
Proanthocyanidins Modulate Rumen Enzyme Activities and Protein Utilization In Vitro

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Extraction and Molecular Characterization of Proanthocyanidins
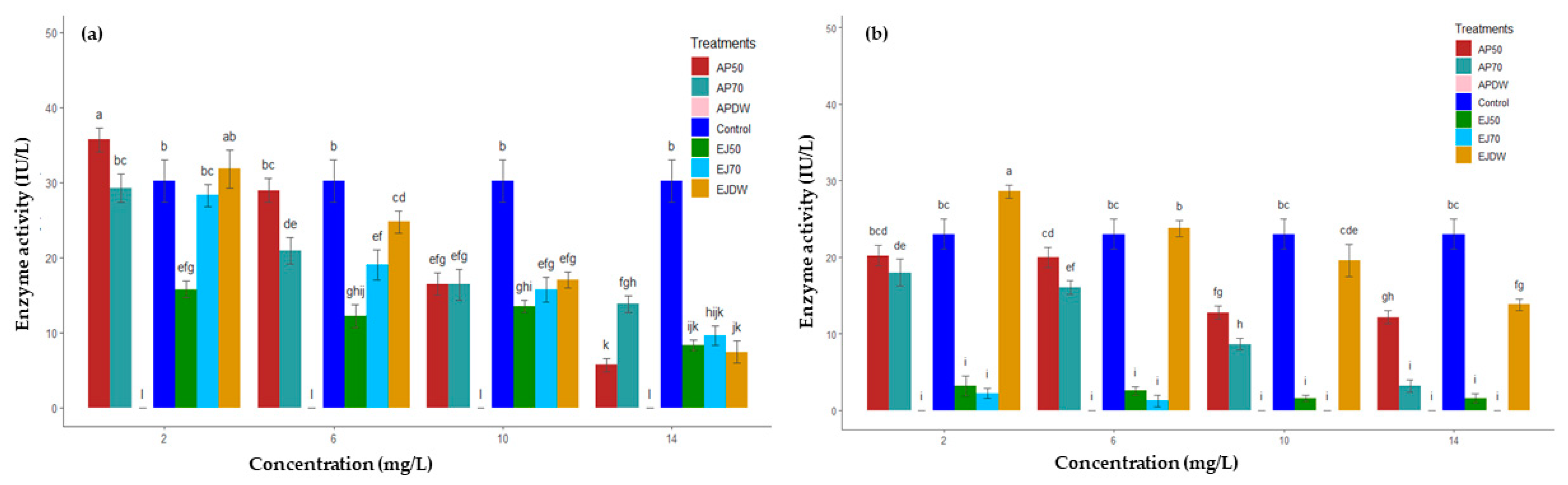
2.2. Effect on Rumen Enzymes Activities
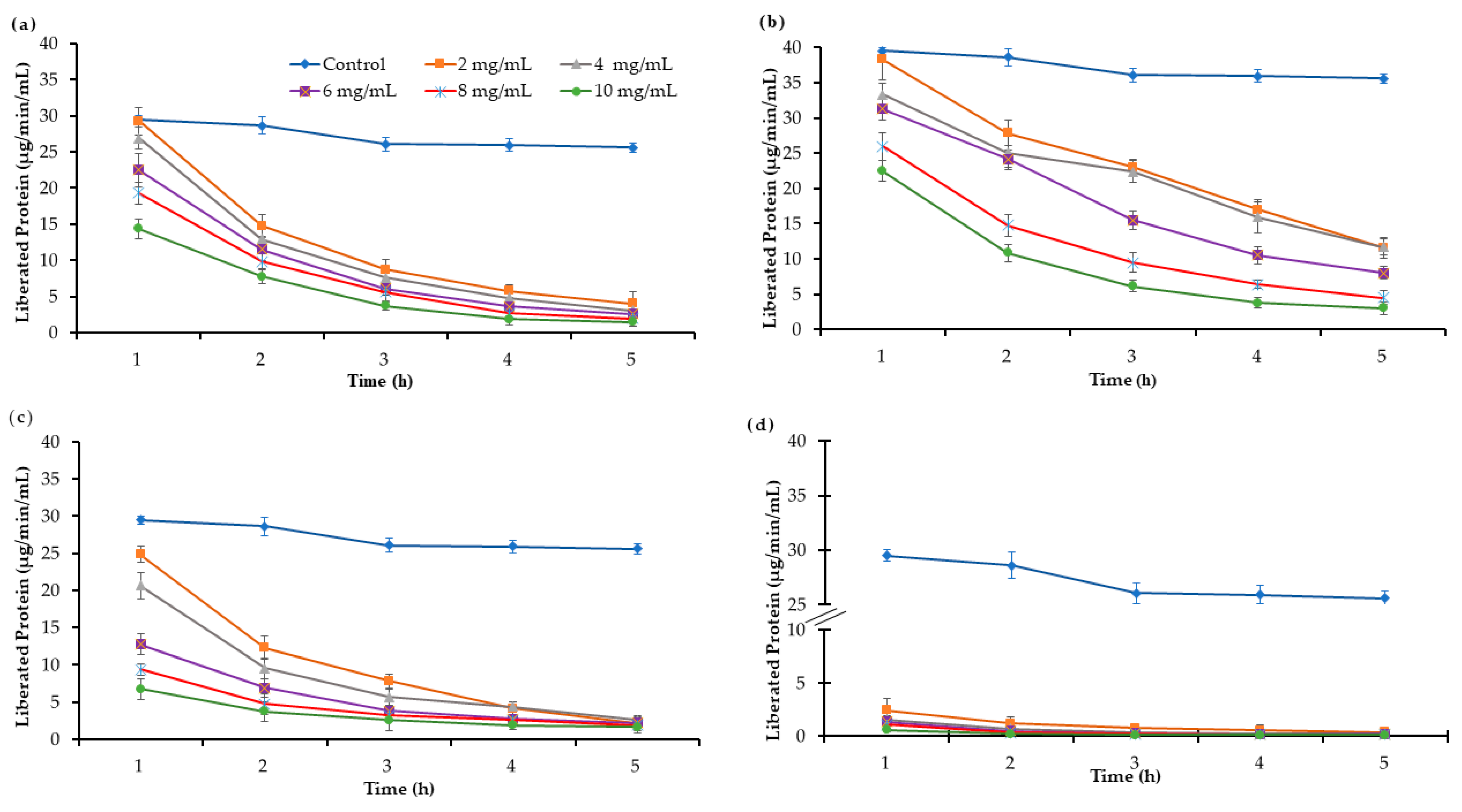
2.3. Effect on Protein Solubilization and Microbial Protein Efficiency
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Collection of Plant Material and Extraction of Proanthocyanidins (PAs)
4.3. Ruminal Microbial Enzyme Activity
4.4. Rubisco Solubilization and Microbial Protein Synthesis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, D.; Baghel, U.S.; Pannu, M.S.; Yadav, R. Ethnopharmacological based Evaluation of Anogeissus pendula Edgew Extracts for Antioxidant and Hepatoprotective Potential. Anci. Sci. Life 2017, 36, 136–140. [Google Scholar]
- Xu, J.; Liu, T.; Li, Y.; Liu, W.; Ding, Z.; Ma, H.; Seeram, N.P.; Mu, Y.; Huang, X.; Li, L. Jamun (Eugenia jambolana lam.) fruit extract prevents obesity by modulating the gut microbiome in high-fat-diet-fed mice. Mol. Nutr. Food Res. 2019, 63, 1801307. [Google Scholar] [CrossRef]
- Abbassy, M.; Salem, M.Z.M.; Rashad, N.M.; Afify, S.M.; Salem, A.Z.M. Nutritive and biocidal properties of agroforestry trees of Moringa oleifera Lam., Cassia fistula L., and Ceratonia siliqua L. as non-conventional edible vegetable oils. Agrofor. Syst. 2020, 94, 1567–1579. [Google Scholar] [CrossRef]
- Leterme, P.; Londoño, A.M.; Estrada, F.; Souffrant, W.B.; Buldgen, A. Chemical composition, nutritive value and voluntary intake of tropical tree foliage and cocoyam in pigs. J. Sci. Food Agric. 2005, 85, 1725–1732. [Google Scholar] [CrossRef]
- Agbaire, P.O. Levels of anti-nutritional factors in some common leafy edible vegetables of southern Nigeria. Afr. J. Food Sci. Technol. 2012, 3, 99–101. [Google Scholar]
- Gemede, H.F.; Ratta, N. Antinutritional factors in plant foods: Potential health benefits and adverse effects. Int. J. Food Sci. Nutr. 2014, 3, 284–289. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Antinutritional factors in foods for livestock. BSAP Occas. Publ. 1993, 16, 69–85. [Google Scholar] [CrossRef]
- Singh, S.; Bhadoria, B.K.; Koli, P.; Singh, A. Nutritional evaluation of top foliages for livestock feeding in semi arid region of India. Indian J. Anim. Sci. 2019, 89, 1389–1398. [Google Scholar]
