Adverse renal effects of anti-inflammatory agents: Evaluation of selective and nonselective cyclooxygenase inhibitors. J. Intern. Med. 2003, 253, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, G.K.; Heyneman, C.A. Nephrotoxic potential of selective cyclooxygenase-2 inhibitors. Ann. Pharmacother. 2004, 38, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Kummer, C.L.; Coelho, T.C.R. Cycloxygenase-2 inhibitors nonsteroid anti-inflammatory drugs: Current issues. Rev. Bras. Anestesiol. 2002, 52, 498–512. [Google Scholar]
- Lucas, G.N.C.; Leitão, A.C.C.; Alencar, R.L.; Xavier, R.M.F.; Daher, E.D.F.; Silva, G.B.D. Pathophysiological aspects of nephropathy caused by non-steroidal anti-inflammatory drugs. Braz. J. Nephrol. 2018, 41, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Kostermans, A.J. A monograph of the genus Neocinnamomum Liou Ho. Reinwardtia 1974, 9, 85–96. [Google Scholar]
- Rohwer, J.G. Lauraceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Rohwer, J.G., Bittrich, V., Eds.; Springer: Berlin, Germany, 1993; Volume 2, pp. 366–391. [Google Scholar]
- Li, X.-W.; Li, J.; van der Werff, H. Neocinnamomum H. Liou. In Flora of China; Wu, Z.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2008; Volume 7, pp. 187–189. [Google Scholar]
- Wu, Z. Flora Yunnanica; Science Press: Beijing, China, 1983; Volume 3. [Google Scholar]
- Doley, B.; Gajurel, P.R.; Rethy, P.; Singh, B.; Hazarika, H. Ethnomedicinal uses of different species of Cinnamomum Schaeffer (Lauraceae) by ethnic communities in Arunachal Pradesh, India. Pleione 2009, 3, 9–12. [Google Scholar]
- Wang, Z.; Yang, J.; Tan, Y.; Hu, G.; Long, C. Comprehensive evaluation of woody oil-bearing plants in Yunnan as sources for biodiesel. Plant Divers. Resour. 2013, 35, 630–640. [Google Scholar]
- Naldarine, M.M. Ethnomedicinal Plants in West Garo Hills District in Meghalaya. Ph.D. Thesis, Mizoram University, Aizawl, India, 2018. [Google Scholar]
- Singh, B.; Borthakur, S.K. Forest issues and challenges in protected area management: A case study from Himalayan Nokrek National Park and Biosphere Reserve. Int. J. Conserv. Sci. 2015, 6, 233–252. [Google Scholar]
- Singh, B.; Sinha, B.K.; Phukan, S.J.; Borthakur, S.K.; Singh, V.N. Wild edible plants used by Garo tribes of Nokrek Biosphere Reserve in Meghalaya, India. Indian J. Tradit. Knowl. 2012, 11, 166–171. [Google Scholar]
- Sherub, K.; Singh, A.P. Notes on the food and feeding habits of rufous-necked hornbill Aceros nipalensis in Zhemgang district, Bhutan. J. Bombay Nat. Hist. Soc. 2020, 117, 50–53. [Google Scholar] [CrossRef]
- Altschul, S.V.R. Drugs and Foods from Little-Known Plants: Notes in Harvard University Herbaria; Harvard University Press: Cambridge, MA, USA, 1973. [Google Scholar]
- Bharali, P.; Sharma, C.L.; Singh, B.; Sharma, M. Ethnobotanical studies of spice and condiment plant used by some communities of Assam. Int. J. Adv. Sci. Res. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Gan, Y.; Song, Y.; Chen, Y.; Liu, H.; Yang, D.; Xu, Q.; Zheng, Z. Transcriptome analysis reveals a composite molecular map linked to unique seed oil profile of Neocinnamomum caudatum (Nees) Merr. BMC Plant Biol. 2018, 18, 303. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.F.; Hsu, H.Y.; Cheng, S.S.; Liu, J.Y.; Chang, S.T. Study on the antiinflammatory activity of essential oil from leaves of Cinnamomum osmophloeum. J. Agric. Food Chem. 2005, 53, 7274–7278. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Chen, C.J.; Lin, T.Y.; Tung, J.C.; Wang, S.Y. Anti-inflammation activity of fruit essential oil from Cinnamomum insularimontanum Hayata. Bioresour. Technol. 2008, 99, 8783–8787. [Google Scholar] [CrossRef]
