
Bioaccessibility and Oxidative Stability of Omega-3 Fatty Acids in Supplements, Sardines and Enriched Eggs Studied Using a Static In Vitro Gastrointestinal Model



Abstract
:1. Introduction
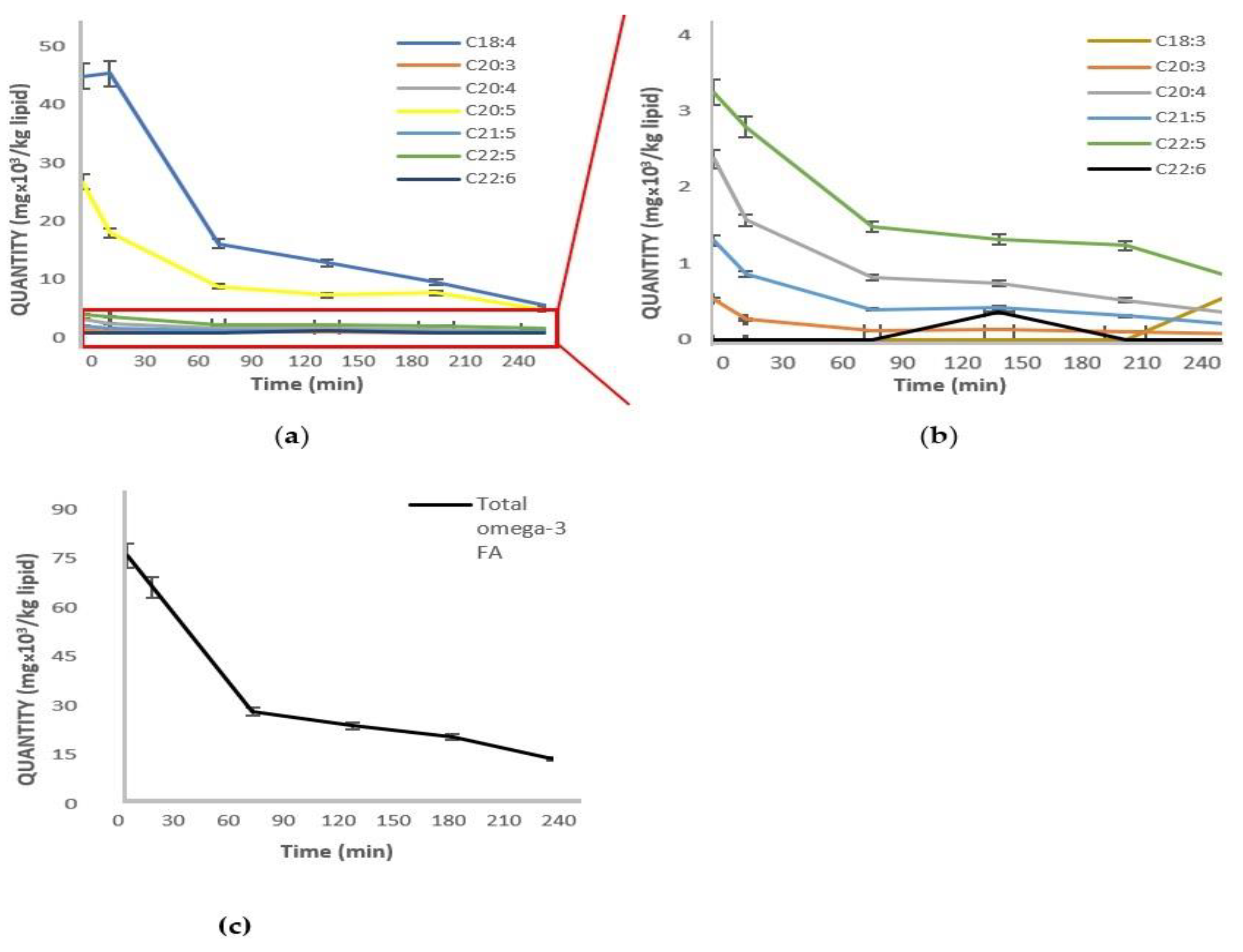
2. Results
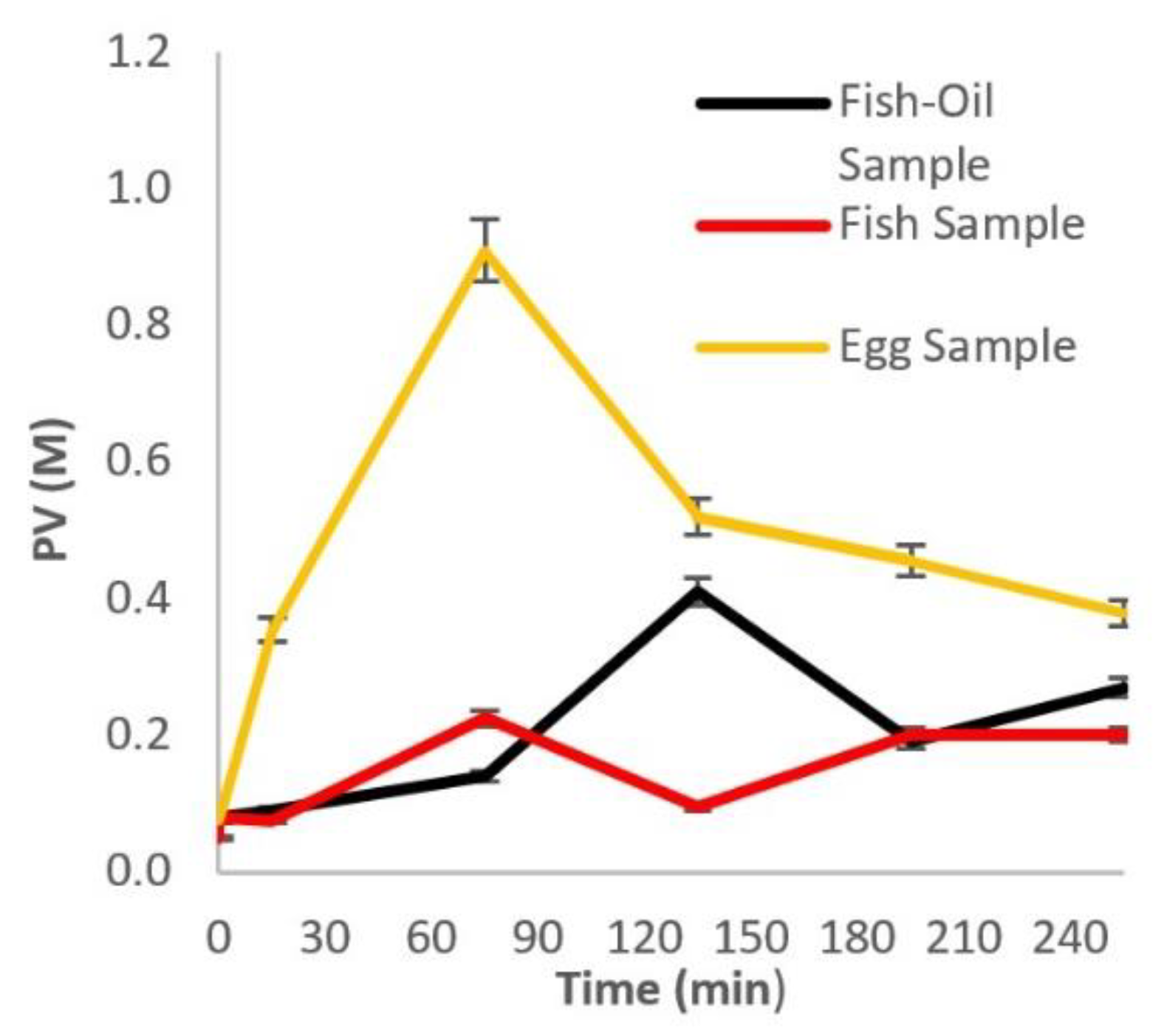
2.1. Primary Oxidation—PV Determination
2.2. Secondary Oxidation—TBARS Determination
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sample Preparation
4.2.1. Enzyme Activity Assays
4.2.2. Stock Solution Preparation
4.3. Static In Vitro Simulation of Gastrointestinal (GI) Digestion
4.3.1. Oral Digestion Phase
4.3.2. Gastric Digestion Phase
4.3.3. Intestinal Digestion Phase
4.4. Sample Treatment and Storage
4.5. Sample Analysis
4.5.1. Peroxide Value (PV)
4.5.2. Thiobarbituric Acid (TBARS) Method
4.5.3. Gas Chromatography—Flame Ionization Detector (GC-FID)
4.6. Bioaccessibility Index (BI)
