


Collection of Hairy Roots as a Basis for Fundamental and Applied Research

 ,
,
Abstract
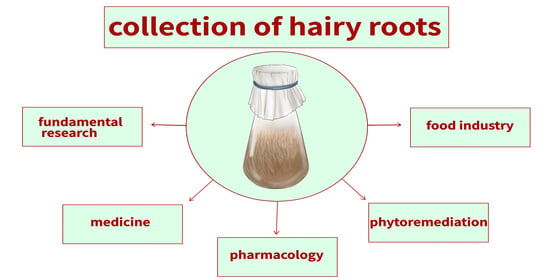
1. Introduction
2. The History of the Development of the Hairy Roots Trend in the World

{kind=link}
{kind=link}
{kind=link}
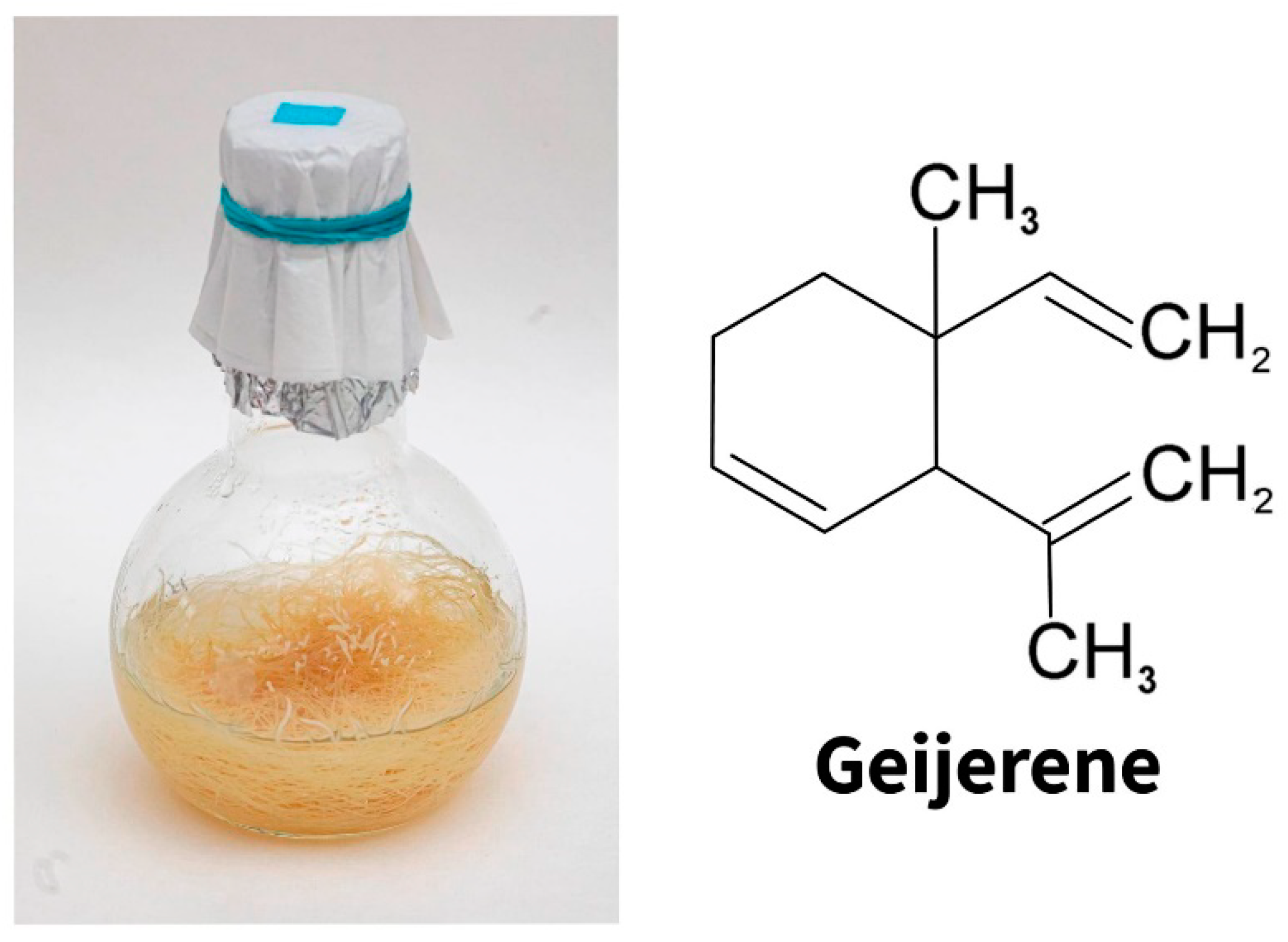{kind=link}
{kind=link}
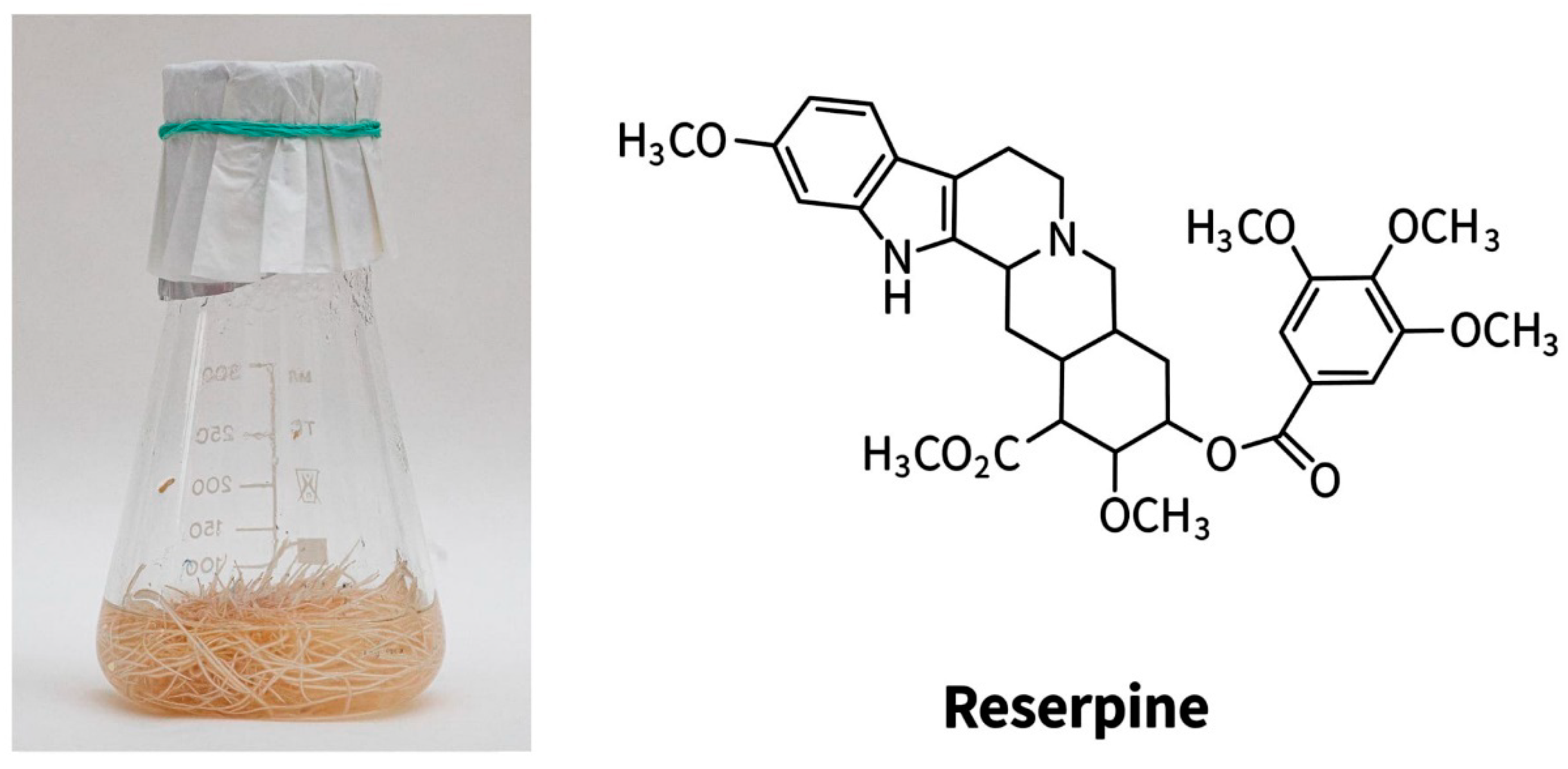{kind=link}
{kind=link}
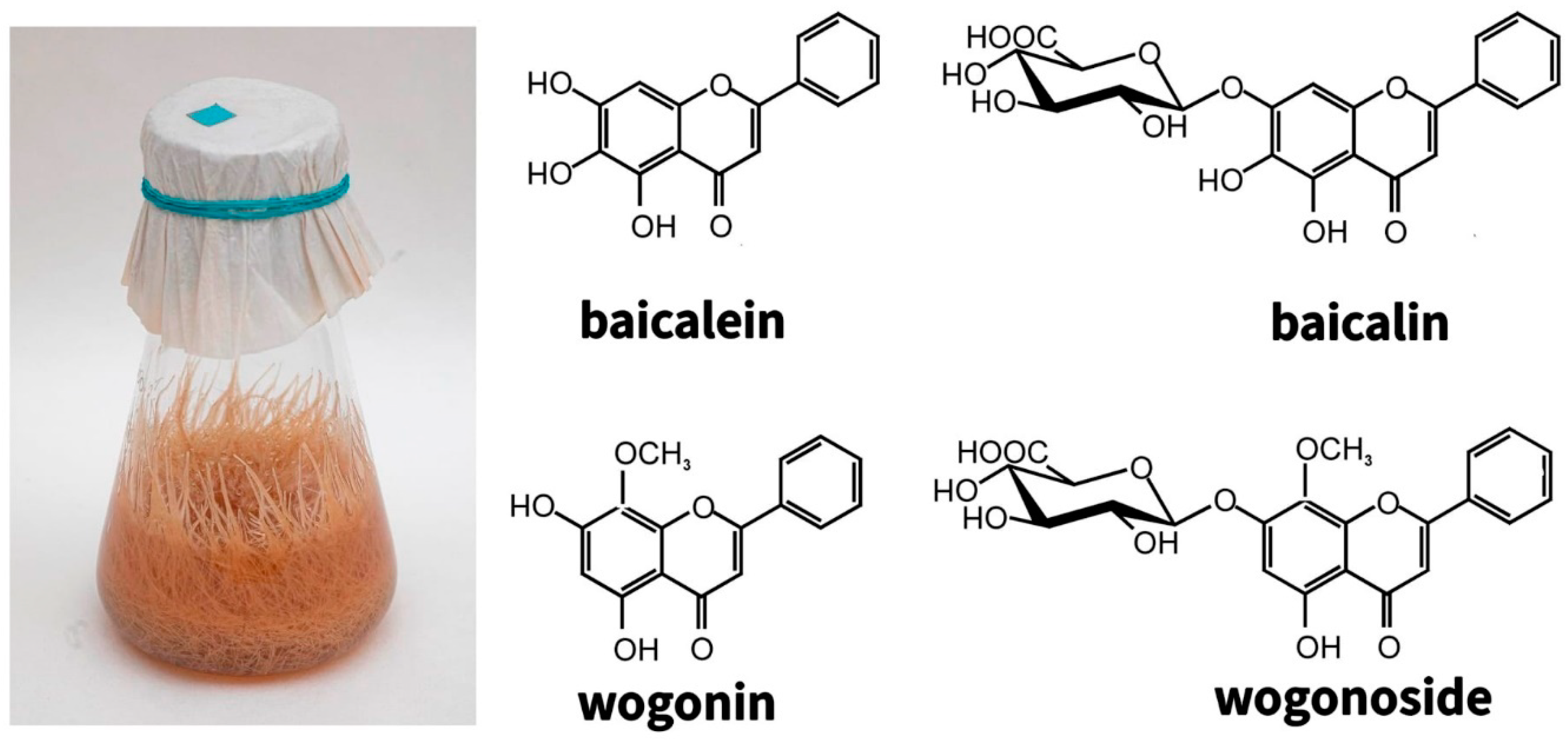{kind=link}
{kind=link}
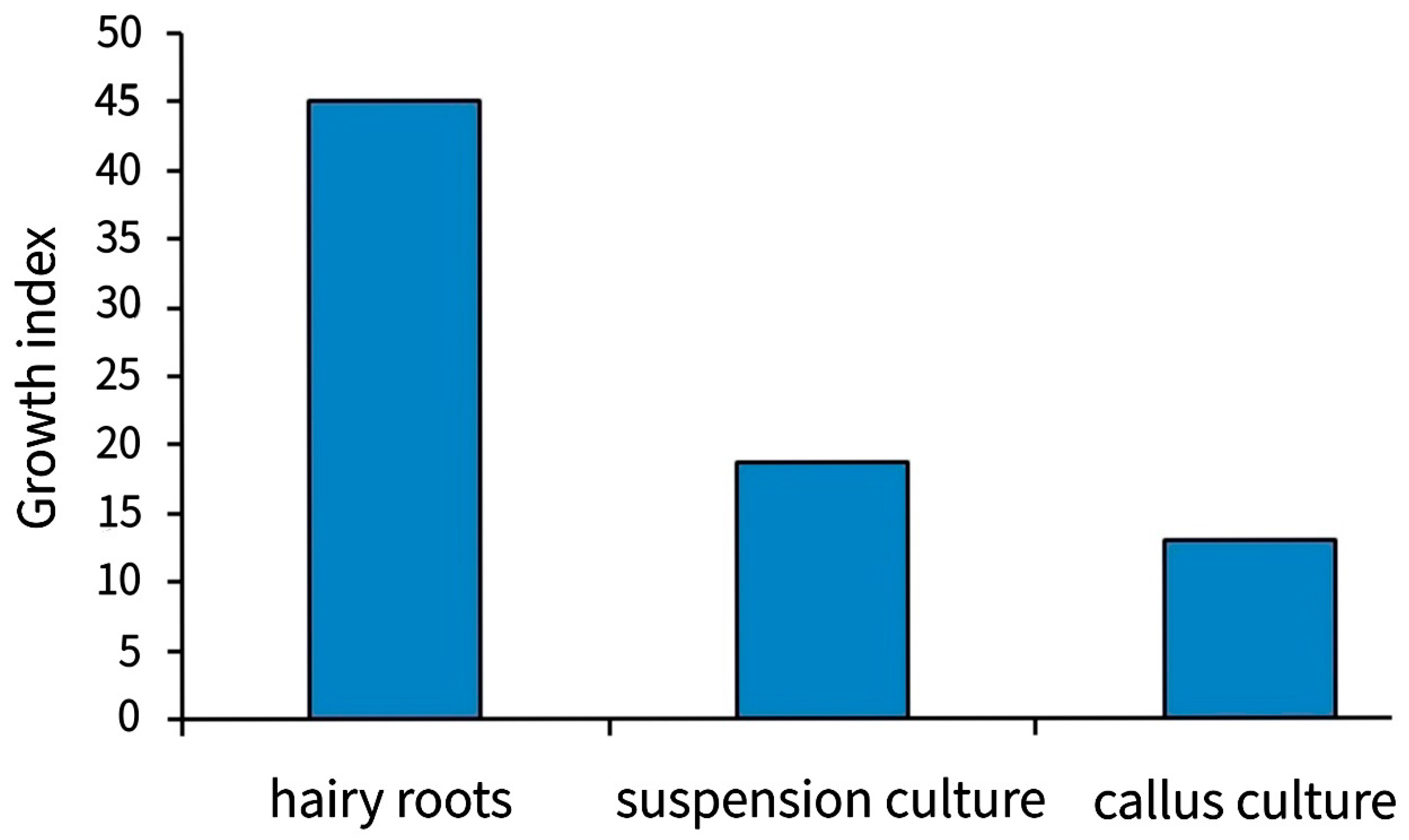{kind=link}
| Species | Analyzed Metabolites | Metabolites Content in Hairy Roots | Metabolites in Plants | Metabolites in Callus Suspension | Reference |
|---|---|---|---|---|---|
| Atropa belladonna | Total alkaloids contents | 1.1–8 mg/gDW | Intact roots—0.3 mg/g DW | [78] | |
| Atropa belladonna | Total alkaloids contents | 1.32% | In untransformed roots—0.8% | [79] | |
| Artemisia dubui | Artemisin | 0.603–0.753%. | In untransformed roots—0.001% | [80] | |
| Artemisia sp. | Artemisinin and its co-products | 1.02 mg/g DW | In untransformed roots—up to 0.687 mg/g DW | [81] | |
| Panax ginseng | Ginsenoside | Total content 5.44 mg/g DW | Total content in untransformed roots—4.55 mg/g DW | [82] | |
| Panax ginseng | Saponins | 2–2.4 times higher compared with native root | [83] | ||
| Panax ginseng | Ginsenoside | 2.88% of dry weight when cultured in 1/8 MS medium for 8 weeks | 2.56% of dry weight (cultivated roots were 5 years old) | [84] | |
| Rubia yunnanensis | Rubiaceae-type cyclopeptides (RAs) | Amount of (RAs) in 1/2 MS liquid medium—4.611 μg/g DW | In plants in vitro—0.331 μg/g and 4.096 μg/g DW for shoots and roots, respectively. Amount of RAs in seed-borne plants—80.296 μg/g, quinones—7409 μg/g DW | In calli—1.082 μg/g DW | [85] |
| Rubia yunnanensis | Quinones | 5067 μg/g DW | 24–132 μg/g DW in plants in vitro; in seed-borne plants 7409 μg/g DW | In calli—338 μg/g DW | [85] |
| Rubia cordifolia Linn | Total phenolic contents | 139.7 mg/g DW | 41.02 mg/g DW of field grown roots | - | [86] |
| Rubia cordifolia Linn | Alizarin | In 5.16-fold than normal roots of field grown plants | 5.16 times lower in the roots of the field-grown plants compared with the hairy roots | [86] | |
| Withania sominifera | Withanolide A | 157.4 μg/g DW | 57.9 μg/g DW | - | [87] |
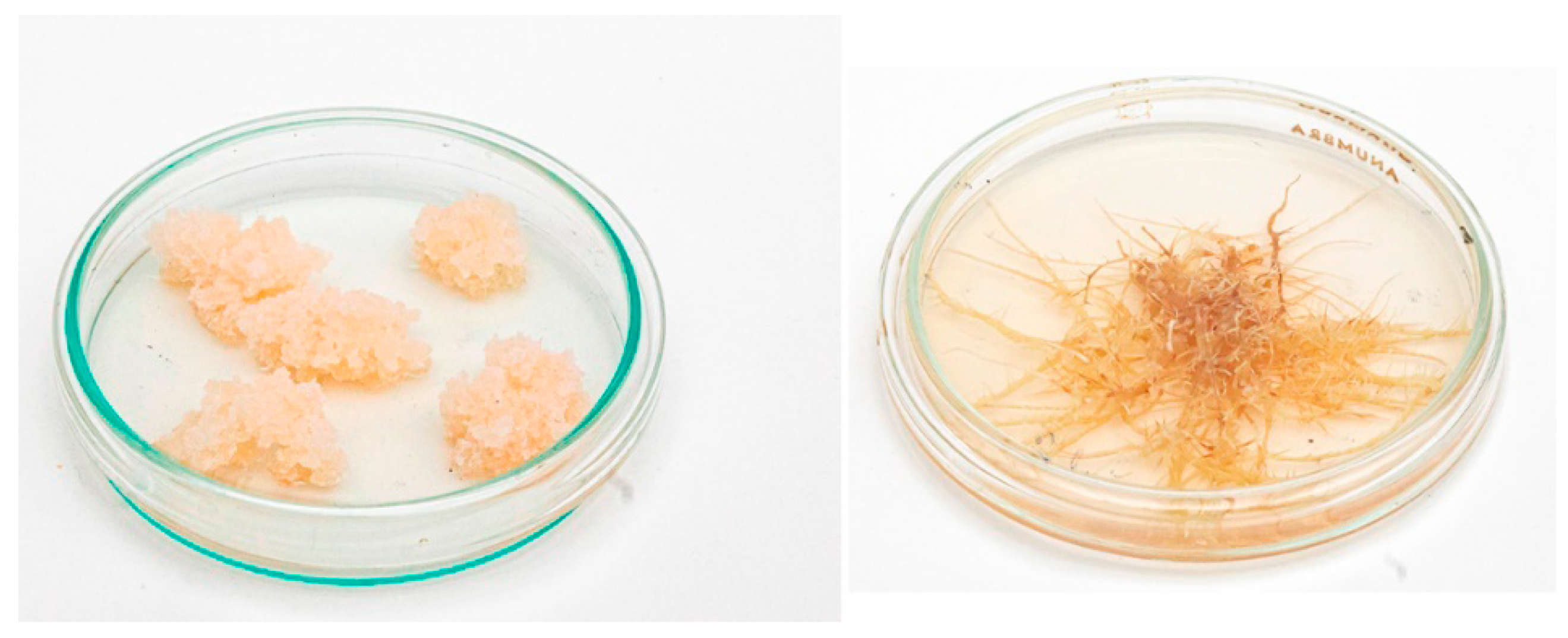
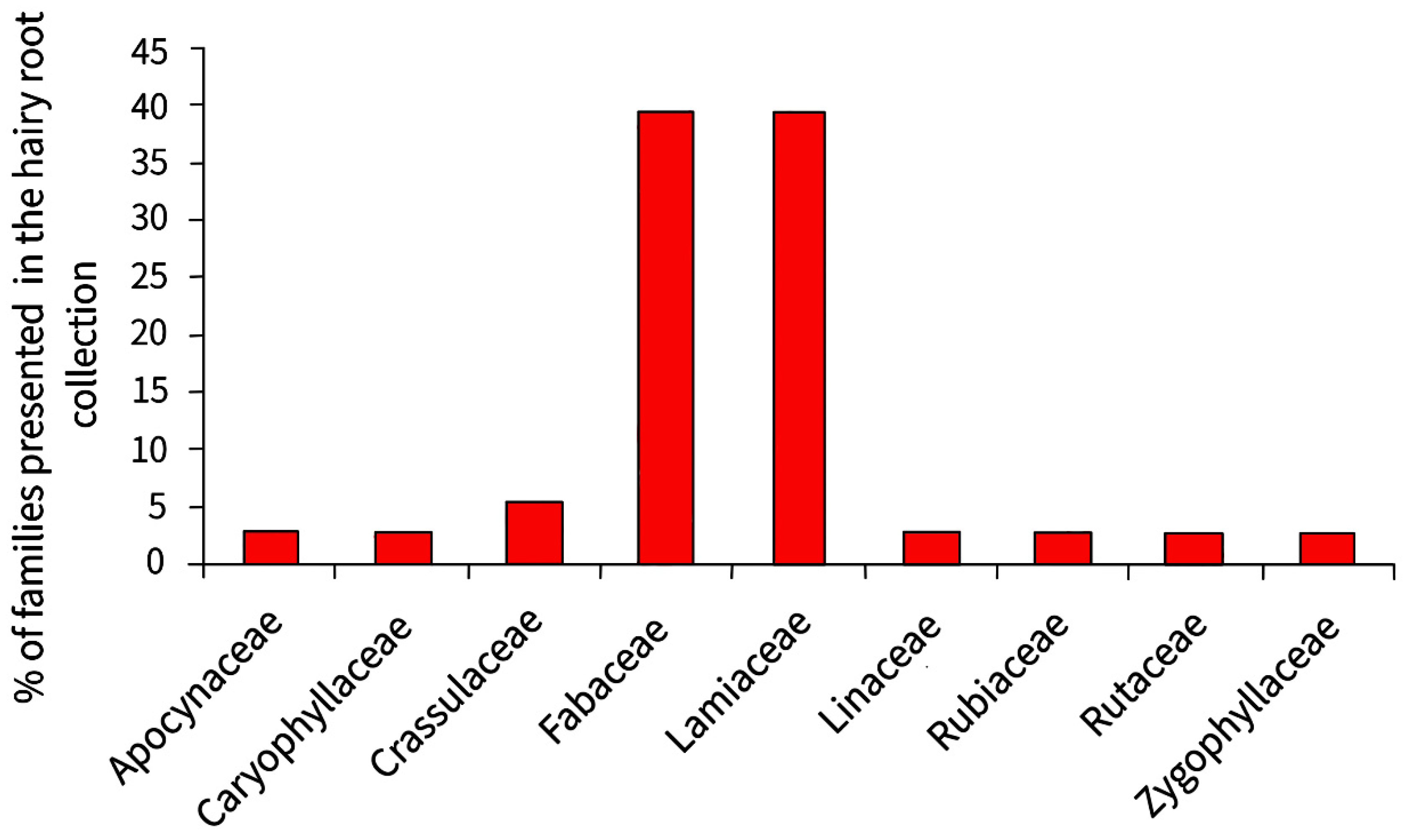
3. The Collection of Hairy Roots of the Institute of Plant Physiology
| Family | Species Name | Number of Lines | Type of the Culture | The Year Obtained | Origin of the Culture | Secondary Metabolites | Note | Possible Use | Literature References in Which This Strain Is Mentioned |
|---|---|---|---|---|---|---|---|---|---|
| Apocynaceae | Rauwolfia serpentina L. (Benth.) | 1 | Hairy roots | 1990 | Leaves of juvenile plants | Contain indole alkaloids (vomilenine, vinorine, perakine) higher than in suspension culture. | The total alkaloid content was 3 times lower compared with the suspension culture. There is no raucaffricine alkaloid, which predominates in suspension culture. | Pharmacology, medicine | [112] |
| Caryophyllaceae | Silene vulgaris L. | 1 | Hairy roots | 2002 | - | - | - | Phytoremediation (accumulation of heavy metals) | - |
| Crassulaceae | Rhodiola quadrifida Pall | 2 | Hairy roots | 2017 | Cotyledons and hypocotyls | Contain salidroside, rosavin | Tyrosol and rosarin are missing. The content is significantly lower than in callus tissue. | Pharmacology, preserving of rare and endangered species | [156] |
| Rhodiola quadrifida Pall | 4 | Calli | 2019 | Hairy roots | Contain salidroside, rosavin, rosin | Thyrosol and rosarin are absent | |||
| Fabaceae | Hedysarum sp. | 5 | Hairy roots | 2001–2019 | Juvenile seedlings | Isoflavones (ononine) | - | Pharmacology, medicine | [157] |
