
Grape Polyphenols in the Treatment of Human Skeletal Muscle Damage Due to Inflammation and Oxidative Stress during Obesity and Aging: Early Outcomes and Promises





Abstract
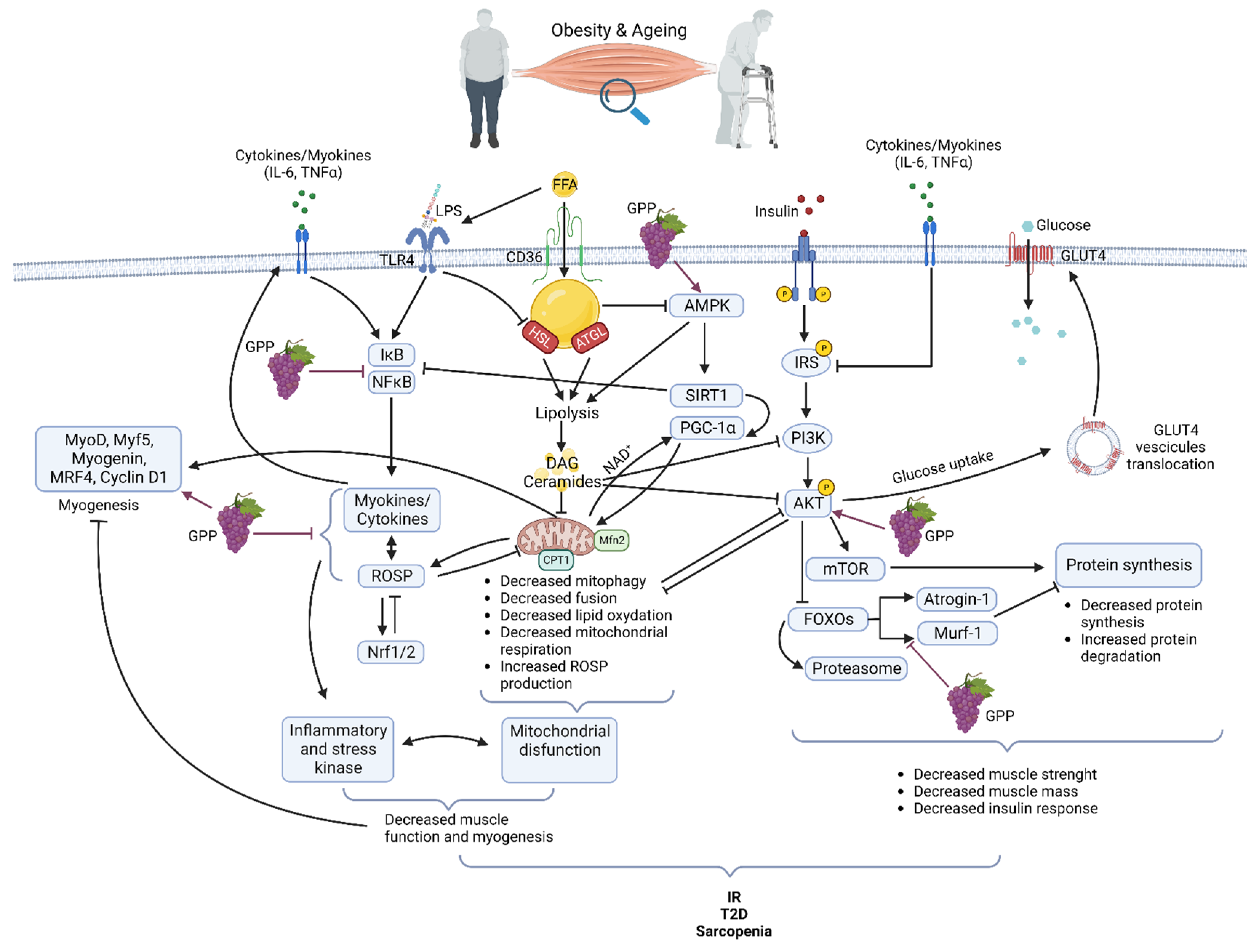
1. Obesity and Aging: Two Major Healthcare Challenges to Solve
1.1. Skeletal Muscle Alterations Are Central
1.2. Muscle Alterations
1.2.1. Muscle Alterations in Obesity
1.2.2. Muscle Alterations in Aging
2. Oxidative Stress and Inflammation: Two Essential Harms
3. Grape Polyphenols: An Effective Tool
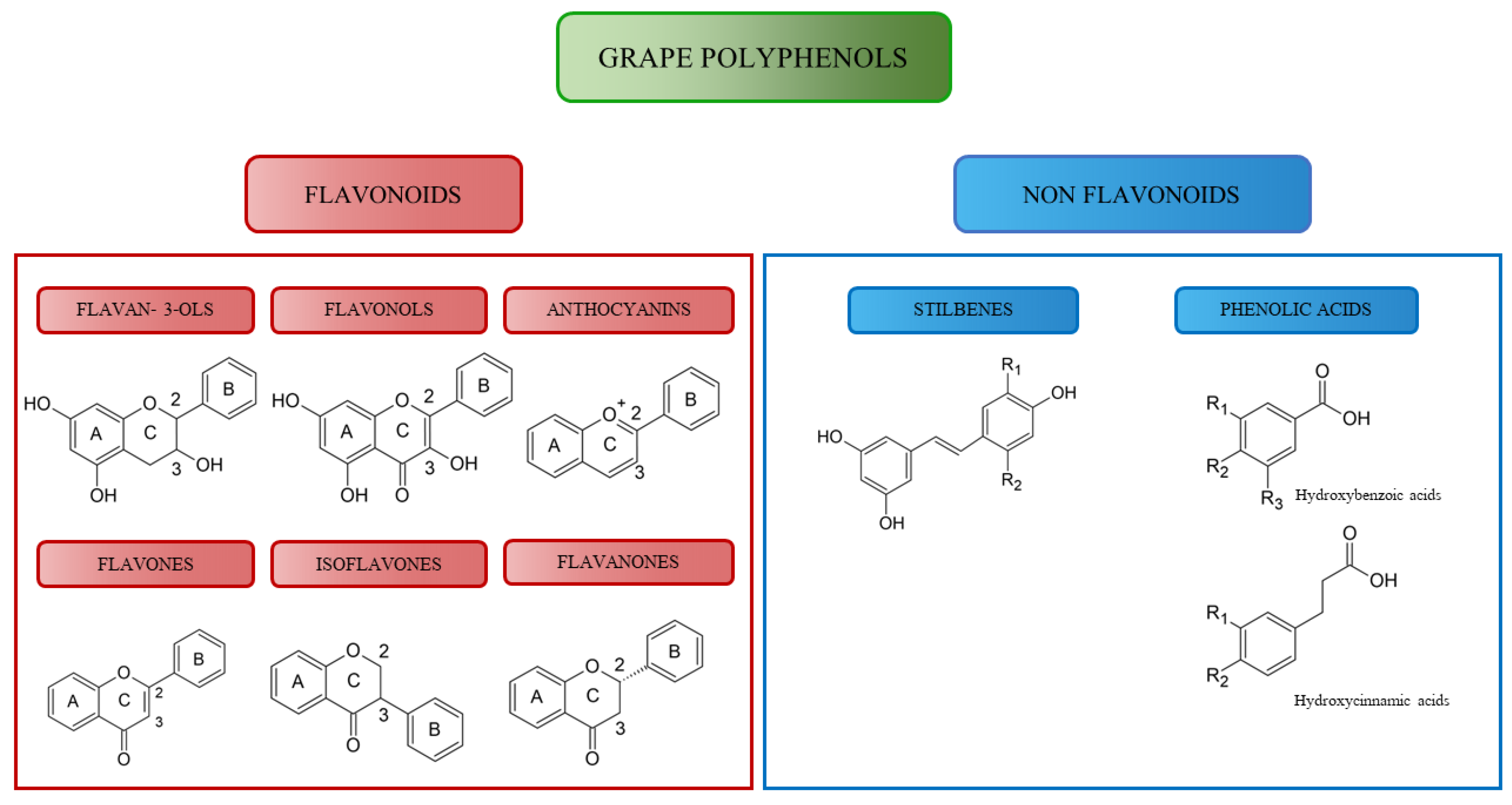
4. Structure and Function of Grape Polyphenol
4.1. The Flavonoids
4.1.1. Flavan-3-ols
4.1.2. Flavonols
4.1.3. Anthocyanes
4.1.4. Flavones
4.1.5. Isoflavones
4.1.6. Flavanones
4.2. Non-Flavonoids
4.2.1. Stilbenes
4.2.2. Phenolic Acids
5. Metabolism of Polyphenols
6. Clinical Studies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zarulli, V.; Sopina, E.; Toffolutti, V.; Lenart, A. Health Care System Efficiency and Life Expectancy: A 140-Country Study. PLoS ONE 2021, 16, e0253450. [Google Scholar] [CrossRef]
- Budreviciute, A.; Damiati, S.; Sabir, D.K.; Onder, K.; Schuller-Goetzburg, P.; Plakys, G.; Katileviciute, A.; Khoja, S.; Kodzius, R. Management and Prevention Strategies for Non-Communicable Diseases (NCDs) and Their Risk Factors. Front. Public Health 2020, 8, 574111. [Google Scholar] [CrossRef]
- Divo, M.J.; Martinez, C.H.; Mannino, D.M. Ageing and the Epidemiology of Multimorbidity. Eur. Respir. J. 2014, 44, 1055–1068. [Google Scholar] [CrossRef]
- Puspitasari, Y.M.; Ministrini, S.; Schwarz, L.; Karch, C.; Liberale, L.; Camici, G.G. Modern Concepts in Cardiovascular Disease: Inflamm-Aging. Front. Cell Dev. Biol. 2022, 10, 882211. [Google Scholar] [CrossRef]
- The GBD 2015 Obesity Collaborators. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in Obesity, Diabetes, and Related Disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Nunan, E.; Wright, C.L.; Semola, O.A.; Subramanian, M.; Balasubramanian, P.; Lovern, P.C.; Fancher, I.S.; Butcher, J.T. Obesity as a Premature Aging Phenotype—Implications for Sarcopenic Obesity. GeroScience 2022, 44, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J.; Piasecki, M.; Atherton, P.J. The Age-Related Loss of Skeletal Muscle Mass and Function: Measurement and Physiology of Muscle Fibre Atrophy and Muscle Fibre Loss in Humans. Ageing Res. Rev. 2018, 47, 123–132. [Google Scholar] [CrossRef]
- Tian, S.; Xu, Y. Association of Sarcopenic Obesity with the Risk of All-Cause Mortality: A Meta-Analysis of Prospective Cohort Studies: Sarcopenic Obesity and Mortality. Geriatr. Gerontol. Int. 2016, 16, 155–166. [Google Scholar] [CrossRef]
- Afonso, C.; Sousa-Santos, A.R.; Santos, A.; Borges, N.; Padrão, P.; Moreira, P.; Amaral, T.F. Frailty Status Is Related to General and Abdominal Obesity in Older Adults. Nutr. Res. 2021, 85, 21–30. [Google Scholar] [CrossRef]
- Huang, P.L. A Comprehensive Definition for Metabolic Syndrome. Dis. Models Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Petersmann, A.; Müller-Wieland, D.; Müller, U.A.; Landgraf, R.; Nauck, M.; Freckmann, G.; Heinemann, L.; Schleicher, E. Definition, Classification and Diagnosis of Diabetes Mellitus. Exp. Clin. Endocrinol. Diabetes 2019, 127, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.-L. The Frailty Syndrome: Definition and Natural History. Clin. Geriatr. Med. 2011, 27, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Sinha, S. Obesity and Aging: Molecular Mechanisms and Therapeutic Approaches. Ageing Res. Rev. 2021, 67, 101268. [Google Scholar] [CrossRef]
- Frontera, W.R.; Ochala, J. Skeletal Muscle: A Brief Review of Structure and Function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef]
- Demontis, F.; Piccirillo, R.; Goldberg, A.L.; Perrimon, N. The Influence of Skeletal Muscle on Systemic Aging and Lifespan. Aging Cell 2013, 12, 943–949. [Google Scholar] [CrossRef]
- Anker, S.D.; Ponikowski, P.; Varney, S.; Chua, T.P.; Clark, A.L.; Webb-Peploe, K.M.; Harrington, D.; Kox, W.J.; Poole-Wilson, P.A.; Coats, A.J. Wasting as Independent Risk Factor for Mortality in Chronic Heart Failure. Lancet 1997, 349, 1050–1053. [Google Scholar] [CrossRef]
- Metter, E.J.; Talbot, L.A.; Schrager, M.; Conwit, R. Skeletal Muscle Strength as a Predictor of All-Cause Mortality in Healthy Men. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2002, 57, B359–B365. [Google Scholar] [CrossRef]
- Gonzalez, A.; Simon, F.; Achiardi, O.; Vilos, C.; Cabrera, D.; Cabello-Verrugio, C. The Critical Role of Oxidative Stress in Sarcopenic Obesity. Oxidative Med. Cell. Longev. 2021, 2021, 4493817. [Google Scholar] [CrossRef] [PubMed]
- Bielecka-Dabrowa, A.; Ebner, N.; Santos, M.R.; Ishida, J.; Hasenfuss, G.; Haehling, S. Cachexia, Muscle Wasting, and Frailty in Cardiovascular Disease. Eur. J. Heart Fail. 2020, 22, 2314–2326. [Google Scholar] [CrossRef]
- Amouzou, C.; Breuker, C.; Fabre, O.; Bourret, A.; Lambert, K.; Birot, O.; Fédou, C.; Dupuy, A.-M.; Cristol, J.-P.; Sutra, T.; et al. Skeletal Muscle Insulin Resistance and Absence of Inflammation Characterize Insulin-Resistant Grade I Obese Women. PLoS ONE 2016, 11, e0154119. [Google Scholar] [CrossRef] [PubMed]
- Aguer, C.; Mercier, J.; Kitzmann, M. Lipid Content and Response to Insulin Are Not Invariably Linked in Human Muscle Cells. Mol. Cell. Endocrinol. 2010, 315, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Aguer, C.; Foretz, M.; Lantier, L.; Hebrard, S.; Viollet, B.; Mercier, J.; Kitzmann, M. Increased FAT/CD36 Cycling and Lipid Accumulation in Myotubes Derived from Obese Type 2 Diabetic Patients. PLoS ONE 2011, 6, e28981. [Google Scholar] [CrossRef]
- Mengeste, A.M.; Rustan, A.C.; Lund, J. Skeletal Muscle Energy Metabolism in Obesity. Obesity 2021, 29, 1582–1595. [Google Scholar] [CrossRef]
- Borén, J.; Taskinen, M.-R.; Olofsson, S.-O.; Levin, M. Ectopic Lipid Storage and Insulin Resistance: A Harmful Relationship. J. Intern. Med. 2013, 274, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.A.; Reed, M.A.; Consitt, L.A.; Martin, O.J.; Haynie, K.R.; Hulver, M.W.; Muoio, D.M.; Dohm, G.L. Lipid Partitioning, Incomplete Fatty Acid Oxidation, and Insulin Signal Transduction in Primary Human Muscle Cells: Effects of Severe Obesity, Fatty Acid Incubation, and Fatty Acid Translocase/CD36 Overexpression. J. Clin. Endocrinol. Metab. 2010, 95, 3400–3410. [Google Scholar] [CrossRef] [PubMed]
