
Variation of the Antioxidative Defense in Elaeis guineensis Jacq. Facing Bud Rot Disease in the Coastal Area of Ecuador
 , and
, and 
Abstract
1. Introduction
2. Results
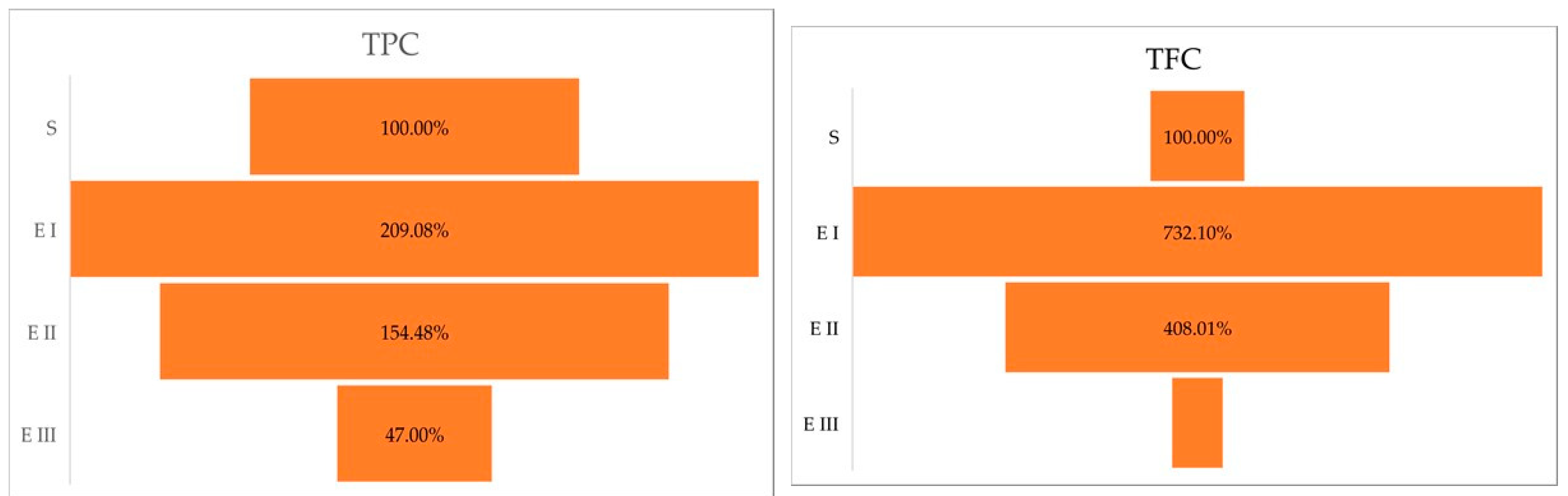
2.1. Total Polyphenolic Content and Total Flavonoid Content
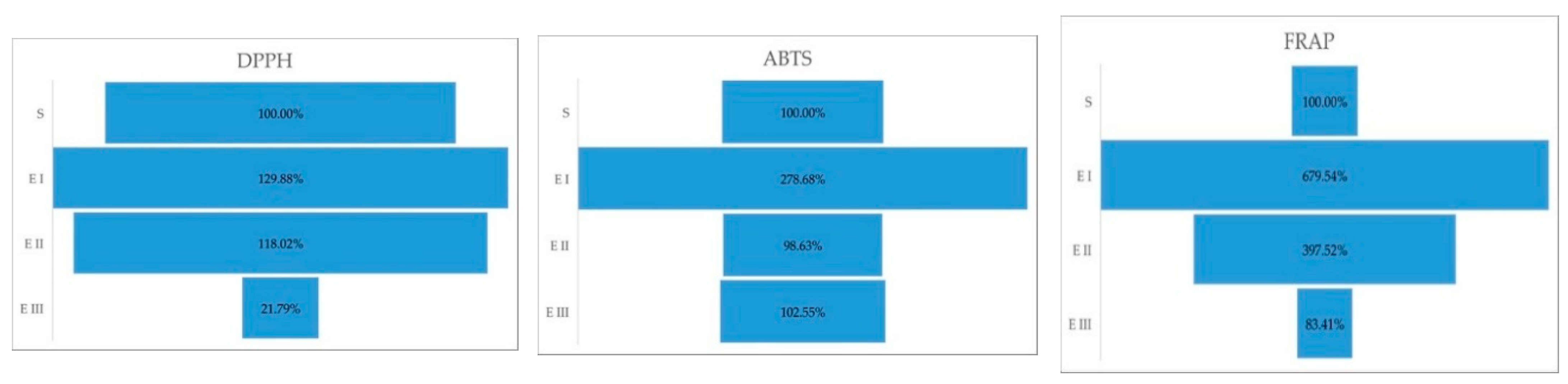
2.2. Antioxidant Activity
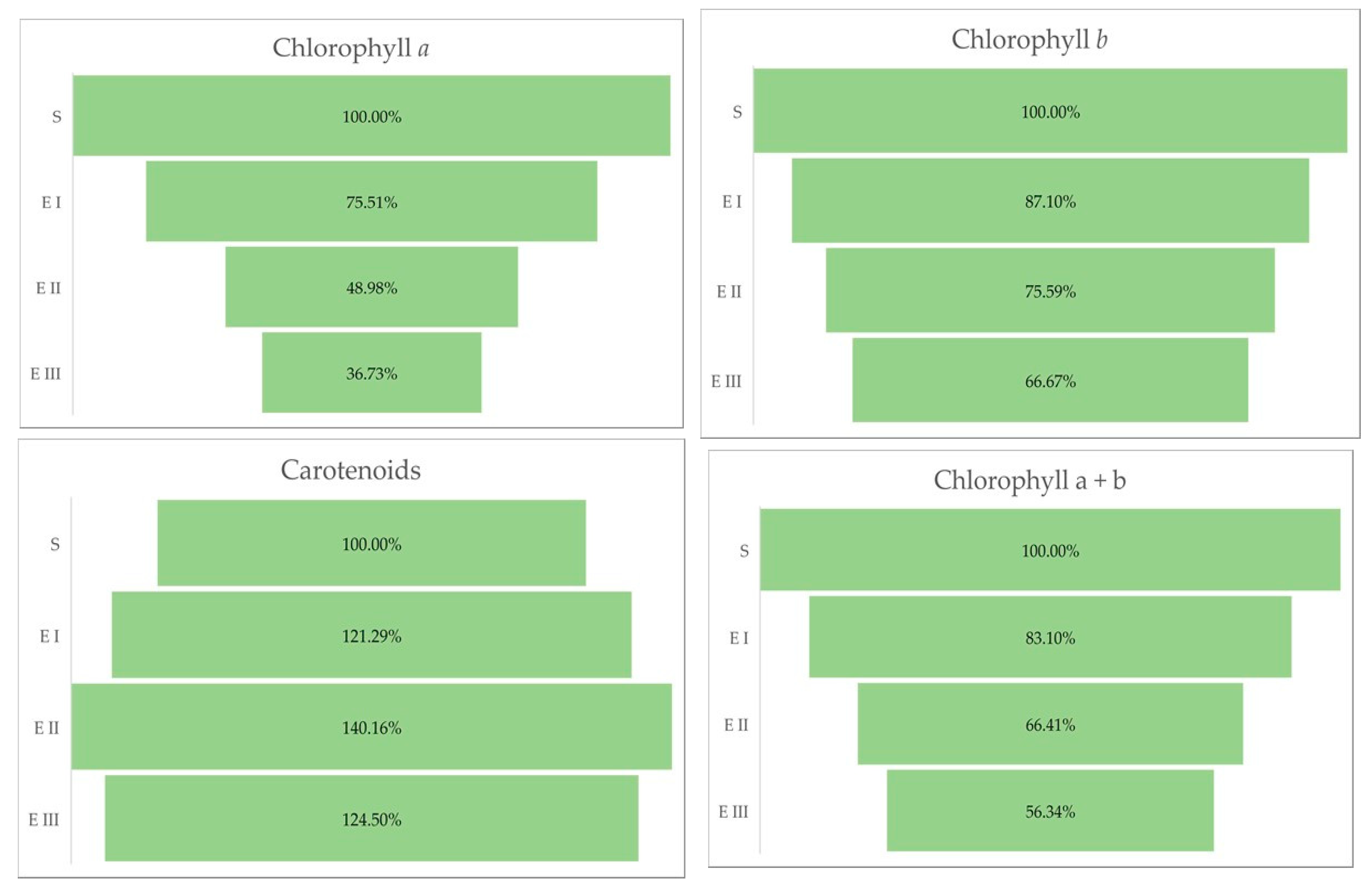
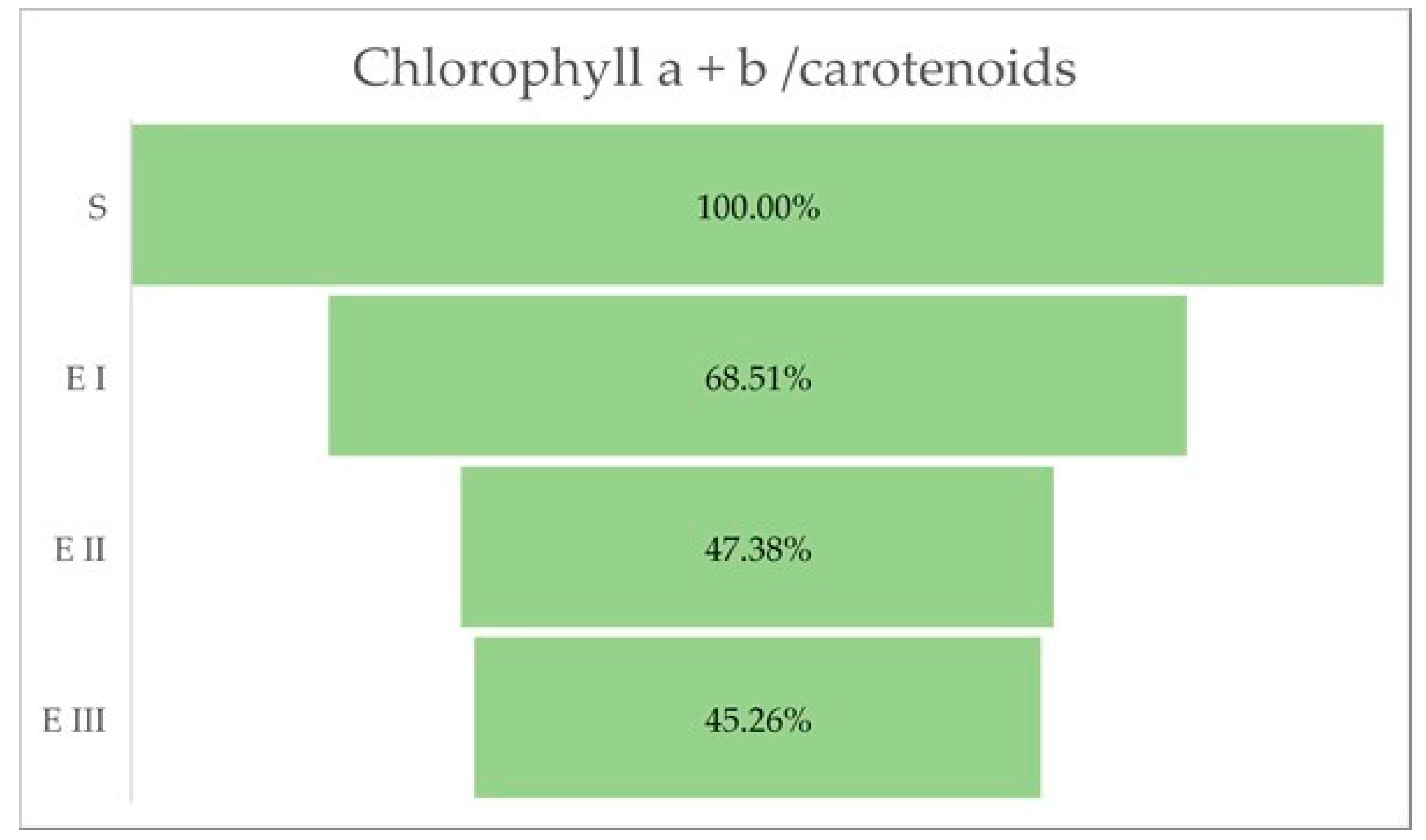
2.3. Chlorophyll a, b and Total Carotenoids Content
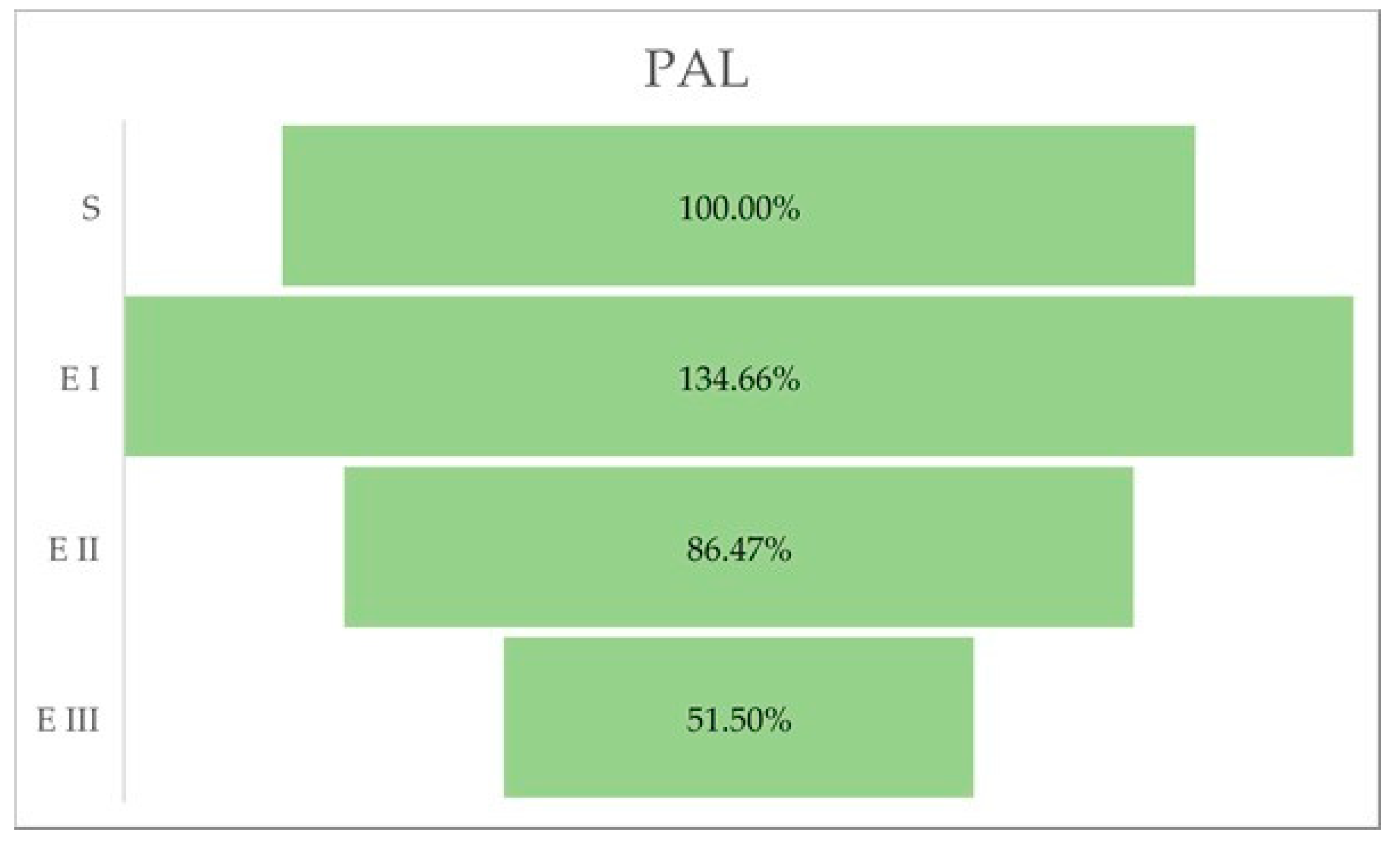
2.4. PAL Activity
3. Discussion
3.1. Total Polyphenol and Total Flavonoid Content Determination
3.2. Antioxidant Activity
3.3. PAL Activity
3.4. Chlorophyll a, b and Total Carotenoids
4. Materials and Methods
4.1. Materials
4.2. Plant Material and Sample Collection
4.3. Palm Extraction
4.4. Determination of the Total Phenolic Content and Total Flavonoid Content
4.5. Antioxidant Activity
4.5.1. DPPH Radical Scavenging Assay
4.5.2. Ferric Reducing Antioxidant Power (FRAP) Assay
4.5.3. ABTS Radical Scavenging Assay
4.6. Quantitative Determination of Chlorophylls a and b and Carotenoids
4.7. Assay of PAL Activity
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Sapey, E.; Adusei-Fosu, K.; Agyei-Dwarko, D.; Okyere-Boateng, G. Collection of Oil Palm (Elaeis guineensis Jacq.) Germplasm in the Northern Regions of Ghana. Asian J. Agric. Sci. 2012, 4, 325–328. [Google Scholar]
- Cosiaux, A.; Gardiner, L.M.; Stauffer, F.W.; Bachman, S.P.; Sonké, B.; Baker, W.J.; Couvreur, T.L. Low extinction risk for an important plant resource: Conservation assessments of continental African palms (Arecaceae/Palmae). Biol. Conserv. 2018, 221, 323–333. [Google Scholar] [CrossRef]