- Lee, Y.S.; Son, E.; Kim, S.H.; Lee, Y.M.; Kim, O.S.; Kim, D.S. Synergistic uric acid-lowering effects of the combination of Chrysanthemum indicum linne flower and Cinnamomum cassia (L.) J. Persl bark extracts. Evid. Based Complement. Altern. Med. 2017, 2017, 9764843. [Google Scholar] [CrossRef] [PubMed]
- Azab, S.S.; Abdel Jaleel, G.A.; Eldahshan, O.A. Anti-inflammatory and gastroprotective potential of leaf essential oil of Cinnamomum glanduliferum in ethanol-induced rat experimental gastritis. Pharm. Biol. 2017, 55, 1654–1661. [Google Scholar] [CrossRef]
- Han, X.; Parker, T.L. Antiinflammatory activity of cinnamon (Cinnamomum zeylanicum) bark essential oil in a human skin disease model. Phytother. Res. 2017, 31, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Sun, W.; Ke, C.; Wang, F.; Xue, Y.; Luo, Z.; Wang, X.; Zhang, J.; Zhang, Y. Anti-inflammatory activities of leaf oil from Cinnamomum subavenium in vitro and in vivo. Biomed Res. Int. 2019, 2019, 1823149. [Google Scholar] [CrossRef]
- Tung, Y.T.; Chua, M.T.; Wang, S.Y.; Chang, S.T. Ant-inflammation activities of essential oil and its constituents from indigenous cinnamon (Cinnamomum osmophloeum) twigs. Bioresour. Technol. 2008, 99, 3908–3913. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Yu, H.; Xie, Y.; Guo, Y.; Fan, J.; Yao, W. The anti-inflammatory potential of Cinnamomum camphora (L.) J. Presl essential oil in vitro and in vivo. J. Ethnopharmacol. 2021, 267, 113516. [Google Scholar] [CrossRef] [PubMed]
- Budiastuti; Nurcholida, R.D.; Primaharinastiti, R.; Sukardiman. Anti-inflammatory activity of Cinnamon bark oil (Cinnamomum burmannii (Nees & T. Nees) Blume from Lombok, Timur, Indonesia. Pharmacogn. J. 2021, 13, 1005–1013. [Google Scholar]
- Esmaeili, F.; Zahmatkeshan, M.; Yousefpoor, Y.; Alipanah, H.; Safari, E.; Osanloo, M. Anti-inflammatory and anti-nociceptive effects of Cinnamon and Clove essential oils nanogels: An in vivo study. BMC Complement. Med. Ther. 2022, 22, 143. [Google Scholar] [CrossRef] [PubMed]
- Pannee, C.; Chandhanee, I.; Wacharee, L. Anti-inflammatory effects of essential oil from the leaves of Cinnamomum cassia and cinnamaldehyde on lipopolysaccharide-stimulated J774A.1 cells. J. Adv. Pharm. Technol. Res. 2014, 5, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.C.; Siani, A.C.; Ramos, M.F.S.; Menezes-de-Lima, O., Jr.; Henriques, M.G.M.O. Evaluation of anti-inflammatory activity of essential oils from two Asteraceae species. Pharmazie 2003, 58, 582–586. [Google Scholar]
- Neves, A.; Rosa, S.; Gonçalves, J.; Rufino, A.; Judas, F.; Salgueiro, L.; Lopes, M.C.; Cavaleiro, C.; Mendes, A.F. Screening of five essential oils for identification of potential inhibitors of IL-1-induced Nf-κB activation and NO production in human chondrocytes: Characterization of the inhibitory activity of α-pinene. Planta Med. 2010, 76, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Bae, G.S.; Park, K.C.; Choi, S.B.; Jo, I.J.; Choi, M.O.; Hong, S.H.; Song, K.; Song, H.J.; Park, S.J. Protective effects of alpha-pinene in mice with cerulein-induced acute pancreatitis. Life Sci. 2012, 91, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Lee, H.J.; Jeon, Y.D.; Han, Y.H.; Kee, J.Y.; Kim, H.J.; Shin, H.J.; Kang, J.; Lee, B.S.; Kim, S.H.; et al. Alpha-pinene exhibits anti-inflammatory activity through the suppression of MAPKs and the NF-κB pathway in mouse peritoneal macrophages. Am. J. Chin. Med. 2015, 43, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Rufino, A.T.; Ribeiro, M.; Sousa, C.; Judas, F.; Salgueiro, L.; Cavaleiro, C.; Mendes, A.F. Evaluation of the anti-inflammatory, anti-catabolic and pro-anabolic effects of E-caryophyllene, myrcene and limonene in a cell model of osteoarthritis. Eur. J. Pharmacol. 2015, 750, 141–150. [Google Scholar] [CrossRef]
- Li, X.J.; Yang, Y.J.; Li, Y.S.; Zhang, W.K.; Tang, H.B. α-pinene, linalool, and 1-octanol contribute to the topical anti-inflammatory and analgesic activities of frankincense by inhibiting COX-2. J. Ethnopharmacol. 2016, 179, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Basholli-Salihua, M.; Schustera, R.; Hajdaric, A.; Mullaa, D.; Viernsteina, H.; Mustafac, B.; Mueller, M. Phytochemical composition, anti-inflammatory activity and cytotoxic effects of essential oils from three Pinus spp. Pharm. Biol. 2017, 55, 1553–1560. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; Orhan, I.E.; Jugran, A.K.; Jayaweera, S.L.D.; Dias, D.A.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N.; et al. Therapeutic potential of α- and β-pinene: A miracle gift of nature. Biomolecule 2019, 9, 738. [Google Scholar] [CrossRef]
- Palariya, D.; Dhami, A.; Singh, A.; Kumar, R.; Chandra, D.; Pant, A.K.; Prakash, O.; Rawat, D.S. β-pinene rich seed essential oil of Zingiber chrysanthum Roscoe (Zingiberaceae) from Kumaun Lesser Himalaya, Uttarakhand and its biological activities. J. Essent. Oil Bear. Plants. 2019, 22, 1349–1361. [Google Scholar] [CrossRef]
- Da Silva, A.C.; Lopes, P.M.; de Azevedo, M.M.; Costa, D.C.; Alviano, C.S.; Alviano, D.S. Biological activities of alpha-pinene and beta-pinene enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef] [PubMed]
- Surendran, S.; Qassadi, F.; Surendran, G.; Lilley, D.; Heinrich, M. Myrcene-What are the potential health benefits of this flavouring and aroma agent? Front. Nutr. 2021, 8, 699666. [Google Scholar] [CrossRef] [PubMed]
- Weston-Green, K.; Clunas, H.; Jimenez Naranjo, C. A review of the potential use of pinene and linalool as terpene-based medicines for brain health: Discovering novel therapeutics in the flavours and fragrances of Cannabis. Front. Psychiatry 2021, 12, 583211. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Choi, W.S.; Kim, K.J.; Eom, C.D.; Park, M.J. Investigation of active anti-inflammatory constituents of essential oil from Pinus koraiensis (Sieb. et Zucc.) Wood in LPS-stimulated RBL-2H3 cells. Biomolecules 2021, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Yu, X.; Ding, Z.; Cheng, B.; Yi, Y.; Yu, W.; Hayashi, N.; Komae, H. Essential oils of some Lauraceae species from the southwestern parts of China. J. Essent. Oil Res. 1994, 6, 577–585. [Google Scholar] [CrossRef]
- Singh, C.; Singh, S.; Pande, C.; Tewari, G.; Pande, V.; Sharma, P. Exploration of antimicrobial potential of essential oils of Cinnamomum glanduliferum, Feronia elephantum, Bupleurum hamiltonii and Cyclospermum leptophyllum against foodborne pathogens. Pharm. Biol. 2013, 51, 1607–1610. [Google Scholar] [CrossRef] [PubMed]