- Dhanasekaran, D.K.; Dias-Silva, T.P.; Sakita, G.Z.; Abdalla, A.L.; Louvandini, H.; Elghandour, M.M.M.Y. Plants extract and bioactive compounds on rumen methanogenesis. Agrofor. Syst. 2020, 94, 1541–1553. [Google Scholar] [CrossRef]
- Valente, I.M.; Maia, M.R.G.; Malushi, N.; Oliveira, H.M.; Papa, L.; Rodrigues, J.A.; Fonseca, A.J.M.; Cabrita, A.R.J. Profiling of phenolic compounds and antioxidant properties of European varieties and cultivars of Vicia faba L. pods. Phytochemistry 2018, 152, 223–229. [Google Scholar] [CrossRef]
- Singh, S.; Bhadoria, B.K.; Koli, P.; Lata, S. Seasonal variation in chemical and biochemical constituents of tropical top feed species: Components in silvipasture system. Range Manag. Agrofor. 2021, 42, 312–319. [Google Scholar]
- Horvarth, P.J. The Nutritional and Ecological Significance of Acer-Tannins and Related Polyphenols; Cornell University: Ithaca, NY, USA, 1981. [Google Scholar]
- Koupai-Abyazani, M.R.; McCallum, J.; Bohm, B.A. Identification of the constituent flavanoid units in sainfoin proanthocyanidins by reversed-phase high-performance liquid chromatography. J. Chromatogr. 1992, 594, 117–123. [Google Scholar] [CrossRef]
- Fulcrand, H.; Mané, C.; Preys, S.; Mazerolles, G.; Bouchut, C.; Mazauric, J.-P.; Souquet, J.-M.; Meudec, E.; Li, Y.; Cole, R.B. Direct mass spectrometry approaches to characterize polyphenol composition of complex samples. Phytochemistry 2008, 69, 3131–3138. [Google Scholar] [CrossRef] [PubMed]
- X Liu, S.; White, E. Extraction and characterization of proanthocyanidins from grape seeds. Open Food Sci. J. 2012, 6, 5–11. [Google Scholar] [CrossRef]
- Ikawa, M.; Schaper, T.D.; Dollard, C.A.; Sasner, J.J. Utilization of Folin−Ciocalteu phenol reagent for the detection of certain nitrogen compounds. J. Agric. Food Chem. 2003, 51, 1811–1815. [Google Scholar] [CrossRef]
- Markham, K. Isolation techniques for flavonoids. In The Flavonoids; Springer: Berlin/Heidelberg, Germany, 1975; pp. 1–44. [Google Scholar]
- Stanek, N.; Jasicka-Misiak, I. HPTLC phenolic profiles as useful tools for the authentication of honey. Food Anal. Methods 2018, 11, 2979–2989. [Google Scholar] [CrossRef]
