


Neuroprotective Effect of Polyphenol Extracts from Terminalia chebula Retz. against Cerebral Ischemia-Reperfusion Injury

 ,
, 
Abstract
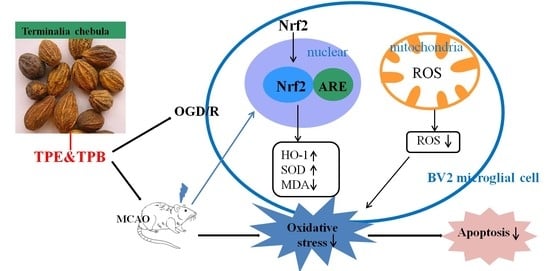
1. Introduction
2. Results
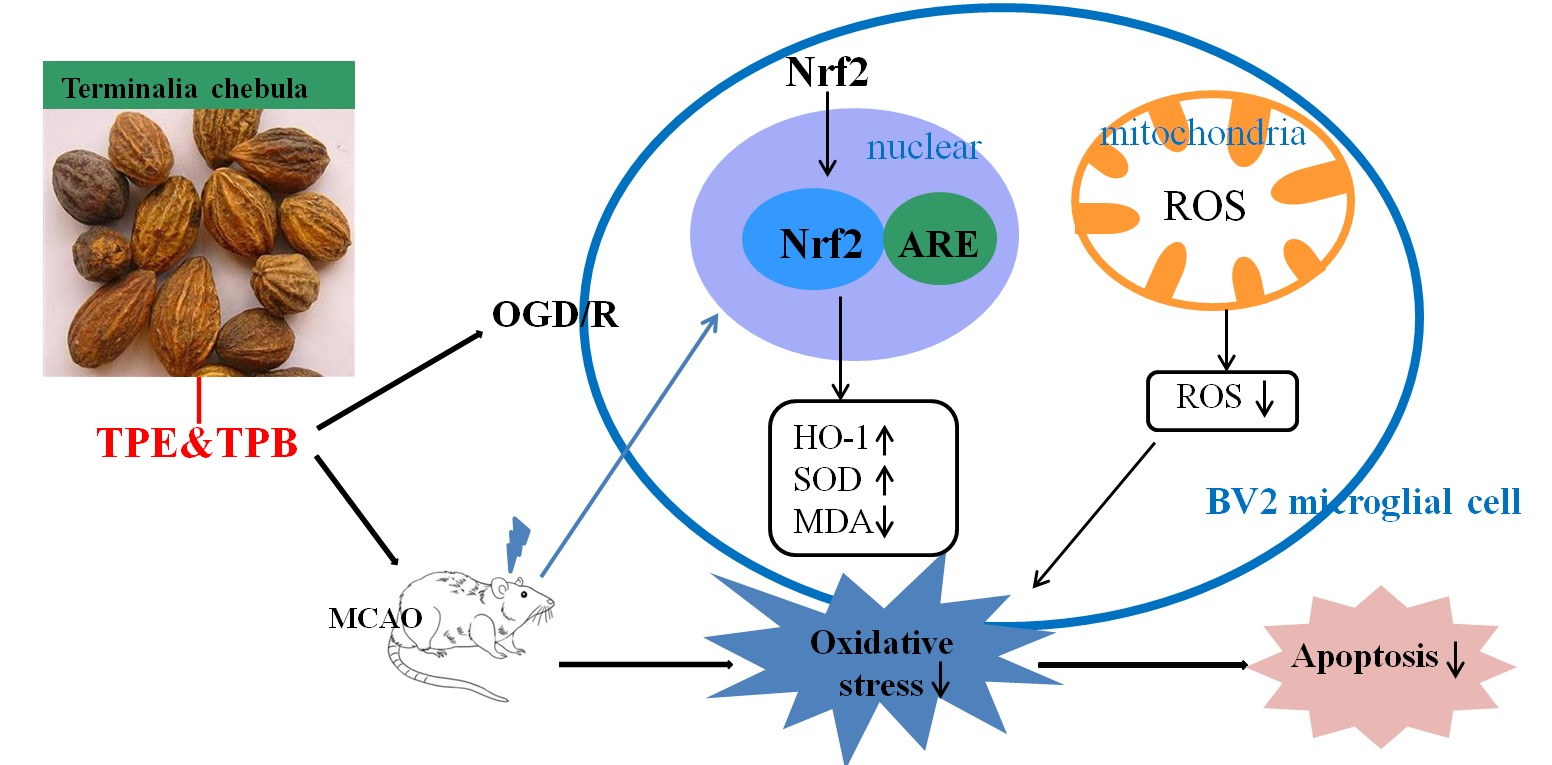
2.1. Different Fractions from T. chebula Extract and Antioxidant Activities
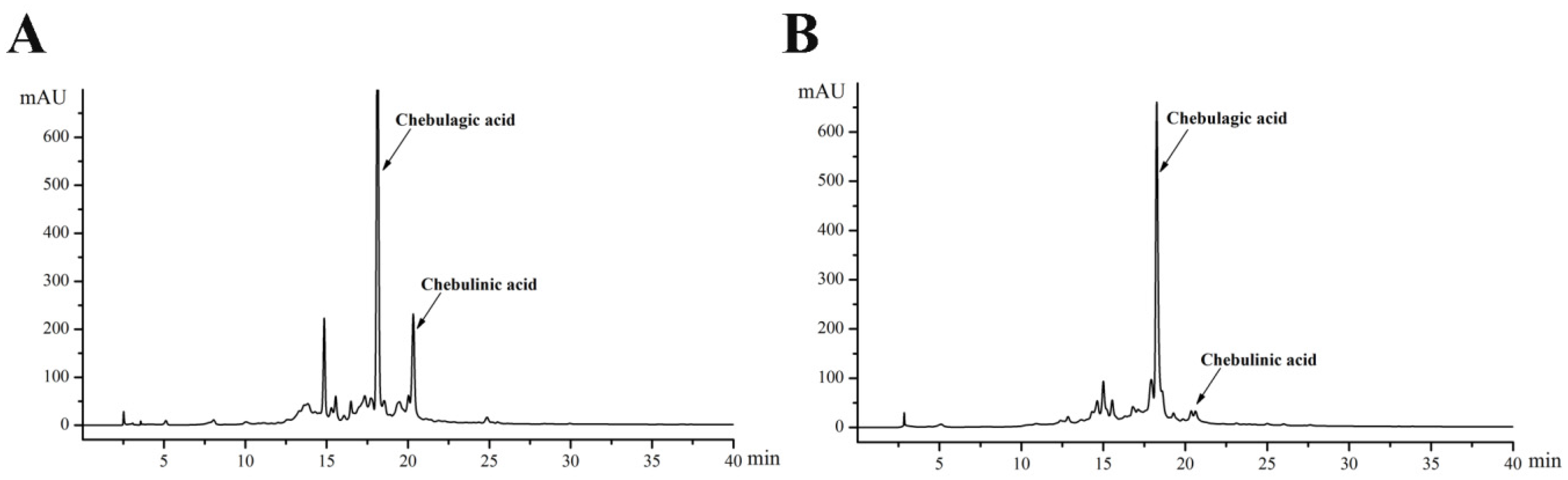
2.2. Effects of TPE and TPB on the Viability of Cells
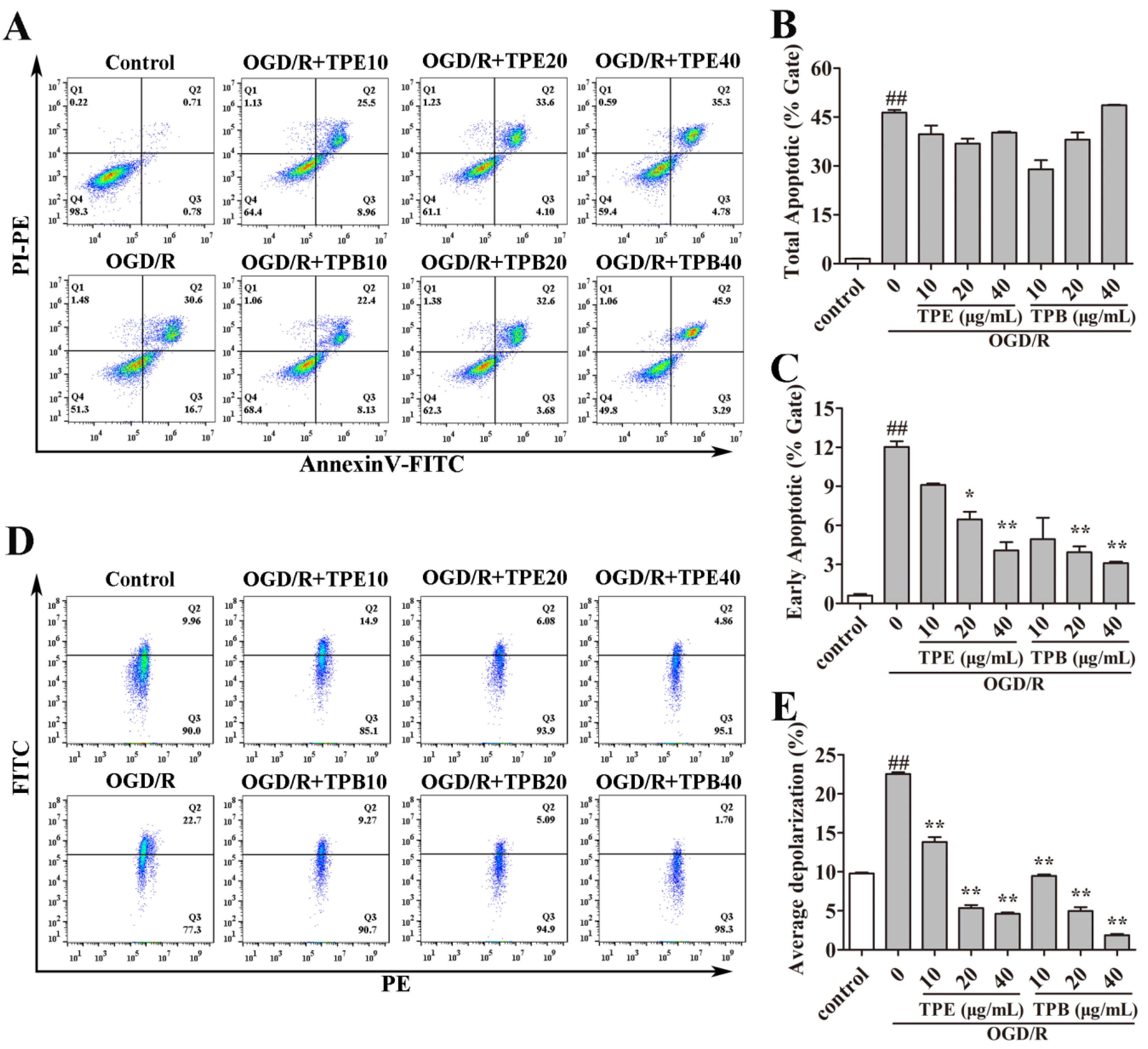
2.3. TPE and TPB Ameliorate OGD/R Injury by Reducing Cell Apoptosis
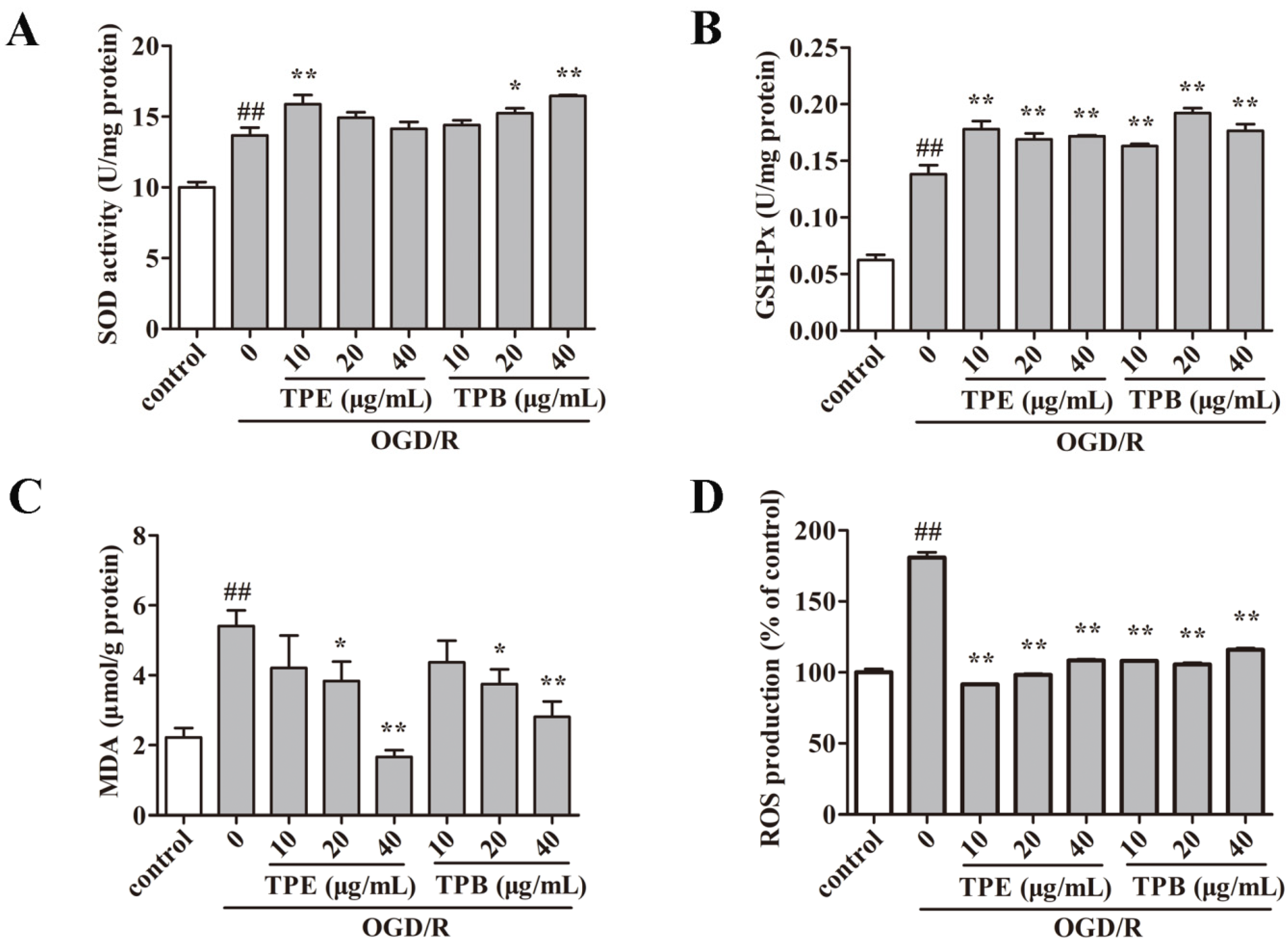
2.4. Enzyme Activities Related to Oxidative Stress
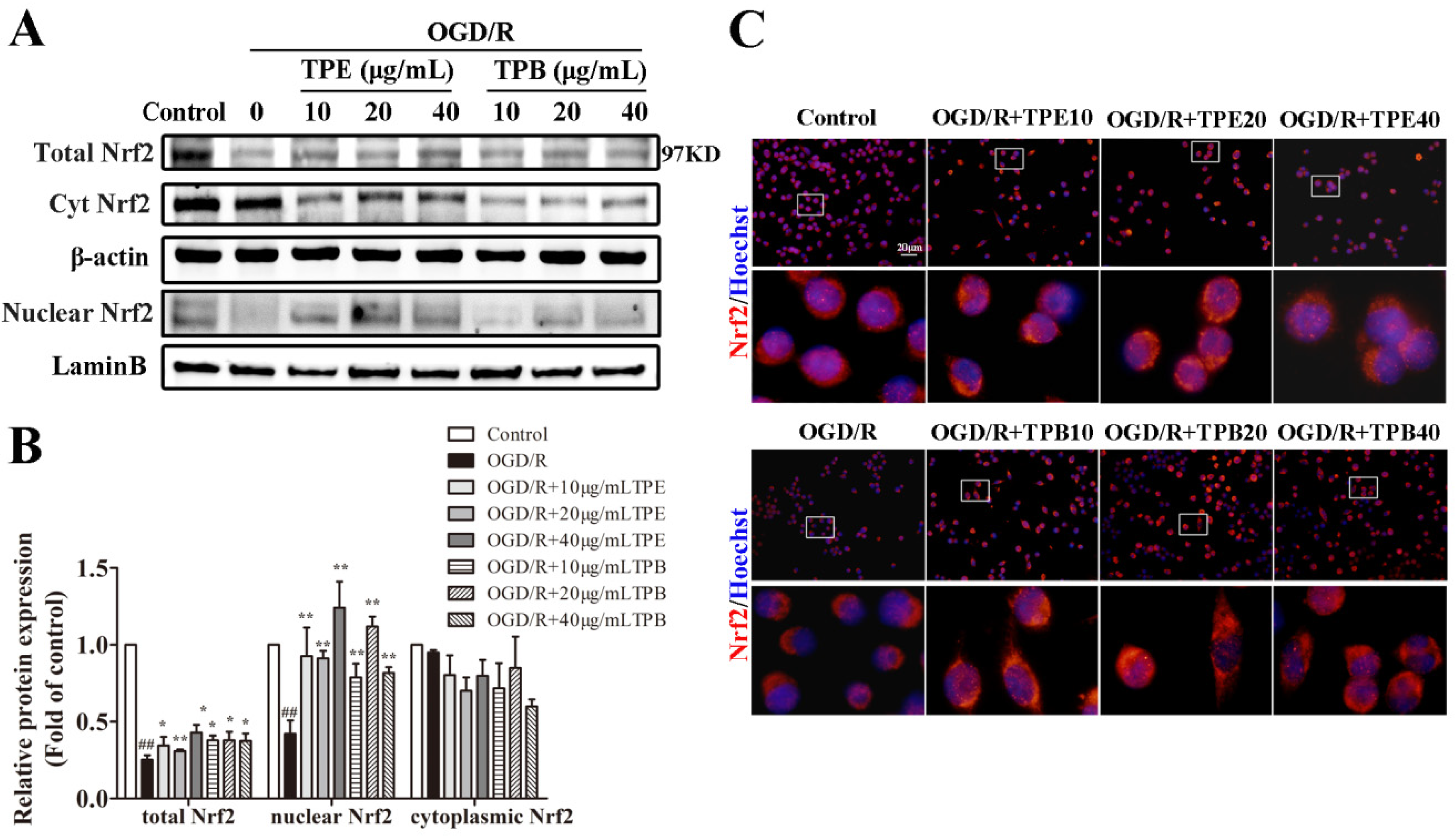
2.5. TPE and TPB Promotes the Nuclear Translocation of Nrf2 in BV2 Cells
2.6. TPE and TPB Stimulate the Nrf2/HO-1 Signaling Pathway in BV2 Cells
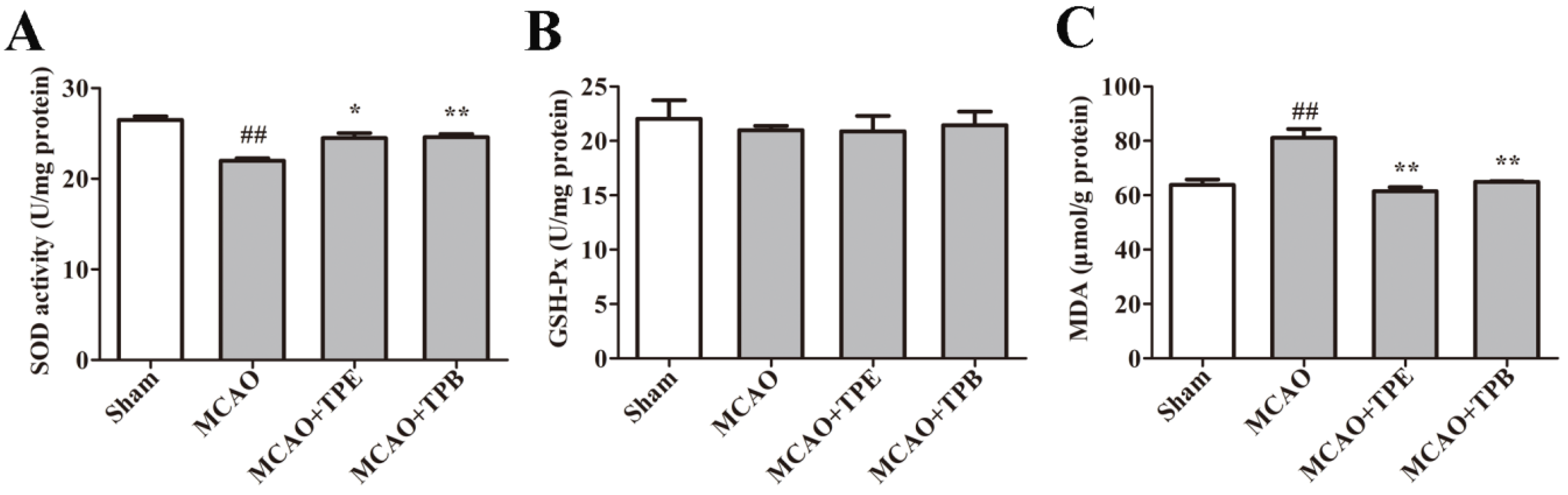
2.7. TPE and TPB Improve Enzyme Activities Related to Oxidative Stress in MCAO Mice
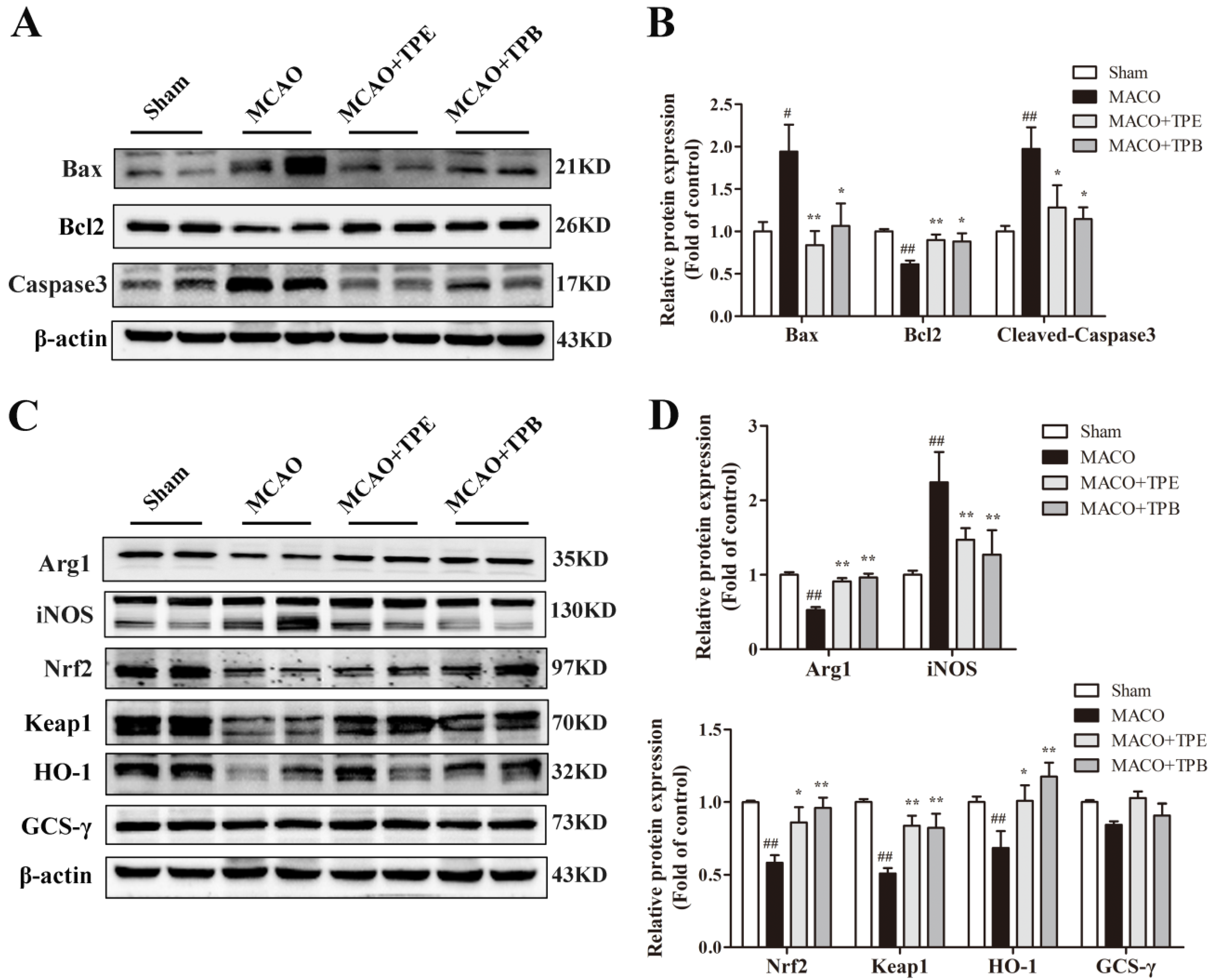
2.8. TPE and TPB Protect Brain from MCAO Injury by Reducing Apoptosis and Stimulating the Nrf2/HO-1 Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material and Preparation of Extracts
4.3. Main Compounds and Antioxidant Activity In Vitro
4.4. Cell Culture
4.5. Measurement of Cell Viability and Enzyme Activities
4.6. Detection of Apoptotic Ratio and Mitochondrial Membrane Potential
4.7. Middle Cerebral Artery Occlusion (MCAO) Model and Drug Treatment
4.8. Western Blotting
4.9. Immunofluorescence Staining
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Lapchak, P.A.; Zhang, J.H. The high cost of stroke and stroke cytoprotection research. Transl. Stroke Res. 2017, 8, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.G.; Mohney, R.P.; Wilson, B.; Jeohn, G.H.; Liu, B.; Hong, J.S. Regional difference in susceptibility to lipopolysaccharide-induced neurotoxicity in the rat brain: Role of microglia. J. Neurosci. 