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Higdon, J. Essential Fatty Acids, Micronutrient Information Center 179. 2019. Available online: https://lpi.oregonstate.edu/mic/other-nutrients/essential-fatty-acids (accessed on 20 September 2021).
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Ahmed, M.; Pickova, J.; Ahmad, T.; Liaquat, M.; Farid, A.; Jahangir, M. Oxidation of Lipids in Foods. Sarhad J. Agric. 2016, 32, 230–238. [Google Scholar] [CrossRef]
- Larsson, K. Oxidation of Fish Lipids During Gastrointestinal In Vitro Digestion; Department of Biology and Biological Engineering, Chalmers University of Technology: Gothenburg, Sweden, 2016. [Google Scholar]
- Helbig, A.; Digestion of Dietary Fat. Gastrointestinal Behaviour of Emulsions and Human Physiological Responses. 2013. Available online: https://www.wur.nl/nl/show/Anna-Helbig-Digestion-of-dietary-fat-Gastrointestinal-behaviour-of-emulsions-and-human-physiological-responses (accessed on 22 September 2021).
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef] [Green Version]
- Minich, D.M.; Vonk, R.J.; Verkade, H.J. Intestinal absorption of essential fatty acids under physiological and essential fatty acid-deficient conditions. J. Lipid Res. 1997, 38, 1709–1721. [Google Scholar] [CrossRef]
- Parada, J.; Aguilera, J.M. Food Microstructure Affects the Bioavailability of Several Nutrients. J. Food Sci. 2007, 72, R21–R32. [Google Scholar] [CrossRef]
- Galanakis, C.M. Bioavailability, bioaccessibility and bioactivity of food components. In Nutraceutical and Functional Food Components: Effects of Innovative Processing Techniques; Academic Press: Cambridge, MA, USA, 2017; Available online: https://www.sciencedirect.com/science/article/pii/B9780128052570000016 (accessed on 15 June 2021).
