Abstract
Sirtuins are NAD+-dependent deac(et)ylases with different subcellular localization. The sirtuins’ family is composed of seven members, named SIRT-1 to SIRT-7. Their substrates include histones and also an increasing number of different proteins. Sirtuins regulate a wide range of different processes, ranging from transcription to metabolism to genome stability. Thus, their dysregulation has been related to the pathogenesis of different diseases. In this review, we discussed the pharmacological approaches based on sirtuins’ modulators (both inhibitors and activators) that have been attempted in in vitro and/or in in vivo experimental settings, to highlight the therapeutic potential of targeting one/more specific sirtuin isoform(s) in cancer, neurodegenerative disorders and type 2 diabetes. Extensive research has already been performed to identify SIRT-1 and -2 modulators, while compounds targeting the other sirtuins have been less studied so far. Beside sections dedicated to each sirtuin, in the present review we also included sections dedicated to pan-sirtuins’ and to parasitic sirtuins’ modulators. A special focus is dedicated to the sirtuins’ modulators identified by the use of virtual screening.
1. Introduction
Sirtuins (SIRTs) are a family of evolutionary, conserved enzymes that depend on nicotinamide adenine dinucleotide (NAD+) and are homologues of the silence information regulator 2 (Sir2) enzyme, thus also being known as Sir2-like proteins [1]. In mammals, the seven known sirtuins, named SIRT-1 to SIRT-7, are classified as class III histone deacetylases (HDACs). Their substrates, however, have been reported to be also non-histone proteins, such as cytoskeletal proteins, signaling molecules, transcription factors, chaperones, p53 and DNA repair proteins [2,3,4]. Sirtuins, moreover, often possess other enzymatic activities apart from the deacetylation one, such as deacylation and mono-ADP-ribosylation [2,3,4,5]. Structurally, the seven isoforms share a central catalytic domain of about 270 amino acids, where a Rossman fold and a smaller domain with the NAD+-binding module and a zinc-binding one create the enzymatic active site; the seven sirtuins then differ in the N-terminal and C-terminal domains [6]. The crystal structures of the seven sirtuins, chosen on the basis of resolution values (<2.00 Å), are represented in Figure 1.
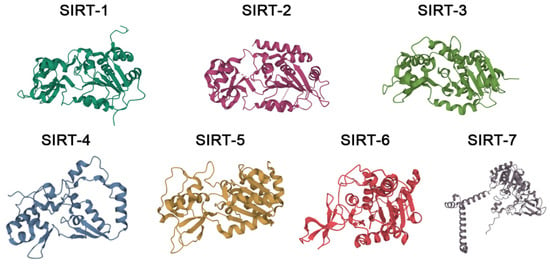
Figure 1.
Crystal structures of the 7 human sirtuins. The PDB identifiers for SIRT-1, SIRT-2, SIRT-3, SIRT-5, and SIRT-6 are 4KXQ, 4Y6O, 4BN4, 6LJK, and 6HOY, respectively. SIRT-4 and SIRT-7 do not exist in a crystal structure; thus, the AlphaFold-software-predicted structures are shown with the identifiers AF-A0A347ZJG7-F1 and AF-Q9NRC8-F1, respectively.
Sirtuins are characterized by diverse subcellular localizations, unique substrate specificity and distinct enzymatic activities (Table 1) [7], and this confers to each isoform’s specific functions. Given their involvement in different biological pathways, ranging from transcription to metabolism to genome stability, their dysregulation is implicated in many diseases, such as cancer, neurodegenerative disorders, diabetes, and cardiovascular and autoimmune diseases [8,9,10,11,12]. Pharmacological modulation of their enzymatic activity becomes, therefore, a promising strategy to modify disease initiation and/or progression. Extensive research has already been conducted on SIRT-1 modulators, while compounds targeting the other sirtuins have been less studied or faced issues such as reduced isoform selectivity, poor water solubility or limited cell membrane permeability.

Table 1.
Enzymatic activities and cellular localizations of the 7 sirtuins.
In the first part of the present review, we summarize the sirtuins’ modulators that have demonstrated biological effects in cellular or animal models in cancer, neurodegenerative diseases and type 2 diabetes (T2D), to demonstrate the great potential of these enzymes as therapeutic/drug targets. In the second part, a special focus will be dedicated to the sirtuins’ modulators identified by the use of virtual screenings (VS).
2. Sirtuins as Targets in Different Pathologies
Sirtuins play a critical role in cancer and have been reported with a dual function in tumorigenesis, behaving as oncopromoter or oncosuppressor, depending on the sirtuin and on the cancer type [28]. In addition, often one same sirtuin has controversial roles within the same cancer type, with independent studies reporting that activators/overexpression or inhibitors/knock down have both anti-cancer effects [28]. The possibility that sirtuins act as a tumor suppressor is mainly corroborated by their role in maintaining genome stability through chromatin regulation and DNA repair. On the other hand, some sirtuins were found to be overexpressed in certain tumors and were also shown to promote angiogenesis. Moreover, sirtuins affect metabolism, thus acquiring pro-tumorigenic or oncosuppressive functions. For instance, one reason to consider SIRT-6 as an oncosuppressor is represented by the shift from aerobic respiration to glycolysis observed in SIRT6-deficient cells resembling the Warburg effect, typical of cancer cells. However, the number of pathways affected by the different sirtuins is so wide that in a different cancer context the final outcome of sirtuins’s contribution can be dual. Of note, changes in sirtuins’ expression are often a consequence of time, and not a cause of different oncogenic pathways. The most studied SIRTs in cancer are SIRT-1, -2, -3 and -6, with modulators that either exhibit an anti-proliferative effect, induce apoptosis, or sensitize cancer cells to existing chemotherapeutics in vitro, or that block tumor growth in vivo.
Sirtuins are also involved in many neurodegenerative diseases, among which Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD), amyotrophic lateral sclerosis (ALS) and multiple sclerosis (MS) are the most common. Neurodegenerative diseases are characterized by neuronal cell death, which leads to progressive loss of mobility, coordination, sensation and memory, and current treatments aim solely at relieving physical or mental symptoms, since no cures exist for any neurodegenerative diseases [29]. Sirtuins’ modulators proved to be effective in reducing the symptoms and in some cases also in preventing progression of neurodegeneration (AD, PD, ALS) or inflammatory relapses (MS). SIRT-1 and -2 (with some research conducted also on SIRT-3 and -6) represent the most investigated SIRTs under a pharmacological perspective. In addition, genetic studies have been also conducted on the other sirtuins, demonstrating the potential of this class of proteins as drug targets in neurodegenerative diseases [11]. All sirtuins, however, do not possess the same function in neurodegenerative diseases, with some that are downregulated and others that are upregulated in disease progression.
Regarding type 2 diabetes (T2D), some studies have been carried in the past decades to define the role of SIRTs [8], while less studies involving SIRTs modulators have been performed. In vivo experiments demonstrated that effective treatments were obtained with SIRT-1 activators, or SIRT-2 and SIRT-6 inhibitors.
The sirtuins’ pharmacological modulation that has been reported to exert therapeutic effects in vitro and in vivo in a selection of diseases, namely, cancer, neurodegenerative diseases and T2D, is summarized in Table 2.
Table 2.
Overview of the pharmacological modulation of sirtuins with a therapeutic effect in cancer, neurodegenerative diseases (Alzheimer’s disease—AD, Huntington’s disease—HD, Parkinson’s disease—PD, Amyotrophic Lateral Sclerosis—ALS, Multiple Sclerosis—MS) and type 2 diabetes (T2D).
In the following summary of sirtuins’ inhibitors and activators, those that also modulate other enzymes (e.g., kinases or others) and therefore that are not specific for the sirtuin family, have not been reported, since their beneficial effects may not be entirely attributed to the sirtuin modulation.
2.1. SIRT-1 Modulators as Therapeutics
SIRT-1 is the best characterized isoform of the sirtuin class of enzymes: it is located in the nucleus and it regulates gene stability, stress response and apoptosis [30,31,32]. Its modulators have been extensively investigated in the past decades [33]; a particular mention should be conducted of the inhibitor EX-527, which is selective for SIRT-1 over the other sirtuins, and furthermore has been also used in clinical trials as treatment for HD. Other inhibitors did not prove to be suitable for clinical studies until now, even though many were explored in in vitro and in vivo settings. Resveratrol and the compounds developed by the company Sirtris Pharmaceuticals, SRT1720, SRT2183, SRT1460, have been extensively used in disease models as SIRT-1 activators. However, Pacholec and colleagues demonstrated that they are not direct activators of SIRT-1 [34], and therefore even though they exhibited therapeutic effects in many diseases and SIRT-1 was modulated [35,36,37,38,39,40,41], they are excluded from the below summary of SIRT-1 activators.
2.1.1. SIRT-1 Modulators in Cancer
In cancer, SIRT-1 has a controversial role, being reported with oncopromoter and oncosuppressor functions. This was also confirmed by several studies with inhibitors that were shown as promising anti-cancer agents, and with activators that had positive biological effects as well, in some other cancer types.
SIRT-1 inhibition has been shown to exert a great therapeutic effect in several types of cancers. In two independent studies, Selisistat (called also EX-527 or SEN0014196 and discovered by a high-throughput screen in 2005) [42] and compound JGB1741 (developed starting from the structure of Sirtinol and modifying its structure on the basis of its probable interaction with the protein SIRT-1) [43] induced apoptosis in leukaemia cells. The induction of apoptosis was also observed upon the treatment of breast cancer and HCC cells with compound JGB1741 [43]. EX-527, which has a selectivity 200 times higher for SIRT-1 with respect to the other sirtuins, on the other hand, has been investigated in several other cancer types: in glioma [44], bladder [45], and endometrial [46] cancers, it exhibited anti-proliferative properties; in pancreatic cancer, it blocked cell proliferation and sensitized its cells to gemcitabine [47]; in endometrial [46] carcinoma, it reduced cancer cell proliferation and tumor growth in mouse studies. In lung cancer, inhibition was demonstrated to have several anti-cancer effects, with research showing that EX-527 increased sensitivity to MK-1775, as well as reduced tumor growth in an in vivo mouse model [48] and increased sensitivity to cisplatin in vitro [49]. Breast cancer has been investigated with SIRT-1 inhibitors: two studies with EX-527 showed that it causes cell cycle arrest [50] and suppresses the resistance to Hsp90 inhibitors in cancer cells [51]. Other works indicated that compound 4d [52], which was discovered by VS as well (see Section 3.2), compound 3a and its family of Aurone derivative, synthetized starting from a thiobarbiturate structure (a promising inhibitor of sirtuins), [53] and compounds 27 and 30, obtained from a synthesized library of pseudopeptides, [54] inhibited cell proliferation; in addition, inhibitor Amuresin G, obtained from a plant extract of V. amurensis, increased cellular uptake of doxorubicin in doxorubicin-resistant breast cancer cells and restored the responsiveness of MCF-7/ADR cells to doxorubicin in vitro and in vivo. [55]
SIRT-1 inhibitors have been investigated in prostate cancer as well: EX-527 increased the sensitivity of cancer cells to vesicular stomatitis virus oncolysis [56], while compound 17 [57] (discovered by VS, see Section 3.1) exhibited anti-proliferative effects in vitro.
In colon and NSCL cancer, inhibitor Inauhzin (found by a computational structure-based screening) decreased cell proliferation, induced senescence and apoptosis and inhibited tumor growth in mouse models. [58] One more compound to mention is inhibitor S1th, which was discovered by VS (see Section 3.2) and was reported to possess anti-proliferative proprieties in cervical cancer cells [59].
The mechanism of action of all these SIRT-1 inhibitors in tumors is the induction of cell cycle arrest and tumor cell apoptosis via the p53 pathway, increasing Bax/Bcl-2 ratio and PARP cleavage.
SIRT-1 pharmacological activation was also demonstrated to have anti-cancer effects. In particular, in neuroblastoma Chikusetsu saponin V (extracted from Panax japonicus) reduced H2O2-induced oxidative stress in cell studies by increasing the activity of SOD and increasing the levels of GSH [60].
2.1.2. SIRT-1 Modulators in Neurodegenerative Disorders
Neurodegenerative diseases treatment may benefit from SIRT-1 inhibitors as well as activators.
In particular, inhibitors EX-527 [61] and Baicalin (isolated from the roots of the Radix Scutellariae plant) [62] were demonstrated to have a neuroprotective effect in mice, by reducing acute stress response and by negatively regulating the expression of IL-6, TNF-α, and Il-1β in the hippocampus and hypothalamus through the regulation of the SIRT-1-NF-kB pathway.
On the other hand, SIRT-1 activators also exhibited protection from neuroinflammation. Specifically, Salidroside (extracted from Rhodiola rosea L.) reduced inflammation-induced cognitive defects in mice through the Nrf-2/HO-1/NF-κB pathway [63], and 17β-estradiol blocked neuroinflammation and neuronal apoptosis, halting cognitive dysfunction and memory impairment in a male aging mouse model [64].
In AD, pieces of evidence revealed that SIRT-1 activation may be protective, while pharmacological inhibition aggravated tau accumulation [65] and abolished the resveratrol-mediated attenuation of autophagy in an AD cell model [66]. Other examples of activators/molecules causing SIRT-1 overexpression with a biological effect in AD are Syzygium aromaticum extract (obtained by the ethanol extraction of a pool of molecules from Syzygium aromaticum), which instead maintains oxidative balance in vitro and in AD patients’ serum by increasing the expression and activity of SOD and GSH [67], and dihydromyricetin (extracted from Ampelopsis grossedentata), which inhibits neuronal cell apoptosis, ameliorating cognitive dysfunction in the AD rat model by activating the AMPK/SIRT-1/PGC-1α pathway [65,68]. Other pieces of evidence include the SLAB51 probiotic formulation, which reduces oxidative species levels in AD mouse, thus having a neuroprotective effect by activating SIRT-1 and increasing the oxidative stress defences [69] and Cilostazol, which suppresses β-amyloid production in vitro by stimulating the expression of α-secretase via the ADAM10/SIRT-1 pathway activation [70], thus improving or protecting cognitive function, through increased glucose metabolism, in AD patients with white matter lesions [71].