- Murphy, D. Oil palm value chain management. In The Oxford Handbook of Food, Water and Society; Allan, T., Bromwich, B., Keulertz, M., Colman, A., Eds.; Oxford University Press: Oxford, UK, 2019; pp. 630–651. [Google Scholar]
- Viera-Torres, M.; Sinde-González, I.; Gil-Docampo, M.; Bravo-Yandún, V.; Toulkeridis, T. Generating the Baseline in the Early Detection of Bud Rot and Red Ring Disease in Oil Palms by Geospatial Technologies. Remote Sens. 2020, 12, 3229. [Google Scholar] [CrossRef]
- De Franqueville, H. Oil paalm bud rot in latín America. Exp. Agric. 2003, 39, 225–240. [Google Scholar] [CrossRef]
- Benítez, É.; García, C. The history of research on oil palm bud rot (Elaeis guineensis Jacq.) in Colombia. Agron. Colomb. 2014, 32, 390–398. [Google Scholar] [CrossRef]
- Gerardo Martínez, G.L.; Torres, G.A.L. Presence of “Bud Rot Diseasses” in Oil Palm at Nursery Stages. PALMAS 2007, 28, 13–20. [Google Scholar]
- Allen, W.; Paulus, A.; van Gundy, S.; Swinburne, T.R.; Ollagnier, M.; Renard, J.L. Report on the mission to Ecuador. In Document Cirad-Cp; Palmeras del Ecuador-Palmoriente: Montpellier, France, 1995. [Google Scholar]
- Gómez, P.; Ayala, L.; Munévar, F. Characteristics and management of bud rot, a disease of oil palm. In Plantation Tree Crops in the New Millennium: The Way Ahead; Pushparajah, E., Ed.; The Incorporated Society of Planters: Kuala Lumpur, Malaysia, 2000; Volume 1, pp. 545–553. [Google Scholar]
- Sarria, G.A.; Torres, G.A.; Aya, H.A.; Ariza, J.G.; Rodríguez, J.; Vélez, D.C.; Varón, F.; Martínez, G. Phytophthora sp. es el responsable de las lesiones iniciales de la pudrición de cogollo (PC) de la palma de aceite en Colombia. Palmas 2008, 29, 31–41. [Google Scholar]