- Werka, J.S.; Boehme, A.K.; Setzer, W.N. Biological activities of essential oils from Monteverde, Costa Rica. Nat. Prod. Commun. 2007, 2, 1215–1219. [Google Scholar] [CrossRef]
- El-Readi, M.Z.; Eid, H.H.; Ashour, M.L.; Eid, S.Y.; Labib, R.M.; Sporer, F.; Wink, M. Variations of the chemical composition and bioactivity of essential oils from leaves and stems of Liquidambar styraciflua (Altingiaceae). J. Pharm. Pharmacol. 2013, 65, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Yang, K.W.; Kim, S.S.; Park, S.M.; Park, K.J.; Kim, K.S.; Choi, Y.H.; Cho, K.K.; Hyun, C.G. Chemical composition and anti-inflammation activity of essential oils from Citrus unshiu flower. Nat. Prod. Commun. 2014, 9, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Popović, V.; Petrović, S.; Tomić, M.; Stepanović-Petrović, R.; Micov, A.; Pavlović-Drobac, M.; Couladis, M.; Niketić, M. Antinociceptive and anti-edematous activities of the essential oils of two Balkan endemic Laserpitium species. Nat. Prod. Commun. 2014, 9, 125–128. [Google Scholar]
- Mühlbauer, R.C.; Lozano, A.; Palacio, S.; Reinli, A.; Felix, R. Common herbs, essential oils, and monoterpenes potently modulate bone metabolism. Bone 2003, 32, 372–380. [Google Scholar] [CrossRef]
- Nam, S.Y.; Chung, C.K.; Seo, J.H.; Rah, S.Y.; Kim, H.M.; Jeong, H.J. The therapeutic efficacy of α-pinene in an experimental mouse model of allergic rhinitis. Int. Immunopharmacol. 2014, 23, 273–282. [Google Scholar] [CrossRef]
- Van Zyl, R.L.; Seatlholo, S.T.; van Vuuren, S.F. The biological activities of 20 nature identical essential oil constituents. J. Essent. Oil Res. 2006, 18, 129–133. [Google Scholar] [CrossRef]
- Santos, E.S.; Abrantes Coelho, G.L.; Saraiva Fontes Loula, Y.K.; Saraiva Landim, B.L.; Fernandes Lima, C.N.; Tavares de Sousa Machado, S.; Pereira Lopes, M.J.; Soares Gomes, A.D.; Martins da Costa, J.G.; Alencar de Menezes, I.R.; et al. Hypoglycemic, hypolipidemic, and anti-Inflammatory effects of beta-pinene in diabetic rats. Evid. Based Complement. Altern. Med. 2022, 2022, 8173307. [Google Scholar] [CrossRef] [PubMed]
- Kamatou, G.P.; Van Zyl, R.L.; Van Vuuren, S.F.; Viljoen, A.M.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Tilney, P.M. Chemical composition, leaf trichome types and biological activities of the essential oils of four related Salvia species indigenous to Southern Africa. J. Essent. Oil Res. 2006, 18, 72–79. [Google Scholar] [CrossRef]
- Syamsir, D.R.B. Essential oils and biological activities of three selected wild Alpinia species. Master’s Thesis, Institute of Biological Sciences, Faculty of Sciences, University of Malaya, Kuala Lumpur, Malaysia, 2009. [Google Scholar]
- Tung, Y.T.; Yen, P.L.; Lin, C.Y.; Chang, S.T. Anti-inflammatory activities of essential oils and their constituents from different provenances of indigenous cinnamon (Cinnamomum osmophloeum) leaves. Pharm. Biol. 2010, 48, 1130–1136. [Google Scholar] [CrossRef]
- Gasparrini, M.; Afrin, S.; Forbes-Hernández, T.Y.; Cianciosi, D.; Reboredo-Rodriguez, P.; Amici, A.; Battino, M.; Giampieri, F. Protective effects of Manuka honey on LPS-treated RAW 264.7 macrophages. Part 2: Control of oxidative stress induced damage, increase of antioxidant enzyme activities and attenuation of inflammation. Food Chem. Toxicol. 2018, 120, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Bian, K.; Murad, F. Discovery of the nitric oxide signaling pathway and targets for drug development. Front. Biosci. 2009, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Blaise, G.A.; Gauvin, D.; Gangal, M.; Authier, S. Nitric oxide, cell signaling and cell death. Toxicology 2005, 208, 177–192. [Google Scholar] [CrossRef]
