
Thermodynamics and Kinetics of Electron Transfer of Electrode-Immobilized Small Laccase from Streptomyces coelicolor

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
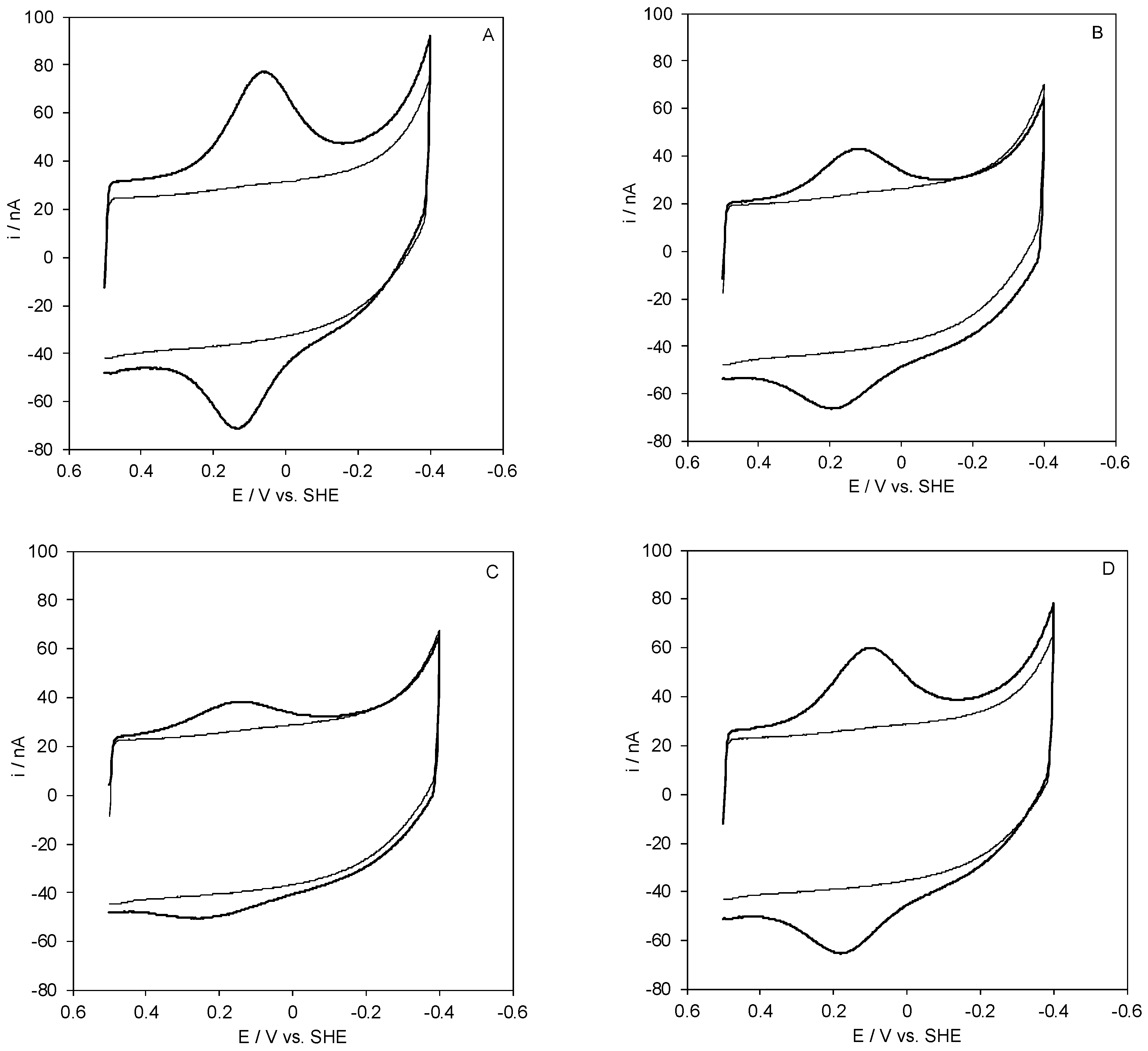
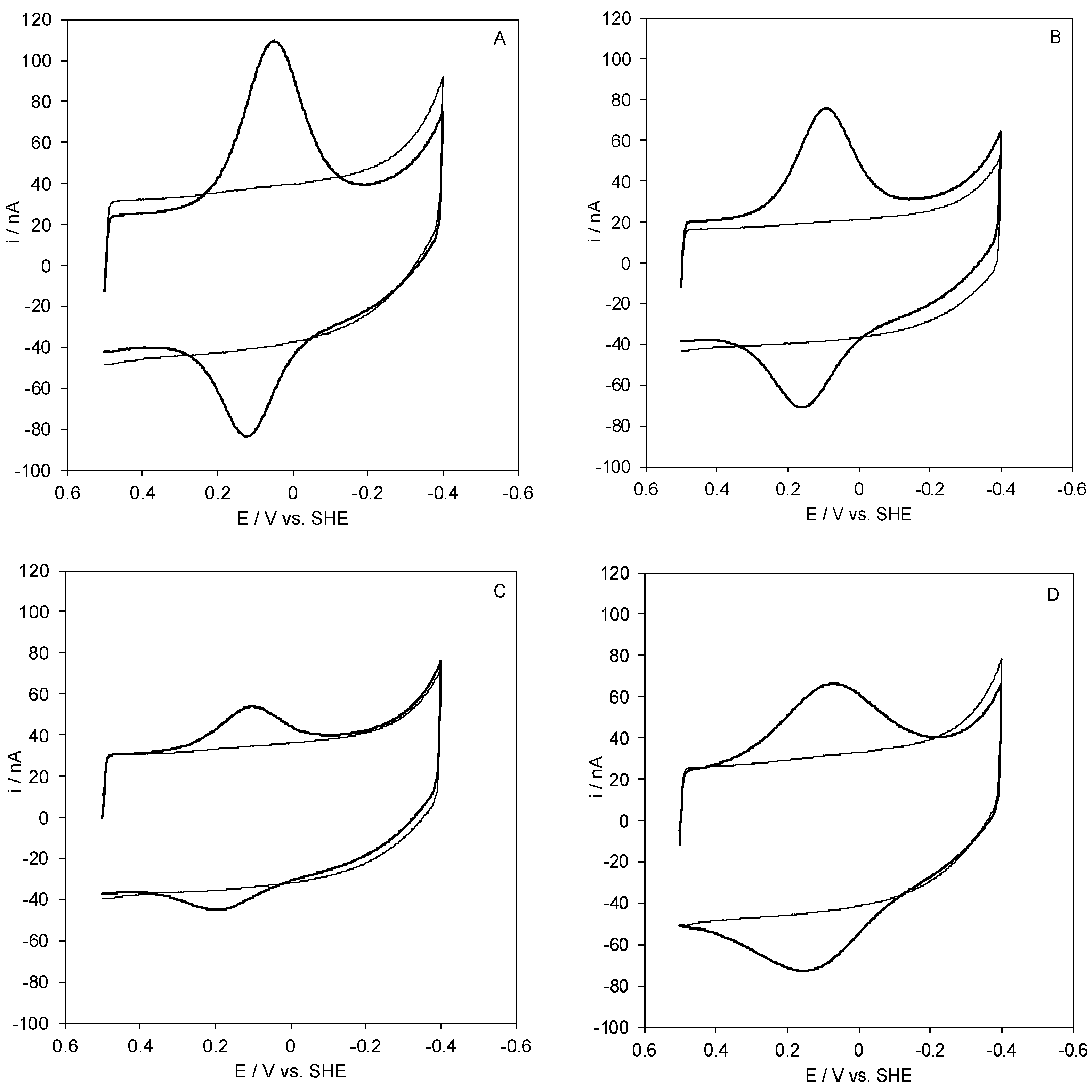
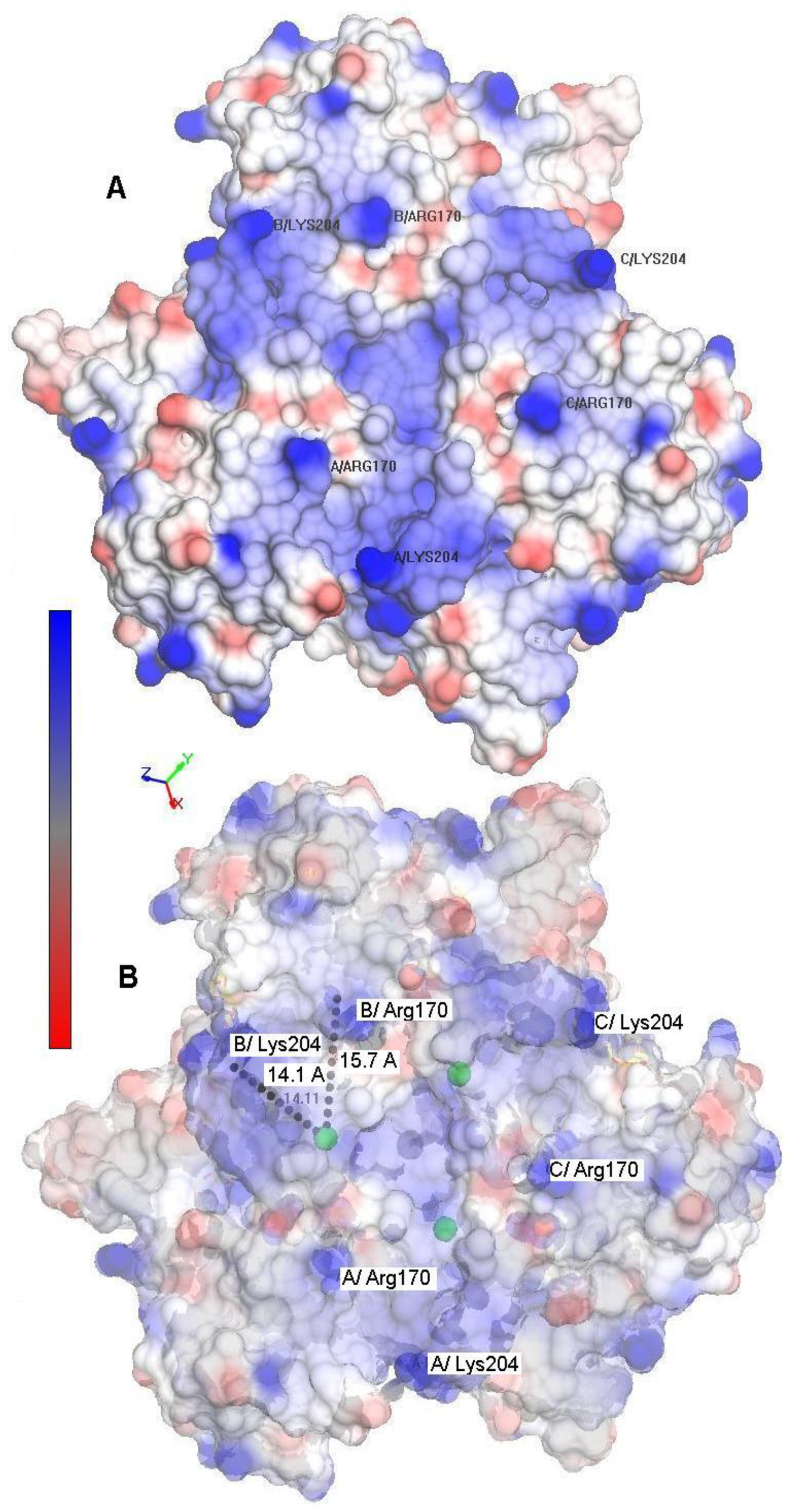
2.1. Voltammetric Responses of Electrode-Immobilized SLAC

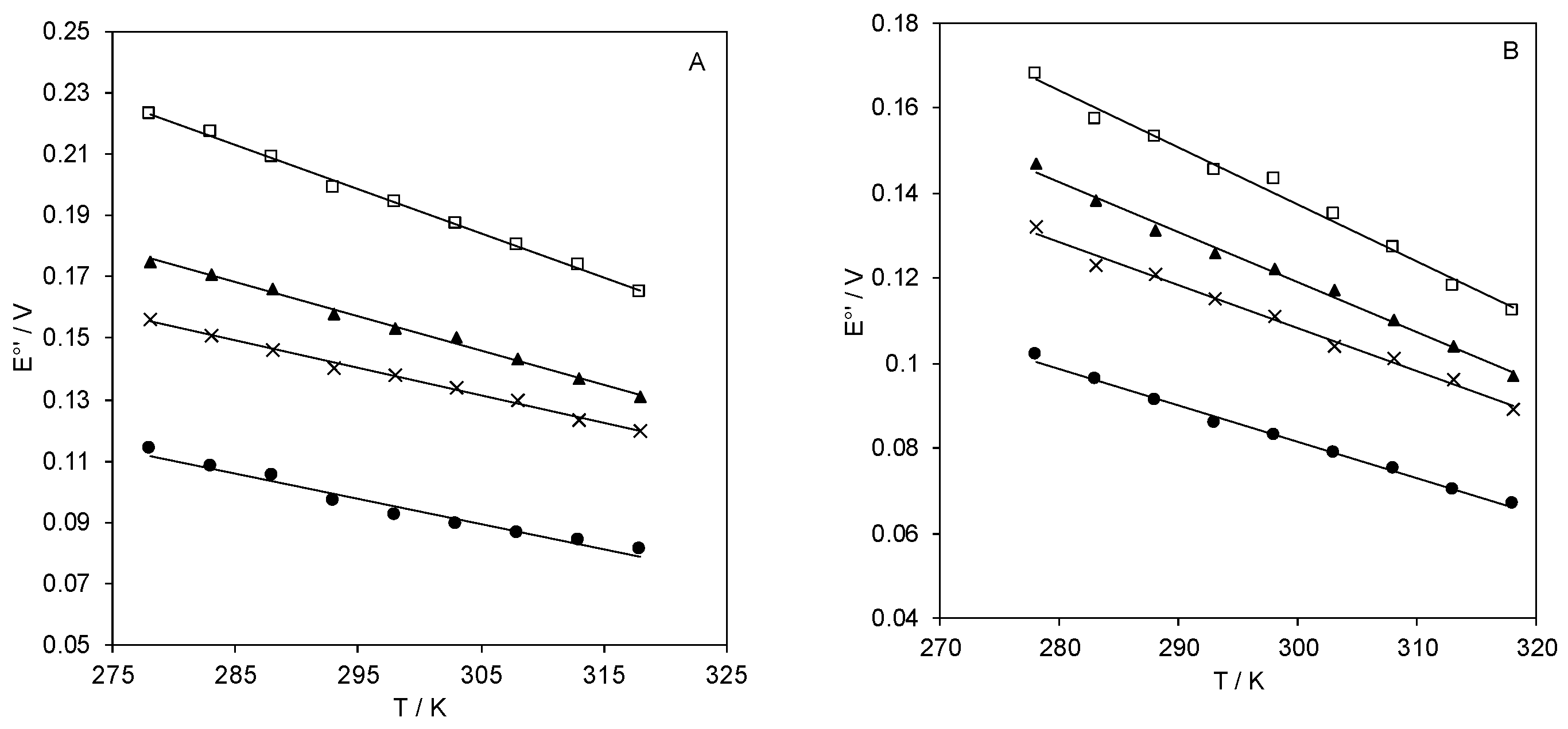
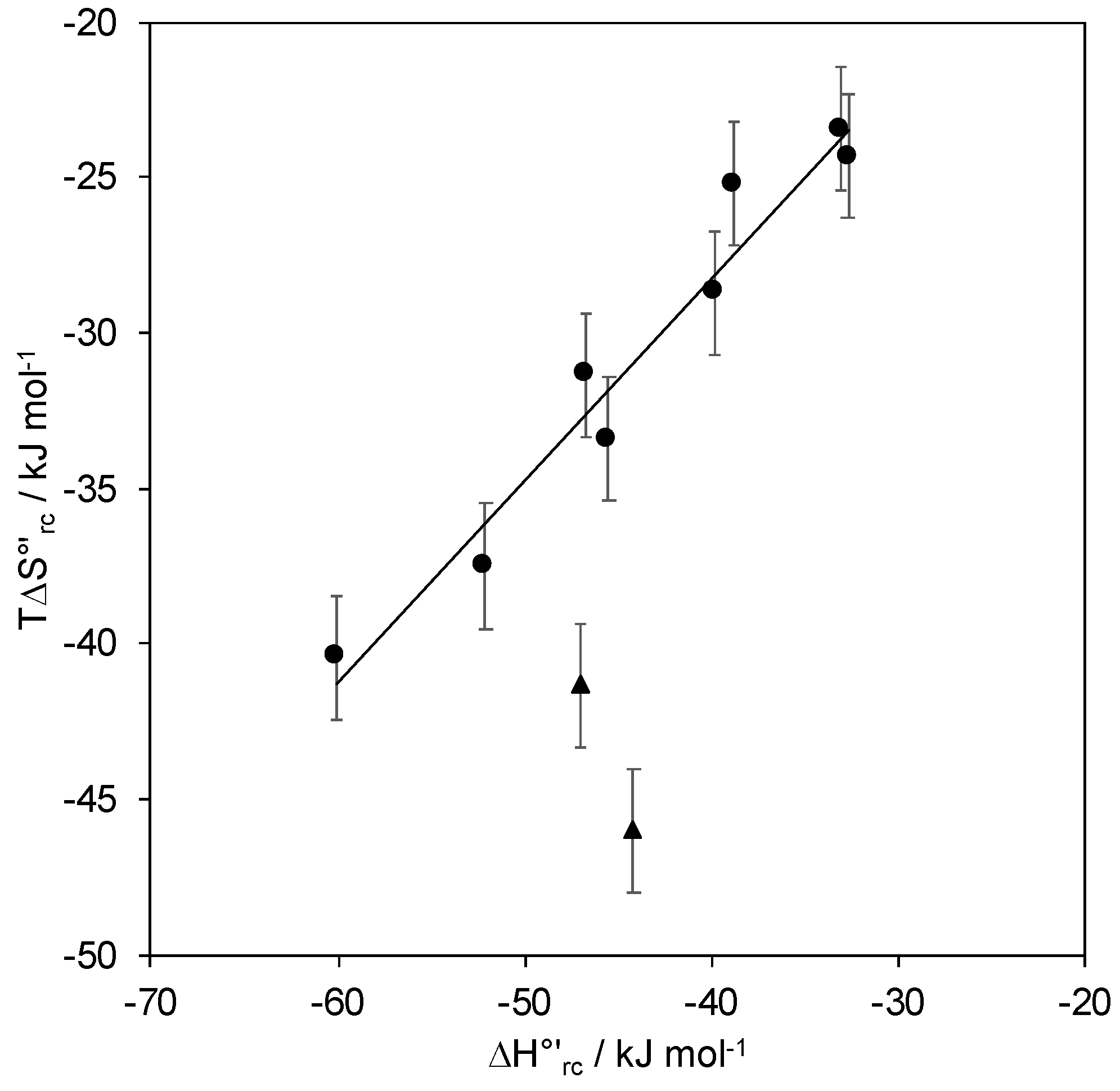
2.2. Reduction Thermodynamics and pH-Induced Changes
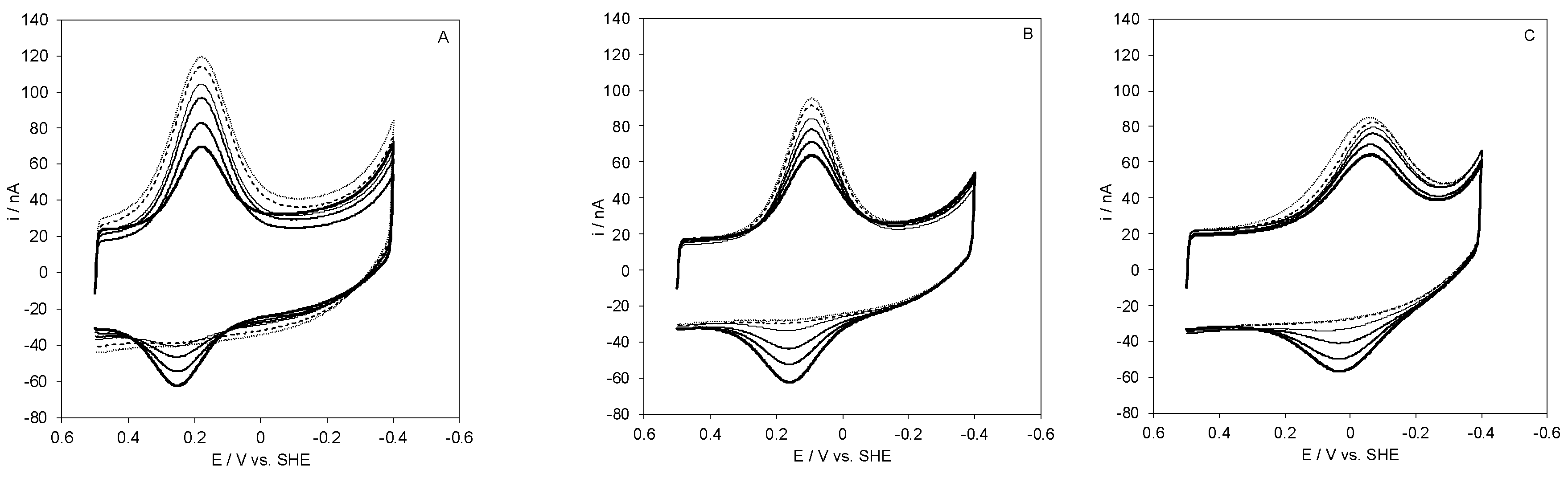
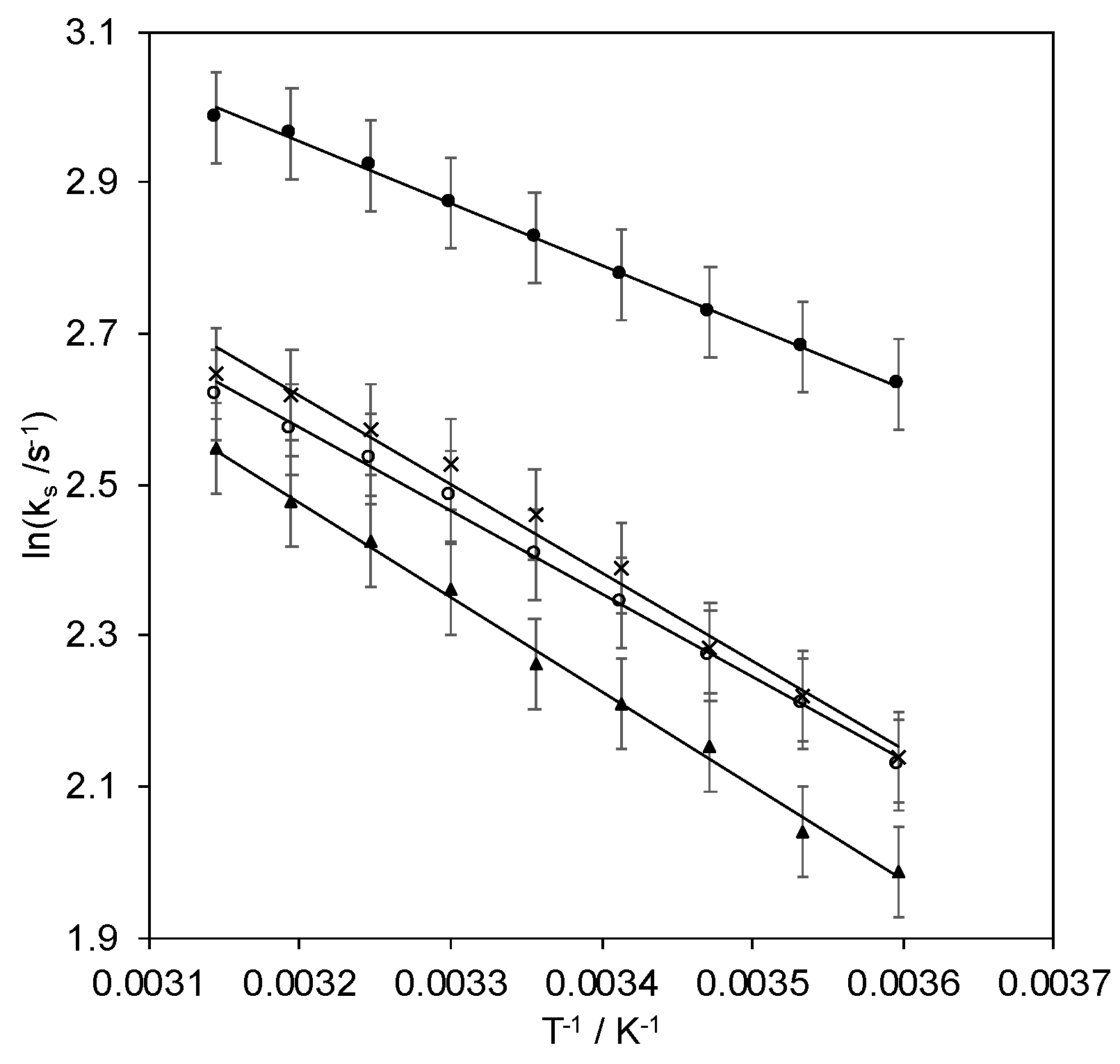
2.3. Kinetics of Heterogeneous Protein-Electrode ET
3. Experimental Section
3.1. Materials
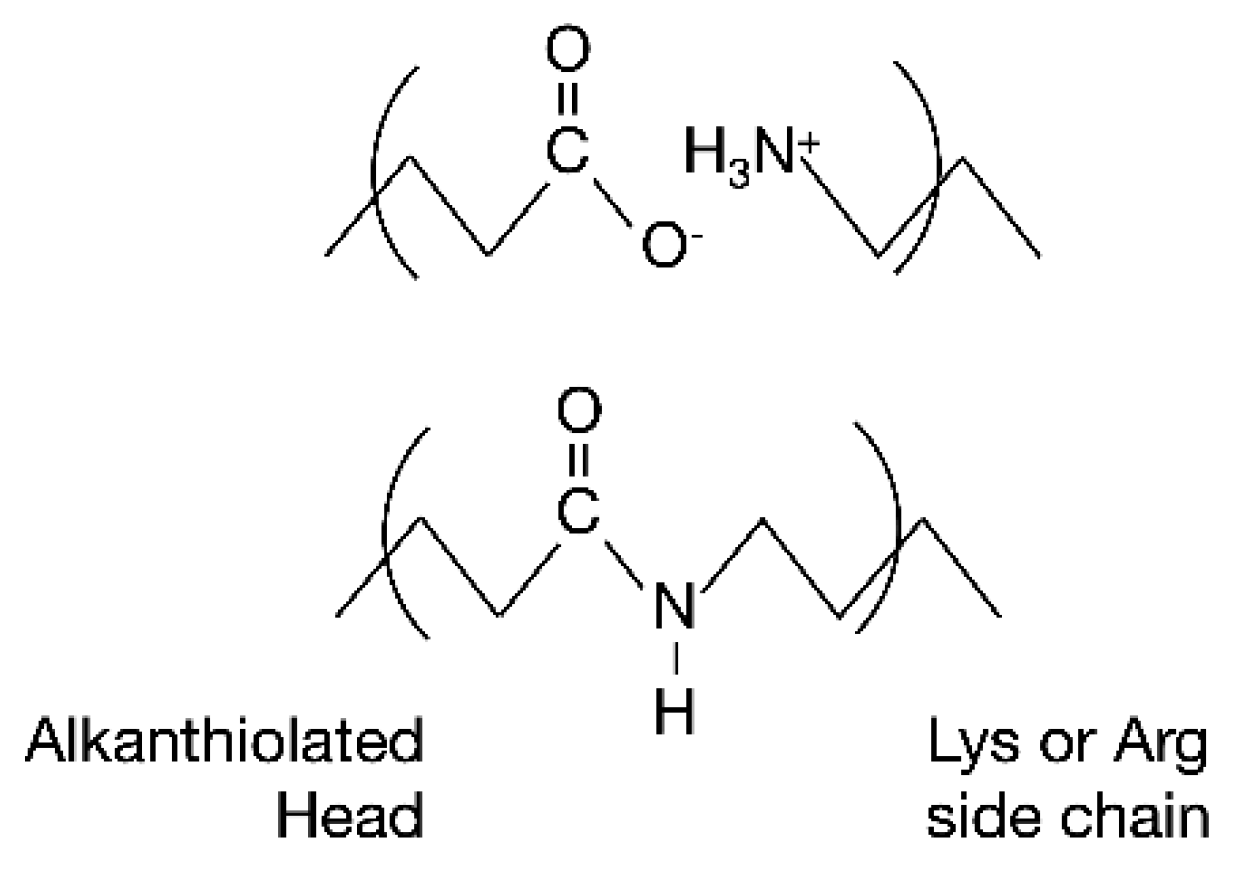
3.2. Electrochemical Measurements
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Yoshida, H. LXIII.—Chemistry of Lacquer (Urushi). Part I. Communication from the Chemical Society of Tokio. J. Chem. Soc. Trans. 1883, 43, 472–486. [Google Scholar] [CrossRef]
- Lontie, R. Copper Proteins and Copper Enzymes; Lontie, R., Ed.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781351070904. [Google Scholar]
- Yaropolov, A.I.; Skorobogat’ko, O.V.; Vartanov, S.S.; Varfolomeyev, S.D. Laccase. Appl. Biochem. Biotechnol. 1994, 49, 257–280. [Google Scholar] [CrossRef]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper Oxidases and Oxygenases. Chem. Rev. 1996, 96, 2563–2606. [Google Scholar] [CrossRef]
- Lee, S.-K.; George, S.D.; Antholine, W.E.; Hedman, B.; Hodgson, K.O.; Solomon, E.I. Nature of the Intermediate Formed in the Reduction of O2 to H2O at the Trinuclear Copper Cluster Active Site in Native Laccase. J. Am. Chem. Soc. 2002, 124, 6180–6193. [Google Scholar] [CrossRef] [PubMed]
- Rulíšek, L.; Solomon, E.I.; Ryde, U. A Combined Quantum and Molecular Mechanical Study of the O2 Reductive Cleavage in the Catalytic Cycle of Multicopper Oxidases. Inorg. Chem. 2005, 44, 5612–5628. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Go, N. Function and Molecular Evolution of Multicopper Blue Proteins. Cell. Mol. Life Sci. 2005, 62, 2050–2066. [Google Scholar] [CrossRef]
- Sanchez-Amat, A.; Solano, F. A Pluripotent Polyphenol Oxidase from the Melanogenic MarineAlteromonas SpShares Catalytic Capabilities of Tyrosinases and Laccases. Biochem. Biophys. Res. Commun. 1997, 240, 787–792. [Google Scholar] [CrossRef]
- Diamantidis, G.; Effosse, A.; Potier, P.; Bally, R. Purification and Characterization of the First Bacterial Laccase in the Rhizospheric Bacterium Azospirillum Lipoferum. Soil Biol. Biochem. 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Alexandre, G.; Zhulin, I.B. Laccases Are Widespread in Bacteria. Trends Biotechnol. 2000, 18, 41–42. [Google Scholar] [CrossRef]
- Grass, G.; Rensing, C. CueO Is a Multi-Copper Oxidase That Confers Copper Tolerance in Escherichia coli. Biochem. Biophys. Res. Commun. 2001, 286, 902–908. [Google Scholar] [CrossRef]