2000, 20, 6309–6316. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ju, F.; Xie, W.; Hafeez, M.T.; Cheng, X.; Yang, Z.; Zhu, L.R.; Li, T.; Zhang, S. Transcriptomic analysis reveals differential activation of microglial genes after ischemic stroke in mice. Neuroscience 2017, 348, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Nie, D.; Wang, L.J.; Qiu, H.C.; Ma, L.; Dong, M.X.; Tu, W.J.; Zhao, J. Microglial polarization: Novel therapeutic strategy against ischemic stroke. Aging Dis. 2021, 12, 466–479. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Ye, X.; Hao, Q.; Zhang, T.; Cui, G.; Yu, M. Nrf2/ARE pathway inhibits ROS-induced NLRP3 inflammasome activation in BV2 cells after cerebral ischemia reperfusion. Inflamm. Res. 2018, 67, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Kelmanson, I.V.; Shokhina, A.G.; Kotova, D.A.; Pochechuev, M.S.; Ivanova, A.D.; Kostyuk, A.I.; Panova, A.S.; Borodinova, A.A.; Solotenkov, M.A.; Stepanov, E.A.; et al. In vivo dynamics of acidosis and oxidative stress in the acute phase of an ischemic stroke in a rodent model. Redox Biol. 2021, 48, 102178. [Google Scholar] [CrossRef]
- Wen, B.; Xu, K.; Huang, R.; Jiang, T.; Wang, J.; Chen, J.; Chen, J.; He, B. Preserving mitochondrial function by inhibiting GRP75 ameliorates neuron injury under ischemic stroke. Mol. Med. Rep. 2022, 25, 165. [Google Scholar] [CrossRef]
- Hu, Y.; Zheng, Y.; Wang, T.; Jiao, L.; Luo, Y. VEGF, a key factor for blood brain barrier injury after cerebral ischemic stroke. Aging Dis. 2022, 13, 647–654. [Google Scholar] [CrossRef]