- Zlatkis, A.; Kaiser, R.E. HPTLC-High Performance Thin-Layer Chromatography; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Kovalikova, Z.; Lnenicka, J.; Andrys, R. The Influence of Locality on Phenolic Profile and Antioxidant Capacity of Bud Extracts. Foods 2021, 10, 1608. [Google Scholar] [CrossRef]
- Barakat, N.A.M.; Laudadio, V.; Cazzato, E.; Tufarelli, V. Potential contribution of Retama raetam (Forssk.) Webb & Berthel as a forage shrub in Sinai, Egypt. Arid Land Res. Manage. 2013, 27, 257–271. [Google Scholar]
- Kökten, K.; Kaplan, M.; Hatipoğlu, R.; Saruhan, V.; Çınar, S. Nutritive value of Mediterranean shrubs. J. Anim. Plant Sci. 2012, 22, 188–194. [Google Scholar]
- Gohlke, A.; Ingelmann, C.J.; Nürnberg, G.; Starke, A.; Wolffram, S.; Metges, C.C. Bioavailability of quercetin from its aglycone and its glucorhamnoside rutin in lactating dairy cows after intraduodenal administration. J. Dairy Sci. 2013, 96, 2303–2313. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of Flavonoids on Rumen Fermentation Activity, Methane Production, and Microbial Population. Biomed Res. Int. 2013, 2013, 349129. [Google Scholar] [CrossRef] [PubMed]
- Terrill, T.H.; Windham, W.R.; Hoveland, C.S.; Amos, H.E. Forage preservation method influences on tannin concentration, intake, and digestibility of sericea lespedeza by sheep. Agron. J. 1989, 81, 435–439. [Google Scholar] [CrossRef]
- Das, A.; Singh, G.P. Effect of partial replacement of groundnut cake with berseem (Trifolium alexandrinum) on nutrient digestibility, rumen fermentation, pool size and passage rate in cattle fed wheat straw based diet. Anim. Nutr. Feed Technol. 2006, 6, 19–28. [Google Scholar]
- Lowry, J.B.; Kennedy, P.M. Fermentation of flavonols by rumen organisms. In Proceedings of the Australian Society of Animal Production 21st biennial conference, Brisbane, QLD, Australia, 8–12 July 1996; ASAP: Armidale, NSW, Australia, 1996; p. 366. Available online: http://hdl.handle.net/102.100.100/226513?index=1 (accessed on 23 August 2022).
- Al-Khshemawee, H.; Du, X.; Agarwal, M.; Yang, J.O.; Ren, Y.L. Application of Direct Immersion Solid-Phase Microextraction (DI-SPME) for Understanding Biological Changes of Mediterranean Fruit Fly (Ceratitis capitata) during Mating Procedures. Molecules 2018, 23, 2951. [Google Scholar] [CrossRef]
- Zhou, H.-C.; Lin, Y.-M.; Li, Y.-Y.; Li, M.; Wei, S.-D.; Chai, W.-M.; Tam, N.F.-y. Antioxidant properties of polymeric proanthocyanidins from fruit stones and pericarps of Litchi chinensis Sonn. Food Res. Int. 2011, 44, 613–620. [Google Scholar] [CrossRef]