- Martins, L.O.; Soares, C.M.; Pereira, M.M.; Teixeira, M.; Costa, T.; Jones, G.H.; Henriques, A.O. Molecular and Biochemical Characterization of a Highly Stable Bacterial Laccase That Occurs as a Structural Component of the Bacillus Subtilis Endospore Coat. J. Biol. Chem. 2002, 277, 18849–18859. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Wildner, G.F.; Grass, G.; Weichsel, A.; Ambrus, A.; Rensing, C.; Montfort, W.R. A Labile Regulatory Copper Ion Lies Near the T1 Copper Site in the Multicopper Oxidase CueO. J. Biol. Chem. 2003, 278, 31958–31963. [Google Scholar] [CrossRef] [PubMed]
- Enguita, F.J.; Martins, L.O.; Henriques, A.O.; Carrondo, M.A. Crystal Structure of a Bacterial Endospore Coat Component. J. Biol. Chem. 2003, 278, 19416–19425. [Google Scholar] [CrossRef] [PubMed]
- Durão, P.; Bento, I.; Fernandes, A.T.; Melo, E.P.; Lindley, P.F.; Martins, L.O. Perturbations of the T1 Copper Site in the CotA Laccase from Bacillus Subtilis: Structural, Biochemical, Enzymatic and Stability Studies. JBIC J. Biol. Inorg. Chem. 2006, 11, 514–526. [Google Scholar] [CrossRef]
- Dalfard, A.B.; Khajeh, K.; Soudi, M.R.; Naderi-Manesh, H.; Ranjbar, B.; Sajedi, R.H. Isolation and Biochemical Characterization of Laccase and Tyrosinase Activities in a Novel Melanogenic Soil Bacterium. Enzyme Microb. Technol. 2006, 39, 1409–1416. [Google Scholar] [CrossRef]
- Li, X.; Wei, Z.; Zhang, M.; Peng, X.; Yu, G.; Teng, M.; Gong, W. Crystal Structures of E. Coli Laccase CueO at Different Copper Concentrations. Biochem. Biophys. Res. Commun. 2007, 354, 21–26. [Google Scholar] [CrossRef]
- Miura, Y.; Tsujimura, S.; Kamitaka, Y.; Kurose, S.; Kataoka, K.; Sakurai, T.; Kano, K. Bioelectrocatalytic Reduction of O 2 Catalyzed by CueO from Escherichia Coli Adsorbed on a Highly Oriented Pyrolytic Graphite Electrode. Chem. Lett. 2007, 36, 132–133. [Google Scholar] [CrossRef]
- Lee, C.-W.; Gray, H.B.; Anson, F.C.; Malmström, B.G. Catalysis of the Reduction of Dioxygen at Graphite Electrodes Coated with Fungal Laccase A. J. Electroanal. Chem. Interfacial Electrochem. 1984, 172, 289–300. [Google Scholar] [CrossRef]
- Ghindilis, A. Direct Electron Transfer Catalysed by Enzymes: Application for Biosensor Development. Biochem. Soc. Trans. 2000, 28, 84–89. [Google Scholar] [CrossRef]
- Calabrese Barton, S.; Gallaway, J.; Atanassov, P. Enzymatic Biofuel Cells for Implantable and Microscale Devices. Chem. Rev. 2004, 104, 4867–4886. [Google Scholar] [CrossRef]
- Heller, A. Miniature Biofuel Cells. Phys. Chem. Chem. Phys. 2004, 6, 209. [Google Scholar] [CrossRef]
- Liu, Y.; Qu, X.; Guo, H.; Chen, H.; Liu, B.; Dong, S. Facile Preparation of Amperometric Laccase Biosensor with Multifunction Based on the Matrix of Carbon Nanotubes–Chitosan Composite. Biosens. Bioelectron. 2006, 21, 2195–2201. [Google Scholar] [CrossRef]
- Tsujimura, S.; Nakagawa, T.; Kano, K.; Ikeda, T. Kinetic Study of Direct Bioelectrocatalysis of Dioxygen Reduction with Bilirubin Oxidase at Carbon Electrodes. Electrochemistry 2004, 72, 437–439. [Google Scholar] [CrossRef]
- Johnson, D.L.; Thompson, J.L.; Brinkmann, S.M.; Schuller, K.A.; Martin, L.L. Electrochemical Characterization of Purified Rhus Vernicifera Laccase: Voltammetric Evidence for a Sequential Four-Electron Transfer. Biochemistry 2003, 42, 10229–10237. [Google Scholar] [CrossRef] [PubMed]