- Rodrigo, R.; Fernandez-Gajardo, R.; Gutierrez, R.; Matamala, J.M.; Carrasco, R.; Merchak, A.M.; Feuerhake, W. Oxidative stress and pathophysiology of ischemic stroke: Novel therapeutic opportunities. CNS Neurol. Disord. Drug Targets 2013, 12, 698–714. [Google Scholar] [CrossRef]
- Jurcau, A.; Ardelean, A.I. Oxidative stress in ischemia/reperfusion injuries following acute ischemic stroke. Biomedicines 2022, 10, 574. [Google Scholar] [CrossRef]
- Wei, N.; Pu, Y.; Yang, Z.; Pan, Y.; Liu, L. Therapeutic effects of melatonin on cerebral ischemia reperfusion injury: Role of Yap-OPA1 signaling pathway and mitochondrial fusion. Biomed. Pharmacother. 2019, 110, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhu, P.; Wang, J.; Zhu, H.; Ren, J.; Chen, Y. Pathogenesis of cardiac ischemia reperfusion injury is associated with CK2α-disturbed mitochondrial homeostasis via suppression of FUNDC1-related mitophagy. Cell Death Differ. 2018, 25, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Zong, N.; Hu, Y.; Chen, Y.; Xu, Y. Neuronal death mechanisms and therapeutic strategy in ischemic stroke. Neurosci. Bull. 2022, 1–19. [Google Scholar] [CrossRef]
- Gao, Y.; Zhuang, Z.; Gao, S.; Li, X.; Zhang, Z.; Ye, Z.; Li, L.; Tang, C.; Zhou, M.; Han, X.; et al. Tetrahydrocurcumin reduces oxidative stress-induced apoptosis via the mitochondrial apoptotic pathway by modulating autophagy in rats after traumatic brain injury. Am. J. Transl. Res. 2017, 9, 887–899. [Google Scholar]
- Uzdensky, A.B. Apoptosis regulation in the penumbra after ischemic stroke: Expression of pro- and antiapoptotic proteins. Apoptosis 2019, 24, 687–702. [Google Scholar] [CrossRef]
- Farina, M.; Vieira, L.E.; Buttari, B.; Profumo, E.; Saso, L. The Nrf2 Pathway in Ischemic Stroke: A Review. Molecules 2021, 26, 5001. [Google Scholar] [CrossRef] [PubMed]