- Zhou, H.-C.; Tam, N.F.-y.; Lin, Y.-M.; Ding, Z.-H.; Chai, W.-M.; Wei, S.-D. Relationships between degree of polymerization and antioxidant activities: A study on proanthocyanidins from the leaves of a medicinal mangrove plant Ceriops tagal. PLoS ONE 2014, 9, e107606. [Google Scholar]
- Yoneda, S.; Nakatsubo, F. Effects of the hydroxylation patterns and degrees of polymerization of condensed tannins on their metal-chelating capacity. J. Wood Chem. Technol. 1998, 18, 193–205. [Google Scholar] [CrossRef]
- Moridani, M.Y.; Pourahmad, J.; Bui, H.; Siraki, A.; O’Brien, P.J. Dietary flavonoid iron complexes as cytoprotective superoxide radical scavengers. Free Radical Biol. Med. 2003, 34, 243–253. [Google Scholar] [CrossRef]
- McAllister, T.A.; Martinez, T.; Bae, H.D.; Muir, A.D.; Yanke, L.J.; Jones, G.A. Characterization of condensed tannins purified from legume forages: Chromophore production, protein precipitation, and inhibitory effects on cellulose digestion. J. Chem. Ecol. 2005, 31, 2049–2068. [Google Scholar] [CrossRef]
- Asam Raza, M.; Shahwar, D.; Khan, T. Radical scavenging, proteases activities, and phenolics composition of bark extracts from 21 medicinal plants. J. Che. 2015, 2015, 951840. [Google Scholar] [CrossRef]
- Koli, P.; Singh, S.; Bhadoria, B.K.; Agarwal, M.; Lata, S.; Ren, Y. Sequential Extraction of Proanthocyanidin Fractions from Ficus Species and Their Effects on Rumen Enzyme Activities In Vitro. Molecules 2022, 27, 5153. [Google Scholar] [CrossRef] [PubMed]
- McNabb, W.C.; Waghorn, G.C.; Peters, J.S.; Barry, T.N. The effect of condensed tannins in Lotus pedunculatus on the solubilization and degradation of ribulose-1,5-bisphosphate carboxylase (EC 4.1.1.39; Rubisco) protein in the rumen and the sites of Rubisco digestion. Br. J. Nutr. 1996, 76, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.J.; Barry, T.N.; McNabb, W.C. Polyphenols and agriculture: Beneficial effects of proanthocyanidins in forages. Agric., Ecosyst. Environ. 1999, 75, 1–12. [Google Scholar] [CrossRef]