- Christenson, A.; Dimcheva, N.; Ferapontova, E.E.; Gorton, L.; Ruzgas, T.; Stoica, L.; Shleev, S.; Yaropolov, A.I.; Haltrich, D.; Thorneley, R.N.F.; et al. Direct Electron Transfer Between Ligninolytic Redox Enzymes and Electrodes. Electroanalysis 2004, 16, 1074–1092. [Google Scholar] [CrossRef]
- Shleev, S.; el Kasmi, A.; Ruzgas, T.; Gorton, L. Direct Heterogeneous Electron Transfer Reactions of Bilirubin Oxidase at a Spectrographic Graphite Electrode. Electrochem. Commun. 2004, 6, 934–939. [Google Scholar] [CrossRef]
- Nazaruk, E.; Michota, A.; Bukowska, J.; Shleev, S.; Gorton, L.; Bilewicz, R. Properties of Native and Hydrophobic Laccases Immobilized in the Liquid-Crystalline Cubic Phase on Electrodes. JBIC J. Biol. Inorg. Chem. 2007, 12, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, B.; Rahmati-Panah, A.; Shleev, S.; Gorton, L. Carbon Ceramic Electrodes Modified with Laccase FromTrametes Hirsuta: Fabrication, Characterization and Their Use for Phenolic Compounds Detection. Electroanalysis 2007, 19, 907–917. [Google Scholar] [CrossRef]
- Weigel, M.C.; Tritscher, E.; Lisdat, F. Direct Electrochemical Conversion of Bilirubin Oxidase at Carbon Nanotube-Modified Glassy Carbon Electrodes. Electrochem. Commun. 2007, 9, 689–693. [Google Scholar] [CrossRef]
- Murata, K.; Nakamura, N.; Ohno, H. Direct Electron Transfer Reaction of Ascorbate Oxidase Immobilized by a Self-Assembled Monolayer and Polymer Membrane Combined System. Electroanalysis 2007, 19, 530–534. [Google Scholar] [CrossRef]
- Ivnitski, D.; Atanassov, P. Electrochemical Studies of Intramolecular Electron Transfer in Laccase FromTrametes Versicolor. Electroanalysis 2007, 19, 2307–2313. [Google Scholar] [CrossRef]
- Shleev, S.; Jarosz-Wilkolazka, A.; Khalunina, A.; Morozova, O.; Yaropolov, A.; Ruzgas, T.; Gorton, L. Direct Electron Transfer Reactions of Laccases from Different Origins on Carbon Electrodes. Bioelectrochemistry 2005, 67, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Shleev, S.; Christenson, A.; Serezhenkov, V.; Burbaev, D.; Yaropolov, A.; Gorton, L.; Ruzgas, T. Electrochemical Redox Transformations of T1 and T2 Copper Sites in Native Trametes Hirsuta Laccase at Gold Electrode. Biochem. J. 2005, 385, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Shleev, S.; Pita, M.; Yaropolov, A.I.; Ruzgas, T.; Gorton, L. Direct Heterogeneous Electron Transfer Reactions OfTrametes Hirsuta Laccase at Bare and Thiol-Modified Gold Electrodes. Electroanalysis 2006, 18, 1901–1908. [Google Scholar] [CrossRef]
- Shleev, S.; Tkac, J.; Christenson, A.; Ruzgas, T.; Yaropolov, A.I.; Whittaker, J.W.; Gorton, L. Direct Electron Transfer between Copper-Containing Proteins and Electrodes. Biosens. Bioelectron. 2005, 20, 2517–2554. [Google Scholar] [CrossRef]
- Thuesen, M.H.; Farver, O.; Reinhammar, B.; Ulstrup, J.; Guirgis, G.A.; Rosendahl, C.N.; Søtofte, I.; Långström, B. Cyclic Voltammetry and Electrocatalysis of the Blue Copper Oxidase Polyporus Versicolor Laccase. Acta Chem. Scand. 1998, 52, 555–562. [Google Scholar] [CrossRef]