| Glycyrrhiza uralensis L. | 1 | Hairy roots | 1990 | Hypocotyle of juvenile plant | Phenolic derivatives | - | Pharmacology, medicine | - | |
| Lupinus polyphyllus L. | 1 | Hairy roots | 1990 | Hypocotyle of juvenile plant | Isoflavone glycoside | Biotechnology, feed additives | |||
| Ononis sp. L. | 4 | Hairy roots | 1993–1994 | Hypocotyle of juvenile plant | isoflavonoids | ||||
| Sophora korolkovii Koehne | 1 | Hairy roots | 2004 | Hypocotyle of juvenile plant | Phenolic compounds | ||||
| Trifolium repens L. | 1 | Hairy roots | 1991 | Hypocotyle of juvenile plant | A model object for studying arbuscular mycorrhizae | ||||
| Medicago sativa L. | 2 | Hairy roots | 2013 | Hypocotyle of juvenile plant | Phytoremediation | ||||
| Lamiaceae | Scutellaria baicalensis Georgi | 3 | Hairy roots | 1993, 2018, 2021 | Leaves of juvenile plant | Contains flavones (baicalin, baicalein, wogonin, wogonoside) | The content of methylated flavones is higher than in the roots of intact plants | Pharmacology, medicine, food industry | [135,136,137,138,158] |
| Scutellaria baicalensis Georgi | 2 | Calli | Hairy roots | Contains flavones (baicalin, baicalein, wogonin, wogonoside) | The main flavones are baicalin and baicalein, as in the roots of intact plants. | ||||
| Scutellaria androchnoides | 1 | Hairy roots | 2006 | Cotyledon and hypocotyl | The dominant compounds are acteoside (phenylethanoids) and the four main flavones of representatives of Scutellaria sp.: baicalin, wogonoside, wogonin and baicalein | The content of acteoside is 10 times higher, and the content of methylated flavone wogonoside is 3 times higher compared with the roots of intact plants | Pharmacology, medicine | [140] | |
| Scutellaria androchnoides | 1 | Calli | 2006 | Hairy roots | The dominant metabolites are acteoside and the methylated flavones wogonoside and wogonin | The content of acteoside is 2.5 times, wogonoside 1.5 times higher than in the roots of intact plants | Pharmacology, medicine | ||
| Scutellaria przewalskii | 1 | Hairy roots | 2014, 2020 | Cotyledons, hypocotyls of sterile-grown plants | Based on HPLC data, 17 flavones were found, among which the main metabolites are baicalin and wogonoside glucuronides | The content of the main metabolites is higher than in all of the above representatives of Scutellaria | Pharmacology, medicine | [136] | |
| Scutellaria lateriflora | 2 | Hairy roots | 2020 | Hypocotyls of sterile-grown plants | Contains flavones (baicalin, baicalein, wogonin, wogonoside) | The content of flavones is 4.57 mg/g DW, that is lower than in all the species of hairy roots studied by us | Pharmacology, medicine | [136] | |
| Scutellaria pycnoclada | 8 | Hairy roots | 2020 | Hypocotyls of sterile-grown plants | In contrast to other lines of skullcaps, the ratio of main flavones is close to that of the roots of intact plants | Pharmacology, medicine | [136] | ||
| Linaceae | Linum usitatissimum L. | 1 | Hairy roots | 1995 | Cotyledons of sterile-grown seedlings | Cyanogenic glycosides, lignan | Pharmacology, medicine | ||
| Rubiaceae | Rubia tictorum L. | 1 | Hairy roots | 1991 | Anthraquinones | The content of anthraquinones is 2.5% by dry weight. In the roots and rhizomes of an intact plant—5.2% | Pharmacology, medicine | [159] | |
| Rutaceae | Ruta graveolens L. | 1 | Hairy roots | 1991 | Hypocotyle of a juvenile plant | Acridon alkaloids | Study of the spatial distribution of acridone alkaloids | [109] | |
| Ruta graveolens L. | 8 | Calli | 1969, 1970, 1978, 1980, 1999 | Stem of whole plants, hypocotyle of juvenile seedlings, roots of juvenile seedling | Acridonalkaloids | Study of the biosynthesis of acridone alkaloids | [160,161] | ||
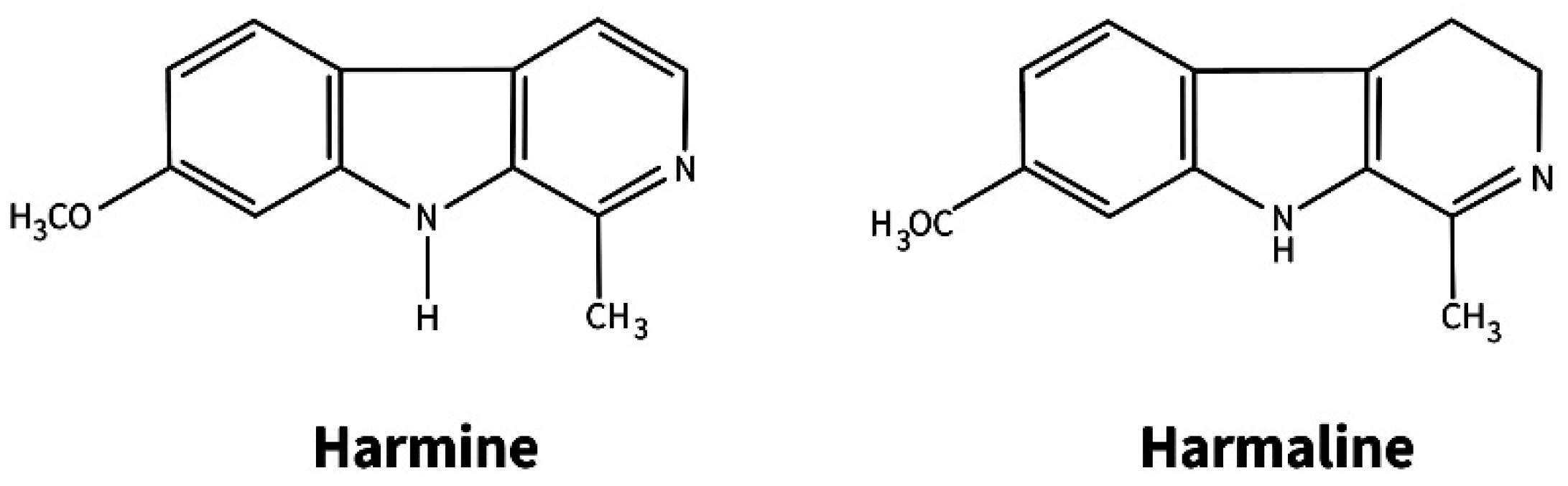