- Løvsletten, N.G.; Rustan, A.C.; Laurens, C.; Thoresen, G.H.; Moro, C.; Nikolić, N. Primary Defects in Lipid Handling and Resistance to Exercise in Myotubes from Obese Donors with and without Type 2 Diabetes. Appl. Physiol. Nutr. Metab. 2020, 45, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Laurens, C.; Moro, C. Intramyocellular Fat Storage in Metabolic Diseases. Horm. Mol. Biol. Clin. Investig. 2016, 26, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Kitessa, S.; Abeywardena, M. Lipid-Induced Insulin Resistance in Skeletal Muscle: The Chase for the Culprit Goes from Total Intramuscular Fat to Lipid Intermediates, and Finally to Species of Lipid Intermediates. Nutrients 2016, 8, 466. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, B.; Summers, S.A. Ceramides in Metabolism: Key Lipotoxic Players. Annu. Rev. Physiol. 2021, 83, 303–330. [Google Scholar] [CrossRef] [PubMed]
- Holloway, G.P.; Thrush, A.B.; Heigenhauser, G.J.F.; Tandon, N.N.; Dyck, D.J.; Bonen, A.; Spriet, L.L. Skeletal Muscle Mitochondrial FAT/CD36 Content and Palmitate Oxidation Are Not Decreased in Obese Women. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1782–E1789. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, I.R.; Joshi, M. CPT1A-Mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Skeletal Muscle Inflammation and Insulin Resistance in Obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Akhmedov, D.; Berdeaux, R. The Effects of Obesity on Skeletal Muscle Regeneration. Front. Physiol. 2013, 4, 371. [Google Scholar] [CrossRef]
- Jiang, A.; Guo, H.; Zhang, L.; Jiang, X.; Zhang, X.; Wu, W.; Liu, H. Free Fatty Acid Impairs Myogenic Differentiation through the AMPKα-MicroRNA 206 Pathway. Mol. Cell. Biol. 2022, 42, e00327-21. [Google Scholar] [CrossRef]
- Tallis, J.; James, R.S.; Seebacher, F. The Effects of Obesity on Skeletal Muscle Contractile Function. J. Exp. Biol. 2018, 221, jeb163840. [Google Scholar] [CrossRef]
- Abdelaal, M.; le Roux, C.W.; Docherty, N.G. Morbidity and Mortality Associated with Obesity. Ann. Transl. Med. 2017, 5, 161. [Google Scholar] [CrossRef]
- Peeters, A.; Barendregt, J.J.; Willekens, F.; Mackenbach, J.P.; Mamun, A.A.; Bonneux, L.; NEDCOM, The Netherlands Epidemiology and Demography Compression of Morbidity Research Group. Obesity in Adulthood and Its Consequences for Life Expectancy: A Life-Table Analysis. Ann. Intern. Med. 2003, 138, 24. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; Simonsick, E.M.; Tylavsky, F.A.; Visser, M.; Newman, A.B.; et al. The Loss of Skeletal Muscle Strength, Mass, and Quality in Older Adults: The Health, Aging and Body Composition Study. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Romanello, V.; Sandri, M. Mitochondrial Quality Control and Muscle Mass Maintenance. Front. Physiol. 2016, 6, 422. [Google Scholar] [CrossRef]
- Tam, B.T.; Morais, J.A.; Santosa, S. Obesity and Ageing: Two Sides of the Same Coin. Obes. Rev. 2020, 21, e12991. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Luo, Q.; Nie, R.; Yang, X.; Tang, Z.; Chen, H. Potential Implications of Polyphenols on Aging Considering Oxidative Stress, Inflammation, Autophagy, and Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 2175–2193. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.; Gatta, P.D.; Cameron-Smith, D. Aging and Its Effects on Inflammation in Skeletal Muscle at Rest and Following Exercise-Induced Muscle Injury. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1485–R1495. [Google Scholar] [CrossRef]
- Visser, M.; Goodpaster, B.H.; Kritchevsky, S.B.; Newman, A.B.; Nevitt, M.; Rubin, S.M.; Simonsick, E.M.; Harris, T.B.; Health ABC Study. Muscle Mass, Muscle Strength, and Muscle Fat Infiltration as Predictors of Incident Mobility Limitations in Well-Functioning Older Persons. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2005, 60, 324–333. [Google Scholar] [CrossRef]
- Milan, G.; Romanello, V.; Pescatore, F.; Armani, A.; Paik, J.-H.; Frasson, L.; Seydel, A.; Zhao, J.; Abraham, R.; Goldberg, A.L.; et al. Regulation of Autophagy and the Ubiquitin–Proteasome System by the FoxO Transcriptional Network during Muscle Atrophy. Nat. Commun. 2015, 6, 6670. [Google Scholar] [CrossRef] [PubMed]
- Romanello, V. The Interplay between Mitochondrial Morphology and Myomitokines in Aging Sarcopenia. Int. J. Mol. Sci. 2020, 22, 91. [Google Scholar] [CrossRef] [PubMed]
- Boengler, K.; Kosiol, M.; Mayr, M.; Schulz, R.; Rohrbach, S. Mitochondria and Ageing: Role in Heart, Skeletal Muscle and Adipose Tissue: Mitochondria and Ageing. J. Cachexia Sarcopenia Muscle 2017, 8, 349–369. [Google Scholar] [CrossRef]
- Romanello, V.; Guadagnin, E.; Gomes, L.; Roder, I.; Sandri, C.; Petersen, Y.; Milan, G.; Masiero, E.; Del Piccolo, P.; Foretz, M.; et al. Mitochondrial Fission and Remodelling Contributes to Muscle Atrophy. EMBO J. 2010, 29, 1774–1785. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Blum, A.; Liu, J.; Finkel, T. The Role of Mitochondria in Aging. J. Clin. Investig. 2018, 128, 3662–3670. [Google Scholar] [CrossRef]
- Huang, D.-D.; Fan, S.-D.; Chen, X.-Y.; Yan, X.-L.; Zhang, X.-Z.; Ma, B.-W.; Yu, D.-Y.; Xiao, W.-Y.; Zhuang, C.-L.; Yu, Z. Nrf2 Deficiency Exacerbates Frailty and Sarcopenia by Impairing Skeletal Muscle Mitochondrial Biogenesis and Dynamics in an Age-Dependent Manner. Exp. Gerontol. 2019, 119, 61–73. [Google Scholar] [CrossRef]
- Conte, M.; Martucci, M.; Sandri, M.; Franceschi, C.; Salvioli, S. The Dual Role of the Pervasive “Fattish” Tissue Remodeling With Age. Front. Endocrinol. 2019, 10, 114. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Proteostasis and Aging. Nat. Med. 2015, 21, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Aoi, W.; Yamaguchi, A. Current Understanding of Sarcopenia: Possible Candidates Modulating Muscle Mass. Pflugers Arch. Eur. J. Physiol. 2015, 467, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Gumucio, J.P.; Mendias, C.L. Atrogin-1, MuRF-1, and Sarcopenia. Endocrine 2013, 43, 12–21. [Google Scholar] [CrossRef]
- Jura, M.; Kozak, L.P. Obesity and Related Consequences to Ageing. Age 2016, 38, 23. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef]
- Meng, S.-J.; Yu, L.-J. Oxidative Stress, Molecular Inflammation and Sarcopenia. Int. J. Mol. Sci. 2010, 11, 1509–1526. [Google Scholar] [CrossRef]
- Rotariu, D.; Babes, E.E.; Tit, D.M.; Moisi, M.; Bustea, C.; Stoicescu, M.; Radu, A.-F.; Vesa, C.M.; Behl, T.; Bungau, A.F.; et al. Oxidative Stress—Complex Pathological Issues Concerning the Hallmark of Cardiovascular and Metabolic Disorders. Biomed. Pharmacother. 2022, 152, 113238. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding Oxidants and Antioxidants: Classical Team with New Players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef] [PubMed]