- Tarasevich, M.R.; Bogdanovskaya, V.A.; Kuznetsova, L.N. Bioelectrocatalytic Reduction of Oxygen in the Presence of Laccase Adsorbed on Carbon Electrodes. Russ. J. Electrochem. 2001, 37, 833–837. [Google Scholar] [CrossRef]
- Tsujimura, S.; Kano, K.; Ikeda, T. Glucose/O2 Biofuel Cell Operating at Physiological Conditions. Electrochemistry 2002, 70, 940–942. [Google Scholar] [CrossRef]
- Kamitaka, Y.; Tsujimura, S.; Kataoka, K.; Sakurai, T.; Ikeda, T.; Kano, K. Effects of Axial Ligand Mutation of the Type I Copper Site in Bilirubin Oxidase on Direct Electron Transfer-Type Bioelectrocatalytic Reduction of Dioxygen. J. Electroanal. Chem. 2007, 601, 119–124. [Google Scholar] [CrossRef]
- Machczynski, M.C.; Vijgenboom, E.; Samyn, B.; Canters, G.W. Characterization of SLAC: A Small Laccase from Streptomyces Coelicolor with Unprecedented Activity. Protein Sci. 2004, 13, 2388–2397. [Google Scholar] [CrossRef]
- Skálová, T.; Dohnálek, J.; Østergaard, L.H.; Østergaard, P.R.; Kolenko, P.; Dušková, J.; Štěpánková, A.; Hašek, J. The Structure of the Small Laccase from Streptomyces Coelicolor Reveals a Link between Laccases and Nitrite Reductases. J. Mol. Biol. 2009, 385, 1165–1178. [Google Scholar] [CrossRef] [PubMed]
- Wheeldon, I.R.; Gallaway, J.W.; Barton, S.C.; Banta, S. Bioelectrocatalytic Hydrogels from Electron-Conducting Metallopolypeptides Coassembled with Bifunctional Enzymatic Building Blocks. Proc. Natl. Acad. Sci. USA 2008, 105, 15275–15280. [Google Scholar] [CrossRef] [PubMed]
- Lörcher, S.; Lopes, P.; Kartashov, A.; Ferapontova, E.E. Direct Bio-Electrocatalysis of O2 Reduction by Streptomyces Coelicolor Laccase Orientated at Promoter-Modified Graphite Electrodes. ChemPhysChem 2013, 14, 2112–2124. [Google Scholar] [CrossRef] [PubMed]
- Gallaway, J.; Wheeldon, I.; Rincon, R.; Atanassov, P.; Banta, S.; Barton, S.C. Oxygen-Reducing Enzyme Cathodes Produced from SLAC, a Small Laccase from Streptomyces Coelicolor. Biosens. Bioelectron. 2008, 23, 1229–1235. [Google Scholar] [CrossRef]
- Schweiss, R.; Welzel, P.B.; Werner, C.; Knoll, W. Dissociation of Surface Functional Groups and Preferential Adsorption of Ions on Self-Assembled Monolayers Assessed by Streaming Potential and Streaming Current Measurements. Langmuir 2001, 17, 4304–4311. [Google Scholar] [CrossRef]
- Vezenov, D.V.; Noy, A.; Rozsnyai, L.F.; Lieber, C.M. Force Titrations and Ionization State Sensitive Imaging of Functional Groups in Aqueous Solutions by Chemical Force Microscopy. J. Am. Chem. Soc. 1997, 119, 2006–2015. [Google Scholar] [CrossRef]
- Tepper, A.W.J.W.; Milikisyants, S.; Sottini, S.; Vijgenboom, E.; Groenen, E.J.J.; Canters, G.W. Identification of a Radical Intermediate in the Enzymatic Reduction of Oxygen by a Small Laccase. J. Am. Chem. Soc. 2009, 131, 11680–11682. [Google Scholar] [CrossRef]
- Guan, D.; Kurra, Y.; Liu, W.; Chen, Z. A Click Chemistry Approach to Site-Specific Immobilization of a Small Laccase Enables Efficient Direct Electron Transfer in a Biocathode. Chem. Commun. 2015, 51, 2522–2525. [Google Scholar] [CrossRef]