| Zygophyllaceae | Peganum harmala L. | 1 | Hairy roots | 1988 | Stem of a juvenile plant | β-carbolinealkaloids (harmine, harmalol, harmaline), serotonin | Study of the distribution of secondary metabolites in plant roots. Pharmacology | [52] | |
| Peganum harmala L. | 1 | Calli | 1988 | Spontaneous callus formation on a juvenile plant stem | β-carboline alkaloids, serotonin |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef] [PubMed]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Kopaei, M.R. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar] [PubMed]
- Quaglio, D.; Talib, W.H.; Daoud, S.; Mahmod, A.I.; Hamed, R.A.; Awajan, D.; Feras Abuarab, S.; Odeh, L.H.; Khater, S.; Al Kury, L.T. Plants as a source of anticancer agents: From bench to bedside. Molecules 2022, 27, 4818. [Google Scholar] [CrossRef]
- Ramachandra Rao, S.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Barrales-Cureño, H.J.; Valdivia, A.C.R.; Hernández, M.S. Increased production of taxoids in suspension cultures of Taxus globosa after elicitation. Futur. Pharmacol. 2022, 2, 45–54. [Google Scholar] [CrossRef]
- Shen, F.; Long, D.; Yu, T.; Chen, X.; Liao, Y.; Wu, Y.; Lin, X. Vinblastine differs from Taxol as it inhibits the malignant phenotypes of NSCLC cells by increasingthe phosphorylation of Op18/stathmin. Oncol. Rep. 2017, 37, 2481–2489. [Google Scholar] [CrossRef]
- Gallego-Jara, J.; Lozano-Terol, G.; Sola-Martínez, R.A.; Cánovas-Díaz, M.; de Diego Puente, T. A Compressive Review about Taxol®: History and Future Challenges. Molecules 2020, 25, 5986. [Google Scholar]
- Hennessy, B.T.; Coleman, R.L.; Markman, M. Ovarian cancer. Lancet 2009, 374, 1371–1382. [Google Scholar] [CrossRef]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; Mcphail, A.T. Plant Antitumor Agents.VI.The Isolation and Structure of Taxol, a Novel Antileukemic and Antitumor Agent from Taxus brevifolia2. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Gozes, I.; Harrington, C.; Lowery, L.A.; Coffey, E.T.; Varidaki, A.; Hong, Y. Repositioning Microtubule Stabilizing Drugs for Brain Disorders. Front. Cell. Neurosci. 2018, 1, 226. [Google Scholar] [CrossRef]
- Zhang, B.; Maiti, A.; Shively, S.; Lakhani, F.; McDonald-Jones, G.; Bruce, J.; Lee, E.B.; Xie, S.X.; Joyce, S.; Li, C.; et al. Microtubule-binding drugs offset tau sequestration by stabilizing microtubules and reversing fast axonal transport deficits in a tauopathy model. Proc. Natl. Acad. Sci. USA 2004, 102, 227–231. [Google Scholar] [CrossRef]
- Success Story: Taxol. Available online: https://dtp.cancer.gov/timeline/flash/success_stories/S2_taxol.htm (accessed on 7 November 2022).
- Barrales-Cureño, H.J.; Reyes, C.R.; García, I.V.; Valdez, L.G.L.; De Jesús, A.G.; Ruíz, J.A.C.; Herrera, L.M.S.; Caballero, M.C.C.; Magallón, J.A.S.; Perez, J.E.; et al. Alkaloids of pharmacological importance in Catharanthus roseus. Alkaloids-Their. Importance Nat. Hum. Life 2019, 1, 18. [Google Scholar] [CrossRef]
- Coderch, C.; Morreale, A.; Gago, F. Tubulin-based Structure-affinity Relationships for Antimitotic Vinca Alkaloids. Anticancer. Agents Med. Chem. 2012, 12, 219–225. [Google Scholar] [CrossRef]
- Heilig, M.L. United States Patent Office. ACM SIGGRAPH Comput. Graph. 1994, 28, 131–134. [Google Scholar] [CrossRef]
- Mehrotra, S.; Rahman, L.U.; Kukreja, A.K. An extensive case study of hairy-root cultures for enhanced secondary-metabolite production through metabolic-pathway engineering. Biotechnol. Appl. Biochem. 2010, 56, 161–172. [Google Scholar] [CrossRef]
- Yue, W.; Ming, Q.L.; Lin, B.; Rahman, K.; Zheng, C.J.; Han, T.; Qin, L.P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef]
- Fazili, M.A.; Bashir, I.; Ahmad, M.; Yaqoob, U.; Geelani, S.N. In vitro strategies for the enhancement of secondary metabolite production in plants: A review. Bull. Natl. Res. Cent. 2022, 46, 35. [Google Scholar] [CrossRef]