- Bouviere, J.; Fortunato, R.S.; Dupuy, C.; Werneck-de-Castro, J.P.; Carvalho, D.P.; Louzada, R.A. Exercise-Stimulated ROS Sensitive Signaling Pathways in Skeletal Muscle. Antioxidants 2021, 10, 537. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.P. Inflammation and Its Role in Regeneration and Repair: A Caution for Novel Anti-Inflammatory Therapies. Circ. Res. 2019, 124, 1166–1168. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Nelke, C.; Dziewas, R.; Minnerup, J.; Meuth, S.G.; Ruck, T. Skeletal Muscle as Potential Central Link between Sarcopenia and Immune Senescence. EBioMedicine 2019, 49, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Guijarro-Muñoz, I.; Compte, M.; Álvarez-Cienfuegos, A.; Álvarez-Vallina, L.; Sanz, L. Lipopolysaccharide Activates Toll-like Receptor 4 (TLR4)-Mediated NF-ΚB Signaling Pathway and Proinflammatory Response in Human Pericytes. J. Biol. Chem. 2014, 289, 2457–2468. [Google Scholar] [CrossRef]
- Lee, S.; Norheim, F.; Gulseth, H.L.; Langleite, T.M.; Kolnes, K.J.; Tangen, D.S.; Stadheim, H.K.; Gilfillan, G.D.; Holen, T.; Birkeland, K.I.; et al. Interaction between Plasma Fetuin-A and Free Fatty Acids Predicts Changes in Insulin Sensitivity in Response to Long-Term Exercise. Physiol. Rep. 2017, 5, e13183. [Google Scholar] [CrossRef]
- Di Paola, M.; Lorusso, M. Interaction of Free Fatty Acids with Mitochondria: Coupling, Uncoupling and Permeability Transition. Biochim. Biophys. Acta BBA Bioenerg. 2006, 1757, 1330–1337. [Google Scholar] [CrossRef]
- Lian, D.; Chen, M.-M.; Wu, H.; Deng, S.; Hu, X. The Role of Oxidative Stress in Skeletal Muscle Myogenesis and Muscle Disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Kma, L.; Baruah, T.J. The Interplay of ROS and the PI3K/Akt Pathway in Autophagy Regulation. Biotech. Appl. Biochem. 2022, 69, 248–264. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J.; Murach, K.A. Anabolic and Catabolic Signaling Pathways That Regulate Skeletal Muscle Mass. In Nutrition and Enhanced Sports Performance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 275–290. ISBN 978-0-12-813922-6. [Google Scholar]
- Jang, Y.C.; Rodriguez, K.; Lustgarten, M.S.; Muller, F.L.; Bhattacharya, A.; Pierce, A.; Choi, J.J.; Lee, N.H.; Chaudhuri, A.; Richardson, A.G.; et al. Superoxide-Mediated Oxidative Stress Accelerates Skeletal Muscle Atrophy by Synchronous Activation of Proteolytic Systems. GeroScience 2020, 42, 1579–1591. [Google Scholar] [CrossRef] [PubMed]
- Nader, G.A. Molecular Determinants of Skeletal Muscle Mass: Getting the “AKT” Together. Int. J. Biochem. Cell Biol. 2005, 37, 1985–1996. [Google Scholar] [CrossRef]
- Kitaoka, Y. The Role of Nrf2 in Skeletal Muscle on Exercise Capacity. Antioxidants 2021, 10, 1712. [Google Scholar] [CrossRef]
- Huang, C.-J.; McAllister, M.J.; Slusher, A.L.; Webb, H.E.; Mock, J.T.; Acevedo, E.O. Obesity-Related Oxidative Stress: The Impact of Physical Activity and Diet Manipulation. Sports Med. Open 2015, 1, 32. [Google Scholar] [CrossRef]
- Renaud, S.; de Lorgeril, M. Wine, Alcohol, Platelets, and the French Paradox for Coronary Heart Disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Aune, D. Plant Foods, Antioxidant Biomarkers, and the Risk of Cardiovascular Disease, Cancer, and Mortality: A Review of the Evidence. Adv. Nutr. 2019, 10, S404–S421. [Google Scholar] [CrossRef]
- Tanaka, T.; Talegawkar, S.A.; Jin, Y.; Bandinelli, S.; Ferrucci, L. Association of Adherence to the Mediterranean-Style Diet with Lower Frailty Index in Older Adults. Nutrients 2021, 13, 1129. [Google Scholar] [CrossRef]
- Amiot, M.-J.; Latgé, C.; Plumey, L.; Raynal, S. Intake Estimation of Phytochemicals in a French Well-Balanced Diet. Nutrients 2021, 13, 3628. [Google Scholar] [CrossRef]
- Neveu, V.; Perez-Jimenez, J.; Vos, F.; Crespy, V.; du Chaffaut, L.; Mennen, L.; Knox, C.; Eisner, R.; Cruz, J.; Wishart, D.; et al. Phenol-Explorer: An Online Comprehensive Database on Polyphenol Contents in Foods. Database 2010, 2010, bap024. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remon, A.; M’Hiri, N.; Garcia-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A Major Update of the Phenol-Explorer Database to Incorporate Data on the Effects of Food Processing on Polyphenol Content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Urpi-Sarda, M.; Boto-Ordonez, M.; Knox, C.; Llorach, R.; Eisner, R.; Cruz, J.; Neveu, V.; Wishart, D.; Manach, C.; et al. Phenol-Explorer 2.0: A Major Update of the Phenol-Explorer Database Integrating Data on Polyphenol Metabolism and Pharmacokinetics in Humans and Experimental Animals. Database 2012, 2012, bas031. [Google Scholar] [CrossRef]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic Biochemical Mechanisms behind the Health Benefits of Polyphenols. Mol. Asp. Med. 2010, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef]
- Youn, H.S.; Lee, J.Y.; Saitoh, S.I.; Miyake, K.; Kang, K.W.; Choi, Y.J.; Hwang, D.H. Suppression of MyD88- and TRIF-Dependent Signaling Pathways of Toll-like Receptor by (−)-Epigallocatechin-3-Gallate, a Polyphenol Component of Green Tea. Biochem. Pharmacol. 2006, 72, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Casanova, E.; Salvadó, J.; Crescenti, A.; Gibert-Ramos, A. Epigallocatechin Gallate Modulates Muscle Homeostasis in Type 2 Diabetes and Obesity by Targeting Energetic and Redox Pathways: A Narrative Review. Int. J. Mol. Sci. 2019, 20, 532. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, S.; Zhang, W.; Zhao, P.; He, B.; Wu, N.; Han, P. Epigallocatechin-3-O-Gallate (EGCG) Attenuates FFAs-Induced Peripheral Insulin Resistance through AMPK Pathway and Insulin Signaling Pathway in Vivo. Diabetes Res. Clin. Pract. 2011, 93, 205–214. [Google Scholar] [CrossRef]
- Pallauf, K.; Rimbach, G. Autophagy, Polyphenols and Healthy Ageing. Ageing Res. Rev. 2013, 12, 237–252. [Google Scholar] [CrossRef]
- Xu, M.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; Luo, Y.; Zheng, P.; Yu, J.; He, J. Grape Seed Proanthocyanidin Extract Promotes Skeletal Muscle Fiber Type Transformation via AMPK Signaling Pathway. J. Nutr. Biochem. 2020, 84, 108462. [Google Scholar] [CrossRef]
- Costa, L.G.; Garrick, J.M.; Roquè, P.J.; Pellacani, C. Mechanisms of Neuroprotection by Quercetin: Counteracting Oxidative Stress and More. Oxidative Med. Cell. Longev. 2016, 2016, 2986796. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Mukai, Y. Modulation of Chronic Inflammation by Quercetin: The Beneficial Effects on Obesity. JIR 2020, 13, 421–431. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Role of Red Grape Polyphenols as Antidiabetic Agents. Integr. Med. Res. 2014, 3, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a Dietary Anti-Inflammatory Agent: Current Therapeutic Standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef] [PubMed]