- Kumar, A.; Verma, J.P. Does plant Microbe interaction confer stress tolerance in plants: A review. Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Singla, P.; Bhardwaj, R.D.; Kaur, S.; Kaur, J. Antioxidant potential of barley genotypes inoculated with five pathotypes of Puccinia striiformis sp. Hordei. Physiol. Mol. Biol. Plants 2019, 25, 145–157. [Google Scholar] [CrossRef]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Diff. 2016, 23, 369–379. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Huang, J.; Waszczak, C.; Jacques, S.; Gevaert, K.; Breusegem, F.V. Cysteines under ROS attack in plants: A proteomics view. J. Exp. Bot. 2015, 66, 2935–2944. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry Advances in Research; Imperato, F., Ed.; Research Signpost: Kollam, India, 2006; pp. 23–67. [Google Scholar]
- Holub, P.; Nezval, J.; Stroch, M.; Spunda, V.; Urban, O.; Jansen, M.A.; Klem, K. Induction of phenolic compounds by UV and PAR is modulated by leaf ontogeny and barley genotype. Plant Physiol. Biochem. 2019, 134, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Cherif, M.; Benhamou, N.; Menzies, J.G.; Belanger, R.R. Silicon-induced cellular defense reactions in cucumber plants attacked with Pythium ultimum. Physiol. Mol. Plant Pathol 1992, 41, 411–425. [Google Scholar] [CrossRef]
- Bhat, S.V.; Nagasampagi, B.A.; Sivakumar, M. Chemistry of Natural Products; Narosa Publishing House: New Delhi, India, 2005. [Google Scholar]