- Waghorn, G.C.; Ulyatt, M.J.; John, A.; Fisher, M.T. The effect of condensed tannins on the site of digestion of amino acids and other nutrients in sheep fed on Lotus corniculatus L. Br. J. Nutr. 1987, 57, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Jonker, A.; Yu, P. The occurrence, biosynthesis, and molecular structure of proanthocyanidins and their effects on legume forage protein precipitation, digestion and absorption in the ruminant digestive tract. Int. J. Mol. Sci. 2017, 18, 1105. [Google Scholar] [CrossRef]
- Min, B.R.; McNabb, W.C.; Barry, T.N.; Peters, J.S. Solubilization and degradation of ribulose-1,5-bisphosphate carboxylase/oxygenase (EC 4.1.1.39; Rubisco) protein from white clover (Trifolium repens) and Lotus corniculatus by rumen microorganisms and the effect of condensed tannins on these processes. The J. Agri. Sci. 2000, 134, 305–317. [Google Scholar] [CrossRef]
- Waghorn, G.C.; McNabb, W.C. Consequences of plant phenolic compounds for productivity and health of ruminants. Proc. Nutr. Soc. 2003, 62, 383–392. [Google Scholar] [CrossRef]
- McNabb, W.C.; Peters, J.S.; Foo, L.Y.; Waghorn, G.C.; Jackson, F.S. Effect of condensed tannins prepared from several forages on the in vitro precipitation of ribulose-1, 5-bisphosphate carboxylase (Rubisco) protein and its digestion by trypsin (EC 2.4. 21.4) and chymotrypsin (EC 2.4. 21.1). J. Sci. Food Agric. 1998, 77, 201–212. [Google Scholar] [CrossRef]
- Getachew, G.; Pittroff, W.; Putnam, D.; Dandekar, A.; Goyal, S.; DePeters, E. The influence of addition of gallic acid, tannic acid, or quebracho tannins to alfalfa hay on in vitro rumen fermentation and microbial protein synthesis. Anim. Feed Sci. Technol. 2008, 140, 444–461. [Google Scholar] [CrossRef]
- Bento, M.H.L.; Makkar, H.P.S.; Acamovic, T. Effect of mimosa tannin and pectin on microbial protein synthesis and gas production during in vitro fermentation of 15N-labelled maize shoots. Anim. Feed Sci. Technol. 2005, 123, 365–377. [Google Scholar] [CrossRef]
- Jain, R.; Singh, M.; Dezman, D.J. Qualitative and quantitative characterization of phenolic compounds from lantana (Lantana camara) leaves. Weed Sci. 1989, 37, 302–307. [Google Scholar] [CrossRef]
- Mandels, M.; Weber, J. The Production of Cellulases; ACS Publications: Washington, DC, USA, 1969. [Google Scholar]
- Singh, S.; Kundu, S.S. Effect of tropical browse leaves supplementation on rumen enzymes of sheep and goats fed Dichanthium annulatum grass-based diets. Trop. Anim. Health Prod. 2010, 42, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Tsubota, H.; Hoshino, S. Transaminase activity in sheep rumen content. J. Dairy Sci. 1969, 52, 2024–2028. [Google Scholar] [CrossRef]