- Mozos, I.; Flangea, C.; Vlad, D.C.; Gug, C.; Mozos, C.; Stoian, D.; Luca, C.T.; Horbańczuk, J.O.; Horbańczuk, O.K.; Atanasov, A.G. Effects of Anthocyanins on Vascular Health. Biomolecules 2021, 11, 811. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. The Influence of Supplementation of Anthocyanins on Obesity-Associated Comorbidities: A Concise Review. Foods 2020, 9, 687. [Google Scholar] [CrossRef]
- Jia, Y.; Wu, C.; Kim, Y.-S.; Yang, S.O.; Kim, Y.; Kim, J.-S.; Jeong, M.-Y.; Lee, J.H.; Kim, B.; Lee, S.; et al. A Dietary Anthocyanin Cyanidin-3-O-Glucoside Binds to PPARs to Regulate Glucose Metabolism and Insulin Sensitivity in Mice. Commun. Biol. 2020, 3, 514. [Google Scholar] [CrossRef]
- Geng, X.; Ji, J.; Liu, Y.; Li, X.; Chen, Y.; Su, L.; Zhao, L. Cyanidin-3-O-Glucoside Supplementation Ameliorates Metabolic Insulin Resistance via Restoration of Nitric Oxide-Mediated Endothelial Insulin Transport. Mol. Nutr. Food Res. 2022, 66, 2100742. [Google Scholar] [CrossRef]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Lv, T.; Cui, C.; Liu, M. Investigation of the Anti-Inflammatory and Antioxidant Activities of Luteolin, Kaempferol, Apigenin and Quercetin. S. Afr. J. Bot. 2021, 137, 257–264. [Google Scholar] [CrossRef]
- Wang, D.; Yang, Y.; Zou, X.; Zhang, J.; Zheng, Z.; Wang, Z. Antioxidant Apigenin Relieves Age-Related Muscle Atrophy by Inhibiting Oxidative Stress and Hyperactive Mitophagy and Apoptosis in Skeletal Muscle of Mice. J. Gerontol. Ser. A 2020, 75, 2081–2088. [Google Scholar] [CrossRef]
- Park, M.-H.; Ju, J.-W.; Park, M.; Han, J. Daidzein Inhibits Carbohydrate Digestive Enzymes in Vitro and Alleviates Postprandial Hyperglycemia in Diabetic Mice. Eur. J. Pharmacol. 2013, 712, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Sarkar, S.; Bordoloi, J.; Wann, S.B.; Kalita, J.; Manna, P. Daidzein, Its Effects on Impaired Glucose and Lipid Metabolism and Vascular Inflammation Associated with Type 2 Diabetes: Prophylactic Role of Daidzein in Type 2 Diabetes. BioFactors 2018, 44, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Cruz, M.; Godinez-Salas, E.T.; Sanchez-Tapia, M.; Torres-Villalobos, G.; Pichardo-Ontiveros, E.; Guizar-Heredia, R.; Arteaga-Sanchez, L.; Gamba, G.; Mojica-Espinosa, R.; Schcolnik-Cabrera, A.; et al. Genistein Stimulates Insulin Sensitivity through Gut Microbiota Reshaping and Skeletal Muscle AMPK Activation in Obese Subjects. BMJ Open Diabetes Res. Care 2020, 8, e000948. [Google Scholar] [CrossRef]
- Rehman, K.; Khan, I.I.; Akash, M.S.H.; Jabeen, K.; Haider, K. Naringenin Downregulates Inflammation-mediated Nitric Oxide Overproduction and Potentiates Endogenous Antioxidant Status during Hyperglycemia. J. Food Biochem. 2020, 44, e13422. [Google Scholar] [CrossRef] [PubMed]
- Dhanya, R.; Jayamurthy, P. In Vitro Evaluation of Antidiabetic Potential of Hesperidin and Its Aglycone Hesperetin under Oxidative Stress in Skeletal Muscle Cell Line. Cell Biochem. Funct. 2020, 38, 419–427. [Google Scholar] [CrossRef]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Kang, W.; Hong, H.J.; Guan, J.; Kim, D.G.; Yang, E.-J.; Koh, G.; Park, D.; Han, C.H.; Lee, Y.-J.; Lee, D.-H. Resveratrol Improves Insulin Signaling in a Tissue-Specific Manner under Insulin-Resistant Conditions Only: In Vitro and in Vivo Experiments in Rodents. Metabolism 2012, 61, 424–433. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, X.; Chen, K.; Lang, H.; Zhang, Y.; Hou, P.; Ran, L.; Zhou, M.; Zheng, J.; Yi, L.; et al. Resveratrol Prevents Sarcopenic Obesity by Reversing Mitochondrial Dysfunction and Oxidative Stress via the PKA/LKB1/AMPK Pathway. Aging 2019, 11, 2217–2240. [Google Scholar] [CrossRef]
- Bai, C.-H.; Alizargar, J.; Peng, C.-Y.; Wu, J.-P. Combination of Exercise Training and Resveratrol Attenuates Obese Sarcopenia in Skeletal Muscle Atrophy. Chin. J. Physiol. 2020, 63, 101. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic Acids: Natural Versatile Molecules with Promising Therapeutic Applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; El-Twab, S.M.A.; Yousef, A.I.; Reheim, E.S.A.; Ashour, M.B. Modulation of Hyperglycemia and Dyslipidemia in Experimental Type 2 Diabetes by Gallic Acid and P-Coumaric Acid: The Role of Adipocytokines and PPARγ. Biomed. Pharmacother. 2018, 105, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.; Lopes, N.M.F. Cardiovascular Effects of Caffeic Acid and Its Derivatives: A Comprehensive Review. Front. Physiol. 2020, 11, 595516. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Park, J.; Kim, H.; Sim, J.; Youn, D.; Kang, J.; Lim, S.; Jeong, M.; Yang, W.M.; Lee, S.; et al. Vanillic Acid Attenuates Obesity via Activation of the AMPK Pathway and Thermogenic Factors in Vivo and in Vitro. FASEB J. 2018, 32, 1388–1402. [Google Scholar] [CrossRef]
- Rashedinia, M.; Alimohammadi, M.; Shalfroushan, N.; Khoshnoud, M.J.; Mansourian, M.; Azarpira, N.; Sabahi, Z. Neuroprotective Effect of Syringic Acid by Modulation of Oxidative Stress and Mitochondrial Mass in Diabetic Rats. BioMed Res. Int. 2020, 2020, 8297984. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, J.; Fezeu, L.; Touvier, M.; Arnault, N.; Manach, C.; Hercberg, S.; Galan, P.; Scalbert, A. Dietary Intake of 337 Polyphenols in French Adults. Am. J. Clin. Nutr. 2011, 93, 1220–1228. [Google Scholar] [CrossRef]
- Faustino, M.; Veiga, M.; Sousa, P.; Costa, E.; Silva, S.; Pintado, M. Agro-Food Byproducts as a New Source of Natural Food Additives. Molecules 2019, 24, 1056. [Google Scholar] [CrossRef]
- Magrone, T.; Magrone, M.; Russo, M.A.; Jirillo, E. Recent Advances on the Anti-Inflammatory and Antioxidant Properties of Red Grape Polyphenols: In Vitro and In Vivo Studies. Antioxidants 2019, 9, 35. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 Signaling in Oxidative and Reductive Stress. Biochim. Biophys. Acta BBA Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and Mitochondria: Activation of Cytoprotective Pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef]
- Annunziata, G.; Jimenez-García, M.; Tejada, S.; Moranta, D.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Sureda, A.; Novellino, E.; Capó, X. Grape Polyphenols Ameliorate Muscle Decline Reducing Oxidative Stress and Oxidative Damage in Aged Rats. Nutrients 2020, 12, 1280. [Google Scholar] [CrossRef] [PubMed]