- Dordoe, C.; Wang, X.; Lin, P.; Wang, Z.; Hu, J.; Wang, D.; Fang, Y.; Liang, F.; Ye, S.; Chen, J.; et al. Non-mitogenic fibroblast growth factor 1 protects against ischemic stroke by regulating microglia/macrophage polarization through Nrf2 and NF-κB pathways. Neuropharmacology 2022, 212, 109064. [Google Scholar] [CrossRef]
- Rana, A.K.; Sharma, S.; Saini, S.K.; Singh, D. Rutin protects hemorrhagic stroke development via supressing oxidative stress and inflammatory events in a zebrafish model. Eur. J. Pharmacol. 2022, 925, 174973. [Google Scholar] [CrossRef]
- Imai, T.; Matsubara, H.; Hara, H. Potential therapeutic effects of Nrf2 activators on intracranial hemorrhage. J. Cereb. Blood Flow Metab. 2021, 41, 1483–1500. [Google Scholar] [CrossRef]
- Bag, A.; Bhattacharyya, S.K.; Chattopadhyay, R.R. The development of Terminalia chebula Retz. (Combretaceae) in clinical research. Asian Pac. J. Trop. Biomed. 2013, 3, 244–252. [Google Scholar] [CrossRef]
- Jin, F.; Cheng, D.; Tao, J.Y.; Zhang, S.L.; Pang, R.; Guo, Y.J.; Ye, P.; Dong, J.H.; Zhao, L. Anti-inflammatory and anti-oxidative effects of corilagin in a rat model of acute cholestasis. BMC Gastroenterol. 2013, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Youn, K.; Lee, S.; Jeong, W.S.; Ho, C.T.; Jun, M. Protective role of corilagin on Aβ 25–35-Induced neurotoxicity: Suppression of NF-κB signaling pathway. J. Med. Food 2016, 19, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, Y.; Li, G.; Xue, J.; Chen, Z.L.; Jin, F.; Luo, L.; Zhou, X.; Ma, Q.; Cai, X.; et al. Effects of corilagin on alleviating cholestasis via farnesoid X receptor-associated pathways in vitro and in vivo. Br. J. Pharmacol. 2018, 175, 810–829. [Google Scholar] [CrossRef] [PubMed]