In HD, conversely, SIRT-1 inhibition seems to be required in order to gain neuroprotection. Indeed, the inhibitor EX-527 displayed neuroprotection in Drosophila and in mouse models of HD, by improving motor function through inactivation of the FOXO3A and CREB pathways and the subsequent decrease in the expression of exon 1 of the Htt protein [72], and has been further studied in a few clinical trials (Phase I and II) [73,74]. Specifically, in two studies in healthy volunteers and HD patients, EX-527 was found to be well tolerated with no adverse effects, even though, unfortunately, the circulating levels of soluble huntingtin were not affected. Nevertheless, two other studies in the HD mouse model showed that increasing NAD+ availability and causing SIRT-1 activation with β-lapachone (a natural o-naphthoquinone compound, and a substrate of NADH:quinone oxidoreductase, NQO1) results in neuroprotection by activating autophagy of Htt exon1-expressing cells [75,76], revealing that further investigations involving sirtuin modulators and HD are still needed.
In PD, SIRT-1 proved to be neuroprotective when activated, as indicated by some pharmacological studies, including the use of the activators Echinacoside [72,73] derived from the fleshy stem of distance, a widely used Chinese herb [72,77], and Embelin, a natural product with structural resemblance to ubiquinone, exhibiting mitochondrial uncoupling and antioxidant effects [73,78], which are neuroprotective in an induced neurotoxic PD mouse model. Echinacoside mediates the autophagic degradation of α-synuclein by the increased expression of FoxO1 via SIRT-1 activation [77]. The mitochondrial uncoupling effect of Embelin increases NAD+/NADH levels, followed by enhanced SIRT-1, PGC1α and mitochondrial biogenesis conferring to Embelin neuroprotective properties [78].
Another neurodegenerative disease to be noted is ALS, for which the inhibitor EX-527 has shown a neuroprotective effect in an in vitro study, by increasing neuron viability [74], although with an action independent from SIRT-1 inhibition [74,79]. No studies have been performed with activators or with other inhibitors; therefore, there is potential for further investigation in this field.
Multiple Sclerosis (MS) is the last neurodegenerative disease covered in this review and a few studies were reported showing the anti-inflammation proprieties of inhibitor EX-527. In one study, EX-527 expanded the endogenous pool of neuronal progenitor cells without affecting their differentiation perhaps by increasing PDGFRα expression and the activity of p38 MAPK and AKT pathways [80]; in an additional study, it was able to suppress the cell differentiation of Th17 by inhibiting RORγt transcriptional activity, decreasing Th17 cell generation and function. Th17 are cells associated with multiple autoimmune diseases, to delay disease onset and reduce lymphocytic infiltration and demyelination in an in vivo mouse model [81].
2.1.3. SIRT-1 Modulators in T2D
SIRT-1 regulates glucose/lipid metabolism, since it deacetylates several proteins of the insulin signaling pathway, and its overexpression improves the sensitivity to insulin. Some SIRT-1 activators, such as JHJ1, JHJ2, and JHJ3 (derived from the structure of OAP) [82], have been reported to modulate the plasma lipid metabolism and blood glucose in high-fat fed mice by upregulating FoxO1, PPARγ, and PGC-1α genes [83], while another activator, named E6155 (identified by a high-throughput screening using the purified recombinant human SIRT-1 and HTRF SIRT-1 assay) by Liu and colleagues, improved blood glucose tolerance and insulin resistance in diabetic mice by activating LKB1/AMPK and IRS1/AKT pathways [84].
2.2. SIRT-2 Modulators as Therapeutics
SIRT-2 is localized in the cytoplasm, although it can shuttle to the nucleus to participate to several physiological and pathological functions [85]. It plays a role in several processes, including cell cycle regulation and metabolism [86,87]. SIRT-2 activators have never been discovered, while many inhibitors have been identified in the past years, with the most studied one being AGK2. Although this compound is the most selective inhibitor for SIRT-2, its poor water solubility creates the need to seek still for new SIRT-2 inhibitors. Since many SIRT-2 inhibitors have been studied also in vitro and in vivo, a selection of them is reported in the next paragraphs.
2.2.1. SIRT-2 Modulators in Cancer
In many types of cancer, SIRT-2 acts as a tumor promotor and therefore its inhibition may be beneficial. Often, SIRT-2 inhibitors act as anti-proliferative agents or have a cytotoxic effect, since this sirtuin is involved in cell cycle regulation, in particular in the G(2)/M transition [87]. Nevertheless, in some other cancers, it was shown that SIRT-2 possesses tumor suppressor functions, which, however, were not further investigated pharmacologically, since no SIRT-2 activators are currently available.
In glioma, SIRT-2 inhibition has shown relevant therapeutic effects: AGK2 [88], as well as NH4-13 [89], reduced cancer cell proliferation, and AGK2, in addition, displayed anti-proliferative activity in cancer stem cells [90]; moreover, another SIRT-2 inhibitor, called AK7, was shown to reduce tumor growth in an in vivo mouse model [88]. In particular, Funato and colleagues showed that the SIRT-2-mediated inactivation of tumor suppressor p73 is crucial in the proliferation and tumorigenicity of glioblastoma cells, thus making SIRT-2 inhibition by AGK2 or AK7 a valid anti-cancer treatment [88].
Compound AC-93253 and Salermide (chosen at a concentration at which SIRT-2, but not SIRT-1, is inhibited) blocked cell proliferation and downregulated the c-Myc and N-Myc oncoproteins in neuroblastoma and in pancreatic cancer [91]. In addition, in this last type of cancer, other inhibitors were reported to possess an anti-proliferative activity, such as NPD11033 [92], AF8 [93] and NH4-13 [89]. One more inhibitor, namely, compound AC-93253, then displayed a cytotoxic effect in pancreatic cancer cells, by triggering apoptosis [94]
Many SIRT-2 inhibitors displayed their anti-cancer activity by blocking cell proliferation. Examples include SirReal2 [95] and TM [96] in colorectal cancer; compounds 2 and 3 in adenocarcinoma (which were discovered by VS, and mentioned in Section 3.2) [97], SirReal2 [95], 4-chromanone compounds 6f and 12a [98], compound 24a [99] and NH4-13 [89] in lung cancer; compound 35 in NSCL cancer [100]; NCO-90 and NCO-141 [101], TM [96], compounds 35 and 39 [100] in leukaemia; SirReal2 [95], compounds 35 and 39 [100], the Cambinol analogues compound 24 [102] and compounds 55 and 56 [103] in lymphoma; AF8 [93], NH4-13 [89], SirReal2 [95], compound 35 [100], compound 6f and 12a [98] RK-91230156 [104], Splitomicin derivatives compounds 5c, 8c and (R)-8c (discovered by VS, see Section 3.2) [105], γ-mangostin [106], Tenovin-D3 [107], RK-9123016 [104], compounds 2 and 3 [97] in breast cancer; SirReal2 [95], compounds 2 and 3 [97] and NH4-13 [89] in cervical cancer.
The anti-proliferative effects of these compounds were described in several works together with other molecular mechanisms affected by SIRT-2 inhibition. For instance, compound TM was reported to promote ubiquitination and degradation of c-Myc oncoprotein in different cancer cell lines [96], while compounds 35 and 39 induced apoptosis in leukaemia and breast cancer cells [100]. Moreover, cell viability of cancer cells was reduced because inhibitor RK-9123016 was accompanied by a decrease in c-Myc expression [104]; in another study, SIRT-2 inhibition by Tenovin-D3 promoted expression of the cell-cycle regulator and p53 target p21WAF1/CIP1 (CDKN1A) in a p53-independent manner, along with a reduction in cell proliferation [107]. NCO-90/141 simultaneously caused apoptosis and autophagy in leukemic cell lines, by inducing apoptosis via caspase activation and mitochondrial superoxide generation, and by increasing the LC3-II level together with autophagosome accumulation, indicating autophagic cell death [101]. In a similar way, the induction of apoptosis was observed along with cell growth arrest when treating lymphoma cells with compounds 55 and 56 [103].
The therapeutic effect of some other SIRT-2 inhibitors relies on the combination of the cytotoxicity of the compounds and their inhibitory activity, having been observed that the compound toxicity was crucial in obtaining anti-cancer effects; this is the mechanism suggested for AC-93253 in prostate, pancreatic and lung cancer [94].
In addition, some inhibitors have been further studied in in vivo mouse models and blocked tumor growth. These studies were performed in colorectal cancer with compounds AGK2 [108], AF8 [93] and NH4-13 [89]; in gastric cancer using SirReal2 [95]; in breast cancer with compound TM [96].
From the point of view of the isoform-selectivity of the compounds, the most selective SIRT-2 inhibitors listed above are AGK2, AK7, NH4-13, NPD11033, AF8, AC-93253, SirReal2, TM and RK-91230156. In particular, AGK2, which was identified from a screening of 200 compounds [109], inhibits SIRT-2 with an IC50 of 3.5 μM, and inhibits SIRT-1 and -3 with IC50 of 30 and 91 μM, respectively [110], while NH4-13 inhibits SIRT-2 with an IC50 of 0.087 μM, and its IC50 for SIRT-1, -3, -5 and -6 is reported to be higher than 50 μM [89]. Inhibitor AK7 was identified after making a substructure search of sulfobenzoic acid derivatives that were predicted to be brain permeable, since the authors were targeting HD and had already identified inhibitors with this core structure. AK7 was reported to be brain permeable in vivo, with an IC50 of 15.5 μM, and to neither modulate SIRT-1 or -3 up to 20-50 μM [111]. NPD11033, which was discovered by a high-throughput screen method has an SIRT-2 IC50 of 0.46 μM, while its IC50 for SIRT-1 and -3 is higher than 100 μM [92]. One more inhibitor is AF8, which is characterized by an IC50 of 0.06 μM for SIRT-2, and of 11 μM or above 50 μM for SIRT-1 and -3, respectively [93]. The inhibitory ability of AC-93253 was tested in vitro and not in enzymatic assays, and it was reported that its IC50 was 6 μM, and that it was 7.5- and 4-fold more potent in inhibiting SIRT-2 than the isoforms SIRT-1 and -3 [94]. SirReal2 was identified through an in vitro compound screening of an in-house developed library [112], and its IC50 for SIRT-2 is 0.23 μM, while it is above 50 μM for SIRT-1 and -3 [95]. TM is another SIRT-2 selective inhibitor, with an IC50 values of 0.4 μM for SIRT-2, and it is at least 650-fold more selective for SIRT-2 compared to SIRT-1 and -3 [95]. RK-91230156, finally, has an IC50 for SIRT-2 of 0.18 μM, while it is above 100 μM for SIR-1 and -3 [104].
Less selective inhibitors, but still to be mentioned, are the following compounds. Compounds 6f and 12a, which were discovered after optimizing the hydrophilicity of an inhibitor previously developed by the group, have IC50 of 3.7 and 12.2 μM, respectively, while their IC50 for SIRT-1 and -3 are above 200 μM [98]. Tenovin-6, instead, was discovered by screening, in a cell-based assay, 30,000 drug-like small molecules present in the Chembridge DIVERSet, making a selection of those that had been reported to activate p53 [113], and exhibits IC50 for SIRT-2, -1 and -3, respectively, of 9, 26 and above 50 μM [95]. Moreover, Yang and colleagues demonstrated that compound 24a, which was identified with X-ray crystal structure-guided structure–activity relationship (SAR) studies, has an IC50 value of 0.815 μM; the modulation of other sirtuins, however, was not investigated [99]. Moreover, compounds 35 and 39 were discovered after SAR studies performed on an existing SIRT-2 inhibitor, and they proved to be quite effective in inhibiting SIRT-2 (IC50 10.4 and 1.5 μM, respectively) and with a good selectivity for this isoform (further studies on compound 39 with SIRT-1, -3, -5 revealed no non-specific effects up to 100 μM) [100]. Splitomicin derivatives, compounds 5c, 8c and (R)-8c, were discovered by VS (see Section 3.2) and have been reported with IC50 ranging from 1.0 to 1.5 μM (the modulation of other sirtuins was not investigated) [105]. γ-mangostin is one other SIRT-2 inhibitor to mention, since it displays ≥6-fold selectivity against SIRT-2 (SIRT-2 IC50 = 3.8 µM; SIRT-1 and -3 IC50 in the 22–26 µM range); this xanthone extracted from the tropical plant Garcinia mangostana was investigated after it was reported that the α-isoform of mangostin was able to modulate sirtuins [106].
2.2.2. SIRT-2 Modulators in Neurodegenerative Disorders
Among different tissues, SIRT-2 expression is most abundant in the brain, at the neurons and oligodendrocytes levels; thus, the role of SIRT-2 in neurodegenerative disorders has been extensively investigated in the past years [77,114]. SIRT-2 inhibitors have been shown to have a neuroprotective effect in the main neurodegenerative disorders. In particular, Wang and colleagues showed that inhibitor AGK2, which was proved to be brain permeable, was able to significantly decrease LPS-induced neuroinflammation markers in cell and mice models [115]. Furthermore, another inhibitor, namely, AK1, provided some neuroprotection in the hippocampus, in which inflammation is associated both to Alzheimer’s disease and tau-associated frontotemporal dementia [116].
It is worth noting that inhibitor AK1 was discovered along with the previously described AGK2 and it is selective for SIRT-2; however, AK1 is slightly less potent than AGK2 (12.5 μM vs 3.5 μM) [109].