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Echevarria, B.N.; Goicoechea, E.; Guillén, M.D. Food lipid oxidation under gastrointestinal digestion conditions: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 461–478. [Google Scholar] [CrossRef]
- Calvo-Lerma, J.; Fornés-Ferrer, V.; Heredia, A.; Andrés, A. In Vitro Digestion of Lipids in Real Foods: Influence of Lipid Organization within the Food Matrix and Interactions with Nonlipid Components. J. Food Sci. 2018, 83, 2629–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClements, D.J.; Decker, E.A.; Park, Y. Controlling Lipid Bioavailability through Physicochemical and Structural Approaches. Crit. Rev. Food Sci. Nutr. 2008, 49, 48–67. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.L.; Hoy, C.E. The digestion of dietary triacylglycerols. Prog. Lipid Res. 2004, 43, 105–133. [Google Scholar] [CrossRef]
- Armand, M. Lipases and lipolysis in the human digestive tract: Where do we stand? Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 156–164. [Google Scholar] [CrossRef]
- Tullberg, C.; Vegarud, G.; Undeland, I. Oxidation of marine oils during in vitro gastrointestinal digestion with human digestive fluids—Role of oil origin, added tocopherols and lipolytic activity. Food Chem. 2019, 270, 527–537. [Google Scholar] [CrossRef]
- Larsson, K.; Cavonius, L.; Alminger, M.; Undeland, I. Oxidation of Cod Liver Oil during Gastrointestinal In Vitro Digestion. J. Agric. Food Chem. 2012, 60, 7556–7564. [Google Scholar] [CrossRef] [PubMed]
- Grotto, D.; Maria, L.S.; Valentini, J.; Paniz, C.; Schmitt, G.; Garcia, S.; Pomblum, V.J.; da Rocha, J.B.T.; Farina, M. Importance of the lipid peroxidation biomarkers and methodological aspects FOR malondialdehyde quantification. Química Nova 2009, 32, 169–174. [Google Scholar] [CrossRef] [Green Version]
- De Leonardis, A.; Macciola, V. A study on the lipid fraction of Adriatic sardine filets (Sardina pilchardus). Food/Nahrung 2004, 48, 209–212. [Google Scholar] [CrossRef]
- Davey, M.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Vellido-Perez, J.A.; Ochando-Pulido, J.M.; Brito-de la Fuente, E.; Martinez-Ferez, A. Novel emulsions–based technological approaches for the protection of omega–3 polyunsaturated fatty acids against oxidation processes—A comprehensive review. Food Struct. 2021, 27, 100175. [Google Scholar] [CrossRef]
- Aguilera, J.M. The food matrix: Implications in processing, nutrition and health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.S.; Galano, J.-M.; Durand, T.; Lee, J.C.-Y. Profiling of Omega-Polyunsaturated Fatty Acids and Their Oxidized Products in Salmon after Different Cooking Methods. Antioxidants 2018, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.; Decker, E.A.; McClements, D.J. Development of food-grade nanoemulsions and emulsions for delivery of omega-3 fatty acids: Opportunities and obstacles in the food industry. Food Funct. 2015, 6, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Čepo, D.V.; Radić, K.; Turčić, P.; Anić, D.; Komar, B.; Šalov, M. Food (Matrix) Effects on Bioaccessibility and Intestinal Permeability of Major Olive Antioxidants. Foods 2020, 9, 1831. [Google Scholar] [CrossRef]
- Ismail, A.; Bannenberg, G.; Rice, H.B.; Schutt, E.; MacKay, D. Oxidation in EPA- and DHA-rich oils: An overview. Lipid Technol. 2016, 28, 55–59. [Google Scholar] [CrossRef]
- Capuano, E. The behavior of dietary fiber in the gastrointestinal tract determines its physiological effect. Crit. Rev. Food Sci. Nutr. 2017, 57, 3543–3564. [Google Scholar] [CrossRef] [Green Version]
- Lavado, G.; Higuero, N.; León-Camacho, M.; Cava, R. Formation of Lipid and Protein Oxidation Products during In Vitro Gastrointestinal Digestion of Dry-Cured Loins with Different Contents of Nitrate/Nitrite Added. Foods 2021, 10, 1748. [Google Scholar] [CrossRef]