- Lambert, K.; Hokayem, M.; Thomas, C.; Fabre, O.; Cassan, C.; Bourret, A.; Bernex, F.; Feuillet-Coudray, C.; Notarnicola, C.; Mercier, J.; et al. Combination of Nutritional Polyphenols Supplementation with Exercise Training Counteracts Insulin Resistance and Improves Endurance in High-Fat Diet-Induced Obese Rats. Sci. Rep. 2018, 8, 2885. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Gerhart-Hines, Z.; Puigserver, P. Metabolic Adaptations through the PGC-1α and SIRT1 Pathways. FEBS Lett. 2008, 582, 46–53. [Google Scholar] [CrossRef]
- Iside, C.; Scafuro, M.; Nebbioso, A.; Altucci, L. SIRT1 Activation by Natural Phytochemicals: An Overview. Front. Pharmacol. 2020, 11, 1225. [Google Scholar] [CrossRef]
- Collier, C.A.; Bruce, C.R.; Smith, A.C.; Lopaschuk, G.; Dyck, D.J. Metformin Counters the Insulin-Induced Suppression of Fatty Acid Oxidation and Stimulation of Triacylglycerol Storage in Rodent Skeletal Muscle. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E182–E189. [Google Scholar] [CrossRef]
- Hokayem, M.; Blond, E.; Vidal, H.; Lambert, K.; Meugnier, E.; Feillet-Coudray, C.; Coudray, C.; Pesenti, S.; Luyton, C.; Lambert-Porcheron, S.; et al. Grape Polyphenols Prevent Fructose-Induced Oxidative Stress and Insulin Resistance in First-Degree Relatives of Type 2 Diabetic Patients. Diabetes Care 2013, 36, 1454–1461. [Google Scholar] [CrossRef]
- Shabani, M.; Sadeghi, A.; Hosseini, H.; Teimouri, M.; Babaei Khorzoughi, R.; Pasalar, P.; Meshkani, R. Resveratrol Alleviates Obesity-Induced Skeletal Muscle Inflammation via Decreasing M1 Macrophage Polarization and Increasing the Regulatory T Cell Population. Sci. Rep. 2020, 10, 3791. [Google Scholar] [CrossRef]
- Alamdari, N.; Aversa, Z.; Castillero, E.; Gurav, A.; Petkova, V.; Tizio, S.; Hasselgren, P.-O. Resveratrol Prevents Dexamethasone-Induced Expression of the Muscle Atrophy-Related Ubiquitin Ligases Atrogin-1 and MuRF1 in Cultured Myotubes through a SIRT1-Dependent Mechanism. Biochem. Biophys. Res. Commun. 2012, 417, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Momken, I.; Stevens, L.; Bergouignan, A.; Desplanches, D.; Rudwill, F.; Chery, I.; Zahariev, A.; Zahn, S.; Stein, T.P.; Sebedio, J.L.; et al. Resveratrol Prevents the Wasting Disorders of Mechanical Unloading by Acting as a Physical Exercise Mimetic in the Rat. FASEB J. 2011, 25, 3646–3660. [Google Scholar] [CrossRef]
- Tuladhar, P.; Sasidharan, S.; Saudagar, P. Role of Phenols and Polyphenols in Plant Defense Response to Biotic and Abiotic Stresses. In Biocontrol Agents and Secondary Metabolites; Elsevier: Amsterdam, The Netherlands, 2021; pp. 419–441. ISBN 978-0-12-822919-4. [Google Scholar]
- Benbouguerra, N.; Valls-Fonayet, J.; Krisa, S.; Garcia, F.; Saucier, C.; Richard, T.; Hornedo-Ortega, R. Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells. Foods 2021, 10, 541. [Google Scholar] [CrossRef] [PubMed]
- Savoia, D. Plant-Derived Antimicrobial Compounds: Alternatives to Antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Cherubim, D.J.; Martins, C.V.; Fariña, L.; Lucca, R.A. Polyphenols as Natural Antioxidants in Cosmetics Applications. J. Cosmet. Dermatol. 2020, 19, 33–37. [Google Scholar] [CrossRef]
- Frankel, E.N.; German, J.B.; Kinsella, J.E.; Parks, E.; Kanner, J. Inhibition of Oxidation of Human Low-Density Lipoprotein by Phenolic Substances in Red Wine. Lancet 1993, 341, 454–457. [Google Scholar] [CrossRef]
- Frankel, E.N.; Waterhouse, A.L.; Kinsella, J.E. Inhibition of Human LDL Oxidation by Resveratrol. Lancet 1993, 341, 1103–1104. [Google Scholar] [CrossRef]
- Sun, A.Y.; Simonyi, A.; Sun, G.Y. The “French Paradox” and beyond: Neuroprotective Effects of Polyphenols. Free Radic. Biol. Med. 2002, 32, 314–318. [Google Scholar] [CrossRef]
- Gutiérrez-Escobar, R.; Aliaño-González, M.J.; Cantos-Villar, E. Wine Polyphenol Content and Its Influence on Wine Quality and Properties: A Review. Molecules 2021, 26, 718. [Google Scholar] [CrossRef]
- Watson, R.R. Polyphenols: Mechanisms of Action in Human Health and Disease, 1st ed.; Elsevier: San Diego, CA, USA, 2018; ISBN 978-0-12-813006-3. [Google Scholar]
- Baskar, V.; Venkatesh, R.; Ramalingam, S. Flavonoids (Antioxidants Systems) in Higher Plants and Their Response to Stresses. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerand, 2018; pp. 253–268. ISBN 978-3-319-75087-3. [Google Scholar]
- Ruiz-Cruz, S.; Chaparro-Hernández, S.; Ruiz, K.L.H.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Ortega, L.E.G.; de Ornelas-Paz, J.J.; Mata, M.A.L. Flavonoids: Important Biocompounds in Food. In Flavonoids—From Biosynthesis to Human Health; Justino, G.C., Ed.; InTech: Rijeka, Croatia, 2017; ISBN 978-953-51-3423-7. [Google Scholar]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant Flavonoids: Classification, Distribution, Biosynthesis, and Antioxidant Activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Aron, P.M.; Kennedy, J.A. Flavan-3-Ols: Nature, Occurrence and Biological Activity. Mol. Nutr. Food Res. 2008, 52, 79–104. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Padilla-González, G.F.; Grosskopf, E.; Sadgrove, N.J.; Simmonds, M.S.J. Chemical Diversity of Flavan-3-Ols in Grape Seeds: Modulating Factors and Quality Requirements. Plants 2022, 11, 809. [Google Scholar] [CrossRef] [PubMed]
- Latos-Brozio, M.; Masek, A. Structure-Activity Relationships Analysis of Monomeric and Polymeric Polyphenols (Quercetin, Rutin and Catechin) Obtained by Various Polymerization Methods. Chem. Biodivers. 2019, 16, e1900426. [Google Scholar] [CrossRef]
- Habib, H.M.; El-Fakharany, E.M.; Kheadr, E.; Ibrahim, W.H. Grape Seed Proanthocyanidin Extract Inhibits DNA and Protein Damage and Labile Iron, Enzyme, and Cancer Cell Activities. Sci. Rep. 2022, 12, 12393. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S. Grapevine Structure and Function. In Wine Science; Elsevier: Amsterdam, The Netherlands, 2008; pp. 50–107. ISBN 978-0-12-373646-8. [Google Scholar]
- Le, N.H.; Kim, C.-S.; Park, T.; Park, J.H.Y.; Sung, M.-K.; Lee, D.G.; Hong, S.-M.; Choe, S.-Y.; Goto, T.; Kawada, T.; et al. Quercetin Protects against Obesity-Induced Skeletal Muscle Inflammation and Atrophy. Mediat. Inflamm. 2014, 2014, 834294. [Google Scholar] [CrossRef] [PubMed]
- Ohmae, S.; Akazawa, S.; Takahashi, T.; Izumo, T.; Rogi, T.; Nakai, M. Quercetin Attenuates Adipogenesis and Fibrosis in Human Skeletal Muscle. Biochem. Biophys. Res. Commun. 2022, 615, 24–30. [Google Scholar] [CrossRef]
- Eseberri, I.; Laurens, C.; Miranda, J.; Louche, K.; Lasa, A.; Moro, C.; Portillo, M.P. Effects of Physiological Doses of Resveratrol and Quercetin on Glucose Metabolism in Primary Myotubes. Int. J. Mol. Sci. 2021, 22, 1384. [Google Scholar] [CrossRef]
- Ding, Y.; Dai, X.; Zhang, Z.; Li, Y. Myricetin Attenuates Hyperinsulinemia-Induced Insulin Resistance in Skeletal Muscle Cells. Eur. Food Res. Technol. 2012, 234, 873–881. [Google Scholar] [CrossRef]