In AD, several SIRT-2 inhibitors have been shown to be promising therapeutics, both in cell and mouse models. In cellular studies, SIRT-2 inhibitors displayed different mechanisms in neuroprotection: in one study, γ-mangostin induced neurite outgrowth in an AD cellular model, [81,106] in another one, AGK2 reduced the reactive gliosis (which is considered one of the hallmarks of AD), [40] while in one other study, AK1 modulated mitochondrial dysfunction in AD cells, by recovering microtubule stabilization, and by eliminating toxic Aβ, thus improving cell survival [84,117]. Collecting instead the evidence of the in vivo research conducted, one study reported that compound 33i was effective in hampering age-related cognitive decline in a senescence-accelerated mouse model, by preventing neuroinflammation, having observed reduced levels of GFAP, IL-1β, Il-6, and Tnf-α [118]. Another work demonstrated that inhibitor AK7 decreased the BACE1 and Aβ production in an AD mouse model, improving the cognitive functional defects of the mice; [86,119] one more study reported that AK7, via SIRT-2 inhibition, decreased the phosphorylated tau levels, a characteristic linked to tau pathology, resulting in increased Tau/tubulin and α-synuclein/tubulin binding, thus reducing tau and α-synuclein aggregation and neurotoxicity [120].
In addition, Biella and colleagues observed a reduction in the Aβ production in neuroglioma cells and a modification in the Amyloid Precursor Protein proteolytic processing, when SIRT-2 was inhibited with AGK2 or AK7 in two AD transgenic mouse models. While the amount of soluble Aβ decreased, the one of soluble α-amyloid protein increased, thus improving the cognitive performance of mice [88,121].
HD is another neurodegenerative disorder in which SIRT-2 inhibition is to seek. Some pieces of evidence are here described. Quinti and colleagues reported the neuroprotective effects of inhibitor MIND4 in ex vivo brain slices and in Drosophila models of HD, where the induction of cryoprotective NRF2 responses in neuronal and non-neuronal cells was seen, accompanied by reduced production of ROS and nitrogen intermediates [89,122]. Likewise, inhibitors AK1 and AGK2 also decreased Huntingtin accumulation and increased neuronal viability in Drosophila [90,123]. Moreover, two more studies in a genetic mouse model of HD demonstrated that inhibition with AK7 disrupted disease progression, by improving motor function, extending survival, reducing brain atrophy, and reducing aggregated mutant Huntingtin in the mice [88,91,111,124].
The inhibitor MIND4 was identified using an iterative structure–activity drug discovery approach, starting from a known SIRT-2 inhibitor and making chemical modifications with the aim of improving potency and selectivity [122].
In PD, several SIRT-2 inhibitors were proved to have neuroprotective effects. In several independent studies, AK1, AGK2 and compounds 86 and 102 protected dopaminergic neurons in vitro and in a Drosophila PD model, rescuing cells from α-synuclein aggregation-induced toxicity [89,92,93,109,125,126]. Moreover, AK7 and AGK2 were further demonstrated to prevent dopaminergic neuronal cell death, to reduce activation of microglia and to attenuate behavioural abnormality in aged mice and rats [94,95,96,127,128,129]. Some other pieces of evidence regarding SIRT-2 inhibition in PD is given by two more studies: Di Fruscia showed that compound 10 prevented neuronal cell death, triggered by lactacystin in an in vitro model of PD, [95,130] while AK7 was neuroprotective in vivo, by down-regulating the RNAs responsible for sterol biosynthesis [98,131].
2.2.3. SIRT-2 Modulators in T2D
The function of SIRT-2 in insulin signalling is still controversial, with some authors reporting that its overexpression improves insulin sensitivity, and others demonstrating the opposite [99,132]. In fact, depending on the tissue type, SIRT-2 may exert even the opposite effect in response to insulin. For example, the SIRT-2 inhibitor AGK2, reproducing the effects obtained by SIRT-2 downregulation, improves insulin sensitivity in C2C12 cells (skeletal muscle cells) [89,133]. Further studies are needed with SIRT-2 activators and in different tissues in order to reach a conclusion regarding the possibility of targeting SIRT-2 in T2D.
2.3. SIRT-3 Modulators as Therapeutics
SIRT-3 is located in the mitochondria and has been shown to act on several metabolic and respiratory enzymes regulating their functions [134]. In addition, SIRT-3 can regulate the production and clearance of ROS by deacetylating numerous mitochondrial enzymes [135,136,137].
2.3.1. SIRT-3 Modulators in Cancer
The role of SIRT-3 in cancer is controversial and there are several reports describing its function as an oncogene as well as an oncosuppressor [138]. Considering its oncogenic role, different inhibitors of the SIRT-3 deacetylase activity were developed and proved to determine cell death by increasing ROS production. This mechanism of action was reported for the compound LC-0296 in head and neck squamous cell carcinoma (HNSCC), and for 3-TYP in acute myeloid leukemia (AML) [139,140]. In addition, LC-0296 works synergistically to increase the sensitivity of HNSCC cells to radiation and cisplatin treatment [139]. The selectivity of LC-0296 for SIRT-3 is approximately ~20- and 10-fold greater compared to SIRT-1 and SIRT-2 (IC50 3.6, 67 and 33 μM for SIRT-3, SIRT-1 and SIRT-2, respectively) [139]. The IC50s of 3-TYP were of 16, 88 and 92 nM for SIRT-3, SIRT-1 and SIRT-2, respectively [140].
Meng Li and colleagues reported that, in lymphoma cells, compound YC8-02 inhibits SIRT-3 deacetylase activity, bringing about an increase in mitochondrial proteins acetylation and subsequently cell death by autophagy [141]. YC8-02 was developed starting from a series of small molecules including an SIRT-2 selective thiomyristoyl lysine compound called TM and Biotin-TM3 (selective to SIRT-1 and SIRT-2). A compound called JH-T4 was developed, which could inhibit SIRT-1, -2 and -3, as measured in in vitro biochemical enzymatic assays, and increase its penetration into mitochondria, JH-T4 was modified by replacing the benzyl carbamoyl group with a triphenylphosphonium mitochondrial targeting moiety, thus producing YC8-02 [141]. This compound was more effective in inhibiting SIRT-3, although the selectivity over SIRT-1 and SIRT-3 was not obtained [141].
On the other hand, as mentioned, there are several pieces of evidence where the activation of SIRT-3 can represent a promising strategy to treat some types of tumors. The mechanism of action of SIRT-3 as a tumor suppressor seems to be different depending on the tumor type. In lung cancer, Wang and colleagues showed that the activation of SIRT-3 by Adjudin inhibits cellular growth and metastasis by regulating the SIRT-3-mediated FOXO3a axis [142]. Zhang and colleagues performed a structure-guided design of SIRT-3 activators and tested their activity on breast cancer. They found that Compound 33c (identified via hit-to-lead optimization starting from VS campaign, see Section 3.3) activates SIRT-3, determines breast cancer cell death by autophagy and reduces cell migration by decreasing ROS production hampering MMP activation [143]. In addition, Compound 33c could not activate SIRT-1, SIRT-2, and SIRT-5 in an enzymatic reaction assay in vitro, thus representing a selective SIRT-3 activator [143].
In conclusion, SIRT-3, as well as other sirtuins, seems to represent a target to be either activated or inhibited in a cancer-specific manner. Thus, the research of new potent SIRT-3 modulators should proceed to look for both activators and inhibitors.
2.3.2. SIRT-3 Modulators in Neurodegenerative Disorders
Regarding the role of SIRT-3 in neurodegenerative disorders, there are no reports about the use of specific modulators, but there are pieces of evidence that increasing the expression/activity of SIRT-3 could be a promising approach in a number of diseases. For example, Gastrodin can increase the levels of SIRT-3 in activated microglia causing a decrease in ROS and exhibiting a protective role [144]. In addition, Park and colleagues showed that the activation of SIRT-3 by the AMPK/CREB-PGC-1α signalling results in reduced αsyn oligomers in PD, suggesting that the use of SIRT-3 activators may represent a potential therapy to restore mitochondrial deficits and decrease αsyn-induced pathophysiology [145]. Another evidence supporting the idea that SIRT-3 activation could be a good approach in neurodegenerative disorders is that Honokiol, a not specific SIRT-3 activator, decreases ROS and lipid peroxidation, enhances antioxidant activities, and mitochondrial function, thereby reducing β-amyloid and sAPPβ production in an AD model. [146] Honokiol did not affect cell viability at concentrations up to 10 μM [146].
2.3.3. SIRT-3 Modulators in T2D
Lee and colleagues showed that the not specific SIRT-3 activator Honokiol improves insulin resistance in adipocytes, by promoting insulin receptor beta (IRβ) and PI3K/AKT/mTOR pathways, resulting in an increase in phosphorylation of the forkhead family FoxO1/FoxO3a/FoxO4 and glycogen synthase kinase-3 (GSK-3β). Conversely, the SIRT-3 inhibitor 3-TYP decreases insulin resistance [147]. These data suggest that the activation of SIRT-3 with more specific activators could represent a promising approach to treat T2D.
2.4. SIRT-4 Modulators as Therapeutics
SIRT-4 is located in the mitochondrial matrix, where it controls several pathways by modifying the activation status of different proteins [148]. To the best of our knowledge, no specific SIRT-4 modulators are available yet. There are instead several reports suggesting the suitable therapeutical approach, targeting SIRT-4 to be adopted in different diseases.
2.4.1. SIRT-4 Modulators in Cancer
Moreover, for SIRT-4, the rule that its overexpression or downregulation is cancer-specific is valid. In head and neck carcinoma, it was reported that SIRT-4 expression is induced during different types of stress, and this helps tumor cell survival [149]. In ovarian cancer, the expression of SIRT-4 correlates with a poor prognosis [150]. On the contrary, in breast cancer, neuroblastoma, liver cancer and pancreatic cancer, SIRT-4 is downregulated [151,152,153,154], suggesting an oncosuppressive role of this sirtuin.
2.4.2. SIRT-4 Modulators in Neurodegenerative Disorders
Regarding the role of SIRT-4 in neurodegenerative disorders, SIRT-4 was suggested to possess potential neuroprotective roles against excitotoxic insults by facilitating glutamate uptake [155]. In a Huntington’s disease (HD) mouse model, SIRT-4 expression varies in different brain regions: it was increased in the striatum, decreased in the cortex, but remained unaltered in the cerebellum [156]. Thus, the correlation between SIRT-4 and neurodegeneration is still vague and it is not possible to take a stand regarding the meaning of an SIRT-4-directed pharmacological strategy in neurodegenerative disorders.
2.4.3. SIRT-4 Modulators in T2D
SIRT-4 was reported to inhibit insulin secretion [157,158,159]. The molecular mechanism relies on SIRT-4 transferring an ADP-ribosyl group to glutamate dehydrogenase (GDH), thereby decreasing its enzymatic activity. In turn, GDH inhibition impairs ATP generation and insulin secretion. Thus, the pharmacologic inhibition of SIRT-4 could be proposed as a therapy for T2D.
2.5. SIRT-5 Modulators as Therapeutics
SIRT-5 is located in the mitochondria and controls several physiological pathways, including the promotion of ammonia detoxification, fatty acid β-oxidation and ketone body production and the regulation of energy production [160]. Several compounds were synthesized and tested as SIRT-5 modulators, but only a few of them are specific and have already been tested on diseases.
2.5.1. SIRT-5 Modulators in Cancer
As for the other sirtuins, SIRT-5 also has a controversial role in cancer, being proposed as an oncogene in some cancers and as an oncosuppressor in others [160].
The most promising and selective SIRT-5 inhibitor is DK1-04e, which is able to suppress mammary tumor growth in vitro and in vivo [161]. IDH2, an NADPH-generating enzyme, was identified as an SIRT-5 substrate in mammary tumors: SIRT-5 was demonstrated to desuccinylate and activate IDH2 [161]. DK1-04e was obtained through chemical modifications of a thiosuccinyllysine peptide, known to inhibit SIRT-5; DK1-04e has an IC50 of 0.34 μM against SIRT-5, and proved to be very selective, since it showed no inhibition of SIRT-1–3 and -6 deacylation activity at 83.3 μM [161]. In addition, Yan and colleagues showed that the SIRT-5 selective inhibitor NRD167 can inhibit AML cell proliferation, both in vitro and in vivo [162]. Cell apoptosis induced by SIRT-5 inhibition is preceded by reductions in oxidative phosphorylation and glutamine utilization, and an increase in mitochondrial superoxide [162]. NRD167 was synthesized starting from a parent known SIRT-5 inhibitor [162].
On the contrary, in pancreatic ductal adenocarcinoma (PDAC), the SIRT-5 activator MC3138, by promoting GOT1 deacetylation and inhibition, showed a reduction in cell viability in vitro and a reduction in tumor growth in vivo in combination with gemcitabine [163]. C3138 exhibited a selective activation of SIRT-5: it activated SIRT5 at 10 μM concentration, whereas it did not affect SIRT-1 or SIRT-3 activity at 100 μM concentration [163].
2.5.2. SIRT-5 Modulators in Neurodegenerative Disorders
No reports are available on the pharmacological modulation of SIRT-5 in neurodegenerative disorders. However, the data obtained by Liu and colleagues suggest that the pharmacological activation of SIRT-5 could represent a good approach to reduce oxidative stress in PD [164]. Similarly, another study suggests that the pharmacological activation of SIRT-5 could ameliorate the progression of AD, since SIRT-5 has a role in promoting autophagy [165].
2.5.3. SIRT-5 Modulators in T2D
Regarding the role of SIRT-5 in T2D, Wang and colleagues showed that SIRT-5 deficiency in brown adipose tissue (BAT) is followed by hyper-succinylation on GDH, SDHA, and UCP1, reducing their activity [166] and causing glucose intolerance. Thus, SIRT-5 activators may be a promising approach to treat T2D via BAT activation. On the contrary, Ma and colleagues showed that SIRT-5 downregulation promotes insulin secretion from β-cells, suggesting inhibition of SIRT5 as a therapeutical approach for T2D [167].