- Yazaki, K. Lithospermum erythrorhizon cell cultures: Present and future aspects. Plant Biotechnol. 2017, 34, 131. [Google Scholar] [CrossRef] [PubMed]
- Alcalde, M.A.; Perez-Matas, E.; Escrich, A.; Cusido, R.M.; Palazon, J.; Bonfill, M. Biotic Elicitors in Adventitious and Hairy Root Cultures: A Review from 2010 to 2022. Molecules 2022, 27, 5253. [Google Scholar] [CrossRef]
- Królicka, A.; Staniszewska, I.; Bielawski, K.; Maliński, E.; Szafranek, J.; Lojkowska, E. Establishment of hairy root cultures of Ammi majus. Plant Sci. 2001, 160, 259–264. [Google Scholar] [CrossRef]
- Purwianingsih, W.; Hidayat, R.Y.; Rahmat, A. Increasing anthraquinone compounds on callus leaf Morinda citrifolia (L.) by elicitation method using chitosan shell of shrimps (Penaeus monodon). J. Phys. Conf. Ser. 2019, 1280, 022001. [Google Scholar] [CrossRef]
- Yamada, Y.; Endo, T. Tropane alkaloid production in cultured cells of Duboisia leichhardtii. Plant Cell Rep. 1984, 3, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Yamada, Y. Scopolamine production in suspension cultures and redifferentiated roots of Hyoscyamus niger. Planta Med. 1983, 47, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Häkkinen, S.T.; Moyano, E.; Cusido, R.M.; Oksman-Caldentey, K.M. Exploring the Metabolic Stability of Engineered Hairy Roots after 16 Years Maintenance. Front Plant Sci. 2016, 7, 1486. [Google Scholar] [CrossRef] [PubMed]
- Thi, L.; Tien, T. Root Cultures for Secondary Products. Plant Roots 2020, 425–574. [Google Scholar] [CrossRef]
- Khanam, M.N.; Anis, M.; Bin Javed, S.; Mottaghipisheh, J.; Csupor, D. Adventitious root culture-an alternative strategy for secondary metabolite production: A review. Agronomy 2022, 12, 1178. [Google Scholar] [CrossRef]
- Praveen, N.; Manohar, S.H.; Naik, P.M.; Nayeem, A.; Jeong, J.H.; Murthy, H.N. Production of andrographolide from adventitious root cultures of Andrographis paniculata. Curr. Sci. 2009, 2009, 694–697. [Google Scholar]
- Ho, T.T.; Lee, J.D.; Jeong, C.S.; Paek, K.Y.; Park, S.Y. Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Shi, M.; Liao, P.; Nile, S.H.; Georgiev, M.I.; Kai, G. Biotechnological Exploration of Transformed Root Culture for Value-Added Products. Trends Biotechnol. 2021, 39, 137–149. [Google Scholar] [CrossRef]
- Sharma, P.; Padh, H.; Shrivastava, N. Hairy root cultures: A suitable biological system for studying secondary metabolic pathways in plants. Eng. Life Sci. 2013, 13, 62–75. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K. Hairy root culture for mass-production of high-value secondary metabolites. Crit. Rev. Biotechnol. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.M.; Ritala, A.; Cardon, F. Hairy Root Cultures—A Versatile Tool With Multiple Applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef]
- Rekha, K.; Thiruvengadam, M. Secondary Metabolite Production in Transgenic Hairy Root Cultures of Cucurbits. Transgenesis Second. Metab. 2017, 267, 267–293. [Google Scholar] [CrossRef]
- Giri, A.; Narasu, M.L. Transgenic hairy roots: Recent trends and applications. Biotech. Adv. 2000, 18, 1–22. [Google Scholar] [CrossRef]
- Morey, K.J.; Peebles, C.A.M. Hairy roots: An untapped potential for production of plant products. Front. Plant Sci. 2022, 13, 2808. [Google Scholar] [CrossRef]
- Sun, J.; Ma, L.; San, K.Y.; Peebles, C.A.M. Still stable after 11 years: A Catharanthus roseus Hairy root line maintains inducible expression of anthranilate synthase. Biotechnol. Prog. 2017, 33, 66–69. [Google Scholar] [CrossRef]
- Smirnov, A.M. Rost i Metabolizm Izolirovannyh Kornej v Steril’noj Kul’ture; Ratner, E.I., Ed.; Izdatel’stvo “Nauka”: Moscow, Russia, 1970. [Google Scholar]
- Smith, E.F.; Brown, N.A.; Townsend, C.O. Crown-Gall of Plants: Its Cause and Remedy; US Government Printing Office: Washington, DC, USA, 1911; pp. 213, 1–215.