- Davies, K.M.; Albert, N.W.; Schwinn, K.E. From landing lights to mimicry: The molecular regulation of flower coloration and mechanisms for pigmentation patterning. Funct. Plant Biol. 2012, 39, 619–638. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol. 2004, 163, 547–561. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Bhaduri, A.M.; Fulekar, M.H. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Biotechnol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Kumar, S.; Abedin, M.M.; Singh, A.K.; Das, S. Role of Phenolic Compounds in Plant-Defensive Mechanisms. In Plant Phenolics in Sustainable Agriculture; Lone, R., Shuab, R., Kamili, A., Eds.; Springer: Singapore, 2020. [Google Scholar]
- Shoib, A.B.; Shahid, A.M. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar]
- Bhattacharjee, P.; Sarkar, J.; Chakraborty, A.P.; Chakraborty, U. Biochemical responses in Sorghum bicolor and Triticum aestivum to spot blotch disease and induction of resistance by plant growth promoting rhizobacteria. J. Mycol. Plant Pathol. 2017, 47, 323–332. [Google Scholar]
- You, X.; Fang, H.; Wang, R.; Wang, G.L.; Ning, Y. Phenylalanine ammonia lyases mediate broad-spectrum resistance to pathogens and insect pests in plants. Sci. Bull. 2020, 65, 1425–1427. [Google Scholar] [CrossRef]
- Kaur, S.; Bhardwaj, R.D.; Kaur, J.; Kaur, S. Induction of defense-related enzymes and pathogenesis-related proteins imparts resistance to barley genotypes against spot blotch disease. J. Plant Growth Regul. 2022, 41, 682–696. [Google Scholar] [CrossRef]
- Jun, S.Y.; Sattler, S.A.; Cortez, G.S.; Vermerris, W.; Sattler, S.E.; Kang, C. Biochemical and structural analysis of substrate specificity of a Phenylalanine Ammonia-Lyase. Plant Physiol. 2018, 176, 1452–1468. [Google Scholar] [CrossRef]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Gogoi, R.; Singh, D.V.; Srivastava, K.D. Phenols as a biochemical basis of resistance in wheat against Karnal bunt. Plant Pathol. 2001, 50, 470–476. [Google Scholar] [CrossRef]