- Han, Z.; Zhao, L.; Yu, P.; Chen, J.; Wu, F.; Mao, L. Comparative Investigation of Small Laccase Immobilized on Carbon Nanomaterials for Direct Bioelectrocatalysis of Oxygen Reduction. Electrochem. Commun. 2019, 101, 82–87. [Google Scholar] [CrossRef]
- Gunne, M.; Höppner, A.; Hagedoorn, P.-L.; Urlacher, V.B. Structural and Redox Properties of the Small Laccase Ssl1 from Streptomyces Sviceus. FEBS J. 2014, 281, 4307–4318. [Google Scholar] [CrossRef]
- Ranieri, A.; Bortolotti, C.A.; di Rocco, G.; Battistuzzi, G.; Sola, M.; Borsari, M. Electrocatalytic Properties of Immobilized Heme Proteins: Basic Principles and Applications. ChemElectroChem 2019, 6, 5172–5185. [Google Scholar] [CrossRef]
- Di Rocco, G.; Battistuzzi, G.; Borsari, M.; Bortolotti, C.A.; Ranieri, A.; Sola, M. The Enthalpic and Entropic Terms of the Reduction Potential of Metalloproteins: Determinants and Interplay. Coord. Chem. Rev. 2021, 445, 214071. [Google Scholar] [CrossRef]
- Ranieri, A.; Battistuzzi, G.; Borsari, M.; Casalini, S.; Fontanesi, C.; Monari, S.; Siwek, M.J.; Sola, M. Thermodynamics and Kinetics of the Electron Transfer Process of Spinach Plastocyanin Adsorbed on a Modified Gold Electrode. J. Electroanal. Chem. 2009, 626, 123–129. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Borsari, M.; di Rocco, G.; Ranieri, A.; Sola, M. Enthalpy/Entropy Compensation Phenomena in the Reduction Thermodynamics of Electron Transport Metalloproteins. JBIC J. Biol. Inorg. Chem. 2004, 9, 23–26. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Borsari, M.; Canters, G.W.; di Rocco, G.; de Waal, E.; Arendsen, Y.; Leonardi, A.; Ranieri, A.; Sola, M. Ligand Loop Effects on the Free Energy Change of Redox and PH-Dependent Equilibria in Cupredoxins Probed on Amicyanin Variants. Biochemistry 2005, 44, 9944–9949. [Google Scholar] [CrossRef]
- Ranieri, A.; Battistuzzi, G.; Borsari, M.; Bortolotti, C.A.; di Rocco, G.; Sola, M. PH and Solvent H/D Isotope Effects on the Thermodynamics and Kinetics of Electron Transfer for Electrode-Immobilized Native and Urea-Unfolded Stellacyanin. Langmuir 2012, 28, 15087–15094. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Bellei, M.; Leonardi, A.; Pierattelli, R.; de Candia, A.; Vila, A.J.; Sola, M. Reduction Thermodynamics of the T1 Cu Site in Plant and Fungal Laccases. JBIC J. Biol. Inorg. Chem. 2005, 10, 867–873. [Google Scholar] [CrossRef]
- Olloqui-Sariego, J.L.; Márquez, I.; Frutos-Beltrán, E.; Díaz-Moreno, I.; de la Rosa, M.A.; Calvente, J.J.; Andreu, R.; Díaz-Quintana, A. Key Role of the Local Hydrophobicity in the East Patch of Plastocyanins on Their Thermal Stability and Redox Properties. ACS Omega 2018, 3, 11447–11454. [Google Scholar] [CrossRef]
- Monari, S.; Battistuzzi, G.; Dennison, C.; Borsari, M.; Ranieri, A.; Siwek, M.J.; Sola, M. Factors Affecting the Electron Transfer Properties of an Immobilized Cupredoxin. J. Phys. Chem. C 2010, 114, 22322–22329. [Google Scholar] [CrossRef]