- Bochkov, V.N.; Oskolkova, O.V.; Birukov, K.G.; Levonen, A.-L.; Binder, C.J.; Stöckl, J. Generation and Biological Activities of Oxidized Phospholipids. Antioxid. Redox Signal. 2010, 12, 1009–1059. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Sun, J.; Liu, Y.; Zeng, H.; Su, Y.; Yang, Y. ACE inhibitory peptides and antioxidant peptides derived from in vitro digestion hydrolysate of hen egg white lysozyme. Food Chem. 2012, 135, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Nau, F.; Pasco, M.; Mine, Y. Identification of Hen Egg Yolk-Derived Phosvitin Phosphopeptides and Their Effects on Gene Expression Profiling against Oxidative Stress-Induced Caco-2 Cells. J. Agric. Food Chem. 2011, 59, 9207–9218. [Google Scholar] [CrossRef] [PubMed]
- Remanan, M.K.; Wu, J. Antioxidant activity in cooked and simulated digested eggs. Food Funct. 2014, 5, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Nimalaratne, C.; Savard, P.; Gauthier, S.F.; Schieber, A.; Wu, J. Bioaccessibility and Digestive Stability of Carotenoids in Cooked Eggs Studied Using a Dynamic In Vitro Gastrointestinal Model. J. Agric. Food Chem. 2015, 63, 2956–2962. [Google Scholar] [CrossRef] [PubMed]
- Nolasco, E.; Yang, J.; Ciftci, O.; Vu, D.C.; Alvarez, S.; Purdum, S.; Majumder, K. Evaluating the effect of cooking and gastrointestinal digestion in modulating the bio-accessibility of different bioactive compounds of eggs. Food Chem. 2021, 344, 128623. [Google Scholar] [CrossRef]
- Fraeye, I.; Bruneel, C.; Lemahieu, C.; Buyse, J.; Muylaert, K.; Foubert, I. Dietary enrichment of eggs with omega-3 fatty acids: A review. Food Res. Int. 2012, 48, 961–969. [Google Scholar] [CrossRef]
- Venugopalan, V.; Gopakumar, L.; Kumaran, A.; Chatterjee, N.; Soman, V.; Peeralil, S.; Mathew, S.; McClements, D.; Nagarajarao, R. Encapsulation and Protection of Omega-3-Rich Fish Oils Using Food-Grade Delivery Systems. Foods 2021, 10, 1566. [Google Scholar] [CrossRef]
- Chang, H.W.; Tan, T.B.; Tan, P.Y.; Abas, F.; Lai, O.M.; Wang, Y.; Wang, Y.; Nehdi, I.A.; Tan, C.P. Microencapsulation of fish oil using thiol-modified β-lactoglobulin fibrils/chitosan complex: A study on the storage stability and in vitro release. Food Hydrocoll. 2018, 80, 186–194. [Google Scholar] [CrossRef]
- Waraho, T.; McClements, D.; Decker, E.A. Impact of free fatty acid concentration and structure on lipid oxidation in oil-in-water emulsions. Food Chem. 2011, 129, 854–859. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Lipid oxidation and improving the oxidative stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Domingos, A.K.; Saad, E.B.; Vechiatto, W.W.D.; Wilhelm, H.M.; Ramos, L.P. The influence of BHA, BHT and TBHQ on the oxidation stability of soybean oil ethyl esters (biodiesel). J. Braz. Chem. Soc. 2007, 18, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Richards, M.P.; Hultin, H.O. Effect of pH on Lipid Oxidation Using Trout Hemolysate as a Catalyst: A Possible Role for Deoxyhemoglobin. J. Agric. Food Chem. 2000, 48, 3141–3147. [Google Scholar] [CrossRef]
- Christie, W.; Han, X. Lipid Analysis, 5th ed.; Woodhead: Oxford, UK, 2012; pp. 181–211. [Google Scholar]
- Bakota, E. Peroxide Value Method. 2014. Available online: https://www.protocols.io/view/Peroxide-Value-Method-kiuujd (accessed on 17 April 2021).
- Shantha, N.C.; Decker, E. Rapid, Sensitive, Iron-Based Spectrophotometric Methods for Determination of Peroxide Values of Food Lipids. J. AOAC Int. 1994, 77, 421–424. [Google Scholar] [CrossRef]
- Lemons, D.W. Fisheries and Marine Service. 1975. Available online: http://icnaf.nafo.int/docs/1974/res-07.pdf (accessed on 10 March 2021).