2.6. SIRT-6 Modulators as Therapeutics
SIRT-6 is a nuclear sirtuin and its role in the regulation of different processes is being increasingly recognized, covering many different functions, including energy metabolism derived both from glucose and lipids, DNA repair, aging, inflammation and immunity. Both activators and inhibitors of SIRT-6′s activity have been identified and evaluated, in in vitro and in vivo experimental settings.
2.6.1. SIRT-6 Modulators in Cancer
Depending on the different cancer type, or even on the different cancer stage, SIRT-6 has been reported to act as an oncopromoter or oncosuppressor. These studies have been performed by overexpressing or silencing SIRT-6 expression and, depending on the obtained results, inhibitors or activators have been evoked as promising strategies.
Compound UBCS039 is an SIRT-6 activator [168], showing no statistically significant effects on basal SIRT-1, -2, and -3 deacetylation activities and a ≈2-fold increase in SIRT-5 desuccinylation activity when added at 100 μM concentration [168]. UBCS039 [168] induced cell death and reduced cell proliferation in cancer cell lines of different origin, including non-small cell lung, colon and epithelial cervix carcinoma, and fibrosarcoma, clearly demonstrating that pharmacological SIRT-6 activation triggers an autophagy-related cell death [169]. At the molecular level, SIRT-6-mediated autophagy was triggered by an increase in ROS levels, which, in turn, resulted in the activation of the AMPK-ULK1-mTOR signaling pathway [169].
MDL-800, another SIRT-6 activator (developed as hit-to-lead optimization process from compounds AN-988/40889624 and AH-487/41802661, discovered by structure-based studies, see Section 3.6), reduced proliferation in vitro, without inducing cell death, and suppressed tumor growth in vivo in a mouse model of hepatocarcinoma [170]. In addition, MDL-800 reduced proliferation in non-small cell Lung cancer cells, and it was suggested that SIRT-6 activation may be promising as a therapeutic approach alone or in combination with epidermal growth factor receptor tyrosine kinase inhibitors [171]. MDL-800 potently activated SIRT-6 at ~10 µM but showed no activity toward SIRT-1, SIRT-3, SIRT-4, and HDAC1-11 at concentrations up to 50 or 100 µM [170].
MDL-811, derived from structural optimization of MDL-800, proved to represent a possible strategy against colorectal cancer, by tests conducted on the HCT116 cell line, patient-derived xenografts as well as on a spontaneous model of this cancer [172]. Mechanistically, Cytochrome P450 family 24 subfamily A member 1 was identified as a new downstream target gene of SIRT-6 in colorectal cancer [172]. As MDL-800, MDL-811 also potently enhanced the deacetylase activity of SIRT-6 but showed little effect on other histone deacetylase enzymes at concentrations up to 100 μM [172].
Compound 12q, designed as optimized analogue of the prototype Hit20, which was identified via structure-based studies (see Section 3.6), significantly inhibited the proliferation and migration of pancreatic ductal adenocarcinoma (PDAC) cells in vitro and it also markedly suppressed the tumor growth in a PDAC tumor xenograft model [173]. Compound 12q exhibited weak or no activity against other HDAC family members as well as 415 kinases, indicating good selectivity for SIRT-6 [100,173].
In the same cancer model, SIRT-6 inhibitors identified by a VS (see Section 3.6), having a quinazolinedione and salicylate-like structure, reduced proliferation and increased sensitivity to gemcitabine [174,175]. The most promising inhibitor with quinazolinedione structure showed a poor selectivity towards SIRT-1 and -2 [174]. Conversely, the family of SIRT-6 inhibitors with a salicylate-like structure, showed a selectivity ranging approximately from 10 to 30 folds towards SIRT-1 and -2 [175].
In addition, it was suggested that a synthetic lethal approach, enhancing DNA damage while concomitantly blocking SIRT-6-mediated repair responses through the use of SIRT-6 inhibitors, provides the rationale for the clinical evaluation of SIRT-6 inhibitors in the treatment of leukemia [176].
2.6.2. SIRT-6 Modulators in Neurodegenerative Disorders
In PD, no studies in vitro or in vivo have been performed until now using an SIRT-6 modulator. However, it has been observed that in an MPTP-induced PD mouse model, brain-specific SIRT-6 knockout conferred neuroprotection, while SIRT-6 overexpression caused a more severe pathology to the mice. In the same study, Nicotine was proved to decrease SIRT-6 expression both in vitro and in vivo, and as a consequence to protect neurons from apoptosis [177]. In addition, inhibition of SIRT-6 was suggested to be a promising strategy to ameliorate PD and neurodegeneration, given that nicotine reduces the abundance of SIRT-6 in neuronal culture and brain tissue, this mediating the nicotine-induced neuroprotection [177].
One SIRT-6 inhibitor with quinazolinedione structure (named compound 1 and identified via in silico screening, see Section 3.6) [174] was administered following both a “preventive” and a “therapeutic” protocol in a mouse model of Experimental Autoimmune Encephalomyelitis (EAE), regarded as an MS animal model. No significant effects were obtained in the therapeutic protocol. Instead, SIRT-6 inhibition strikingly delayed EAE onset, impaired dendritic cell migration, downregulated pathogenic T cell inflammatory responses. Therefore, SIRT-6 inhibitors were suggested to represent novel therapeutic agents for the treatment of early stages of MS, or of other autoimmune disorders [178].
2.6.3. SIRT-6 Modulators in T2D
SIRT-6 inhibitors identified by VS (see Section 3.6) in vitro induced GLUT1 upregulation and consequent augmented glucose uptake in L6 rat myoblasts and BxPC3 cells [179]. When tested in a murine model of T2D, one SIRT-6 inhibitor having a quinazolinedione scaffold, improved glucose tolerance and reduced plasma levels of insulin, triglycerides, and cholesterol [180].
The potential beneficial effect of using SIRT-6 inhibitors in T2D came also from an independent study, in which Compound 6d (a newly identified SIRT-6 inhibitor) was used in a mouse model of T2D and significantly increased the level of glucose transporter GLUT-1, thereby reducing blood glucose [181]. Compound 6d was identified by performing a screening study against an in-house chemical library containing about 2000 compounds by Fluor de Lys assay and showed good selectivity over other deacetylases including SIRT-1-3 and HDAC1-11 [181].
2.7. SIRT-7 Modulators as Therapeutics
SIRT-7 is localized in the nucleus, where it is involved in the activity of RNA polymerase I and it is important for cell viability [182,183]. Very few reports are available for this sirtuin. The interest in SIRT-7 has increased in the last 10 years, but a lot of work still needs to be accomplished to understand its role in the etiology of pathologies.
2.7.1. SIRT-7 Modulators in Cancer
Zhang and colleagues showed that compounds 2800Z and 40569Z (identified by VS, see Section 3.7) are specific SIRT-7 inhibitors exerting an antiproliferative effect on liver cancer in vitro and in vivo [184].
In addition, Kim and colleagues identified compound 97491 as a specific SIRT-7 inhibitor able to induce apoptosis in uterine sarcoma [185]. This compound was identified by an in vitro enzyme activity assay using compounds obtained from the Korea chemical bank [185].
2.7.2. SIRT-7 Modulators in Neurodegenerative Disorders
SIRT-7 possesses a major role in numerous neuronal pathways. For instance, it regulates rRNA synthesis and assembly of ribosomes via the changes in NAD+/NADH ratio [183] and promotes the repair mechanism of non-homologous DNA damage [186]. Nevertheless, there are no reports explaining its role in neurodegenerative diseases.
2.7.3. SIRT-7 Modulators in T2D
The role of SIRT-7 in T2D is still obscure and there are no reports on the use of SIRT-7 inhibitors in this disease. Li and colleagues showed that the overexpression of SIRT-7 increases hyperglycemia and renal dysfunction in rats [187]. These data suggest that the use of SIRT-7 inhibitors could help T2D treatment.
2.8. Pan-Sirtuin Modulators as Therapeutics
Several pan-sirtuin modulators have been identified and assessed in different cellular and animal settings. The modulators identified have mainly an inhibitory action, since the discovery of activators is more complex and just a few SIRT activators have been discovered in the past decades. The main issues related to the use of non-specific sirtuin inhibitors are represented by the ineffectiveness of the treatment due to the inhibition of two or more sirtuins exerting an opposite effect in the cells [89], until further side effects on non-sirtuin targets are found, which may cause over-toxicity. Thus, in principle, it is preferable not to use pan-sirtuin modulators and, in case no other options are available, extra caution may be needed with these modulators.
2.8.1. Pan-Sirtuin Modulators in Cancer
Cambinol (or NSC-112546) is an SIRT-1 and SIRT-2 inhibitor with equal efficiency (IC50-SIRT1 = 56µM, IC50-SIRT2 = 59 µM) and a less efficient SIRT-5 inhibitor (IC50-SIRT5 > 300 µM). Cambinol is a chemically stable compound related to splitomicin, able to reduce cell proliferation, migration, and invasion and can induce apoptosis and cell cycle arrest in lung cancer, lymphoma and cervical cancer [188], multiple myeloma [189], hepatocarcinoma, neuroblastoma, and breast cancer [190,191,192].
Tenovin-1 and -6 inhibit SIRT-1 and SIRT-2 and decrease cell growth in Burkitt’s lymphoma and melanoma cells [193], gastric cancer cells [194], and NSCLC [195]. In addition, promising results were obtained in decreasing tumor growth in leukemia and melanoma [113,196]. Tenovins were originally identified through a high-throughput screen designed to detect compounds that activate the tumor suppressor p53 [193].
Schnekenburger and colleagues showed that their newly synthesized Compound 18 (identified via in silico screening, see Section 3.8) inhibits SIRT-1 and SIRT-2 with an anti-proliferative effect in glioma cells, both in in vitro and in vivo settings [197].
Compound 3g, designed by Laaroussi and colleagues, proved to be an SIRT-1 and SIRT-2 inhibitor with cytotoxic effects on leukemia, colorectal, lung and breast cancer cell lines [198].
Sirtinol, an SIRT-1 and SIRT-2 inhibitor, originally identified through a high-throughput, phenotypic screen in cells [199] determined a senescence-like growth arrest and decreased activation of the RAS-MAPK pathway in breast and NSCLC [200]. In breast cancer, Sirtinol induces cell death and destabilizes the Slug protein, thus antagonizing the effect on the metastasis capabilities of basal-like breast cancer [50,201]. In addition, Sirtinol enhanced chemosensitivity to camptothecin and cisplatin in the prostate and cervical cancer cells, resulting in a significant reduction in viable cells due to enhanced apoptotic cell death [200,202].
Salermide and its two analogs, Compound 4b and 6a, identified by Rotili and colleagues applying hit-to-lead structural variations, showed a potent anti-proliferative effect on leukemia, lymphoma, colon, breast, NSCLC and glioblastoma cancer cells [90,203,204].
BZD9L1 is another SIRT-1 and SIRT-2 inhibitor able to induce apoptosis in colorectal, leukemia, and breast cancer cell lines [205].
Compounds 27 and 30, discovered by Mellini and colleagues upon synthesis of a compound library of 30 pseudopeptides, were reported to inhibit SIRT-1, -2, and -3 and they are able to arrest cell growth in breast and lung cancer cells [54].
JH-T4 inhibits SIRT-1, -2, and -3 and it has an antiproliferative effect on breast, colorectal, and lung cancer cells. JH-T4 was identified by carrying out structure–activity relationship studies based on the structure of TM, a thiomyristoyl lysine compound [206]. [206]
George and colleagues showed that the dual inhibitor of SIRT-1 and SIRT-3, 4-Bromo resveratrol, obtained with a chemical modification from resveratrol [207] is able to reduce cell growth and induce apoptosis in melanoma cells in vitro and in vivo through metabolic reprogramming [208].
Hui and colleagues designed and synthesized a series of 2-(4-acrylamidophenyl)-quinoline-4-carboxylic acid derivatives as putative SIRT-3 inhibitors, the best of which is Compound P6, which acts also on SIRT-1 and SIRT-2. This compound is able to arrest the cell cycle of mixed-lineage leukemia (MLL) cells in G0/G1 phase [209].
Finally, nicotinamide, the pan-sirtuin inhibitor par excellence, exerts antiproliferative effects and induces apoptosis in leukemic, oral squamous cell carcinoma, lung, and prostate cancer cells [210,211,212,213].
2.8.2. Pan-Sirtuin Modulators in Neurodegenerative Disorders
The only pan-sirtuin inhibitor reported in the literature with an effect on neurodegenerative disorders is nicotinamide, which seems to have the ability to restore cognitive deficit by reducing tau phosphorylation in mouse models of AD. The mechanism of action mediating the nicotinamide-induced beneficial effect is unknown. However, the increased expression of SIRT-2 with age suggests that the preferential target of nicotinamide in AD is SIRT-2 [214,215]. Nicotinamide was also evaluated in a transgenic mouse model of HD, and it caused the improvement of motor deficits [216]. The detailed mechanism of action was not investigated; nevertheless, the treatment with nicotinamide is able to increase the expression of brain-derived neurotrophic factor (BDNF) and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α).
2.8.3. Pan-Sirtuin Modulators in T2D
In INS-1 beta pancreatic cells, nicotinamide was reported to reduce the high glucose- and palmitate-induced cell death. The mechanism of action is not fully understood; however, the knockdown of SIRT-3 or SIRT-4 determines the same effect as nicotinamide treatment [217], suggesting that nicotinamide’s protection may be due to the inhibition of SIRT-3 and/or SIRT-4. Therefore, the use of inhibitors for these two sirtuins may represent a promising therapeutic approach.
3. Virtual Screening Strategies Guiding the Discovery of Sirtuins’ Modulators
During the last years, the rational design of novel bioactive compounds towards a specific biological target, such as enzymes and G-protein-coupled receptors (GPCRs), was deeply accelerated by computational strategies. In this context, the search of sirtuins’ modulators can be pursued, taking advantage of a consistent number of experimental data, such as X-ray crystallographic information, which turn in a viable support for drug design. In addition, collecting a number of known modulators allowed us to develop further ligand-based strategies, such as pharmacophore modelling, to support the preliminary evaluation of a high number of compounds, prior to VS.