- Yatzidis, H. Measurement of transaminases in serum. Nature 1960, 186, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Strecker, H.J. L-Glutamic dehydrogenase from liver. Meth. Enzymol. 1955, 2, 220–221. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with folin phenol reagent. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Blackburn, T.H.; Hobson, P.N. Isolation of proteolytic bacteria from the sheep rumen. Microbiology 1960, 22, 282–289. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, M. Tannins: Their adverse role in ruminant nutrition. J. Agric. Food Chem. 1984, 32, 447–453. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zinn, R.A.; Owens, F.N. A rapid procedure for purine measurement and its use for estimating net ruminal protein synthesis. Can. J. Anim. Sci. 1986, 66, 157–166. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Components (mg/g) | AP–50 | AP–70 | AP–DW | EJ–50 | EJ–70 | EJ–DW |
|---|---|---|---|---|---|---|
| (+) Catechin | 1.03 ± 0.07 a | 1.21 ± 0.38 a | 0.01 ± 0.00 c | 0.22 ± 0.01 bc | 0.12 ± 0.02 bc | 0.47 ± 0.04 b |
| (−) Epicatechin | 0.32 ± 0.04 ab | 0.44 ± 0.18 a | 0.44 ± 0.04 a | 0.12 ± 0.02 b | 0.24 ± 0.03 ab | 0.33 ± 0.01 ab |
| (+) Gallocatechin | 0.09 ± 0.01 b | 0.51 ± 0.05 a | 0.21 ± 0.02 b | 0.12 ± 0.08 b | 0.10 ± 0.08 b | 0.21 ± 0.03 b |
| (−) Epigallocatechin | 0.25 ± 0.05 ab | 0.23 ± 0.07 ab | 0.00 ± 0.00 c | 0.26 ± 0.06 ab | 0.14 ± 0.02 b | 0.35 ± 0.03 a |
| (+) Catechin-4-phloroglucinol | 3.58 ± 1.24 a | 4.40 ± 0.94 a | 0.00 ± 0.00 c | 3.30 ± 0.34 ab | 1.56 ± 0.10 bc | 3.96 ± 0.38 a |
| (+) Gallocatechin-4-phloroglucinol | 1.45 ± 0.09 b | 0.85 ± 0.13 c | 2.86 ± 0.12 a | 1.24 ± 0.11 b | 0.65 ± 0.09 c | 1.36 ± 0.09 b |
| (−) Epigallocatechin-4-phloroglucinol | 1.27 ± 0.10 b | 1.10 ± 0.12 bc | 1.12 ± 0.03 b | 0.86 ± 0.09 c | 1.02 ± 0.03 bc | 1.54 ± 0.13 a |
| Enzymes EC50 (mg/mL) | Control | TA | GA | AP–50 | AP–70 | AP–DW | EJ–50 | EJ–70 | EJ–DW |
|---|---|---|---|---|---|---|---|---|---|
| R–GOT (P) | 22.22 ± 0.64 a | 20.54 ± 1.13 ab | 21.77 ± 0.80 a | 20.27 ± 2.06 ab | 5.45 ± 0.71 d | 20.79 ± 1.03 ab | 18.5 ± 1.25 b | 13.78 ± 0.86 c | 4.44 ± 0.57 d |
| R–GOT (B) | 23.31 ± 2.09 b | 17.54 ± 0.13 cd | 22.78 ± 0.80 b | 9.02 ± 0.63 e | 19.58 ± 0.46 bc | 19.6 ± 1.78 bc | 34.24 ± 1.58 a | 18.39 ± 2.32 c | 14.05 ± 1.86 d |