- Chilton, M.D.; Tepfer, D.A.; Petit, A.; David, C.; Casse Delbart, F.; Tempé, J. Agrobacterium rhizogenes inserts T-DNA into the genomes of the host plant root cells. Nature 1982, 295, 432–434. [Google Scholar] [CrossRef]
- Doran, P.M. Hairy Roots: Culture and Applications; Doran, P.M., Ed.; CRC Press: Sydney, Australia, 2020; ISBN 9781000725339. [Google Scholar]
- Kuzovkina, I.N.; Schneider, B. Genetically transformed root cultures—Generation, properties and application in plant sciences. Prog. Bot. 2006, 67, 275–314. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, Y.; Verma, P.K.; Vakhlu, J. Establishment of Agrobacterium rhizogenes-mediated hairy root transformation of Crocus sativus L. 3 Biotech 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Rahimi, K.; Haghbeen, K.; Marefatjo, J.; Jazii, F.R.; Sheikhani, R. Successful production of hairy root of Valeriana sisymbriifolium by Agrobacterium rhizogenes. Biotechnology 2008, 7, 200–204. [Google Scholar] [CrossRef]
- Chandra, S.; Chandra, R. Engineering secondary metabolite production in hairy roots. Phytochem. Rev. 2011, 10, 371–395. [Google Scholar] [CrossRef]
- Akutsu, M.; Ishizaki, T.; Sato, H. Transformation of the monocotyledonous Alstroemeria by Agrobacterium tumefaciens. Plant Cell Rep. 2004, 22, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Bathoju, G.; Rao, K.; Giri, A. Production of sapogenins (stigmasterol and hecogenin) from genetically transformed hairy root cultures of Chlorophytum borivilianum (Safed musli). Plant Cell Tissue Organ Cult. 2017, 131, 369–376. [Google Scholar] [CrossRef]
- Xu, H.; Zhou, X.; Lu, J.; Wang, J.; Wang, X. Hairy roots induced by Agrobacterium rhizogenes and production of regenerative plants in hairy root cultures in maize. Sci. China Ser. C Life Sci. 2006, 49, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Sanford, J.C. Biolistic plant transformation. Physiol. Plant. 1990, 79, 206–209. [Google Scholar] [CrossRef]
- Yasybaeva, G.; Vershinina, Z.; Kuluev, B.; Mikhaylova, E.; Baymiev, A.; Chemeris, A. Biolistic-mediated plasmid-free transformation for induction of hairy roots in tobacco plants. Plant Root 2017, 11, 33–39. [Google Scholar] [CrossRef][Green Version]
- Vogt, T.; Liu, Q.; John Loake, G.; Hou, W.; Shakya, P.; Franklin, G. A Perspective on Hypericum perforatum Genetic Transformation. Front. Plant Sci. 2016, 1, 879. [Google Scholar] [CrossRef]
- Christou, P. Strategies for variety-independent genetic transformation of important cereals, legumes and woody species utilizing particle bombardment. Euphytica 1995, 85, 13–27. [Google Scholar] [CrossRef]
- Barampuram, S.; Zhang, Z.J. Recent advances in plant transformation. Methods Mol. Biol. 2011, 701, 1–35. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Chen, S.T.; Agnolet, S.; Hegelund, J.N.; Stanstrup, J.; Christensen, J.H.; Müller, R.; Lütken, H. Ethephon-induced changes in antioxidants and phenolic compounds in anthocyanin-producing black carrot hairy root cultures. J. Exp. Bot. 2020, 71, 7030–7045. [Google Scholar] [CrossRef]
- Wawrosch, C.; Zotchev, S.B. Production of bioactive plant secondary metabolites through in vitro technologies-status and outlook. Appl. Microbiol. Biotechnol. 2021, 105, 6649–6668. [Google Scholar] [CrossRef]
- Massa, S.; Paolini, F.; Marino, C.; Franconi, R.; Venuti, A. Bioproduction of a therapeutic vaccine against human papillomavirus in tomato hairy root cultures. Front. Plant Sci. 2019, 10, 452. [Google Scholar] [CrossRef]
- Skarjinskaia, M.; Ruby, K.; Araujo, A.; Taylor, K.; Gopalasamy-Raju, V.; Musiychuk, K.; Chichester, J.A.; Palmer, G.A.; de la Rosa, P.; Mett, V.; et al. Hairy Roots as a Vaccine Production and Delivery System. Adv. Biochem. Eng. Biotechnol. 2013, 134, 115–134. [Google Scholar] [CrossRef]
- Donini, M.; Marusic, C. Hairy roots as bioreactors for the production of biopharmaceuticals. In Hairy Roots: An Effective Tool of Plant Biotechnology; Springer: Singapore, 2018; pp. 213–225. ISBN 9789811325625. [Google Scholar]
- Zhang, N.; Wright, T.; Wang, X.; Karki, U.; Savary, B.J.; Xu, J. Engineering ‘designer’ glycomodules for boosting recombinant protein secretion in tobacco hairy root culture and studying hydroxyproline-O-glycosylation process in plants. Plant Biotechnol. J. 2019, 17, 1130–1141. [Google Scholar] [CrossRef]
- Majumder, A.; Ray, S.; Jha, S. Hairy Roots and Phytoremediation. Bioprocess. Plant Vitr. Syst. 2016, 1–24. [Google Scholar] [CrossRef]
- Moola, A.K.; Balasubramanian, P.; Satish, L.; Shamili, S.; Ramesh, M.; Kumar, T.S.; Kumari, B.D.R.; Moola, A.K.; Kumar, T.S.; Kumari, B.D.R.; et al. Hairy Roots as a Source for Phytoremediation. In Strategies and Tools for Pollutant Mitigation; Springer: Cham, Switzerland, 2021; pp. 29–47. [Google Scholar] [CrossRef]
- Bulgakov, V.P. Functions of rol genes in plant secondary metabolism. Biotechnol. Adv. 2008, 26, 318–324. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Veremeichik, G.N.; Grigorchuk, V.P.; Rybin, V.G.; Shkryl, Y.N. The rolB gene activates secondary metabolism in Arabidopsis calli via selective activation of genes encoding MYB and bHLH transcription factors. Plant Physiol. Biochem. PPB 2016, 102, 70–79. [Google Scholar] [CrossRef]
- Kim, Y.; Wyslouzil, B.E.; Weathers, P.J. Secondary metabolism of hairy root cultures in bioreactors. Vitr. Cell. Dev. Biol.-Plant 2002, 38, 1–10. [Google Scholar] [CrossRef]
- Bakkali, A.T.; Jaziri, M.; Foriers, A.; Vander Heyden, Y.; Vanhaelen, M.; Homès, J. Lawsone accumulation in normal and transformed cultures of henna, Lawsonia inermis. Plant Cell. Tissue Organ Cult. 1997, 51, 83–87. [Google Scholar] [CrossRef]
- Wallaart, T.E.; Pras, N.; Quax, W.J. Isolation and identification of dihydroartemisinic acid hydroperoxide from Artemisia annua: A novel biosynthetic precursor of artemisinin. J. Nat. Prod. 1999, 62, 1160–1162. [Google Scholar] [CrossRef] [PubMed]
- Patra, N.; Srivastava, A.K. Artemisinin production by plant hairy root cultures in gas- and liquid-phase bioreactors. Plant Cell Rep. 2016, 35, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, R.; Xavier, L.S.E.; Udayakumaran, G. Biotic elicitors: A boon for the in-vitro production of plant secondary metabolites. Plant Cell Tiss Organ Cult. 2022, 149, 7–24. [Google Scholar] [CrossRef]
- Cao, W.; Wang, Y.; Shi, M.; Hao, X.; Zhao, W.; Wang, Y.; Ren, J.; Kai, G. Transcription factor SmWRKY1 positively promotes the biosynthesis of tanshinones in Salvia miltiorrhiza. Front. Plant Sci. 2018, 9, 554. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Hao, X.; Shi, M.; Fu, R.; Wang, Y.; Zhang, Y.; Zhou, W.; Feng, Y.; Makunga, N.P.; Kai, G. Tanshinone production could be increased by the expression of SmWRKY2 in Salvia miltiorrhiza hairy roots. Plant Sci. 2019, 284, 1–8. [Google Scholar] [CrossRef]
- Hao, X.; Pu, Z.; Cao, G.; You, D.; Zhou, Y.; Deng, C.; Shi, M.; Nile, S.H.; Wang, Y.; Zhou, W.; et al. Tanshinone and salvianolic acid biosynthesis are regulated by SmMYB98 in Salvia miltiorrhiza hairy roots. J. Adv. Res. 2020, 23, 1–12. [Google Scholar] [CrossRef]
- Shi, M.; Luo, X.; Ju, G.; Yu, X.; Hao, X.; Huang, Q.; Xiao, J.; Cui, L.; Kai, G. Increased accumulation of the cardio-cerebrovascular disease treatment drug tanshinone in Salvia miltiorrhiza hairy roots by the enzymes 3-hydroxy-3-methylglutaryl CoA reductase and 1-deoxy-d-xylulose 5-phosphate reductoisomerase. Funct. Integr. Genom. 2014, 14, 603–615. [Google Scholar] [CrossRef]