Herein, we report and discuss the computational studies and VS approaches so far applied towards the discovery of novel sirtuin modulators, as described in the literature.
The analyzed articles were retrieved by means of a cross-search on PubMed [218] and Web of Science [219]. The queries combined the terms “sirtuin” AND “virtual screening” (VS). The merge of the two results produced as outcome a list of 77 articles, then refined to exclude non-relevant data (review articles, VS conducted on other targets, articles that do not contain VS, etc.) and duplicates. A similar method was followed using the queries “sirtuin” AND “docking” to include docking-based VS. Four articles were added. In total, 39 articles containing VS experiments applied to one or more sirtuins were obtained. Figure 2 summarizes the article selection procedure (last check 14-04-22), while Table 3 reports the applied computational methods and the obtained results.
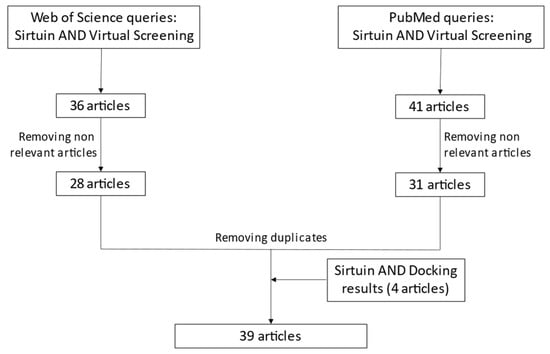
Figure 2.
Workflow for article selection.
3.1. Virtual Screening of SIRT-1 Modulators
To our knowledge, the first application of VS techniques to the discovery of SIRT-1 binders appears in 2008, when Huhtiniemi et al. performed a pharmacophore-based VS [220] using a previously reported structure-based model for SIRT-1 inhibitors [221]. A new oxadiazole-carbonylaminothiourea scaffold was individuated, and 47 analogues bearing this group were assessed in vitro against SIRT-1 and SIRT-2 [220]. The most potent compound showed a comparable potency with respect to the reference compound (an EX-527 analogue). In 2009, Sugunadevi Sakkiah et al. [222] explored the allosteric activation of SIRT-1 with a similar approach. In this case, the pharmacophore was modelled using three known SIRT-1 activators belonging to the imidazothiazoles class, and the best hypothesis was used to screen the Maybridge database [223]. Seven compounds were selected as promising hits. A few years later, the same group combined an analogous ligand-based pharmacophoric screening with the use of a Bayesian model [224]. A drug-like database was screened with both the models, and 16 novel candidates were selected as best hits based on the calculation of their energy gap (DFT application). A ligand-based (LB) pharmacophore model was also used in 2015 by Vyas’ group [225] to screen the Zinc database [226] in search of new SIRT-1 activators. The model was generated using the structures of 10 known SIRT-1 activators, and top hits were submitted to docking in an HM of the target. A few years later, A. Azminah et al. combined the use of ligand-based and structure-based pharmacophoric models to identify novel SIRT-1 activators [227]. In particular, a structure-based pharmacophore was generated by the crystal structure of SIRT-1 co-crystalized with a small molecule sirtuin-activating compound (STAC, PDB code: 4ZZJ) [228]. The ligand-based model was instead built using 12 actives retrieved by the literature. A database of 1377 compounds by the HerbalDB [229] was screened with the two pharmacophore hypotheses. The ligand-based approach retrieved quinine, quinidine and gartarin as best candidates for SIRT-1 activation. The SB model individuated mulberrin as best hit. In vitro studies confirmed the bioactivity of these compounds. A similar approach was chosen by Pulla’s group [57] in 2016. In particular, an energy-based pharmacophore model was built on the crystal structure of SIRT-1 in complex with an EX527 analogue and NAD+ (PDB code: 4i5i) [230], while a ligand-based pharmacophore model was developed using a training set of 79 molecules. The pharmacophore features were used to pre-filter a large database from Asinex [231] and a smaller inhouse library. The resulting compounds were then subjected to molecular docking using a multi-step procedure. A benzimidazole derivative was identified as a promising scaffold on the basis of in vitro and in vivo screenings (see Compound 17 mentioned in Section 2.2.1).
In 2012, Alvala et al. [52] performed an SBVS of an in-house database against a homology model of the SIRT-1 catalytic domain. A new isonicotinamidic scaffold was used, and three putative inhibitors were selected for in vitro testing. The positive outcome of the analysis allowed further optimization, leading to a new potent acridinedione with anticancer potential (as mentioned in Section 2.1.1). In 2014 [232] the same group used a structure-based approach to identify novel SIRT-1 modulators. In particular, the crystal structure of SIRT-1 in complex with an analogue of EX-527 (a SIRT-1 inhibitor) and NAD+ (PDB code 4I5I) [230] was used to screen the Asinex database [231], resulting in the individuation of two promising inhibitors. In the same article, a similar procedure was followed for the design of SIRT-1 activators, using a previously reported homology model [233] of the target as structural information. A new activator was proposed, and the compound efficacy was confirmed by in vitro tests. A few years later, Padmanabhan et al. [234] used the hSIRT-1 crystal structure [235] in complex with ADPR (pdb code: 4KXQ) to screen a Drug Bank [236] library containing 1716 compounds. Two scaffolds (diphenyl derivatives and oxycoumarin derivatives) were selected and submitted to MD simulation analysis. Four compounds resulted to inhibit SIRT-1 deacetylase activity in vitro. More recently, Wössner et al. [59] proposed an iterative in silico-in vitro screening procedure for the individuation of novel SIRT-1 inhibitors. Initially, an SBVS against the EX-527-SIRT-1-NAD+ complex [230] (PDB ID 4I5I) was performed. A first series of in vitro experiments validated the individuation of a thienopyrimidone with a thiocyanate moiety as promising scaffold. Based on this result, phenyl thiocyanates-containing compounds were selected by the Princeton BioMolecular Research Compound collection [237] and the resulting 113 thiocyanates were docked in the SIRT-1 active site. The in vitro test of the best docked compounds led to the individuation of an improved thienopyrimidone-thiocyanate. Interestingly, these compounds showed high selectivity towards SIRT-2 and -3. In addition, the compounds discovered showed anti-cancer activity in vitro (see Section 2.2.1).
A similar method was used by Tugba Ertan-Bolelli and Kayhan Bolelli [238] to identify novel hSIRT-1 activators. In particular, they performed an SBVS of a zinc [226] library on the SIRT-1 allosteric domain, individuating seven putative activators. Among them, acebutolol was selected for MD studies.
A less classic approach is reported in the study of Wang et al. [239], which developed an innovative sequence-based prediction model. This method relies on the idea that as the sequence determined the three-dimensional fold of the target, it can be correlated to the ligand bioactivities with no need for structural information. A general (target-unspecific) model was therefore developed with a training set of protein–ligand interactions, using a support vector machine (SVM) [240,241,242] approach. The model output was the prediction of unknown protein–ligand interactions using as input a protein sequence and a small molecule library. In the present case, a drug-like Specs database [243] and SIRT-1 sequence were used. Five novel inhibitors were individuated by in vitro tests, highlighting the success of this strategy. In 2016, Sun et al. [244] used Inductive logic programming (ILP) to screen a large database of compounds belonging to the Traditional Chinese Medicines-Taiwan database [245] and Traditional Chinese Medicine Integrated Database [246]. Briefly, the used ILP-based software combined both the input data (ligand information by the literature) and background knowledge to produce an inhibitor structure pattern, which was then used as a query for the screening. In 2019, a contest-based study was described by Chiba et al. [247]. Different groups were proposed to prioritize a subset of putative SIRT-1 inhibitors among a 2.5-million-compound database through different VS methods, with the aim to perform a comparative study. The experimental testing of approximately 50% of the proposed compounds led to the discovery of seven structurally distinct hits.
3.2. Virtual Screening of SIRT-2 Modulators
In 2008, W. Sippl et al. performed a first example of VS on SIRT-2 [248], combining ligand-based and structure-based techniques. In particular, the Chembridge database [249] was initially filtered according to lead-like features and to similarity with respect to Cambinol, a known SIRT-2 inhibitor (see Section 2.2.1 and Section 2.8.1). The resulting compounds were then docked in the SIRT-2 catalytic pocket (crystal structure in the apo-form, 1J8F [6]), and compounds showing a H bond to Gln167 (a key residue) were retained. Compounds with a low logP were prioritized, and the resulting molecules were submitted to in vitro validation. Five barbiturate and thiobarbiturate derivatives were proposed. The authors further optimized this scaffold in 2012 through a similar multi-step VS procedure [250]. Most active thiobarbiturates from the initial work were selected, and their fingerprints were used as queries to screen the Chembridge database [249]. The results were filtered according to drug-like properties and docked in the apo-SIRT-2 active site [6] (PDB code 1J8F). Top hits were subjected to additional in silico and in vitro studies, highlighting an ameliorate potency with respect to the previously proposed analogues. A similarity-based approach was again followed by the same group to design novel splitomicin-related SIRT-2 inhibitors [105]. The authors performed an SAR study on the splitomicin scaffold and reported the best activity for β-aryl derivatives of splitomicin. The fingerprints of the most active compounds were used as queries to screen the Chembridge [249] database. The top-ranked compounds were docked in the apo form of SIRT-2 [6] (PDB code 1J8F) and four candidates were evaluated in vitro (the best inhibitors, namely, 8c and (R)-8c, showed potent anti-cancer effects, see Section 2.2.1). The outcomes revealed that the substitution of the original lactone with a lactam is well tolerated.
A pure structure-based technique was instead chosen by P. Sivaraman et al. [251], which virtually screened an NCI [252] Diversity Set using the apo-form of SIRT-2 (PDB code 1J8F) [6] as a docking template. Every ligand was submitted to 150 separate docking calculation, using the Lamarckian genetic algorithm local search (GALS) method: the system generates a set of solution and propagates the most suitable answers to the following generations. The top hits were tested in vitro, revealing a new potent nucleoside-like inhibitor of SIRT-2. In 2016, Sacconnay et al. [253] performed an SBVS against the ADPR-bound form of SIRT-2 (PDB code 3ZGV) [254] in search of new SIRT-2 inhibitors. A 197,477-compound library from Specs was employed to this aim. The top ranked compounds were filtered according to drug-like properties and submitted to a cluster analysis. The best ranked compounds of each cluster were purchased and evaluated in vitro, leading to the identification of two novel scaffolds. In particular, the 5-benzylidene-hydantoin was reported to be a promising hit with anti-cancer potential.
In some cases, SB screening has been coupled with MD to generate suitable conformation of the target. This requirement has been dictated by the unavailability of an inhibitor-bound form of SIRT-2 until 2015 [112] (PDB code 4RMG). In 2004, Tervo et al. [255] generated an artificial SIRT-2 conformation by submitting to MD the apo-form of SIRT-2 [6] (PDB code 1J8F) and calculating the average structure on the equilibrated portion of the dynamics. The generated construct exhibited an enlargement of the binding pocket, whose interaction fields and properties were computed. The Maybridge database [223] was then screened according to these properties (hydrophobicity, shape, favourable interactions with polar probes). The most promising compounds were submitted to docking in SIRT-2 structure, and the compounds with a similar pose and key interactions with respect to sirtinol were retained. Five compounds resulted to be active against SIRT-2 by in vitro tests. The same authors used a similar approach [256] again in 2006. After the generation of the MD conformation, the docking of a known inhibitor was performed, identifying key interactions for binding. According to this analysis, two queries were developed and used to screen the Maybridge Screening Collection [223] and the Lead Quest databases. Four out of the eleven tested compounds resulted to be active in vitro at the micromolar concentration. One of these compounds bears a new scaffold, not previously reported in relation to SIRT-2 inhibition, featuring the indole ring as the main core of the inhibitor. In 2012 [257], Sakkiah et al. generated an inhibitor-bound conformation by docking sirtinol in the NAD+ binding site of SIRT-2 apo-form [6] (PDB code 1J8F) and submitting the complex to 5-ns MD simulation. Three different representative conformations of the protein were obtained by means of a cluster analysis. They were merged together to form a single dynamic pharmacophore model, which was then used to screen the CHEMDIV database [258]. According to their drug-likeliness, ADMET properties and docking analysis, the authors proposed 21 leads as putative SIRT-2 inhibitors. In the same year, the same group combined this coupled MD-SB approach with ligand-based techniques [259]. In particular, a pharmacophore model built on a training set of 19 inhibitors was used to screen various databases (NCI [252], Maybridge [223] and Chembridge [249]), and the best candidates were filtered according to drug-like and calculated ADMET parameters. Then, an MD-derived structure of the sirtinol-SIRT-2 complex was used as a docking template to refine the results. The candidates with a comparable or higher score with respect to known inhibitors of SIRT-2 were retained, prioritizing the poses which established significant H-bonds. Twenty-nine candidates were individuated. In 2019, a similar approach was used by Eren et al. [97]. An LB pharmacophore was developed starting from a series of 31 SirReal analogues and applied to screen a 13-million-compound ZINC drug-like database. The retrieved molecules were then submitted to docking, this time in the finally available inhibitor-bound form of SIRT-2 (SIRT-2-SirReal analogue-NAD+, PDB code: 5DY4) [260]. The Glide [261,262] Virtual Screening Workflow (VSW) was used to this aim. The MM-GBSA technique was applied to increase the accuracy of the predicted pose. According to a PCA-based cluster analysis, 31 compounds were selected for in vitro evaluation. Two compounds showed good in vitro inhibitory potency and a certain degree of selectivity toward SIRT-1,-3,-5 (see Section 2.2.2).