| R–GPT (P) | 20.49 ± 1.21 a | 17.52 ± 0.70 d | 6.89 ± 0.12 e | 9.67 ± 1.32 e | 17.75 ± 1.92 cd | 16.23 ± 0.79 d | 24.03 ± 0.89 b | 20.66 ± 0.65 c | 7.83 ± 1.0 e |
| R–GPT (B) | 24.96 ± 1.74 a | 17.12 ± 0.15 c | 15.52 ± 0.53 c | 15.91 ± 0.24 c | 17.43 ± 0.71 c | 16.86 ± 1.43 c | 3.23 ± 0.26 d | 23 ± 1.03 ab | 21.04 ± 2.06 b |
| R–Cellulase | 82.84 ± 2.89 b | 124.32 ± 1.63 a | 6.62 ± 0.56 d | 9.08 ± 1.02 d | 23.36 ± 1.34 c | 81.09 ± 1.54 b | 21.21 ± 1.86 c | 82.50 ± 2.32 b | – |
| Diet | Solubilization (%) | NRNAQ | |
|---|---|---|---|
| AP | 16.60 ± 1.97 c | 0.25 ± 0.01 fgh | |
| EJ | 15.03 ± 1.06 cd | 0.26 ± 0.02 efg | |
| WS | 8.95 ± 0.95 e | 0.16 ± 0.01 i | |
| BH | 3.04 ± 0.08 f | 0.17 ± 0.01 i | |
| AP + WS | 1:1 | 15.39 ± 1.67 c | 0.42 ± 0.03 a |
| 2:1 | 31.46 ± 1.06 a | 0.33 ± 0.01 cd | |
| 3:1 | 23.05 ± 0.51 b | 0.16 ± 0.01 i | |
| AP + BH | 1:1 | 10.55 ± 0.83 e | 0.37 ± 0.01 bc |
| 2:1 | 11.30 ± 0.28 e | 0.41 ± 0.03 ab | |
| 3:1 | 8.87 ± 1.21 e | 0.31 ± 0.02 de | |
| EJ + WS | 1:1 | 10.77 ± 1.45 e | 0.25 ± 0.01 fgh |
| 2:1 | 23.28 ± 1.23 b | 0.23 ± 0.01 gh | |
| 3:1 | 28.19 ± 0.62 a | 0.20 ± 0.01 hi | |
| EJ + BH | 1:1 | 11.75 ± 1.55 de | 0.32 ± 0.03 cd |
| 2:1 | 9.76 ± 0.87 e | 0.29 ± 0.01 def | |
| 3:1 | 9.71 ± 0.76 e | 0.24 ± 0.02 fgh | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Koli, P.; Bhadoria, B.K.; Agarwal, M.; Lata, S.; Ren, Y.; Du, X. Proanthocyanidins Modulate Rumen Enzyme Activities and Protein Utilization In Vitro. Molecules 2022, 27, 5870. https://doi.org/10.3390/molecules27185870
Singh S, Koli P, Bhadoria BK, Agarwal M, Lata S, Ren Y, Du X. Proanthocyanidins Modulate Rumen Enzyme Activities and Protein Utilization In Vitro. Molecules. 2022; 27(18):5870. https://doi.org/10.3390/molecules27185870
Chicago/Turabian StyleSingh, Sultan, Pushpendra Koli, Brijesh K. Bhadoria, Manjree Agarwal, Suman Lata, Yonglin Ren, and Xin Du. 2022. "Proanthocyanidins Modulate Rumen Enzyme Activities and Protein Utilization In Vitro" Molecules 27, no. 18: 5870. https://doi.org/10.3390/molecules27185870
APA StyleSingh, S., Koli, P., Bhadoria, B. K., Agarwal, M., Lata, S., Ren, Y., & Du, X. (2022). Proanthocyanidins Modulate Rumen Enzyme Activities and Protein Utilization In Vitro. Molecules, 27(18), 5870. https://doi.org/10.3390/molecules27185870