- Sousa, C. Anthocyanins, Carotenoids and Chlorophylls in Edible Plant Leaves Unveiled by Tandem Mass Spectrometry. Foods 2022, 11, 1924. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental Significance of Anthocyanins in Plant Stress Responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Hogan, S.; Canning, C.; Sun, S.; Sun, X.; Zhou, K. Effects of Grape Pomace Antioxidant Extract on Oxidative Stress and Inflammation in Diet Induced Obese Mice. J. Agric. Food Chem. 2010, 58, 11250–11256. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef]
- Valletta, A.; Iozia, L.M.; Leonelli, F. Impact of Environmental Factors on Stilbene Biosynthesis. Plants 2021, 10, 90. [Google Scholar] [CrossRef]
- Pecyna, P.; Wargula, J.; Murias, M.; Kucinska, M. More Than Resveratrol: New Insights into Stilbene-Based Compounds. Biomolecules 2020, 10, 1111. [Google Scholar] [CrossRef] [PubMed]
- Babazadeh, A.; Taghvimi, A.; Hamishehkar, H.; Tabibiazar, M. Development of New Ultrasonic–Solvent Assisted Method for Determination of Trans-Resveratrol from Red Grapes: Optimization, Characterization, and Antioxidant Activity (ORAC Assay). Food Biosci. 2017, 20, 36–42. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic Potential of Resveratrol: The in Vivo Evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Koushki, M.; Amiri-Dashatan, N.; Ahmadi, N.; Abbaszadeh, H.-A.; Rezaei-Tavirani, M. Resveratrol: A Miraculous Natural Compound for Diseases Treatment. Food Sci. Nutr. 2018, 6, 2473–2490. [Google Scholar] [CrossRef] [PubMed]
- Shadfar, S.; Couch, M.E.; McKinney, K.A.; Weinstein, L.J.; Yin, X.; Rodríguez, J.E.; Guttridge, D.C.; Willis, M. Oral Resveratrol Therapy Inhibits Cancer-Induced Skeletal Muscle and Cardiac Atrophy In Vivo. Nutr. Cancer 2011, 63, 749–762. [Google Scholar] [CrossRef]
- Jackson, J.R.; Ryan, M.J.; Alway, S.E. Long-Term Supplementation With Resveratrol Alleviates Oxidative Stress but Does Not Attenuate Sarcopenia in Aged Mice. J. Gerontol. Ser. A 2011, 66A, 751–764. [Google Scholar] [CrossRef]
- Sirago, G.; Toniolo, L.; Crea, E.; Giacomello, E. A Short-Term Treatment with Resveratrol Improves the Inflammatory Conditions of Middle-Aged Mice Skeletal Muscles. Int. J. Food Sci. Nutr. 2022, 73, 630–637. [Google Scholar] [CrossRef]
- Ali, G.; Neda, G. Flavonoids and Phenolic Acids: Role and Biochemical Activity in Plants and Human. J. Med. Plants Res. 2011, 5, 6697–6703. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic Acids from Vegetables: A Review on Processing Stability and Health Benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef]
- Doan, K.V.; Ko, C.M.; Kinyua, A.W.; Yang, D.J.; Choi, Y.-H.; Oh, I.Y.; Nguyen, N.M.; Ko, A.; Choi, J.W.; Jeong, Y.; et al. Gallic Acid Regulates Body Weight and Glucose Homeostasis Through AMPK Activation. Endocrinology 2015, 156, 157–168. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Cipolletti, M.; Solar Fernandez, V.; Montalesi, E.; Marino, M.; Fiocchetti, M. Beyond the Antioxidant Activity of Dietary Polyphenols in Cancer: The Modulation of Estrogen Receptors (ERs) Signaling. Int. J. Mol. Sci. 2018, 19, 2624. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.-B.; Lee, H.-S.; Hong, J.S.; Kim, D.H.; Moon, J.M.; Park, Y. Effects of Tannase-Converted Green Tea Extract on Skeletal Muscle Development. BMC Complement. Med. Ther. 2020, 20, 47. [Google Scholar] [CrossRef] [PubMed]
- Nikawa, T.; Ulla, A.; Sakakibara, I. Polyphenols and Their Effects on Muscle Atrophy and Muscle Health. Molecules 2021, 26, 4887. [Google Scholar] [CrossRef] [PubMed]
- Dudoit, A.; Benbouguerra, N.; Richard, T.; Hornedo-Ortega, R.; Valls-Fonayet, J.; Coussot, G.; Saucier, C. α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages. Biomolecules 2020, 10, 1088. [Google Scholar] [CrossRef]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding Phenolic Acids Inhibition of α-Amylase and α-Glucosidase and Influence of Reaction Conditions. Food Chem. 2022, 372, 131231. [Google Scholar] [CrossRef]
- Arora, I.; Sharma, M.; Sun, L.Y.; Tollefsbol, T.O. The Epigenetic Link between Polyphenols, Aging and Age-Related Diseases. Genes 2020, 11, 1094. [Google Scholar] [CrossRef]
- Shirazi-Tehrani, E.; Chamasemani, A.; Firouzabadi, N.; Mousaei, M. NcRNAs and Polyphenols: New Therapeutic Strategies for Hypertension. RNA Biol. 2022, 19, 575–587. [Google Scholar] [CrossRef]
- Scalbert, A.; Morand, C.; Manach, C.; Rémésy, C. Absorption and Metabolism of Polyphenols in the Gut and Impact on Health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef]
- Roblegg, E.; Coughran, A.; Sirjani, D. Saliva: An All-Rounder of Our Body. Eur. J. Pharm. Biopharm. 2019, 142, 133–141. [Google Scholar] [CrossRef]
- Morzel, M.; Canon, F.; Guyot, S. Interactions between Salivary Proteins and Dietary Polyphenols: Potential Consequences on Gastrointestinal Digestive Events. J. Agric. Food Chem. 2022, 70, 6317–6327. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, I.; Kohen, R.; Koren, E. Saliva: A ‘Solubilizer’ of Lipophilic Antioxidant Polyphenols. Oral Dis. 2013, 19, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, H.; Xiao, J. Polyphenols. In Polyphenols: Properties, Recovery, and Applications; Elsevier: Amsterdam, The Netherlands, 2018; pp. 45–67. ISBN 978-0-12-813572-3. [Google Scholar]
- Chen, L.; Teng, H.; Xie, Z.; Cao, H.; Cheang, W.S.; Skalicka-Woniak, K.; Georgiev, M.I.; Xiao, J. Modifications of Dietary Flavonoids towards Improved Bioactivity: An Update on Structure–Activity Relationship. Crit. Rev. Food Sci. Nutr. 2018, 58, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Absorption and Metabolism of Flavonoids. Free Radic. Biol. Med. 2004, 36, 829–837. [Google Scholar] [CrossRef]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; De Filippis, A.; Xiao, H.; Quiles, J.L.; Xiao, J.; et al. Interaction of Dietary Polyphenols and Gut Microbiota: Microbial Metabolism of Polyphenols, Influence on the Gut Microbiota, and Implications on Host Health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.-S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In Vivo and in Vitro Metabolism of Trans-Resveratrol by Human Gut Microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef]
- Cortés-Martín, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, 1900952. [Google Scholar] [CrossRef]
- Stevens, J.F.; Maier, C.S. The Chemistry of Gut Microbial Metabolism of Polyphenols. Phytochem. Rev. 2016, 15, 425–444. [Google Scholar] [CrossRef]