- Mishra, V.K.; Biswas, S.K.; Rajik, M. Biochemical mechanism of resistance to Alternaria blight by different varieties of wheat. Int. J. Plant Pathol. 2011, 2, 72–80. [Google Scholar] [CrossRef][Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Paznocht, L.; Kotíková, Z.; Šulc, M.; Lachman, J.; Orsák, M.; Eliášová, M.; Martinek, P. Free and esterified carotenoids in pigmented wheat, tritordeum and barley grains. Food Chem. 2018, 240, 670–678. [Google Scholar] [CrossRef]
- Young, A.; Lowe, G. Carotenoids—Antioxidant Properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef]
- Serrano, I.; Audran, C.; Rivas, S. Chloroplasts at work during plant innate immunity. J. Exp. Bot. 2016, 67, 3845–3854. [Google Scholar] [CrossRef] [PubMed]
- Hafiza, N.; Nabil, S.; Muhammad, F.; McCullagh, J.S.O. Measurement of total phenolic content and antioxidant activity of aerial parts of medicinal plant Coronopus didymus. Asian Pac. J. Trop. Med. 2017, 10, 792–801. [Google Scholar]
- Arvouet-Grand, A.; Vennat, B.; Pourrat, A.; Legret, P. Standardization of propolis extract and identification of principal constituents. J. Pharm. Belg. 1994, 49, 462–468. [Google Scholar] [PubMed]
- Simirgiotis, M.J.; Schmeda-Hirschmann, G. Determination of phenolic composition and antioxidant activity in fruits, rhizomes and leaves of the white strawberry (Fragaria chiloensis spp. chiloensis form chiloensis) using HPLC-DAD-ESI-MS and free radical quenching techniques. J. Food Compos. Anal. 2010, 23, 545–553. [Google Scholar]