In 2021, Kessler et al. chose a machine learning approach with the aim to identify new selective inhibitors of SIRT-2 [263]. In particular, a Multilayer Perceptron (MLP) model [242] was built using a set of 234 inhibitors of SIRT-2 and 234 putative inactive molecules. The model was applied to screen the ZINC [226] collection of FDA-approved drugs, in a drug-repositioning perspective. The ML approach was then validated in silico through docking in SIRT-2 crystal structure (5YQL) [99]. The study reveals a promising ligand-based technique, which may be applied to other targets, even in the absence of a target 3D structure. In the same year, Khanfar and Alqtaishat [264] reported an interesting combination of pharmacophore-based screening and QSAR techniques. An ensemble of structure-based pharmacophoric models was built using a set of 18 SIRT-2 co-crystals. Nineteen models were regarded as satisfying by means of ROC curves [265] analysis. In parallel, a set of SIRT-2 inhibitors was submitted to the calculation of their physicochemical descriptors. Genetic Function Algorithm [266] and Multiple Linear Regression analysis [267,268,269] were used to select the best combination of pharmacophores and descriptors, building a single mathematical equation able to predict and explain activities. In particular, the resulting equation reports three descriptors and the fit value of the most suitable pharmacophore model. The latter was used to screen the AnalytiCon Discovery database of purified natural products [270]. The matching compounds were ranked according to their predicted IC50, according to the integrated QSAR model. Among the 10 bioassayed compounds, two candidates (asperphenamate and salvianolic acid B) showed inhibitions with IC50 in the low micromolar concentration.
Table 3.
Perspective of the computational studies leading to the identification of selective and/or pan-Sirtuins modulators (shown in green). The chemical structure and the explored biological activity of the discovered hit compounds are reported. The applied virtual screening (VS) strategy is specified as structure-based (SBVS) or ligand-based (LBVS) methodology. The results are listed based on the sirtuin type (alternatively in gray and cyan), according to SBVS followed by LBVS and combined SB-LB approaches, as chronological order. Data about parasitic sirtuins (depicted in coral) are also detailed, referring to the Leishmania (Lm-Sirt), Trypanosoma cruzi (Tc-Sirt) and Schistosoma mansoni (Sm-Sirt) sirtuins.
Table 3.
Perspective of the computational studies leading to the identification of selective and/or pan-Sirtuins modulators (shown in green). The chemical structure and the explored biological activity of the discovered hit compounds are reported. The applied virtual screening (VS) strategy is specified as structure-based (SBVS) or ligand-based (LBVS) methodology. The results are listed based on the sirtuin type (alternatively in gray and cyan), according to SBVS followed by LBVS and combined SB-LB approaches, as chronological order. Data about parasitic sirtuins (depicted in coral) are also detailed, referring to the Leishmania (Lm-Sirt), Trypanosoma cruzi (Tc-Sirt) and Schistosoma mansoni (Sm-Sirt) sirtuins.
| Year | Ref. | Title | SIRT(s) | Type of VS | Notes | Selectivity Over Other Isoforms | Experimental Validation | Screened Database (n. of Compounds) | Software(s) | Most Active Compound/Proposed Compound | Activator/Inhibitor | Potency [n. of Proposed Compounds by Computational Study] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2008 | [220] | Oxadizole-carbonylaminothioureas as SIRT-1 and SIRT-2 inhibitors | 1 | SBVS | Pharmacophore based | 1,2 | YES | Maybridge and Leadquest libraries | Unity 4.3.1/Sybyl 7.1 |  oxadizole-carbonylaminothiourea | I | 13 μM (IC50, SIRT-1) |
| 2012 | [52] | Novel acridinedione derivatives: design, synthesis, SIRT-1 enzyme and tumor cell growth inhibition studies. | 1 | SBVS | Target: HM | Not tested | YES | In house database (2500) | Glide, Gold, AutoDock 4.0 |  acridinedione derivatives | I | 0.25 μM (IC50) |
| 2014 | [232] | Structure-based drug design of small molecule SIRT-1 modulators to treat cancer and metabolic disorders | 1 | SBVS | Target model for inhibitors: crystal structureTarget model for activators: HM of the allosteric site | Not tested | YES | Asinex (>600000) | Glide 5.0 |  | I, A | 16.35 μM (IC50) |
| 2016 | [234] | Identification of New Inhibitors for Human SIRT-1: An in-silico Approach | 1 | SBVS | Not tested | YES | Drug bank library from ZINC (1716) | AutoDock Vina 1.1.2 |  diphenyl and oxycoumarin derivatives | I | 77.7% inhibition @5μM | |
| 2020 | [59] | Sirtuin 1 Inhibiting Thiocyanates (S1th)-A New Class of Isotype Selective Inhibitors of NAD(+) Dependent Lysine Deacetylases | 1 | SBVS | Iterative in vitro-in silico screenings | 2,3,5 | YES | Small library of previously identified putative SIRT-1 inhibitors | GOLD 5.6 |  Thiocyanates | I | 5.2 μM (IC50) |
| 2021 | [238] | In Silico Design of Novel SIRT-1 Enzyme Activators for the Treatment of Age-related Diseases and Life Span | 1 | SBVS | Not tested | NO | Zinc (150 000) | Information not available |  acebutolol and others | A | NC [7] | |
| 2009 | [222] | Pharmacophore Mapping and Virtual Screening for SIRT-1 Activators | 1 | LBVS | Pharmacophore based | Not tested | NO | Maybridge | HipHop module/CATALYST | isothiazole scaffold benzimidazole scaffold | A | NC† [7] |
| 2014 | [224] | Theoretical approaches to identify the potent scaffold for human SIRT-1 activator: Bayesian modeling and density functional theory | 1 | LBVS | Bayesian model, pharmacophore model | Not tested | NO | Maybridge (60,000), Chembridge (50,000), NCI (200,000), and ChemDiv (700 000) | Discovery Studio v 3.1 | Various | A | NC [16] |
| 2016 | [244] | Ligand-based virtual screening and inductive learning for identification of SIRT-1 inhibitors in natural products | 1 | LBVS | Inductive logic programming | Not tested | NO | Traditional Chinese Medicines-Taiwan Database and Traditional Chinese Medicine Integrated Database (1 444 880) | DMax Chemistry Assistant software | Various | I | NC [3] |
| 2015 | [225] | Ligand and structure-based approaches for the identification of SIRT-1 activators. | 1 | LB-SB | LBVS: Pharmacophore basedSBVS Target model: HM of Sirt-1 | Not tested | NO | ZINC database | DISCOtech, GASP/SYBYL X 1.2 (pharmacophore)Surflex-Dock/SYBYL X 1.2 (docking) | Various | A | NC [2] |
| 2016 | [57] | Energy-Based Pharmacophore and Three-Dimensional Quantitative Structure--Activity Relationship (3D-QSAR) Modeling Combined with Virtual Screening To Identify Novel Small-Molecule Inhibitors of Silent Mating-Type Information Regulation 2 Homologue 1 (SIRT-1). | 1 | LB-SB | LB and SB pharmacophore models combined with docking-based VS | Not tested | YES | ASINEX (5 000 000), in-house (971) | PHASE 3.4/Maestro 9.3 (pharmacophore) Glide 5.8/Maestro 9.4 (docking) |  benzimidazole derivative | I | 4.34 μM (IC50) |
| 2019 | [227] | In silico and in vitro identification of candidate SIRT-1 activators from Indonesian medicinal plants compounds database | 1 | LB-SB | LB and SB pharmacophore models | Not tested | YES | Indonesian Herbal Database (1377) | LigandScout 4.2 |  mulberrin, quinine, quinidine, and gartanin | A | 1.14 μM (EC50) |
| 2011 | [239] | Computational screening for active compounds targeting protein sequences: methodology and experimental validation. | 1 | Other | Sequence-based VS | Not tested | YES | SPECS drug-like library (85 000) | LIBSVM |  various | I | 5.72 μM (IC50) |
| 2019 | [247] | A prospective compound screening contest identified broader inhibitors for SIRT-1 | 1 | Other | Contest-based approach | Not tested | YES | ENAMINE (2 459 912) | various |  various various | I | 4.1 μM (IC50) |
| 2004 | [255] | An in silico approach to discovering novel inhibitors of human SIRT-2 | 2 | SBVS | Queries: (SB) features calculated on MD generated conformation + Docking | Not tested | YES | Maybridge | Unity 4.3.1/Sybyl v6.8 (feature-based), GOLD v1.2 (docking) |  Phenol derivatives | I | 56.7 μM (IC50) |
| 2006 | [256] | Discovering inhibitors of human SIRT-2: Novel structural scaffolds | 2 | SBVS | Queries: (SB) features calculated on MD generated conformation + Docking | Not tested | YES | Maybridge Screening Collection and LeadQuest libraries | Unity 4.4/Sybyl 6.9 (feature-based), GOLD 2.0 (docking) |  indole derivatives, phenol derivatives | I | 51 μM (IC50) |
| 2010 | [251] | Design of a novel nucleoside analog as potent inhibitor of the NAD+ dependent deacetylase, SIRT-2. | 2 | SBVS | 1 | YES | NCI Diversity Set II | AutoDock 4.0 |  nucleoside analog (thieno [2,3-d]pyrimidine) | I | 8.7 μM (IC50) | |
| 2012 | [257] | Molecular Docking and Dynamics Simulation, Receptor-based Hypothesis: Application to Identify Novel SIRT-2 Inhibitors | 2 | SBVS | SBVS pharmacophoric model based on MD-generated conformations + docking | Not tested | NO | CHEMDIV database (700 000) | Ligand Pharmacophore Mapping/DS (screening), LigandFit/DS and GOLD (docking) | Various | I | NC [21] |
| 2016 | [253] | 5-Benzylidene-hydantoin is a new scaffold for SIRT inhibition: From virtual screening to activity assays. | 2 | SBVS | 1 | YES | Specs library (197 477) | GOLD v5.2 |  5-Benzylidene-hydantoin | I | 37.7 μM (IC50) | |
| 2008 | [248] | Thiobarbiturates as Sirtuin Inhibitors:Virtual Screening, Free-Energy Calculations, and Biological Testing | 2 | LB-SB | LB: similarity based | 1 | YES | Chembridge database (∼328 000) | MOE (fingerprints) GOLD 3.2 (docking) |  thiobarbiturates | I | 9.1 μM (IC50) |
| 2008 | [105] | Structure-activity studies on splitomicin derivatives as sirtuin inhibitors and computational prediction of binding mode | 2 | LB-SB | LB: similarity based | Not tested | YES | Chembridge (∼328 000) | MOE (fingerprint), GOLD 3.0 (docking) |  lactame analogues of beta-Phenylsplitomicins | I | 6.4 μM (IC50) |
| 2012 | [259] | Pharmacophore modeling and molecular dynamics simulation to identify the critical chemical features against human SIRT2 inhibitors | 2 | LB-SB | LB: pharmacophore basedSB: VS on MD-derived protein conformation | Not tested | NO | NCI (5672), Maybridge (26 490), Chembridge (17 885) | Discovery Studio v2.5 (pharmacophore), GOLD (SB) | Not reported | I | NC [29] |
| 2012 | [250] | Binding free energy calculations and biological testing of novel thiobarbiturates as inhibitors of the human NAD(+) dependent histone deacetylase SIRT-2 | 2 | LB-SB | LB: similarity-based | Not tested | YES | Chembridge | MOE (fingerprint), GOLD 4.0 (SBVS) |  Thiobarbiturates | I | 1.5 μM (IC50) |
| 2019 | [97] | Pharmacophore modeling and virtual screening studies to identify novel selective SIRT-2 inhibitors | 2 | LB-SB | LB: Pharmacophore-based | 1,3,5 | YES | ZINC drug-like database (13 000 000) | Schrodinger Small-Molecule Drug Discovery Suite (pharmacophore) Glide (SBVS) |  various (triazines and dimethylfuran derivatives among others) | I | 84.28% inhibition @300μM |
| 2021 | [264] | Discovery of Potent Natural-Product-Derived SIRT-2 Inhibitors Using Structure- Based Exploration of SIRT-2 Pharmacophoric Space Coupled With QSAR Analyses. | 2 | LB-SB | SBVS pharmacophoric model combined with (SB+LB) QSAR | Not tested | YES | AnalytiCon Discovery database of purified natural products (5637) | DISCOVERY STUDIOv2.5 |  asperphenamate and salvianolic acid B | I | 1.94 μM (IC50) |
| 2021 | [263] | Targeting Epigenetic Regulators Using Machine Learning: Potential SIRT-2 Inhibitors | 2 | LB-SB | LB: Machine learning model | Not tested | NO | ZINC/FDA library | WEKA (ML), AutoDock Vina/PyRx (SBVS) | Various | I | NC [43] |
| 2015 | [271] | Virtual screening approach of sirtuin inhibitors results in two new scaffolds | 3 | SBVS | 1,2 | YES | ZINC database (> 8 000 000) | Glide v5.8 |  3-oxo-1,4-benzothiazinyl compounds and 4-(1-piperidyliminomethyl)benzene-1,3-dioles | I | 40% inhibition @200 μM | |
| 2013 | [272] | Identification of novel SIRT-3 inhibitor scaffolds by virtual screening | 3 | LB-SB | LB: similarity based | 1,2 | YES | Chembridge EXPRESS-Pick Collection database | Phase 3.2/ Schrodinger (similarity) + Glide 5.7 (SBVS) |  benzoxazoles, furopyrimidines | I | 71% inhibition (200 μM) |
| 2021 | [143] | Structure-Guided Design of a Small-Molecule Activator of SIRT-3 that Modulates Autophagy in Triple Negative Breast Cancer | 3 | SBVS | SB compound optimization | Not tested | YES | ZINC database (> 8 000 000) | Glide |  ZINC03830212 | A | EC50 = 3.25 μM |
| 2017 | [273] | Molecular modeling, dynamics studies and density functional theory approaches to identify potential inhibitors of SIRT-4 protein from Homo sapiens: a novel target for the treatment of type 2 diabetes. | 4 | SBVS | Target: HM | Not tested | NO | LifeChem, Specs, ZINC and MayBridge libraries | Glide/Maestro/Schrodinger v10.1 |  Indole derivative | I | NC [1] |
| 2018 | [274] | Structure-based discovery of new selective small-molecule SIRT-5 inhibitors | 5 | SBVS | SBVS + protein–ligand interaction fingerprint (IFP)-based filter | 2,6 | YES | In-house database (>15,000) | AutoDock Vina |  (E)-2-cyano-N-phenyl-3-(5-phenylfuran-2-yl)acrylamide derivatives | I | 5,59 μM (IC50) |