- Kim, H.; Darwish, I.; Monroy, M.-F.; Prockop, D.J.; Liles, W.C.; Kain, K.C. Mesenchymal stromal (stem) cells suppress pro-inflammatory cytokine production but fail to improve survival in experimental staphylococcal toxic shock syndrome. BMC Immunol. 2014, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Chung, T.W.; Jeong, Y.J.; Kwak, C.H.; Ha, S.H.; Kwon, K.M.; Abekura, F.; Cho, S.H.; Lee, Y.C.; Ha, K.T.; et al. Ascofuranone inhibits lipopolysaccharide-induced inflammatory response via NF-kappaB and AP-1, p-ERK, TNF-α, IL-6 and IL-1β in RAW 264.7 macrophages. PLoS ONE 2017, 12, e0171322. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.J.; Moon, J.Y.; Song, G.; Lee, Y.K.; Han, M.S.; Lee, J.S.; Ihm, B.S.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Artemisia fukudo essential oil attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Yamazaki, M.; Katagata, Y. Kuromoji (Lindera umbellata) essential oil inhibits LPS-induced inflammation in RAW 264.7 cells. Biosci. Biotechnol. Biochem. 2013, 77, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, N.; Echchannaoui, H.; Landmann, R.; Rivest, S. Cooperation between toll-like receptor 2 and 4 in the brain of mice challenged with cell wall components derived from gram-negative and gram-positive bacteria. Eur. J. Immunol. 2003, 33, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, X.; Bi, Y.; Miao, R.; Zhang, Z.; Su, H. Anti-inflammatory effects of Cumin essential oil by blocking JNK, ERK, and NF-𝜅B signaling pathways in LPS-stimulated RAW 264.7 cells. Evid. Based Complement. Altern. Med. 2015, 2015, 474509. [Google Scholar]
- Liou, H.C. Regulation of the immune system by NF-κB and IκB. BMB Rep. 2002, 35, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.D.O.; Yaochite, J.N.U.; Braga, M.A.; Sousa, A.R.D.; Sasahara, G.L.; Fonseca, S.G.D.C.; Araújo, T.D.D.V.; Santiago, G.M.P.; Sousa, L.M.D.; Carvalho, J.L.D.; et al. Antioxidant and anti-inflammatory activities of Bauhinia ungulata L. (Fabaceae) on LPS-stimulated RAW 264.7 Cells. Pharmacogn. J. 2019, 11, 37–42. [Google Scholar] [CrossRef]
- Tian, Y.; Zhou, S.; Takeda, R.; Okazaki, K.; Sekita, M.; Sakamoto, K. Anti-inflammatory activities of amber extract in lipo-polysaccharide-induced RAW 264.7 macrophages. Biomed. Pharmacother. 2021, 141, 111854. [Google Scholar] [CrossRef]
- Nogueira, V.; Hay, N. Molecular pathways: Reactive oxygen species homeostasis in cancer cells and implications for cancer therapy. Clin. Cancer Res. 2013, 19, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, X.; Bao, Y.; Wang, X.; Zhai, J.; Zhan, X.; Zhang, H. Characterization and anti-inflammation of a polysaccharide produced by Chaetomium globosum CGMCC 6882 on LPS-induced RAW 264.7 cells. Carbohydr. Polym. 2021, 251, 117129. [Google Scholar] [CrossRef] [PubMed]
- Bognar, E.; Sarszegi, Z.; Szabo, A.; Debreceni, B.; Kalman, N.; Tucsek, Z.; Sumegi, B.; Gallyas, F., Jr. Antioxidant and anti-inflammatory effects in RAW264. 7 macrophages of malvidin, a major red wine polyphenol. PLoS ONE 2013, 8, e65355. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Chen, L.; Ma, X.; Jiao, R.; Li, X.; Wang, Y. Protective effects of kaempferol against reactive oxygen species-induced hemolysis and its antiproliferative activity on human cancer cells. Eur. J. Med. Chem. 2016, 114, 24–32. [Google Scholar] [CrossRef]