- Loizzo, M.R.; Pacetti, D.; Lucci, P.; Núñez, O.; Menichini, F.; Frega, N.G.; Tundis, R. Prunus persica var. platycarpa (Tabacchiera Peach): Bioactive compounds and antioxidant activity of pulp, peel and seed ethanolic extracts. Plant Foods Hum. Nutr. 2015, 70, 331–337. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Wellburn, A. Determination of total carotenoids and chlorophylls A and B of leaf in different solvents. Biochem. SocTrans 1985, 11, 591–592. [Google Scholar] [CrossRef]
- Lister, C.E.; Lancaster, J.E. Phenylalanine Ammonia-lyase (PAL) Activity and its Relationship to Anthocyanin and Flavonoid Levels in New Zealand-grown Apple Cultivars. J. Am. Soc. Hortic. Sci. 1996, 121, 281–285. [Google Scholar] [CrossRef]
- Data Analysis and Statistical Solution for Microsoft Excel. In XLSTAT Pro; Addinsoft: Paris, France, 2013.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPC (mg GAE/g DW) | TFC (mg QE/g DW) | |
|---|---|---|
| S | 21.619 ± 3.721 a | 0.178 ± 0.032 a |
| E I | 45.203 ± 1.162 b | 1.310 ± 0.108 b |
| E II | 33.397 ± 0.932 c | 0.730 ± 0.062 c |
| E III | 10.161 ± 0.628 d | 0.096 ± 0.018 a |
| DPPH | ABTS | FRAP | |
|---|---|---|---|
| S | 58.5 ± 0.14 a | 15.28 ± 0.056 a | 36.44 ± 0.02 a |
| E I | 75.981 ± 0.003 b | 42.582 ± 0.0085 b | 247.64 ± 0.03 b |
| E II | 69.042 ± 0.001 c | 15.07 ± 0.019 a | 144.86 ± 0.06 c |
| E III | 12.747 ± 0.01 d | 5.623 ± 0.007 c | 30.39 ± 0.0001 a |
| Chlorophyll a (mg/g DW) | Chlorophyll b (mg/g DW) | Carotenoids (mg/g DW) | Chlorophyll a + b | Chlorophyll a + b/Carotenoids | |
|---|---|---|---|---|---|
| S | 4.9 ± 0.01 | 9.3 ± 0.128 | 2.49 ± 0.204 | 14.2 ± 0.125 | 5.703 ± 0.441 |