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, S.; Huang, Y.; Huang, J.; Li, Y. Effect of in vitro gastrointestinal digestion on phenolic compounds and antioxidant properties of soluble and insoluble dietary fibers derived from hulless barley. J. Food Sci. 2021, 86, 628–634. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Peroxide Concentration (M) | ||||
|---|---|---|---|---|
| Sample | Raw State (Before GI Tract) | Heat Treated State (Before GI Tract) | Digested State (After GI Tract) | Increase (%) |
| Fish Oil Supplement: Brand A (n = 3) | 0.013 ± 0.006 a | N/A | 0.093 ± 0.045 b | 615 |
| Fish Oil Supplement: Brand B (n = 3) | 0.026 ± 0.011 a | N/A | 0.140 ± 0.026 b | 438 |
| Fish Oil Supplement: Brand C (n = 3) | 0.056 ± 0.011 a | N/A | 0.193 ± 0.025 b | 245 |
| Fish Oil Supplement: Brand D (n = 3) | 0.083 ± 0.005 a | N/A | 0.273 ±0.015 b | 229 |
| Sardine (n = 9) | 0.026 ± 0.005 a | 0.035 ± 0.006 a | 0.190 ± 0.010 b | 630 |
| Egg (n = 9) | 0.141 ± 0.027 a | 0.144 ± 0.053 a | 0.280 ± 0.077 b | 98 |
| MDAeq Concentration (M × 10−6) | ||||
|---|---|---|---|---|
| Sample | Raw State (Before GI Tract) | Heat Treated State (Before GI Tract) | Digested State (After GI Tract) | Increase (%) |
| Fish Oil Supplement: Brand A (n = 3) | 13.46 ± 2.92 a | N/A | 189.72 ± 99.84 b | 1310 |
| Fish Oil Supplement: Brand B (n = 3) | 7.14 ± 0.69 a | N/A | 147.65 ± 17.49 b | 1968 |
| Fish Oil Supplement: Brand C (n = 3) | 15.80 ± 9.76 a | N/A | 385.14 ± 36.50 b | 2338 |
| Fish Oil Supplement: Brand D (n = 5) | 13.05 ± 3.60 a | N/A | 232.29 ± 52.70 b | 1680 |
| Sardine (n = 9) | 64.09 ± 3.71 a | 48.27 ± 8.80 a | 196.99 ± 37.02 b | 207 |
| Egg (n = 9) | 6.13 ± 1.32 a | 6.17 ± 1.37 a | 19.75 ± 5.68 b | 222 |
| Fish Oil Supplement, Brand A | Fish Oil Supplement, Brand B | Fish Oil Supplement, Brand C | |||||||||
| PUFA | Ca (mg/kg Lipid) | Cb (mg/kg Lipid) | BI (%) | PUFA | Ca (mg/kg Lipid) | Cb (mg/kg Lipid) | BI (%) | PUFA | Ca (mg/kg Lipid) | Cb (mg/kg Lipid) | BI (%) |
| C22:6 n-3 (DHA) | 102,880 | 31,193 | 30.3 | C22:6 n-3 (DHA) | 269,396 | 185,991 | 69.0 | C22:6 n-3 (DHA) | 142,982 | 46,858 | 32.6 |
| C20:5 n-3 (EPA) | 137,695 | 49,958 | 36.3 | C20:5 n-3 (EPA) | 58,620 | 43,965 | 75.0 | C20:5 n-3 (EPA) | 197,368 | 61,695 | 31.0 |
| C20:3 n-3 | 1234 | 0 | 0 | C20:3 n-3 | 1939 | 0 | 0 | C20:3 n-3 | 20,833 | 0 | 0 |
| C18:3 n-3 | 1939 | 4669 | Ν/A | N/A | |||||||
| TOTAL (n-3) | 241,809 | 81,151 | 33.5 | TOTAL (n-3) | 329,955 | 229,956 | 69.0 | TOTAL (n-3) | 361,183 | 108,553 | 30.0 |