- Kher, M.N.; Sheth, N.R.; Bhatt, V.D. In vitro antibacterial evaluation of Terminalia chebula as an alternative of antibiotics against bovine subclinical mastitis. Anim. Biotechnol. 2019, 30, 151–158. [Google Scholar] [CrossRef]
- Nigam, M.; Mishra, A.P.; Devkota, A.A.; Dirar, A.I.; Hassan, M.M.; Adhikari, A.; Belwal, T.; Devkota, H.P. Fruits of Terminalia chebula Retz.: A review on traditional uses, bioactive chemical constituents and pharmacological activities. Phytother. Res. 2020, 10, 2518–2533. [Google Scholar] [CrossRef]
- Zhang, X.R.; Qiao, Y.J.; Zhu, H.T.; Kong, Q.H.; Wang, D.; Yang, C.R.; Zhang, Y.J. Multiple in vitro biological effects of phenolic compounds from Terminalia chebula var. tomentella. J. Ethnopharmacol. 2021, 275, 114135. [Google Scholar] [CrossRef]
- Park, J.H.; Joo, H.S.; Yoo, K.Y.; Shin, B.N.; Kim, I.H.; Lee, C.H.; Choi, J.H.; Byun, K.; Lee, B.; Lim, S.S.; et al. Extract from Terminalia chebula seeds protect against experimental ischemic neuronal damage via maintaining SODs and BDNF levels. Neurochem. Res. 2011, 36, 2043–2050. [Google Scholar] [CrossRef]
- Zhao, L.H.; Duan, Z.Y.; Wang, Y.; Wang, M.Z.; Liu, Y.; Wang, X.; Li, H.Y. Protective effect of Terminalia chebula Retz. extract against Aβ aggregation and Aβ-induced toxicity in Caenorhabditis elegans. J. Ethnopharmacol. 2021, 268, 113640. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, D.Y.; Lee, J.; Kim, H.W.; Sung, S.H.; Han, J.S.; Jeon, W.K. Terminalia chebula extract prevents scopolamine-induced amnesia via cholinergic modulation and anti-oxidative effects in mice. BMC Complement. Altern. Med. 2018, 18, 36. [Google Scholar] [CrossRef]