- Ferraroni, M.; Westphal, A.H.; Borsari, M.; Tamayo-Ramos, J.A.; Briganti, F.; de Graaff, L.H.; van Berkel, W.J.H. Structure and Function of Aspergillus Niger Laccase McoG. Biocatalysis 2017, 3, 1–21. [Google Scholar] [CrossRef]
- Alvarez-Paggi, D.; Zitare, U.A.; Szuster, J.; Morgada, M.N.; Leguto, A.J.; Vila, A.J.; Murgida, D.H. Tuning of Enthalpic/Entropic Parameters of a Protein Redox Center through Manipulation of the Electronic Partition Function. J. Am. Chem. Soc. 2017, 139, 9803–9806. [Google Scholar] [CrossRef] [PubMed]
- Laviron, E. General Expression of the Linear Potential Sweep Voltammogram in the Case of Diffusionless Electrochemical Systems. J. Electroanal. Chem. Interfacial Electrochem. 1979, 101, 19–28. [Google Scholar] [CrossRef]
- Song, S.; Clark, R.A.; Bowden, E.F.; Tarlov, M.J. Characterization of Cytochrome c/Alkanethiolate Structures Prepared by Self-Assembly on Gold. J. Phys. Chem. 1993, 97, 6564–6572. [Google Scholar] [CrossRef]
- Yee, E.L.; Weaver, M.J. Functional Dependence upon Ligand Composition of the Reaction Entropies for Some Transition-Metal Redox Couples Containing Mixed Ligands. Inorg. Chem. 1980, 19, 1077–1079. [Google Scholar] [CrossRef]
- Weaver, M.J. Activation Parameters for Simple Electrode Reactions. Application to the Elucidation of Ion-Solvent Interactions in the Transition State for Heterogeneous Electron Transfer. J. Phys. Chem. 1979, 83, 1748–1757. [Google Scholar] [CrossRef]
- Tarlov, M.J.; Bowden, E.F. Electron-Transfer Reaction of Cytochrome c Adsorbed on Carboxylic Acid Terminated Alkanethiol Monolayer Electrodes. J. Am. Chem. Soc. 1991, 113, 1847–1849. [Google Scholar] [CrossRef]
- Nahir, T.M.; Clark, R.A.; Bowden, E.F. Linear-Sweep Voltammetry of Irreversible Electron Transfer in Surface-Confined Species Using the Marcus Theory. Anal Chem 1994, 66, 2595–2598. [Google Scholar] [CrossRef]
- Becka, A.M.; Miller, C.J. Electrochemistry at.Omega.-Hydroxy Thiol Coated Electrodes. 3. Voltage Independence of the Electron Tunneling Barrier and Measurements of Redox Kinetics at Large Overpotentials. J. Phys. Chem. 1992, 96, 2657–2668. [Google Scholar] [CrossRef]
- Finklea, H.O.; Hanshew, D.D. Electron-Transfer Kinetics in Organized Thiol Monolayers with Attached Pentaammine(Pyridine)Ruthenium Redox Centers. J. Am. Chem. Soc. 1992, 114, 3173–3181. [Google Scholar] [CrossRef]
- Ullmann, G.M.; Kostic, N.M. Electron-Tunneling Paths in Various Electrostatic Complexes between Cytochrome c and Plastocyanin. Anisotropy of the Copper-Ligand Interactions and Dependence of the Iron-Copper Electronic Coupling on the Metalloprotein Orientation. J. Am. Chem. Soc. 1995, 117, 4766–4774. [Google Scholar] [CrossRef]
- Monari, S.; Battistuzzi, G.; Bortolotti, C.A.; Yanagisawa, S.; Sato, K.; Li, C.; Salard, I.; Kostrz, D.; Borsari, M.; Ranieri, A.; et al. Understanding the Mechanism of Short-Range Electron Transfer Using an Immobilized Cupredoxin. J. Am. Chem. Soc. 2012, 134, 11848–11851. [Google Scholar] [CrossRef] [PubMed]