- Selma, M.V.; Tomás-Barberán, F.A.; Romo-Vaquero, M.; Cortés-Martín, A.; Espín, J.C. Understanding Polyphenols’ Health Effects Through the Gut Microbiota. In Dietary Polyphenols; Tomás-Barberán, F.A., González-Sarrías, A., García-Villalba, R., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 497–531. ISBN 978-1-119-56375-4. [Google Scholar]
- Russell, W.R.; Scobbie, L.; Chesson, A.; Richardson, A.J.; Stewart, C.S.; Duncan, S.H.; Drew, J.E.; Duthie, G.G. Anti-Inflammatory Implications of the Microbial Transformation of Dietary Phenolic Compounds. Nutr. Cancer 2008, 60, 636–642. [Google Scholar] [CrossRef]
- Serreli, G.; Deiana, M. In Vivo Formed Metabolites of Polyphenols and Their Biological Efficacy. Food Funct. 2019, 10, 6999–7021. [Google Scholar] [CrossRef]
- Hoda, M.; Hemaiswarya, S.; Doble, M. Pharmacokinetics and Pharmacodynamics of Polyphenols. In Role of Phenolic Phytochemicals in Diabetes Management; Springer: Singapore, 2019; pp. 159–173. ISBN 9789811389962. [Google Scholar]
- Yang, Y.; Trevethan, M.; Wang, S.; Zhao, L. Beneficial Effects of Citrus Flavanones Naringin and Naringenin and Their Food Sources on Lipid Metabolism: An Update on Bioavailability, Pharmacokinetics, and Mechanisms. J. Nutr. Biochem. 2022, 104, 108967. [Google Scholar] [CrossRef] [PubMed]
- Chalet, C.; Rubbens, J.; Tack, J.; Duchateau, G.S.; Augustijns, P. Intestinal Disposition of Quercetin and Its Phase-II Metabolites after Oral Administration in Healthy Volunteers. J. Pharm. Pharmacol. 2018, 70, 1002–1008. [Google Scholar] [CrossRef]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. [Google Scholar] [CrossRef]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, Á.; Halmai, R.; et al. Resveratrol Improves Insulin Sensitivity, Reduces Oxidative Stress and Activates the Akt Pathway in Type 2 Diabetic Patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie Restriction-like Effects of 30 Days of Resveratrol Supplementation on Energy Metabolism and Metabolic Profile in Obese Humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Goh, K.P.; Lee, H.Y.; Lau, D.P.; Supaat, W.; Chan, Y.H.; Koh, A.F.Y. Effects of Resveratrol in Patients with Type 2 Diabetes Mellitus on Skeletal Muscle SIRT1 Expression and Energy Expenditure. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 2–13. [Google Scholar] [CrossRef]
- Timmers, S.; de Ligt, M.; Phielix, E.; van de Weijer, T.; Hansen, J.; Moonen-Kornips, E.; Schaart, G.; Kunz, I.; Hesselink, M.K.C.; Schrauwen-Hinderling, V.B.; et al. Resveratrol as Add-on Therapy in Subjects With Well-Controlled Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2016, 39, 2211–2217. [Google Scholar] [CrossRef] [PubMed]
- Woerdeman, J.; Del Rio, D.; Calani, L.; Eringa, E.C.; Smulders, Y.M.; Serné, E.H. Red Wine Polyphenols Do Not Improve Obesity-associated Insulin Resistance: A Randomized Controlled Trial. Diabetes Obes. Metab. 2018, 20, 206–210. [Google Scholar] [CrossRef]
- Segrestin, B.; Delage, P.; Nemeth, A.; Seyssel, K.; Disse, E.; Nazare, J.-A.; Lambert-Porcheron, S.; Meiller, L.; Sauvinet, V.; Chanon, S.; et al. Polyphenol Supplementation Did Not Affect Insulin Sensitivity and Fat Deposition During One-Month Overfeeding in Randomized Placebo-Controlled Trials in Men and in Women. Front. Nutr. 2022, 9, 854255. [Google Scholar] [CrossRef]
- Alway, S.E.; McCrory, J.L.; Kearcher, K.; Vickers, A.; Frear, B.; Gilleland, D.L.; Bonner, D.E.; Thomas, J.M.; Donley, D.A.; Lively, M.W.; et al. Resveratrol Enhances Exercise-Induced Cellular and Functional Adaptations of Skeletal Muscle in Older Men and Women. J. Gerontol. Ser. A 2017, 72, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.M.; Leeuwenburgh, C.; Guralnik, J.M.; Tian, L.; Sufit, R.; Zhao, L.; Criqui, M.H.; Kibbe, M.R.; Stein, J.H.; Lloyd-Jones, D.; et al. Effect of Resveratrol on Walking Performance in Older People With Peripheral Artery Disease: The RESTORE Randomized Clinical Trial. JAMA Cardiol. 2017, 2, 902. [Google Scholar] [CrossRef]
- Gliemann, L.; Schmidt, J.F.; Olesen, J.; Biensø, R.S.; Peronard, S.L.; Grandjean, S.U.; Mortensen, S.P.; Nyberg, M.; Bangsbo, J.; Pilegaard, H.; et al. Resveratrol Blunts the Positive Effects of Exercise Training on Cardiovascular Health in Aged Men: Adverse Effects of Resveratrol on Cardiovascular Health. J. Physiol. 2013, 591, 5047–5059. [Google Scholar] [CrossRef]
- Olesen, J.; Gliemann, L.; Biensø, R.; Schmidt, J.; Hellsten, Y.; Pilegaard, H. Exercise Training, but Not Resveratrol, Improves Metabolic and Inflammatory Status in Skeletal Muscle of Aged Men: Resveratrol and Exercise Training in Aged Human Subjects. J. Physiol. 2014, 592, 1873–1886. [Google Scholar] [CrossRef]
- Harper, S.A.; Bassler, J.R.; Peramsetty, S.; Yang, Y.; Roberts, L.M.; Drummer, D.; Mankowski, R.T.; Leeuwenburgh, C.; Ricart, K.; Patel, R.P.; et al. Resveratrol and Exercise Combined to Treat Functional Limitations in Late Life: A Pilot Randomized Controlled Trial. Exp. Gerontol. 2021, 143, 111111. [Google Scholar] [CrossRef]
- Buchanan, A.; Villani, A. Association of Adherence to a Mediterranean Diet with Excess Body Mass, Muscle Strength and Physical Performance in Overweight or Obese Adults with or without Type 2 Diabetes: Two Cross-Sectional Studies. Healthcare 2021, 9, 1255. [Google Scholar] [CrossRef] [PubMed]
- De La Cruz Cortés, J.P.; Vallejo-Carmona, L.; Arrebola, M.M.; Martín-Aurioles, E.; Rodriguez-Pérez, M.D.; Ortega-Hombrados, L.; Verdugo, C.; Fernández-Prior, M.Á.; Bermúdez-Oria, A.; González-Correa, J.A. Synergistic Effect of 3′,4′-Dihidroxifenilglicol and Hydroxytyrosol on Oxidative and Nitrosative Stress and Some Cardiovascular Biomarkers in an Experimental Model of Type 1 Diabetes Mellitus. Antioxidants 2021, 10, 1983. [Google Scholar] [CrossRef]
- Kurin, E.; Atanasov, A.; Donath, O.; Heiss, E.; Dirsch, V.; Nagy, M. Synergy Study of the Inhibitory Potential of Red Wine Polyphenols on Vascular Smooth Muscle Cell Proliferation. Planta Med. 2012, 78, 772–778. [Google Scholar] [CrossRef]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef]
- Brat, P.; Mennen, L.; Georgé, S.; Scalbert, A.; Bellamy, A.; Amiot-Carlin, M.-J.; Du Chaffaut, L. Determination of the polyphenol content of fruits and vegetables. Establishment of a database and estimation of the polyphenol intake in the french diet. Acta Hortic. 2007, 744, 61–70. [Google Scholar] [CrossRef]
- Chuang, C.-C.; McIntosh, M.K. Potential Mechanisms by Which Polyphenol-Rich Grapes Prevent Obesity-Mediated Inflammation and Metabolic Diseases. Annu. Rev. Nutr. 2011, 31, 155–176. [Google Scholar] [CrossRef]
- Shahbandeh, M. Statista: Global Production of Fruit by Variety Selected 2020. Available online: https://www.statista.com/statistics/264001/worldwide-production-of-fruit-by-variety (accessed on 2 August 2022).