| Fish Oil Supplement, Brand D | Fish | Egg | |||||||||
| PUFA | Ca (mg/kg Lipid) | Cb (mg/kg Lipid) | BI (%) | PUFA | Ca (mg/kg Lipid) | Cb (mg/kg Lipid) | BI (%) | PUFA | Ca (mg/kg Lipid) | Cb (mg/kg Lipid) | BI (%) |
| C22:5 n-3 (DPA) | 3089 | 708 | 22.9 | C22:6 n-3 (DHA) | 389 | 93 | 23.9 | C22:6 n-3 (DHA) | 5102 | 0 | 0 |
| C21:5 n-3 | 1242 | 175 | 14.1 | C20:5 n-3 (EPA) | 254 | 106 | 41.7 | C22:5 n-3 | 100 | 0 | 0 |
| C20:5 n-3 (EPA) | 24,991 | 3817 | 15.3 | C18:3 n-3 | 254 | 40 | 15.7 | C21:5 n-3 | 171 | 0 | 0 |
| C20:4 n-3 | 2257 | 296 | 13.1 | C18:4 n-3 | 0 | 240 | N/A | C18:4 n-3 | 211 | 0 | 0 |
| C20:3 n-3 | 506 | 71 | 14.4 | C18:3 n-3 | 21 | 579 | Ν/A | ||||
| C18:4 n-3 | 42,269 | 4523 | 10.7 | ||||||||
| C18:3 n-3 | 0 | 669 | N/A | ||||||||
| TOTAL (n-3) | 74,354 | 9590 | 12.9 | TOTAL (n-3) | 897 | 239 | 26.6 | TOTAL (n-3) | 5605 | 0 | 0 |
| Fatty Acid Concentration | |||
|---|---|---|---|
| Fatty Acid | Ca (ppm) | C75 (ppm) | Rate |
| C18:4 n-3 | 42,269 | 14,693 | −368 |
| C20:5 n-3 (EPA) | 24,991 | 7700 | −231 |
| C22:5 n-3 (DPA) | 3089 | 1412 | −22 |
| C20:4 n-3 | 2257 | 779 | −20 |
| C21:5 n-3 | 1242 | 379 | −12 |
| C20:3 n-3 | 506 | 118 | −5 |
| r = −0.9993 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floros, S.; Toskas, A.; Pasidi, E.; Vareltzis, P. Bioaccessibility and Oxidative Stability of Omega-3 Fatty Acids in Supplements, Sardines and Enriched Eggs Studied Using a Static In Vitro Gastrointestinal Model. Molecules 2022, 27, 415. https://doi.org/10.3390/molecules27020415
Floros S, Toskas A, Pasidi E, Vareltzis P. Bioaccessibility and Oxidative Stability of Omega-3 Fatty Acids in Supplements, Sardines and Enriched Eggs Studied Using a Static In Vitro Gastrointestinal Model. Molecules. 2022; 27(2):415. https://doi.org/10.3390/molecules27020415
Chicago/Turabian StyleFloros, Stylianos, Alexandros Toskas, Evagelia Pasidi, and Patroklos Vareltzis. 2022. "Bioaccessibility and Oxidative Stability of Omega-3 Fatty Acids in Supplements, Sardines and Enriched Eggs Studied Using a Static In Vitro Gastrointestinal Model" Molecules 27, no. 2: 415. https://doi.org/10.3390/molecules27020415
APA StyleFloros, S., Toskas, A., Pasidi, E., & Vareltzis, P. (2022). Bioaccessibility and Oxidative Stability of Omega-3 Fatty Acids in Supplements, Sardines and Enriched Eggs Studied Using a Static In Vitro Gastrointestinal Model. Molecules, 27(2), 415. https://doi.org/10.3390/molecules27020415