- Wan, F.; Lenardo, M.J. The nuclear signaling of NF-κB: Current knowledge, new insights, and future perspectives. Cell Res. 2010, 20, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Nam, N.H. Naturally occurring NF-κB inhibitors. Mini-Rev. Med. Chem. 2006, 6, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-kappaB signaling pathways in neurological inflammation: A mini review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Held, S.; Schieberle, P.; Somoza, V. Characterization of α-terpineol as an anti-inflammatory component of orange juice by in vitro studies using oral buccal cells. J. Agric. Food Chem. 2007, 55, 8040–8046. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.J.; Kim, S.S.; Oh, T.H.; Lee, N.H.; Hyun, C.G. Cryptomeria japonica essential oil inhibits the growth of drug-resistant skin pathogens and LPS-induced nitric oxide and pro-inflammatory cytokine production. Pol. J. Microbiol. 2009, 58, 61–68. [Google Scholar] [PubMed]
- Council of Europe; European Pharmacopoeia Commission; European Directorate for the Quality of Medicines and Healthcare. European Pharmacopoeia, 7th ed.; European Directorate for the Quality of Medicines and Healthcare, Council of Europe: Strasbourg, France, 2010. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Ray, A.; Jena, S.; Dash, B.; Sahoo, A.; Kar, B.; Patnaik, J.; Panda, P.C.; Nayak, S.; Mahapatra, N. Hedychium coronarium extract arrests cell cycle progression, induces apoptosis, and impairs migration and invasion in HeLa cervical cancer cells. Cancer Manag. Res. 2019, 11, 483–500. [Google Scholar] [CrossRef] [PubMed]
- Jena, S.; Ray, A.; Sahoo, A.; Das, P.K.; Dash, K.T.; Kar, S.K.; Nayak, S.; Panda, P.C. Chemical composition and biological activities of leaf essential oil of Syzygium myrtifolium from eastern India. J. Essent. Oil Bear. Plants. 2021, 24, 582–595. [Google Scholar] [CrossRef]
- Syam, S.; Bustamam, A.; Abdullah, R.; Sukari, M.A.; Hashim, N.M.; Mohan, S.; Looi, C.Y.; Wong, W.F.; Yahayu, M.A.; Abdelwahab, S.I. β Mangostin suppress LPS-induced inflammatory response in RAW 264.7 macrophages in vitro and carrageenan-induced peritonitis in vivo. J. Ethnopharmacol. 2014, 153, 435–445. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | RI a | RI b | Peak Area (%) | |
| Leaf | Bark | ||||
| 1 | α-Thujene | 921 | 930 | 0.89 | 0.55 |
| 2 | α-Pinene | 930 | 939 | 9.27 | 10.99 |
| 3 | α-Fenchene | 945 | 952 | 0.29 | 0.58 |
| 4 | Benzaldehyde | 957 | 960 | 0.14 | - |
| 5 | Sabinene | 966 | 975 | - | 2.30 |
| 6 | β-Pinene | 971 | 979 | 45.21 | 13.11 |
| 7 | Myrcene | 984 | 990 | 9.97 | 7.81 |
| 8 | α-Phellandrene | 1002 | 1002 | - | 0.26 |
| 9 | δ-3-Carene | 1003 | 1011 | 0.06 | 0.77 |
| 10 | α-Terpinene | 1012 | 1017 | 1.61 | 2.09 |
| 11 | p-Cymene | 1019 | 1024 | 0.29 | 0.15 |
| 12 | Limonene | 1024 | 1029 | 2.18 | 1.12 |
| 13 | 1,8-Cineole | 1028 | 1031 | 2.66 | - |
| 14 | (Z)-β-Ocimene | 1039 | 1037 | 0.24 | - |
| 15 | (E)-β-Ocimene | 1052 | 1050 | 2.30 | 3.00 |
| 16 | γ-Terpinene | 1066 | 1059 | 1.38 | - |
| 17 | Terpinolene | 1098 | 1088 | 1.55 | - |
| 18 | 6-Camphenol | 1120 | 1113 | 0.40 | 0.28 |
| 19 | α-Terpineol | 1176 | 1188 | 3.95 | 6.79 |
| 20 | γ-Terpineol | 1190 | 1199 | 0.30 | 1.55 |