- Shi, M.; Luo, X.; Ju, G.; Li, L.; Huang, S.; Zhang, T.; Wang, H.; Kai, G. Enhanced Diterpene Tanshinone Accumulation and Bioactivity of Transgenic Salvia miltiorrhiza Hairy Roots by Pathway Engineering. J. Agric. Food Chem. 2016, 64, 2523–2530. [Google Scholar] [CrossRef]
- Paul, P.; Singh, S.K.; Patra, B.; Sui, X.; Pattanaik, S.; Yuan, L. A differentially regulated AP2/ERF transcription factor gene cluster acts downstream of a MAP kinase cascade to modulate terpenoid indole alkaloid biosynthesis in Catharanthus roseus. New Phytol. 2017, 213, 1107–1123. [Google Scholar] [CrossRef]
- Park, C.H.; Xu, H.; Yeo, H.J.; Park, Y.E.; Hwang, G.S.; Park, N., II; Park, S.U. Enhancement of the flavone contents of Scutellaria baicalensis hairy roots via metabolic engineering using maize Lc and Arabidopsis PAP1 transcription factors. Metab. Eng. 2021, 64, 64–73. [Google Scholar] [CrossRef]
- Singh, S.; Pandey, P.; Akhtar, M.Q.; Negi, A.S.; Banerjee, S. A new synthetic biology approach for the production of curcumin and its glucoside in Atropa belladonna hairy roots. J. Biotechnol. 2021, 328, 23–33. [Google Scholar] [CrossRef]
- Bonhomme, V.; Laurain-Mattar, D.; Lacoux, J.; Fliniaux, M.A.; Jacquin-Dubreuil, A. Tropane alkaloid production by hairy roots of Atropa belladonna obtained after transformation with Agrobacterium rhizogenes 15834 and Agrobacterium tumefaciens containing rol A, B, C genes only. J. Biotechnol. 2000, 81, 151–158. [Google Scholar] [CrossRef]
- Jung, G.; Tepfer, D. Use of genetic transformation by the Ri T-DNA of Agrobacterium rhizogenes to stimulate biomass and tropane alkaloid production in Atropa belladonna and Calystegia sepium roots grown in vitro. Plant Sci. 1987, 50, 145–151. [Google Scholar] [CrossRef]
- Mannan, A.; Shaheen, N.; Arshad, W.; Qureshi, R.A.; Zia, M.; Mirza, B. Hairy roots induction and artemisinin analysis in Artemisia dubia and Artemisia indica. Afr. J. Biotechnol. 2010, 7, 3288–3292. [Google Scholar] [CrossRef]
- Drobot, K.O.; Matvieieva, N.A.; Ostapchuk, A.M.; Kharkhota, M.A.; Duplij, V.P. Study of artemisinin and sugar accumulation in Artemisia vulgaris and Artemisia dracunculus “hairy” root cultures. Prep. Biochem. Biotechnol. 2017, 47, 776–781. [Google Scholar] [CrossRef]
- Mallol, A.; Cusidó, R.M.; Palazón, J.; Bonfill, M.; Morales, C.; Piñol, M.T. Ginsenoside production in different phenotypes of Panax ginseng transformed roots. Phytochemistry 2001, 57, 365–371. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Furuya, T. Saponin production by cultures of Panax ginseng transformed with Agrobacterium rhizogenes. Plant Cell Rep. 1987, 6, 449–453. [Google Scholar] [CrossRef]
- Washida, D.; Shimomura, K.; Nakajima, Y.; Takido, M.; Kitanaka, S. Ginsenosides in hairy roots of a panax hybrid. Phytochemistry 1998, 49, 2331–2335. [Google Scholar] [CrossRef]
- Miao, Y.; Hu, Y.; Yi, S.; Zhang, X.; Tan, N. Establishment of hairy root culture of Rubia yunnanensis Diels: Production of Rubiaceae-type cyclopeptides and quinones. J. Biotechnol. 2021, 341, 21–29. [Google Scholar] [CrossRef]
- Kudale, S.; Ghatge, S.; Desai, N. Quantification of Phytochemicals in hairy root cultures of Rubia cordifolia Linn. Int. J. Adv. Res. 2015, 3, 903–913. [Google Scholar]
- Murthy, H.N.; Dijkstra, C.; Anthony, P.; White, D.A.; Davey, M.R.; Power, J.B.; Hahn, E.J.; Paek, K.Y. Establishment of Withania somnifera Hairy Root Cultures for the Production of Withanolide A. J. Integr. Plant Biol. 2008, 50, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Berlin, J.; Rügenhagen, C.; Greidziak, N.; Kuzovkina, I.N.; Witte, L.; Wray, V. Biosynthesis of serotonin and β-carboline alkaloids in hairy root cultures of Peganum harmala. Phytochemistry 1993, 33, 593–597. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Gohar, A.; Alterman, I.E. Production of ß-Carboline Alkaloids in Transformed Root Cultures of Peganum harmala L. Zeitschrift Fur Naturforsch.-Sect. C J. Biosci. 1990, 45, 727–728. [Google Scholar] [CrossRef][Green Version]
- Sharifi-Rad, J.; Quispe, C.; Herrera-Bravo, J.; Semwal, P.; Painuli, S.; Özçelik, B.; Hacihasanoǧlu, F.E.; Shaheen, S.; Sen, S.; Acharya, K.; et al. Peganum spp.: A Comprehensive Review on Bioactivities and Health-Enhancing Effects and Their Potential for the Formulation of Functional Foods and Pharmaceutical Drugs. Oxid. Med. Cell. Longev. 2021, 2021, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.-M.; Zhao, T.; Yang, T.; Wang, C.-H.; Bligh, S.W.A.; Wang, Z.-T.; Key, M. HPLC Fingerprints Combined with Principal Component Analysis, Hierarchical Cluster Analysis and Linear Discriminant Analysis for the Classifi cation and Diff erentiation of Peganum sp. Indigenous to China. Phytochem. Anal 2010, 21, 279–289. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, X.; Xie, X.; Laster, K.V.; Pang, M.; Liu, K.; Hwang, J.; Kim, D.J. Harmaline isolated from Peganum harmala suppresses growth of esophageal squamous cell carcinoma through targeting mTOR. Phyther. Res. 2021, 35, 6377–6388. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhao, S.; Wang, C. Antibacterial, Antifungal, Antiviral, and Antiparasitic Activities of Peganum harmala and Its Ingredients: A Review. Molecules 2022, 27, 4161. [Google Scholar] [CrossRef]
- Li, S.; Cheng, X.; Wang, C. A review on traditional uses, phytochemistry, pharmacology, pharmacokinetics and toxicology of the genus Peganum. J. Ethnopharmacol. 2017, 203, 127–162. [Google Scholar] [CrossRef]
- Khadraoui, N.; Essid, R.; Jallouli, S.; Damergi, B.; Ben Takfa, I.; Abid, G.; Jedidi, I.; Bachali, A.; Ayed, A.; Limam, F.; et al. Antibacterial and antibiofilm activity of Peganum harmala seed extract against multidrug-resistant Pseudomonas aeruginosa pathogenic isolates and molecular mechanism of action. Arch. Microbiol. 2022, 204, 133. [Google Scholar] [CrossRef]
- Abbas, M.W.; Hussain, M.; Qamar, M.; Ali, S.; Shafiq, Z.; Wilairatana, P.; Mubarak, M.S. Antioxidant and Anti-Inflammatory Effects of Peganum harmala Extracts: An In Vitro and In Vivo Study. Molecules 2021, 26, 6084. [Google Scholar] [CrossRef]
- Sarpeleh, A.; Sharifi, K.; Sonbolkar, A. Evidence of antifungal activity of wild rue (Peganum harmala L.) on phytopathogenic fungi. J. Plant Dis. Prot. 2009, 116, 208–213. [Google Scholar] [CrossRef]
- Samaha, A.A.; Fawaz, M.; Salami, A.; Baydoun, S.; Eid, A.H. Antihypertensive Indigenous Lebanese Plants: Ethnopharmacology and a Clinical Trial. Biomolecules 2019, 9, 292. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, X.; Wang, Y.; Li, S.; Zheng, T.; Gao, Y.; Wang, G.; Qi, S.; Wang, J.; Ni, J.; et al. In vivo evaluation of the antitussive, expectorant and bronchodilating effects of extract and fractions from aerial parts of Peganum harmala linn. J. Ethnopharmacol. 2015, 162, 79–86. [Google Scholar] [CrossRef]
- Abd El Baky, H.H.; Ahemd, A.A.; Mekawi, E.M.; Ibrahem, E.A.; Shalapy, N.M. The anti-diabetic and anti-lipidemic effects of Peganum harmala seeds in diabetic rats. Der Pharm. Lett. 2016, 8, 1–10. [Google Scholar]
- Astulla, A.; Zaima, K.; Matsuno, Y.; Hirasawa, Y.; Ekasari, W.; Widyawaruyanti, A.; Zaini, N.C.; Morita, H. Alkaloids from the seeds of Peganum harmala showing antiplasmodial and vasorelaxant activities. J. Nat. Med. 2008, 62, 470–472. [Google Scholar] [CrossRef]