| E I | 3.7 ± 0.04 | 8.1 ± 0.115 | 3.02 ± 0.231 | 11.8 ± 0.147 | 3.907 ± 0.279 |
| E II | 2.4 ± 0.08 | 7.03 ± 0.026 | 3.49 ± 0.304 | 9.43 ± 0.079 | 2.702 ± 0.217 |
| E III | 1.8 ± 0.111 | 6.2 ± 0.815 | 3.1 ± 0.096 | 8 ± 0.139 | 2.581 ± 0.113 |
| PAL-Specific Activity (U/mg Protein) | |
|---|---|
| S | 1315.694 ± 71.108 |
| E I | 1771.667 ± 33.868 |
| E II | 1137.639 ± 57.010 |
| E III | 677.6389 ± 187.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihai, R.A.; Guacollantes, G.M.C.; Villacrés Mesias, S.A.; Florescu, L.I.; Catana, R.D. Variation of the Antioxidative Defense in Elaeis guineensis Jacq. Facing Bud Rot Disease in the Coastal Area of Ecuador. Molecules 2022, 27, 7314. https://doi.org/10.3390/molecules27217314
Mihai RA, Guacollantes GMC, Villacrés Mesias SA, Florescu LI, Catana RD. Variation of the Antioxidative Defense in Elaeis guineensis Jacq. Facing Bud Rot Disease in the Coastal Area of Ecuador. Molecules. 2022; 27(21):7314. https://doi.org/10.3390/molecules27217314
Chicago/Turabian StyleMihai, Raluca A., Galo M. Canchignia Guacollantes, Sebastián A. Villacrés Mesias, Larisa I. Florescu, and Rodica D. Catana. 2022. "Variation of the Antioxidative Defense in Elaeis guineensis Jacq. Facing Bud Rot Disease in the Coastal Area of Ecuador" Molecules 27, no. 21: 7314. https://doi.org/10.3390/molecules27217314
APA StyleMihai, R. A., Guacollantes, G. M. C., Villacrés Mesias, S. A., Florescu, L. I., & Catana, R. D. (2022). Variation of the Antioxidative Defense in Elaeis guineensis Jacq. Facing Bud Rot Disease in the Coastal Area of Ecuador. Molecules, 27(21), 7314. https://doi.org/10.3390/molecules27217314