- Dai, B.; Yan, T.; Shen, Y.X.; Xu, Y.J.; Shen, H.B.; Chen, D.; Wang, J.R.; He, S.H.; Dong, Q.R.; Zhang, A.L. Edaravone protects against oxygen-glucose-serum deprivation/restoration-induced apoptosis in spinal cord astrocytes by inhibiting integrated stress response. Neural Regen. Res. 2017, 12, 283–289. [Google Scholar] [CrossRef]
- Chen, B.; Zhao, J.; Zhang, R.; Zhang, L.; Zhang, Q.; Yang, H.; An, J. Neuroprotective effects of natural compounds on neurotoxin-induced oxidative stress and cell apoptosis. Nutr. Neurosci. 2020, 25, 1078–1099. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Qin, Y.; Li, D.; Cai, N.; Wu, J.; Jiang, L.; Jie, L.; Zhou, Z.; Xu, J.; Wang, H. Inhibition of PDE4 protects neurons against oxygen-glucose deprivation induced endoplasmic reticulum stress through activation of the Nrf-2/HO-1 pathway. Redox Biol. 2020, 28, 101342. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, S.; Fan, X. Role of Polyphenols as Antioxidant Supplementation in Ischemic Stroke. Oxid. Med. Cell. Longev. 2021, 2021, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gasparovic, A.C.; Zarkovic, N.; Bottari, S.P. Biomarkers of nitro-oxidation and oxidative stress. Curr. Opin. Toxicol. 2017, 7, 73–80. [Google Scholar] [CrossRef]
- Kovac, S.; Angelova, P.R.; Holmström, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 regulates ROS production by mitochondria and NADPH oxidase. Biochim. Biophys. Acta 2015, 1850, 794–801. [Google Scholar] [CrossRef]
- Wang, W.; Yang, L.; Liu, T.; Ma, Y.; Huang, S.; He, M.; Wang, J.; Wen, A.; Ding, Y. Corilagin ameliorates sleep deprivation-induced memory impairments by inhibiting NOX2 and activating Nrf2. Brain Res. Bull. 2020, 160, 141–149. [Google Scholar] [CrossRef]