- Murgida, D.H.; Hildebrandt, P. Heterogeneous Electron Transfer of Cytochrome c on Coated Silver Electrodes. Electric Field Effects on Structure and Redox Potential. J. Phys. Chem. B 2001, 105, 1578–1586. [Google Scholar] [CrossRef]
- Hildebrandt, P.; Murgida, D.H. Electron Transfer Dynamics of Cytochrome c Bound to Self-Assembled Monolayers on Silver Electrodes. Bioelectrochemistry 2002, 55, 139–143. [Google Scholar] [CrossRef]
- Blanford, C.F.; Heath, R.S.; Armstrong, F.A. A Stable Electrode for High-Potential, Electrocatalytic O2 Reduction Based on Rational Attachment of a Blue Copper Oxidase to a Graphite Surface. Chem. Commun. 2007, 1710, 1710–1712. [Google Scholar] [CrossRef] [PubMed]
- Collman, J.P.; Devaraj, N.K.; Decréau, R.A.; Yang, Y.; Yan, Y.-L.; Ebina, W.; Eberspacher, T.A.; Chidsey, C.E.D. A Cytochrome c Oxidase Model Catalyzes Oxygen to Water Reduction Under Rate-Limiting Electron Flux. Science (1979) 2007, 315, 1565–1568. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.-J.; Brunner, S.; Wieland, M. Reactions of Surface Amines with Heterobifunctional Cross-Linkers Bearing Both Succinimidyl Ester and Maleimide for Grafting Biomolecules. J. Phys. Chem. B 2004, 108, 16508–16517. [Google Scholar] [CrossRef]
- Sharma, R.; Deacon, S.E.; Nowak, D.; George, S.E.; Szymonik, M.P.; Tang, A.A.S.; Tomlinson, D.C.; Davies, A.G.; McPherson, M.J.; Wälti, C. Label-Free Electrochemical Impedance Biosensor to Detect Human Interleukin-8 in Serum with Sub-Pg/Ml Sensitivity. Biosens. Bioelectron. 2016, 80, 607–613. [Google Scholar] [CrossRef]
- Tsai, T.-C.; Liu, C.-W.; Wu, Y.-C.; Ondevilla, N.A.P.; Osawa, M.; Chang, H.-C. In Situ Study of EDC/NHS Immobilization on Gold Surface Based on Attenuated Total Reflection Surface-Enhanced Infrared Absorption Spectroscopy (ATR-SEIRAS). Colloids Surf. B Biointerfaces 2019, 175, 300–305. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Borsari, M.; Loschi, L.; Sola, M. Redox Thermodynamics, Acid-Base Equilibria and Salt-Induced Effects for the Cucumber Basic Protein. General Implications for Blue-Copper Proteins. JBIC J. Biol. Inorg. Chem. 1997, 2, 350–359. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Borsari, M.; Loschi, L.; Righi, F.; Sola, M. Redox Thermodynamics of Blue Copper Proteins. J. Am. Chem. Soc. 1999, 121, 501–506. [Google Scholar] [CrossRef]
- Yee, E.L.; Cave, R.J.; Guyer, K.L.; Tyma, P.D.; Weaver, M.J. A Survey of Ligand Effects upon the Reaction Entropies of Some Transition Metal Redox Couples. J. Am. Chem. Soc. 1979, 101, 1131–1137. [Google Scholar] [CrossRef]
- Taniguchi, V.T.; Sailasuta-Scott, N.; Anson, F.C.; Gray, H.B. Thermodynamics of Metalloprotein Electron Transfer Reactions. Pure Appl. Chem. 1980, 52, 2275–2281. [Google Scholar] [CrossRef]
- Britz, D. IR Elimination in Electrochemical Cells. J. Electroanal. Chem. Interfacial Electrochem. 1978, 88, 309–352. [Google Scholar] [CrossRef]
- Kwan, P.; Schmitt, D.; Volosin, A.M.; McIntosh, C.L.; Seo, D.-K.; Jones, A.K. Spectroelectrochemistry of Cytochrome c and Azurin Immobilized in Nanoporous Antimony-Doped Tin Oxide. Chem. Commun. 2011, 47, 12367. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E°′ (V) (vs. SHE) b | Γ0 (pmol cm−2) b | ΔH°’rc (kJ/mol) | ΔS°′rc (J/mol K) | |