- Nasibova, T.; Garaev, E. Potential anti-Alzheimer alkaloids of Peganum harmala. Alzheimer’s Dement. 2021, 17, e056722. [Google Scholar] [CrossRef]
- Kempster, P.; Ma, A. Parkinson’s disease, dopaminergic drugs and the plant world. Front. Pharmacol. 2022, 13, 3216. [Google Scholar] [CrossRef]
- Malik, S.; Moraes, D.F.C.; do Amaral, F.M.M.; Ribeiro, M.N.S. Ruta graveolens: Phytochemistry, Pharmacology, and Biotechnology. Ref. Ser. Phytochem. 2017, 4, 177–204. [Google Scholar] [CrossRef]
- Pollio, A.; De Natale, A.; Appetiti, E.; Aliotta, G.; Touwaide, A. Continuity and change in the Mediterranean medical tradition: Ruta spp. (rutaceae) in Hippocratic medicine and present practices. J. Ethnopharmacol. 2008, 116, 469–482. [Google Scholar] [CrossRef]
- Jinous Asgarpanah Phytochemistry and pharmacological properties of Ruta graveolens L. J. Med. Plants Res. 2012, 6, 3942–3949. [CrossRef]
- Shikov, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Wagner, H.; Verpoorte, R.; Heinrich, M. Medicinal Plants of the Russian Pharmacopoeia; their history and applications. J. Ethnopharmacol. 2014, 154, 481–536. [Google Scholar] [CrossRef] [PubMed]
- Semerdjieva, I.B.; Burducea, M.; Astatkie, T.; Zheljazkov, V.D.; Dincheva, I. Essential oil composition of ruta graveolens l. fruits and hyssopus officinalis subsp. aristatus (godr.) nyman biomass as a function of hydrodistillation time. Molecules 2019, 24, 4047. [Google Scholar] [CrossRef] [PubMed]
- Kuzovkina, I.; Al’terman, I.; Schneider, B. Specific accumulation and revised structures of acridone alkaloid glucosides in the tips of transformed roots of Ruta graveolens. Phytochemistry 2004, 65, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Kuzovkina, I.N.; Szarka, S.; Héthelyi, É.; Lemberkovics, E.; Szöke, É. Composition of essential oil in genetically transformed roots of Ruta graveolens. Russ. J. Plant Physiol. 2009, 56, 846–851. [Google Scholar] [CrossRef]
- Ali, J.G.; Alborn, H.T.; Stelinski, L.L. Constitutive and induced subterranean plant volatiles attract both entomopathogenic and plant parasitic nematodes. J. Ecol. 2011, 99, 26–35. [Google Scholar] [CrossRef]
- Falkenhagen, H.; Stockigt, J.; Kuzovkina, I.N.; Alterman, I.E.; Kolshorn, H. Indole alkaloids from “hairy roots” of Rauwolfia serpentina. Can. J. Chem. 1993, 71, 2201–2203. [Google Scholar] [CrossRef]
- Lobay, D. Rauwolfia in the Treatment of Hypertension. Integr. Med. A Clin. J. 2015, 14, 40. [Google Scholar]
- Sourabh, P. Ethnomedicinal Uses and Cultivation of Rauvolfia serpentina. Recent Adv. Med. Plants Their Cultiv. 2012, 40, 153–159. [Google Scholar]
- Arjariya, A.; Chaurasia, K. Some Medicinal Plants among the Tribes of Chhatarpur District (M.P.) India. Ecoprint An Int. J. Ecol. 1970, 16, 43–50. [Google Scholar] [CrossRef][Green Version]
- Alshahrani, M.Y.; Rafi, Z.; Alabdallah, N.M.; Shoaib, A.; Ahmad, I.; Asiri, M.; Zaman, G.S.; Wahab, S.; Saeed, M.; Khan, S. A Comparative Antibacterial, Antioxidant, and Antineoplastic Potential of Rauwolfia serpentina (L.) Leaf Extract with Its Biologically Synthesized Gold Nanoparticles (R-AuNPs). Plants 2021, 10, 2278. [Google Scholar] [CrossRef]
- Singh, H.K.; Charan, A.A.; Charan, A.I.; Prasad, S.M. Antifungal and antibacterial activity of methanolic, ethanolic and acetonic leaf extracts of sarpagandha (Rauwolfia serpentina). J. Pharmacogn. Phytochem. 2017, 6, 152–156. [Google Scholar]
- Rao, B.G.; Rao, P.U.; Rao, E.S.; Rao, T.M.; Praneeth, D.V.S. Evaluation of in-vitro antibacterial activity and anti-inflammatory activity for different extracts of Rauvolfia tetraphylla L. root bark. Asian Pac. J. Trop. Biomed. 2012, 2, 818–821. [Google Scholar] [CrossRef]
- Ezeigbo, I.I.; Ezeja, M.I.; Madubuike, K.G.; Ifenkwe, D.C.; Ukweni, I.A.; Udeh, N.E.; Akomas, S.C. Antidiarrhoeal activity of leaf methanolic extract of Rauwolfia serpentina. Asian Pac. J. Trop. Biomed. 2012, 2, 430. [Google Scholar] [CrossRef]
- Weerakoon, S.W.; Arambewela, L.S.R.; Premakumara, G.A.S.; Ratnasooriya, W.D. Sedative activity of the crude extract of Rauvolfia densiflora. Pharm. Biol. 1998, 36, 360–361. [Google Scholar] [CrossRef]
- Gupta, A.K.; Chitme, H.; Dass, S.K.; Misra, N. Hepatoprotective Activity of Rauwolfia serpentina Rhizome in Paracetamol Intoxicated Rats. J. of Pharmacol. Toxicol. 2006, 1, 82–88. [Google Scholar] [CrossRef]
- Kaur, J.; Gulati, S. Therapeutic potential of Rauwolfia serpentina. Indian J. Adv. 2017, 2, 99–104. [Google Scholar]
- Eltamany, E.E.; Nafie, M.S.; Khodeer, D.M.; El-Tanahy, A.H.H.; Abdel-Kader, M.S.; Badr, J.M.; Abdelhameed, R.F.A. Rubia tinctorum root extracts: Chemical profile and management of type II diabetes mellitus. RSC Adv. 2020, 10, 24159–24168. [Google Scholar] [CrossRef]
- Taha, K.; Abu, M. A Natural Anthraquinone Plants with Multi-Pharmacological Activities. Texas J. Med. Sci. 2022; 10, 23–32. [Google Scholar]
- Kalyoncu, F.; Cetin, B.; Saglam, H. Antimicrobial activity of common madder (Rubia tinctorum L.). Phyther. Res. 2006, 20, 490–492. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, J.; Qi, W.; Su, R.; He, Z.; Peng, X. Alizarin and Purpurin from Rubia tinctorum L. Suppress Insulin Fibrillation and Reduce the Amyloid-Induced Cytotoxicity. ACS Chem. Neurosci. 2021, 12, 2182–2193. [Google Scholar] [CrossRef]
- Ahmed, H.E.; Tahoun, I.F.; Elkholy, I.; Shehata, A.B.; Ziddan, Y. Identification of natural dyes in rare Coptic textile using HPLC- DAD and mass spectroscopy in museum of Faculty of Arts, Alexandria University, Egypt. Dye. Pigment. 2017, 145, 486–492. [Google Scholar] [CrossRef]
- Karapanagiotis, I.; Abdel-kareem, O.; Kamaterou, P.; Mantzouris, D. Identification of dyes in coptic textiles from the museum of faculty of archaeology, cairo university. Heritage 2021, 4, 3147–3156. [Google Scholar] [CrossRef]
- Palagina, M.V.; Abramova, G.A. Solodka ural’skaya i ee ispol’zovanie v pishchevoj i farmacevticheskoj promyshlennosti. Nov. v Pishchevyh Tekhnologiyah 2005, 1, 77–87. [Google Scholar]
- Zhu, L.; Xie, S.; Geng, Z.; Yang, X.; Zhang, Q. Evaluating the Potential of Glycyrrhiza uralensis (Licorice) in Treating Alcoholic Liver Injury: A Network Pharmacology and Molecular Docking Analysis Approach. Processes 2022, 10, 1808. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, X.; Feng, Y.; Rui, W.; Shi, Z.; Wu, L. Bioactive components of Glycyrrhiza uralensis mediate drug functions and properties through regulation of CYP450 enzymes. Mol. Med. Rep. 2014, 10, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ye, M. Chemical analysis of the Chinese herbal medicine Gan-Cao (licorice). J. Chromatogr. A 2009, 1216, 1954–1969. [Google Scholar] [CrossRef]
- Deutch, M.R.; Grimm, D.; Wehland, M.; Infanger, M.; Krüger, M. Bioactive Candy: Effects of Licorice on the Cardiovascular System. Foods 2019, 8, 495. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Guseva, A.V.; Kovács, D.; Szöke, É.; Vdovitchenko, M.Y. Flavones in genetically transformed Scutellaria baicalensis roots and induction of their synthesis by elicitation with methyl jasmonate. Russ. J. Plant Physiol. 2005, 52, 77–82. [Google Scholar] [CrossRef]
- Olina, A.V.; Solovyova, A.I.; Solovchenko, A.E.; Orlova, A.V.; Stepanova, A.Y. Physiologically active flavones contentin Scutellaria baicalensis georgiinvitro cultures. Biotekhnologiya 2017, 33, 29–37. [Google Scholar] [CrossRef]