- Li, J.; Zhao, T.; Qiao, H.; Li, Y.; Xia, M.; Wang, X.; Liu, C.; Zheng, T.; Chen, R.; Xie, Y.; et al. Research progress of natural products for the treatment of ischemic stroke. J. Integr. Neurosci. 2022, 21, 14. [Google Scholar] [CrossRef]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Lee, H.S.; Won, N.H.; Kim, K.H.; Lee, H.; Jun, W.; Lee, K.W. Antioxidant effects of aqueous extract of Terminalia chebula in vivo and in vitro. Biol. Pharm. Bull. 2005, 28, 1639–1644. [Google Scholar] [CrossRef]
- Chandrasekhar, Y.; Kumar, G.P.; Navya, K.; Ramya, E.M.; Anilakumar, K.R. Tannins from Terminalia chebula fruits attenuates GABA antagonist-induced anxiety-like behaviour via modulation of neurotransmitters. J. Pharm. Pharmacol. 2018, 70, 1662–1674. [Google Scholar] [CrossRef]
- Prada, A.L.; Amado, J.R.R.; Keita, H.; Zapata, E.P.; Carvalho, H.; Lima, E.S.; Sousa, T.P.; Carvalho, J.C.T. Cassia grandis fruit extract reduces the blood glucose level in alloxan-induced diabetic rats. Biomed. Pharmacother. 2018, 103, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lin, K.; Zhou, M.; Xiong, Q.; Li, C.; Ru, Q. Polysaccharides from Opuntia milpa alta alleviate alloxan-induced INS-1 cells apoptosis via reducing oxidative stress and upregulating Nrf2 expression. Nutr. Res. 2020, 77, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Lin, P.; Chen, M.; Zhang, Y.; Chen, J.; Zheng, M.; Liu, J.; Du, H.; Chen, R.; Pan, X.; et al. Restoration of L-OPA1 alleviates acute ischemic stroke injury in rats via inhibiting neuronal apoptosis and preserving mitochondrial function. Redox Biol. 2020, 34, 101503. [Google Scholar] [CrossRef] [PubMed]