|---|---|---|---|---|
| MUA | +0.097 | 1.00 | −33.1 | −80 |
| 3:1 MUA/MU | +0.140 | 0.71 | −38.8 | −86 |
| 1:1 MUA/MU | +0.158 | 0.32 | −46.7 | −107 |
| 1:2 MUA/MU | +0.199 | 0.15 | −60.1 | −138 |
| MUA (EDC/NHS) | +0.086 | 4.63 | −32.7 | −83 |
| 3:1 MUA/MU (EDC/NHS), pH 7.0 | +0.115 | 1.22 | −39.8 | −98 |
| 1:2 MUA/MU (EDC/NHS), pH 7.0 | +0.145 | 0.49 | −52.1 | −128 |
| 1:1 MUA/MU (EDC/NHS), pH 7.0 | +0.126 | 1.51 | −45.5 | −114 |
| 1:1 MUA/MU (EDC/NHS), pH 8.4 | +0.057 | 1.43 | −47.0 | −141 |
| 1:1 MUA/MU (EDC/NHS), pH 9.6 | −0.021 | 1.58 | −44.2 | −157 |
| Protein | E°′ (V) (vs. SHE) a | ΔS°′rc (J/mol K) | ΔH°’rc (kJ/mol) | Immobilization | Ref |
|---|---|---|---|---|---|
| SLAC (S. coelicolor) | +0.126 | −114 | −45.5 | covalent 1:1 MUA:MU | this work |
| SLAC (S. coelicolor) | +0.158 | −107 | −46.7 | electrostatic 1:1 MUA:MU | this work |
| Plastocyanin (S. oleracea) | +0.090 | −61 | −26.8 | electrostatic 1:1 MUA:MU | [54] |
| Plastocyanin (P. laminosum) | +0.369 | −65 | −54 | cysteamine | [59] |
| Synechocystis sp. PCC 6803 | +0.365 | −41 | −47 | cysteamine | [59] |
| Azurin (P. aeruginosa) | +0.272 | −14 | −30.4 | Decane-1-thiol | [60] |
| Stellacyanin (Rhus vernicifera) | +0.143 | −54 | −29.6 | decane-1-thiol | [57] |
| Laccase McoG (A. niger) | +0.453 | −28 | −51.9 | decane-1-thiol | [61] |
| Cytochrome c oxidase CuA | +0.261 | −42 | −38 | 6-mercapto-1-hexanol | [62] |
| ks (5 °C) | ks (15 °C) | ks (25 °C) | ΔH# | λ | Lower-Upper Tunneling Distance | |
|---|---|---|---|---|---|---|
| s−1 | s−1 | s−1 | kJ/mol | eV | Å | |
| pH 7.0 a | 8 | 10 | 11 | 9.1 | 0.361 | 20.3–23.3, |
| pH 8.4 a | 14 | 15 | 17 | 6.8 | 0.270 | 20.8–23.8 |
| pH 9.6 a | 7 | 9 | 10 | 10.4 | 0.413 | 19.9–22.9 |
| pH 7.0 b | 9 | 10 | 12 | 9.7 | 0.385 | 20.1–23.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Rocco, G.; Battistuzzi, G.; Ranieri, A.; Bortolotti, C.A.; Borsari, M.; Sola, M. Thermodynamics and Kinetics of Electron Transfer of Electrode-Immobilized Small Laccase from Streptomyces coelicolor. Molecules 2022, 27, 8079. https://doi.org/10.3390/molecules27228079
Di Rocco G, Battistuzzi G, Ranieri A, Bortolotti CA, Borsari M, Sola M. Thermodynamics and Kinetics of Electron Transfer of Electrode-Immobilized Small Laccase from Streptomyces coelicolor. Molecules. 2022; 27(22):8079. https://doi.org/10.3390/molecules27228079
Chicago/Turabian StyleDi Rocco, Giulia, Gianantonio Battistuzzi, Antonio Ranieri, Carlo Augusto Bortolotti, Marco Borsari, and Marco Sola. 2022. "Thermodynamics and Kinetics of Electron Transfer of Electrode-Immobilized Small Laccase from Streptomyces coelicolor" Molecules 27, no. 22: 8079. https://doi.org/10.3390/molecules27228079
APA StyleDi Rocco, G., Battistuzzi, G., Ranieri, A., Bortolotti, C. A., Borsari, M., & Sola, M. (2022). Thermodynamics and Kinetics of Electron Transfer of Electrode-Immobilized Small Laccase from Streptomyces coelicolor. Molecules, 27(22), 8079. https://doi.org/10.3390/molecules27228079