- Stepanova, A.Y.; Solov’eva, A.I.; Malunova, M.V.; Salamaikina, S.A.; Panov, Y.M.; Lelishentsev, A.A. Hairy roots scutellaria spp. (lamiaceae) as promising producers of antiviral flavones. Molecules 2021, 26, 3927. [Google Scholar] [CrossRef]
- Solov’eva, A.I.; Evsyukov, S.V.; Sidorov, R.A.; Stepanova, A.Y. Correlation of endogenous β-glucuronidase activity with differentiation of in vitro cultures of Scutellaria baicalensis. Acta Physiol. Plant. 2020, 42, 106. [Google Scholar] [CrossRef]
- Dikaya, V.S.; Solovyeva, A.I.; Sidorov, R.A.; Solovyev, P.A.; Stepanova, A.Y. The Relationship Between Endogenous β-Glucuronidase Activity and Biologically Active Flavones-Aglycone Contents in Hairy Roots of Baikal Skullcap. Chem. Biodivers. 2018, 15, e1700409. [Google Scholar] [CrossRef]
- Elkin, Y.N.; Kulesh, N.I.; Stepanova, A.Y.; Solovieva, A.I.; Kargin, V.M.; Manyakhin, A.Y. Methylated flavones of the hairy root culture Scutellaria baicalensis. J. Plant Physiol. 2018, 231, 277–280. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Prokof’eva, M.Y.; Umralina, A.R.; Chernysheva, T.P. Morphological and biochemical characteristics of genetically transformed roots of Scutellaria andrachnoides. Russ. J. Plant Physiol. 2014, 61, 697–706. [Google Scholar] [CrossRef]
- Li-Weber, M. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Herrera-Bravo, J.; Salazar, L.A.; Shaheen, S.; Abdulmajid Ayatollahi, S.; Kobarfard, F.; Imran, M.; Imran, A.; Custódio, L.; Dolores López, M.; et al. The Therapeutic Potential of Wogonin Observed in Preclinical Studies. Evid.-Based Complement. Altern. Med. 2021, 2021, 9935451. [Google Scholar] [CrossRef]
- Lin, C.C.; Lin, J.J.; Wu, P.P.; Lu, C.C.; Chiang, J.H.; Kuo, C.L.; Ji, B.C.; Lee, M.H.; Huang, A.C.; Chung, J.G. Wogonin, a natural and biologically-active flavonoid, influences a murine WEHI-3 leukemia model in vivo through enhancing populations of T-And B-cells. Vivo 2013, 27, 733–738. [Google Scholar]
- Qi, Q.; Peng, J.; Liu, W.; You, Q.; Yang, Y.; Lu, N.; Wang, G.; Guo, Q. Toxicological studies of wogonin in experimental animals. Phyther. Res. 2009, 23, 417–422. [Google Scholar] [CrossRef]
- Huang, D.S.; Yu, Y.C.; Wu, C.H.; Lin, J.Y. Protective Effects of Wogonin against Alzheimer’s Disease by Inhibition of Amyloidogenic Pathway. Evid. Based. Complement. Alternat. Med. 2017, 2017, 3545169. [Google Scholar] [CrossRef]
- Tronina, T.; Mrozowska, M.; Bartmańska, A.; Popłoński, J.; Sordon, S.; Huszcza, E. Simple and rapid method for wogonin preparation and its biotransformation. Int. J. Mol. Sci. 2021, 22, 8973. [Google Scholar] [CrossRef]
- Tong, T.; Wu, Y.Q.; Ni, W.J.; Shen, A.Z.; Liu, S. The potential insights of Traditional Chinese Medicine on treatment of COVID-19. Chin. Med. 2020, 15, 51. [Google Scholar] [CrossRef]
- Huang, Y.F.; Bai, C.; He, F.; Xie, Y.; Zhou, H. Review on the potential action mechanisms of Chinese medicines in treating Coronavirus Disease 2019 (COVID-19). Pharmacol. Res. 2020, 158, 104939. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.; Zheng, H.; Soo Park, M.; Eog Ji, G. Optimization of β-Glucuronidase Activity from Lactobacillus delbrueckii Rh2 and Its Use for Biotransformation of Baicalin and Wogonoside. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 275–280. [Google Scholar] [CrossRef]
- Levy, G.A. Baicalinase, a plant beta-glucuronidase. Biochem. J. 1954, 58, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, F.; Matsunae, K.; Hisamitsu, M.; Murakoshi, I.; Kurihara, T.; Yamamoto, T. Purification and properties of a plant beta-D-glucuronidase form Scutellaria root. Biol. Pharm. Bull. 1995, 18, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Shimada, H.; Shimoda, H.; Matsuda, H.; Yamahara, J.; Murakami, N. Rhodiocyanosides A and B, New Antiallergic Cyanoglycosides from Chinese Natural Medicine “Si Lie Hong Jing Tian”, The Underground Part of Rhodiola Quadrifida (Pall.) Fisch. Et Mey. Chem. Pharm. Bull. 1995, 43, 1245–1247. [Google Scholar] [CrossRef][Green Version]
- Yoshikawa, M.; Shimada, H.; Shimoda, H.; Murakami, N.; Yamahara, J.; Matsuda, H. Bioactive constituents of Chinese natural medicines. II. Rhodiolae radix. (1). Chemical structures and antiallergic activity of rhodiocyanosides A and B from the underground part of Rhodiola quadrifida (Pall.) Fisch. et Mey. (Crassulaceae). Chem. Pharm. Bull. 1996, 44, 2086–2091. [Google Scholar] [CrossRef]
- Wiedenfeld, H.; Dumaa, M.; Malinowski, M.; Furmanowa, M.; Narantuya, S. Erratum: Phytochemical and analytical studies of extracts from Rhodiola rosea and Rhodiola quadrifida. Pharmazie 2007, 62, 308–311. [Google Scholar]
- You, J.; Qin, X.; Ranjitkar, S.; Lougheed, S.C.; Wang, M.; Zhou, W.; Ouyang, D.; Zhou, Y.; Xu, J.; Zhang, W.; et al. Response to climate change of montane herbaceous plants in the genus Rhodiola predicted by ecological niche modelling. Sci. Rep. 2018, 8, 5879. [Google Scholar] [CrossRef]
- Stepanova, A.; Malunova, M.; Salamaikina, S.; Selimov, R.; Solov’eva, A. Establishment of Rhodiola quadrifida Hairy Roots and Callus Culture to Produce Bioactive Compounds. Phyton (B. Aires). 2021, 90, 543–552. [Google Scholar] [CrossRef]
- Vdovitchenko, M.Y.; Kuzovkina, I.N.; Paetz, C.; Schneider, B. Formation of phenolic compounds in the roots of Hedysarum theinum cultured in vitro. Russ. J. Plant Physiol. 2007, 54, 536–544. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Guseva, A.V.; Alterman, I.E.; Karnachuk, R.A. Flavonoid production in transformed scutellaria baicalensis roots and ways of its regulation. Russ. J. Plant Physiol. 2001, 48, 448–452. [Google Scholar] [CrossRef]
- Bányai, P.; Kuzovkina, I.N.; Kursinszki, L.; Szoke, É. HPLC analysis of alizarin and purpurin produced by Rubia tinctorum L. hairy root cultures. Chromatographia 2006, 63, S111–S114. [Google Scholar] [CrossRef]
- Baumert, A.; Gröger, D.; Kuzovkina, I.N.; Reisch, J. Secondary metabolites produced by callus cultures of various Ruta species. Plant Cell Tissue Organ Cult. 1992, 28, 159–162. [Google Scholar] [CrossRef]
- Baumert, A.; Kuzovkina, I.N.; Krauss, G.; Hieke, M.; Gröger, D. Biosynthesis of rutacridone in tissue cultures of Ruta graveolens L. Plant Cell Rep. 1982, 1, 168–171. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stepanova, A.Y.; Malunova, M.V.; Gladkov, E.A.; Evsyukov, S.V.; Tereshonok, D.V.; Solov’eva, A.I. Collection of Hairy Roots as a Basis for Fundamental and Applied Research. Molecules 2022, 27, 8040. https://doi.org/10.3390/molecules27228040
Stepanova AY, Malunova MV, Gladkov EA, Evsyukov SV, Tereshonok DV, Solov’eva AI. Collection of Hairy Roots as a Basis for Fundamental and Applied Research. Molecules. 2022; 27(22):8040. https://doi.org/10.3390/molecules27228040
Chicago/Turabian StyleStepanova, Anna Yurievna, Maria Viktorovna Malunova, Evgeny Aleksandrovich Gladkov, Sergey Viktorovich Evsyukov, Dmitry Viktorovich Tereshonok, and Aleksandra Ivanovna Solov’eva. 2022. "Collection of Hairy Roots as a Basis for Fundamental and Applied Research" Molecules 27, no. 22: 8040. https://doi.org/10.3390/molecules27228040
APA StyleStepanova, A. Y., Malunova, M. V., Gladkov, E. A., Evsyukov, S. V., Tereshonok, D. V., & Solov’eva, A. I. (2022). Collection of Hairy Roots as a Basis for Fundamental and Applied Research. Molecules, 27(22), 8040. https://doi.org/10.3390/molecules27228040