| 21 | Linalool formate | 1204 | 1216 | 0.08 | - |
| 22 | endo-Fenchyl acetate | 1220 | 1220 | 0.20 | - |
| 23 | Linalool acetate | 1277 | 1257 | - | 0.39 |
| 24 | δ-Terpinyl acetate | 1327 | 1317 | 1.36 | 0.19 |
| 25 | δ-Elemene | 1352 | 1338 | 0.14 | - |
| 26 | β-Patchoulene | 1380 | 1381 | 0.97 | 1.10 |
| 27 | β-Cubebene | 1397 | 1388 | - | 0.26 |
| 28 | β-Elemene | 1404 | 1390 | 0.11 | - |
| 29 | (Z)-Caryophyllene | 1410 | 1408 | 2.96 | 9.30 |
| 30 | (E)-Caryophyllene | 1431 | 1419 | 0.52 | 0.12 |
| 31 | γ-Elemene | 1444 | 1436 | 0.23 | 0.66 |
| 32 | α-Humulene | 1446 | 1454 | 0.28 | - |
| 33 | Aromadendrene | 1459 | 1441 | 0.11 | 0.22 |
| 34 | allo-Aromadendrene | 1464 | 1460 | 0.07 | 0.76 |
| 35 | 9-epi-(E)-Caryophyllene | 1470 | 1466 | 0.38 | - |
| 36 | γ-Gurjunene | 1477 | 1477 | 0.12 | 0.47 |
| 37 | γ-Muurolene | 1484 | 1479 | 0.59 | 1.24 |
| 38 | ar-Curcumene | 1488 | 1480 | 1.10 | 1.31 |
| 39 | Germacrene D | 1501 | 1481 | 0.12 | 1.51 |
| 40 | γ-Curcumene | 1505 | 1482 | 0.41 | 5.50 |
| 41 | trans-Calamenene | 1512 | 1522 | 0.19 | 0.18 |
| 42 | δ-Cadinene | 1514 | 1523 | 0.43 | - |
| 43 | cis-Calamenene | 1521 | 1529 | 0.57 | 0.29 |
| 44 | γ- Cuprenene | 1525 | 1533 | 0.08 | 0.43 |
| 45 | Germacrene B | 1571 | 1561 | 1.30 | 1.07 |
| 46 | Caryophyllene oxide | 1582 | 1583 | - | 0.16 |
| 47 | Humulene epoxide II | 1603 | 1608 | - | 0.24 |
| 48 | Guaiol | 1630 | 1600 | 0.08 | 3.15 |
| 49 | 10-epi-γ-Eudesmol | 1632 | 1623 | 0.07 | 3.49 |
| 50 | 1-epi-Cubenol | 1637 | 1628 | - | 2.29 |
| 51 | α-Cadinol | 1644 | 1654 | 0.17 | 11.18 |
| 52 | α-Muurolol | 1660 | 1646 | - | 0.28 |
| 53 | α-Bisabolol | 1684 | 1685 | 0.22 | 1.79 |
| Monoterpene hydrocarbons (1–3, 5–12, 14–17) | 75.24 | 42.73 | |||
| Monoterpene aldehyde (4) | 0.14 | - | |||
| Monoterpene ether (13) | 2.66 | - | |||
| Monoterpene alcohol (18–20) | 4.65 | 8.62 | |||
| Monoterpene ester (21–24) | 1.64 | 0.58 | |||
| Sesquiterpene hydrocarbon (25–45) | 10.68 | 24.42 | |||
| Sesquiterpene ether (46, 47) | - | 0.4 | |||
| Sesquiterpene alcohol (48–53) | 0.54 | 22.18 | |||
| Total identified | 95.55 | 98.93 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jena, S.; Ray, A.; Mohanta, O.; Das, P.K.; Sahoo, A.; Nayak, S.; Panda, P.C. Neocinnamomum caudatum Essential Oil Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Cells by Inhibiting NF-κB Activation and ROS Production. Molecules 2022, 27, 8193. https://doi.org/10.3390/molecules27238193
Jena S, Ray A, Mohanta O, Das PK, Sahoo A, Nayak S, Panda PC. Neocinnamomum caudatum Essential Oil Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Cells by Inhibiting NF-κB Activation and ROS Production. Molecules. 2022; 27(23):8193. https://doi.org/10.3390/molecules27238193
Chicago/Turabian StyleJena, Sudipta, Asit Ray, Omprakash Mohanta, Prabhat Kumar Das, Ambika Sahoo, Sanghamitra Nayak, and Pratap Chandra Panda. 2022. "Neocinnamomum caudatum Essential Oil Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Cells by Inhibiting NF-κB Activation and ROS Production" Molecules 27, no. 23: 8193. https://doi.org/10.3390/molecules27238193
APA StyleJena, S., Ray, A., Mohanta, O., Das, P. K., Sahoo, A., Nayak, S., & Panda, P. C. (2022). Neocinnamomum caudatum Essential Oil Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Cells by Inhibiting NF-κB Activation and ROS Production. Molecules, 27(23), 8193. https://doi.org/10.3390/molecules27238193