| 2014 | [179] | Discovery of Novel and Selective SIRT-6 Inhibitors | 6 | SBVS | Target: modified structure of Sirt-6 | 1,2 | YES | ASINEX subset of CoCoCo database | Glide v.5.8 |  salicylate-based derivative | I | 89 μM (IC50) |
| 2015 | [174] | Quinazolinedione SIRT-6 inhibitors sensitize cancer cells to chemotherapeutics | 6 | LB-SB | LB: pharmacophore, similarity based | 1,2 | YES | CoCoCo database | Instant JChem (Chemaxon) ver. 5.11.5 |  Compound 1 as Quinazolinedione prototype | I | 106 μM (IC50) |
| 2018 | [170] | Identification of a cellularly active SIRT-6 allosteric activator | 6 | SBVS | Allosteric site search + docking | Not tested | YES | Various DB (> 5,000,000) | GLIDE software (Schrödinger suite 2009, v5.5) |  AN-988/40889624 | A | EC50 = 173 μM |
| 2020 | [173] | Discovery of Potent Small-Molecule SIRT-6 Activators: Structure–Activity Relationship and Anti-Pancreatic Ductal Adenocarcinoma Activity | 6 | SBVS | Allosteric site search + docking | Not tested | YES | Specs, ChemDiv, Selleck, and MedChemExpress and in-house database | GOLD software |  HIT20 | A | 147.60 (% Peptide Demyristoylation Activation @20 μM) |
| 2021 | [275] | Screening of SIRT-6 inhibitors and activators: A novel activator has an impact on breast cancer cells | 6 | LB-SB | LB: pharmacophore, similarity based | 1,2 | YES | ENAMINE (4 103 115), Chembridge (1 022 400), in house library of 1,4-dihydropyridine derivatives (∼100) | MOE (pharmacophore, similarity, docking), Glide/Maestro/ Schrodinger (docking) |  4H-chromen analogs (A),  1,4-dihydropyridine (I) | A, I | 80 μM (EC50, activator) 60% @200 μM (IC50, inhibitor) |
| 2022 | [184] | Discovery of SIRT-7 Inhibitor as New Therapeutic Options Against Liver Cancer | 7 | SBVS | Target: protein modeling via fold recognition | 1,6 | YES | Chemdiv database (939319) | AutoDock Vina |  Compound 2800Z | I | Inhibition of SIRT-7 deacetylase activity |
| 2011 | [276] | Structure-based development of novel sirtuin inhibitors | 2,3,5,6 | SBVS | 2,3,5,6 | YES | NCI diversity set (1990) | AutoDock Vina |  SIRT-2 specific: tetracyclic compounds | I | 4.8 μM (IC50, SIRT-2) | |
| 2017 | [197] | Discovery and Characterization of R/S-N-3-Cyanophenyl-N’-(6-tertbutoxycarbonylamino-3,4-dihydro-2,2-dimethyl-2H-1-benzopyran-4- yl)urea, a New Histone Deacetylase Class III Inhibitor Exerting Antiproliferative Activity against Cancer Cell Lines | 1,2 | SBVS | 1,2,3 | YES | In-house library (17) | AutoDock Vina |  Compound 18 | 6.2 μM (IC50, SIRT-1) 4.2 μM (IC50, SIRT-2) | ||
| 2018 | [277] | Identification of Bichalcones as Sirtuin Inhibitors by Virtual Screening and In Vitro Testing | 1,2 | SBVS | SBVS on different conformations of sirt-1 and sirt-2 | 3 | YES | pan-African Natural Products Library (463) | GOLD, Glide/Maestro/ Schrödinger v5.8 |  Bichalcones | I | 40.8 μM (IC50, SIRT-1) |
| 2019 | [278] | Structure-based identification of novel sirtuin inhibitors against triple negative breast cancer: An in silico and in vitro study | 1,2,3,4,5,6,7 | SBVS | Not applicable | YES | Plant-derived inhibitors (24), synthetic inhibitors (3) with reported epigenetic modulatory and anticancer potential, PubChem | Glide/Maestro/ Schrödinger |  Sulforaphane (SIRT-1,-5), Kaempferol (SIRT-3) Apigenin (SIRT-6) | I | 12,5 μM (IC50) | |
| 2021 | [279] | In silico Repurposing of Drugs for pan-HDAC and pan-SIRT Inhibitors: Consensus Structure-based Virtual Screening and Pharmacophore Modeling Investigations | 1,2,3,5,6 | SBVS | Consensus SBVS on sirt-1,2,3,5,6 with 3 software | Not applicable | NO | (FDA)-approved drugs (1502) | Glide, FRED v3.3.1.2, AutoDock Vina/PyRx v1.1.2 |  Alosetron, cinacalcet, indacaterol (virtual pan-SIRT I) | I | NC [3] |
| 2008 | [280] | Structure function analysis of Leishmania sirtuin: An ensemble of In silico and biochemical studies | Lm Sir2 | LB-SB | LB: Fingerprint basedSBVS target: LmSir2 HM | 2 | YES | National Cancer Institute (NCI) 3D database (~200 000) | MOE, FlexX/SYBYL 6.9 |  Nicotinamide derivative | I | 1,49 mM (IC50) |
| 2012 | [281] | Anti-Trypanosoma cruzi activity of nicotinamide | Tc Sir2 | LB-SB | LB: fingerprint based (nicotinamide)SBVS target: TcSir2 HM | Not tested | YES | Zinc database | MolDock |  Nicotinamide | I | ~100 μM (IC50) |
| 2014 | [282] | Computational Studies on Sirtuins from Trypanosoma cruzi: Structures, Conformations and Interactions with Phytochemicals | Tc Sir2 | SBVS | Targets: 2 HM of the 2 Tc sirtuins (closed state) | 2,5 | NO | Phytochemicals with antitrypanosomal activity collected by the literature (50) | GOLD v5.1 |  Anacardic acid derivative, aculeatin D, 16-acetoxy-11-hydroxyoctadeca-17-ene12,14-diynylethanoate, vismione D | I | NC [4] |
| 2016 | [283] | In-silico analysis of SIRT-2 from Schistosoma mansoni: structures, conformations, and interactions with inhibitors | Sm Sir2 | SBVS | Target: SmSir2 HM | 2 | NO | ZINC derived database (18 560) | idock |  Phenol derivative | I | NC [4] |
3.3. Virtual Screening of SIRT-3 Modulators
In 2013, H.S. Salo [272] et al. developed a combined LB/SB workflow for the discovery of SIRT-3 inhibitors. The group previously identified some thioacetyl-lysine-containing SIRT-3 inhibitors [284]. The shape and some pharmacophoric features of these molecules were used as a query to pre-filter the Chembridge [249] EXPRESS-Pick Collection database. The obtained small molecules were docked in the active site of SIRT-3 [285] (3GLR), and top hits were selected for in vitro evaluation. Two novel scaffolds were individuated and the 5-amino-2-phenyl-benzoxazole one was selected for further studies. Some years later [271], the same group performed again a VS toward this target with the aim to individuate a putative binding site that may be exploited for the design of allosteric inhibitors. The crystal structure of SIRT-3 bound with a reaction intermediate (PDB ID: 3GLT) [285] was submitted to this computational search [286]. The ZINC [226] database (over 8 million compounds) was then screened with the VSW procedure with Glide [261,262]. The presented strategy produced three moderately potent sirtuin inhibitors, which shows a certain selectivity toward SIRT-2 and SIRT-1. In 2021, exploring the X-ray crystallographic data of SIRT-3 (PDB code = 4FVT) [287] allowed to screen in silico 1.4 million small-molecule compounds, leading to the hit compound ZINC03830212 as an SIRT-3 modulator. This derivative has been modified based on structure-guided studies towards the development of more effective compounds featuring anti-cancer ability, such as Compound 33c [143] (see the previous Section 2.3.1).
3.4. Virtual Screening of SIRT-4 Modulators
To individuate new inhibitors of SIRT-4, a VS campaign was carried out by Choubey et al. [273], together with docking experiments. Quantum Polarized Ligand Docking [288] was employed to optimize the geometry of the docked compounds. Three ligands were selected for binding free energy evaluation and stereoelectronic analysis. An indole derivative was proposed as lead molecule, according to the in silico evaluated parameters.
3.5. Virtual Screening of SIRT-5 Modulators
In 2018, Liu et al. performed an SBVS approach to discover new inhibitors of SIRT-5 [274]. The crystal of SIRT-5 in complex with a bicyclic intermediate [289] (4F56) was chosen as a docking template to screen an in-house library of more than 15,000 compounds. An interaction fingerprint analysis involving several protein–ligand interactions was carried out on the best docked compounds. As Tyr102 and Arg105 were individuated as exclusive residues of SIRT-5, being also key residues for the recognition of acidic acyl-lysine substrates [22,289,290], the compounds which showed H-bonds or electrostatic interactions with these amino acids were selected as potentially selective hits. After visual inspection, 20 candidates were evaluated in vitro.
3.6. Virtual Screening of SIRT-6 Modulators
In 2014, a structure-based screening focused on the SIRT-6 X-ray crystallographic structure (PDB code = 3K35) [291] and the CoCoCo database [292] led to the discovery of selective SIRT-6 inhibitors bearing the salicylate-like structure [179]. These derivatives were further optimized by Sociali et al. in 2015 [174]. In particular, different substructure searches were performed against the CoCoCo database [292], with the aim to fulfill the structural requirements exhibited by the previously identified SIRT-6 modulators [179], and to find more potent analogues. As a result, Compound 1 has been identified and described for the treatment of cancer, MS and T2D (see Section 2.6) [174].
More recently, in Tenhunen et al. [275] (2021), a further combination of SB and LB techniques was pursued towards the identification of novel SIRT-6 modulators. Enamine [293] and Chembridge databases [249] were screened according to a ligand-based pharmacophore model built on a small set of known Sirtuin modulators, such as resveratrol, Ex-527, sirtinol, and quercetin, taking into account as reference compounds those previously described by Parenti and coll [179].
The obtained hits from both the procedures together with a small library of synthetic 1,4-dihydropyridine derivatives were docked in SIRT-6 active site [291] (3K35). In vitro tests individuated 4H-chromen as SIRT-6 activator and a novel inhibitor with a 1,4-dihydropyridine scaffold. 1,4-dihydropyridines were already reported in relation to SIRT-1 [294,295].
In 2018, Huang et al. combined in silico studies and experimental approaches to individuate novel SIRT-6 activators [170]. By using the Allosite method developed by the same group [296], the putative SIRT-6 allosteric site was individuated and used to screen a library of more than 5,000,000 compounds. Among the (20) selected and purchased molecules, Huang et al. identified two hits, AN-988/40889624 and AH-487/41802661, featuring halfmaximal effective concentration (EC50) values of 173.8 ± 1.3 μM and 217.6 ± 1.1 μM (mean ± s.d.) acting as SIRT-6 activators (see Section 2.6.1). More recently, in 2020, compound Hit20 was also discovered as SIRT-6 activator, thanks to molecular docking adopted to screen commercial chemical databases including Specs, ChemDiv, Selleck, and MedChemExpress, as well as an in-house database (see Section 2.6.1) [173]. The prototype has been then further optimized towards the lead analogue compound 12q.
3.7. Virtual Screening of SIRT-7 modulators
The search for putative SIRT-7 modulators via in silico screening or structure-based studies is hampered by the absence of complete X-ray crystallographic data of the enzyme, in the presence or not of ligand. Indeed, the crystal structure of the only N-terminal domain of the protein is available (PDB code = 5IQZ) [297], making, so far, VS campaigns quite complicated.
On the other hand, no ligand-based approaches have been attempted, probably because of the limited number of known selective SIRT-7 modulators.
Very recently (2022), Zhang et al. described a predicted model of the SIRT-7 structure, by using the fold recognition (or threading) method, followed by structure-based VS [184]. The structural model of the human SIRT-7 was managed by I-tasser [298] following a procedure described in the literature [299,300]. Subsequent VS screening was carried out by Autodock Vina [301] by taking into account the Chemdiv database [258] containing 939,319 structurally diverse compounds.
The study led to the selective SIRT-7 inhibitors 2800Z and 40569Z. These compounds exhibit high affinities toward SIRT-7 and low affinities to other SIRT proteins, and were shown to inhibit SIRT-7 deacetylation activity in vitro [184]. The related potential role in cancer has been investigated (see Section 2.7.1)
3.8. Pan-Sirtuin Modulators
With the aim to find isoform-specific compounds, Schlicker et al. [276] screened the National Cancer Institute (NCI) [252] diversity set against the crystal structures of SIRT-2 [6], -3 [285], -5 [302] and -6 [291]. The structures were modified in silico to accommodate an ADPR molecule, using as a reference structure the SIRT-6 complex (PDB code: 3K35) [291]. The top compounds from each screening were selected for experimental validation and isoform selectivity evaluation. Among the 20 active compounds, 14 compounds had selectivity toward SIRT-2, while no isoform-selective modulators were retrieved for the other sirtuins. On the opposite, multiple sirtuin VS was applied to individuate non-selective/pan-sirtuins inhibitors, which proved to be beneficial in some cancer conditions [303,304].
In 2017, Schnekenburger et al. identified Compound 18 as an SIRT-1/-2 modulator. The study involved a molecular docking step, useful to compare the predicted binding affinities of the candidates. The in-house library was screened against the X-ray crystal structure of SIRT-1 (PDB code = 4I5I) [230] and SIRT-2 (PDB code = 4RMG) [112], using AutoDock Vina [301]. The compound showed an anti-proliferative effect in glioma cells, both in in vitro and in vivo settings (see Section 2.8.1) [197].