- Song, X.R.; Gong, Z.X.; Liu, K.L.; Kou, J.P.; Liu, B.L.; Liu, K. Baicalin combats glutamate excitotoxicity via protecting glutamine synthetase from ROS-induced 20S proteasomal degradation. Redox Biol. 2020, 34, 101559. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.X.; Zhang, L.Y.; Chen, Y.L.; Yu, S.S.; Zhao, Y.; Zhao, J. Curcumin pretreatment and post-treatment both improve the antioxidative ability of neurons with oxygen-glucose deprivation. Neural Regen. Res. 2015, 10, 481–489. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | IC50 (μ g/mL) | ||
|---|---|---|---|
| DPPH• | ABTS•+ | OH• | |
| TPE | 3.15 ± 0.02 ** | 58.71 ± 1.88 ** | 36.79 ± 2.44 ** |
| TPB | 3.48 ± 0.16 ** | 76.77 ± 1.67 | 253.16 ± 19.37 ** |
| VC | 5.52 ± 0.18 | 76.36 ± 2.33 | 2904 ± 97.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, K.; Zhou, M.; Leng, C.; Tao, X.; Zhou, R.; Li, Y.; Sun, B.; Shu, X.; Liu, W. Neuroprotective Effect of Polyphenol Extracts from Terminalia chebula Retz. against Cerebral Ischemia-Reperfusion Injury. Molecules 2022, 27, 6449. https://doi.org/10.3390/molecules27196449
Lin K, Zhou M, Leng C, Tao X, Zhou R, Li Y, Sun B, Shu X, Liu W. Neuroprotective Effect of Polyphenol Extracts from Terminalia chebula Retz. against Cerebral Ischemia-Reperfusion Injury. Molecules. 2022; 27(19):6449. https://doi.org/10.3390/molecules27196449
Chicago/Turabian StyleLin, Kuan, Mei Zhou, Changlong Leng, Xiaoqing Tao, Rong Zhou, Youwei Li, Binlian Sun, Xiji Shu, and Wei Liu. 2022. "Neuroprotective Effect of Polyphenol Extracts from Terminalia chebula Retz. against Cerebral Ischemia-Reperfusion Injury" Molecules 27, no. 19: 6449. https://doi.org/10.3390/molecules27196449
APA StyleLin, K., Zhou, M., Leng, C., Tao, X., Zhou, R., Li, Y., Sun, B., Shu, X., & Liu, W. (2022). Neuroprotective Effect of Polyphenol Extracts from Terminalia chebula Retz. against Cerebral Ischemia-Reperfusion Injury. Molecules, 27(19), 6449. https://doi.org/10.3390/molecules27196449