{kind=link}
{kind=link}
| Source | Family of Polyphenol | Amount | Mean TPC (Folin Assay) |
|---|---|---|---|
| Apple | Anthocyanins | 0.93 mg/100 g FW | 200.96 mg/100 g FW |
| Dihydrochalcones | 5.38 mg/100 g FW | ||
| Flavanols | 24.12 mg/100 g FW | ||
| Flavonols | 6.86 mg/100 g FW | ||
| Phenolic acids | 19 mg/100 g FW | ||
| Artichoke, heads, raw | Flavones | 57.8 mg/100 g FW | 1142.40 mg/100 g FW |
| Phenolic acids | 202.23 mg/100 g FW | ||
| Blueberries | Flavonols | 12.23 mg/100 g FW | 151.33 mg/100 g FW |
| Phenolic acids | 162.47 mg/100 g FW | ||
| Phenolic acids | 37.06 mg/100 g FW | ||
| Other polyphenols | 0.45527 mg/100 g FW | ||
| Cocoa, powder | Flavanols | 511.62 mg/100 g FW | 5624.23 mg/100 g FW |
| Phenolic acids | 37.06 mg/100 g FW | ||
| Other polyphenols | 0.45527 mg/100 g FW | ||
| Grape | Anthocyanins | 72.1 mg/100 g FW | 184.97 mg/100 g FW |
| Flavanols | 17.11 mg/100 g FW | ||
| Flavonols | 3.08 mg/100 g FW | ||
| Phenolic acids | 1.69 mg/100 g FW | ||
| Stilbenes | 0.3362 mg/100 g FW | ||
| Green tea | Flavanols | 71.18 mg/100 g FW | 61.86 mg/100 ml |
| Flavonols | 5.29 mg/100 g FW | ||
| Phenolic acids | 12.53 mg/100 g FW | ||
| Olives, green | Flavones | 0.56 mg/100 g FW | 161.24 mg/100 g FW |
| Phenolic acids | 134.94 mg/100 g FW | ||
| Other polyphenols | 211.05 mg/100 g FW | ||
| Persil, fresh | Other polyphenols | 13.95 mg/100 g FW | 89.27 mg/100 g FW |
| Strawberries | Anthocyanins | 73.01 mg/100 g FW | 289.20 mg/100 g FW |
| Flavanols | 9.1375 mg/100 g FW | ||
| Flavonols | 2.32 mg/100 g FW | ||
| Phenolic acids | 10.74 mg/100 g FW | ||
| Stilbenes | 0.35 mg/100 g FW |
| Family and Subfamily | Compound | Effect and Mechanism | References |
|---|---|---|---|
| Flavonoids/Flavan-3-ols | EGCG | ● Antioxidant Radical scavenging Metal ion chelation ↑ CAT, ↑ SOD1 e SOD2, ↑ GPx ● Anti-inflammatory ↓ NFĸB via ↓ Iĸβ ↓ COX-2 ↓ IRF3 via ↓ TBK1 ● Anti-diabetic ↓ insulin resistance ↑ lipid oxidation in muscle ↑ NFĸB, ↑ AMPK, ↑ MAPK ● Anti-aging/pro-apoptotic ↑ Beclin-1 and ↑ caspases | ● Fraga et al. [85] Bernatoniene et al. [86] Meng et al. [57] ● Youn et al. [87] ● Casanova et al. [88] Li et al. [89] ● Pallauf and Rimbach [90] |
| Grape seed proanthocyanidins | ● Anti-diabetic ↑ Nrf1, ↑SIRT1, and ↑PGC-1α, ↑ slow myosin heavy chain, ↑ succinic dehydrogenase and malate dehydrogenase activities, ↑ resistance to fatigue | ● Xu et al. [91] | |
| Flavonoids/Flavanols | Quercetin | ● Anti-inflammatory ↑ Nrf2/ARE pathways ↑ Antioxidant enzymes ↓ TNF-α, ↓ IL-6, ↓ IL-1β, ↓ COX-2, ↓ iNOS, ↓ NFĸB in adipocytes and macrophages | ● Costa et al. [92] ● Sato et al. [93] |
| Myricetin | ● Anti-diabetic ↑ glucose uptake, ↓ insulin resistance, ↑Akt and ↑AMPK signaling pathways | ● Pandey et al. [94] | |
| Kaempferol | ● Anti-inflammatory ↓ IL-6, IL-1β, 18 and TNF-α ↑ Nrf2 and synthesis targets Inhibition TLR4 | ● Alam et al. [95] | |
| Flavonoids/Anthocyanes | Anthocyanins | ● Anti-inflammatory ↓ COX-1 and COX-2 ↓ C-reactive protein | ● Mozos et al. [96] ● Sivamaruthi et al. [97] |
| Cyanidin-3-O-glucoside | ● Antidyslipidemic ↑ PPARs ● Anti-diabetic ↑ Insulin sensitivity → ↑ PPARs ↑ Insulin secretion → ↓ IL-1β and IL-6 ↑ TLR4/IĸBα pathway | ● Jia et al. [98] ● Geng et al. [99] | |
| Flavonoids/Flavones | Luteolin/Apigenin | ● Anti-inflammatory ↓ NO and ↓ PGE2 | ● Tian et al. [100] |
| Apigenin | ● Anti-obesity Radical scavenger ↑ Increase muscle fibers size ↑ number and volume mitochondria ↑ SOD and GPx | ● Wang et al. [101] | |
| Flavonoids/Isoflavones | Daidzein | ● Anti-diabetic Inhibition α-amylase and α-glycosidase ↑ AMPK, ↑ GK, ↓ G6Pase, ↓ PEPCK, ↑ GLUT4, ↑ IRS1, ↑ IRS2, ↑ PPARγ ● Anti-inflammatory ↑ PPARγ, ↓ TNFα, ↓ NFĸB, ↓ IL-6, ↓ Ccl2, ↓ Cxcl2 | ● Park et al. [102] ● Das et al. [103] |
| Genistein | ● Anti-diabetic ↑ AMPK in skeletal muscle ↑ insulin sensitivity ↑ lipid oxidation | ● Guevara-Cruz et al. [104] | |
| Flavonoids/Flavanones | Naringenin | ● Anti-diabetic ↑ Insulin secretion ● Anti-inflammatory ↓ TNF-α and IL-6 ↑ SOD | ● Rehman et al. [105] |
| Hesperidin | ● Anti-diabetic ↑ IRS, ↑ Akt, and ↑ GLUT4 in muscle cells | ● Dhanya et al. [106] | |
| Stilbenes | Resveratrol | ● Anti-diabetic ↑ SIRT1 and ↑ PGC-1α ↑ mitochondrial activity (exercise mimetic effect) ↑ Akt and AMPK pathways → ↑ insulin sensitivity ● Anti-obesity ↓ fat accumulation ↑ lipolysis ● Anti-aging ↓ caspase 3 | ● Lagouge et al. [107] ● Lagouge et al. [107] Kang et al. [108] ● Huang et al. [109] ● Bai et al. [110] |
| Phenolic acids | Phenolic acids | ● Anti-diabetic ↑ GLUT2 in pancreatic β-cells ↑ PI3K/Akt and ↑ GLUT4 in adipose and muscle tissues ↓ α-glucosidase activity | ● Kumar et al. [111] ● Duboit et al. [96] |
| Gallic acid/p-coumaric acid | ● Anti-diabetic and anti-obesity ↓ TNF-α and ↓ PPAR γ in adipose tissue | ● Abdel-Moneim et al. [112] | |
| Caffeic acid phenetyl ester | ● Anti-inflammatory ↓ COX and ↓ LOX Inhibition detachment arachidonic acid. | ● Silva et al. [113] | |
| Vanillic acid | ● Anti-obesity ↓ PPAR and C/EBPα ↑ Lipid oxidation through ↑ AMPKα | ● Jung et al. [114] | |
| Syringic acid | ● Anti-diabetic ↑ PGC-1α and Nrf2 ↑ increased mitochondrial biogenesis. ↓ TNF-α, IL-1β, and IL-6 | ● Rashedinina et al. [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capozzi, A.; Saucier, C.; Bisbal, C.; Lambert, K. Grape Polyphenols in the Treatment of Human Skeletal Muscle Damage Due to Inflammation and Oxidative Stress during Obesity and Aging: Early Outcomes and Promises. Molecules 2022, 27, 6594. https://doi.org/10.3390/molecules27196594
Capozzi A, Saucier C, Bisbal C, Lambert K. Grape Polyphenols in the Treatment of Human Skeletal Muscle Damage Due to Inflammation and Oxidative Stress during Obesity and Aging: Early Outcomes and Promises. Molecules. 2022; 27(19):6594. https://doi.org/10.3390/molecules27196594
Chicago/Turabian StyleCapozzi, Adriana, Cédric Saucier, Catherine Bisbal, and Karen Lambert. 2022. "Grape Polyphenols in the Treatment of Human Skeletal Muscle Damage Due to Inflammation and Oxidative Stress during Obesity and Aging: Early Outcomes and Promises" Molecules 27, no. 19: 6594. https://doi.org/10.3390/molecules27196594
APA StyleCapozzi, A., Saucier, C., Bisbal, C., & Lambert, K. (2022). Grape Polyphenols in the Treatment of Human Skeletal Muscle Damage Due to Inflammation and Oxidative Stress during Obesity and Aging: Early Outcomes and Promises. Molecules, 27(19), 6594. https://doi.org/10.3390/molecules27196594