In 2018, Karaman et al. [277] performed a consensus SBVS to identify new SIRT-1 and -2 inhibitors. In particular, the p-ANAPL database [305] was prefiltered to exclude PAINs [306] and submitted to docking against two SIRT-1 [228,230] and four SIRT-2 [112,307,308,309] crystal structures (PDB IDs SIRT-1: 4I5I, 4ZZJ; SIRT-2: 4R8M, 4L3O, 4RMH, 5D7P). Top-scored compounds by the two tranches were combined and the duplicates were removed. Seven compounds were tested in vitro on SIRT-1, SIRT-2 and SIRT-3, and two bichalcones were individuated as moderately active inhibitors. More recently, a repositioning study [279] was performed to individuate (pan-)inhibitors of SIRT-1 (4I5I) [230], -2 (5DY4) [260], -3 (4BV3) [310], -5 (5XHS) [290] and -6 (6HOY) [311]. In particular, a set of 1502 FDA-approved drugs was screened against the crystal structures of the targets using three different pieces of software (Glide [261,262], FRED [312], and AutoDock Vina/PyRx [313]). A consensus score was produced by calculating the average value of the three scores. Alosetron, cinacalcet, and indacaterol were individuated as the best pan-SIRT putative inhibitors. In 2019, Sinha et al. [278] performed an SBVS of plant-derived inhibitors against multiple sirtuin isoforms (SIRT-1 PDB:4I5I [230], SIRT-2 PDB:1J8F [6], SIRT-3 PDB:5D7N [309], SIRT-5 PDB:2B4Y [314], SIRT-6 PDB:3K35 [291] and SIRT-7 PDB:5IQZ [297]). Three hits were selected on the basis of their drug-like and ADME profile: in particular, Sulforaphane showed therapeutic potential against SIRT-1 and -5, Kaempferol on SIRT-3 and Apigenin against SIRT-6. In vitro studies confirmed the hypothesized activities.
3.9. Parasitic Sirtuins
In addition to human isoforms, parasitic sirtuins represent an important target to treat various protozoa-related diseases [315,316,317,318,319,320]. This is due to the importance of sirtuins in different physiological processes of the parasite [321], but an important warning is represented by selectivity issues toward the host’s isoforms. Here, we report VS studies targeting Trypanosoma cruzi, Schistosoma mansoni and Leishmania sirtuins.
In 2012, Soares et al. performed a docking-based VS of a series of nicotine analogues toward Trypanosoma cruzi sirtuin TcSir2rp3 [281]. An HM of the target was used as a receptor model for the VS. The top hit of the screening resulted to be nicotinamide itself. In vitro tests confirmed the antitrypanosomal activity of this compound. A few years later, Sacconnay et al. [282] performed an SBVS of 50 natural trypanocydal compounds against both of the two sirtuins of Trypanosoma cruzi (TcSIr2rp1 and TcSir2rp3). Two homology models corresponding to the two isoforms were employed as a docking template for the VS. The aim of the study was to evaluate if the anti-parasite action of the screened compounds could be exerted via Sirtuin inhibition. The score of four molecules (an anacardic acid derivative, aculeatin D, 16-acetoxy-11-hydroxyoctadeca-17-ene-12,14-diynylethanoate and vismione D) exceeded the one of AGK2 and thiobarbiturate 6 (two potent and selective inhibitors of sirtuins, respectively). For these compounds, a sirtuin-mediated mechanism was hypothesized: nevertheless, experimental validation is required. VS techniques were further applied to develop new inhibitors of Schistosoma mansoni Sirtuin 2, a protein involved in the reproductive functions of this parasite [283]. An HM was used to screen a zinc [226]-derived library. As species-selective compounds are desired, an analogous screening was performed on the human SIRT-2. Four leads were selected by comparing the two screening results, and by score comparison with respect to the AGK2-hSIRT-2 complex. In 2008, Kadam et al. performed a combined LB-SB VS targeting Leishmania sirtuin [280]. In particular, the fingerprint of nicotinamide was used as a query to screen the NCI database [322]. The retrieved molecules were docked in the hSir2 crystal structure [6] and in a previously reported [323] HM of LmSir2. The results were grouped according to their putative selectivity, and members of each group were submitted to experimental validation. Despite the fact that a truly potent and selective lead was not individuated, some of the tested compounds showed potential for further optimization.
4. Conclusions
Sirtuins are involved in a wide range of different processes, ranging from transcription to metabolism to genome stability. Their dysregulation is thought to be related to the pathogenesis and/or progression of different diseases, such as cancer, neurodegenerative disorders and T2D. In this context, molecules targeting selectively different SIRTs are considered as promising compounds for the search for new drugs. Up to now, the discovery of novel SIRT modulators was mainly addressed by applying different rational methods and VS strategies, in the attempt of identifying hit compounds for the further hit-to-lead optimization process. This approach allowed for the discovery of different chemo-types, worthy of further investigation and development.
Most of the VS herein presented regarding the search for SIRT-1 modulators relied on structure-based studies, a number of X-ray crystallographic data being available about SIRT-1 in the presence of activator/inhibitors. Indeed, six and three of the reported studies, out of 14 reported analyses (see Table 3), have been performed by SBVS and combined LB-SB VS strategies, respectively. In most of them, inhibitors as well as activators have been successfully identified. Conversely, SIRT-2 modulators have been discovered mainly thanks to combined LB-SB VS rather than SBVS (5 and 7 studies out of the 12 reported), highlighting on the other hand the predominant effective role played by the high number of structural information on SIRT-2-inhibitor complexes. During the last years, docking programs have been successfully embedded in automated workflows for ultra-large compound library screening, confirming the first steps of the VS search as viable. However, the selection of promising virtual hits (usually less than 100 compounds) from many high-scoring compounds in the library remains a challenge, since different selection protocols usually lead to different results.
Despite the consistent number of similar data regarding SIRT-6, few series of novel derivatives have been so far identified, the search for deepening studies including molecular dynamic simulations being an urgent need to properly manage this issue as well as the selectivity profile of any new putative SIRT-6 modulator. Lastly, performing in silico the prediction of the pharmacokinetic and toxicity (ADMET) properties of compounds should be performed in parallel, starting from the first steps of the drug design process and VS studies: compound library collection and the following hit-to-lead analysis would be efficiently oriented, as more drug-like compounds, prior to chemical synthesis.
Author Contributions
Conceptualization, S.B. and E.C.; writing—original draft preparation, E.A. and N.S.; writing—review and editing, S.B. and F.P.; editing, E.M.; funding acquisition, S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by European Commission: Horizon 2020 Marie Skłodowska-Curie grant No 671881 (INTEGRATA) and Ministry of Education, Universities and Research: PRIN 2017Z5LR5Z_003.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Finkel, T.; Deng, C.-X.; Mostoslavsky, R. Recent Progress in the Biology and Physiology of Sirtuins. Nature 2009, 460, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Makkar, R.; Behl, T.; Arora, S. Role of HDAC Inhibitors in Diabetes Mellitus. Curr. Res. Transl. Med. 2020, 68, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin Activators and Inhibitors: Promises, Achievements, and Challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, R.L.; Daniels, E.G.; Molenaars, M.; Houtkooper, R.H.; Janssens, G.E. From Molecular Promise to Preclinical Results: HDAC Inhibitors in the Race for Healthy Aging Drugs. EMBO Mol. Med. 2019, 11, e9854. [Google Scholar] [CrossRef] [PubMed]
- Hull, E.E.; Montgomery, M.R.; Leyva, K.J. HDAC Inhibitors as Epigenetic Regulators of the Immune System: Impacts on Cancer Therapy and Inflammatory Diseases. BioMed Res. Int. 2016, 2016, 1–15. [Google Scholar] [CrossRef]
- Finnin, M.S.; Donigian, J.R.; Pavletich, N.P. Structure of the Histone Deacetylase SIRT2. Nat. Struct. Biol. 2001, 8, 621–625. [Google Scholar] [CrossRef]
- Haigis, M.C.; Guarente, L.P. Mammalian Sirtuins—Emerging Roles in Physiology, Aging, and Calorie Restriction. Genes Dev. 2006, 20, 2913–2921. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. Sirtuins and Type 2 Diabetes: Role in Inflammation, Oxidative Stress, and Mitochondrial Function. Front. Endocrinol. 2019, 10, 187. [Google Scholar] [CrossRef]
- Wang, W.; Li, J.; Cai, L. Research Progress of Sirtuins in Renal and Cardiovascular Diseases. Curr. Opin. Nephrol. Hypertens. 2021, 30, 108–114. [Google Scholar] [CrossRef]
- Jurkowska, K.; Szymańska, B.; Knysz, B.; Kuźniarski, A.; Piwowar, A. Sirtuins as Interesting Players in the Course of HIV Infection and Comorbidities. Cells 2021, 10, 2739. [Google Scholar] [CrossRef]
- Leite, J.A.; Ghirotto, B.; Targhetta, V.P.; Lima, J.; Câmara, N.O.S. Sirtuins as Pharmacological Targets in Neurodegenerative and Neuropsychiatric Disorders. Br. J. Pharmacol. 2022, 179, 1496–1511. [Google Scholar] [CrossRef]
- Zhao, E.; Hou, J.; Ke, X.; Abbas, M.N.; Kausar, S.; Zhang, L.; Cui, H. The Roles of Sirtuin Family Proteins in Cancer Progression. Cancers 2019, 11, 1949. [Google Scholar] [CrossRef]
- Mei, Z.; Zhang, X.; Yi, J.; Huang, J.; He, J.; Tao, Y. Sirtuins in Metabolism, DNA Repair and Cancer. J. Exp. Clin. Cancer Res. 2016, 35, 182. [Google Scholar] [CrossRef] [PubMed]
- Bonkowski, M.S.; Sinclair, D.A. Slowing Ageing by Design: The Rise of NAD+ and Sirtuin-Activating Compounds. Nat. Rev. Mol. Cell Biol. 2016, 17, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Mautone, N.; Zwergel, C.; Mai, A.; Rotili, D. Sirtuin Modulators: Where Are We Now? A Review of Patents from 2015 to 2019. Expert Opin. Ther. Pat. 2020, 30, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, T.; Li, X.; Peng, T.; Hang, H.C.; Li, X.D. Integrative Chemical Biology Approaches for Identification and Characterization of “Erasers” for Fatty-Acid-Acylated Lysine Residues within Proteins. Angew. Chem. Int. Ed. Engl. 2015, 54, 1149–1152. [Google Scholar] [CrossRef] [PubMed]
- Pillai, V.B.; Kanwal, A.; Fang, Y.H.; Sharp, W.W.; Samant, S.; Arbiser, J.; Gupta, M.P. Honokiol, an Activator of Sirtuin-3 (SIRT3) Preserves Mitochondria and Protects the Heart from Doxorubicin-Induced Cardiomyopathy in Mice. Oncotarget 2017, 8, 34082–34098. [Google Scholar] [CrossRef]
- Hebert, A.S.; Dittenhafer-Reed, K.E.; Yu, W.; Bailey, D.J.; Selen, E.S.; Boersma, M.D.; Carson, J.J.; Tonelli, M.; Balloon, A.J.; Higbee, A.J.; et al. Calorie Restriction and SIRT3 Trigger Global Reprogramming of the Mitochondrial Protein Acetylome. Mol. Cell 2013, 49, 186–199. [Google Scholar] [CrossRef]
- Li, S.; Zheng, W. Mammalian Sirtuins SIRT4 and SIRT7. Prog. Mol. Biol. Transl. Sci. 2018, 154, 147–168. [Google Scholar] [CrossRef]
- Bheda, P.; Jing, H.; Wolberger, C.; Lin, H. The Substrate Specificity of Sirtuins. Annu. Rev. Biochem. 2016, 85, 405–429. [Google Scholar] [CrossRef]
- Tan, M.; Peng, C.; Anderson, K.A.; Chhoy, P.; Xie, Z.; Dai, L.; Park, J.; Chen, Y.; Huang, H.; Zhang, Y.; et al. Lysine Glutarylation Is a Protein Posttranslational Modification Regulated by SIRT5. Cell Metab. 2014, 19, 605–617. [Google Scholar] [CrossRef]
- Du, J.; Zhou, Y.; Su, X.; Yu, J.J.; Khan, S.; Jiang, H.; Kim, J.; Choi, B.H.; He, B.; Chen, W.; et al. Sirt5 Is a NAD-Dependent Protein Lysine Demalonylase and Desuccinylase. Science 2011, 334, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Rardin, M.J.; He, W.; Nishida, Y.; Newman, J.C.; Carrico, C.; Danielson, S.R.; Guo, A.; Gut, P.; Sahu, A.K.; Li, B.; et al. SIRT5 Regulates the Mitochondrial Lysine Succinylome and Metabolic Networks. Cell Metab. 2013, 18, 920–933. [Google Scholar] [CrossRef] [PubMed]
- van Meter, M.; Mao, Z.; Gorbunova, V.; Seluanov, A. SIRT6 Overexpression Induces Massive Apoptosis in Cancer Cells but Not in Normal Cells. Cell Cycle 2011, 10, 3153–3158. [Google Scholar] [CrossRef] [PubMed]
- Liszt, G.; Ford, E.; Kurtev, M.; Guarente, L. Mouse Sir2 Homolog SIRT6 Is a Nuclear ADP-Ribosyltransferase. J. Biol. Chem. 2005, 280, 21313–21320. [Google Scholar] [CrossRef] [PubMed]
- Feldman, J.L.; Dittenhafer-Reed, K.E.; Denu, J.M. Sirtuin Catalysis and Regulation. J. Biol. Chem. 2012, 287, 42419–42427. [Google Scholar] [CrossRef]
- Jiang, H.; Khan, S.; Wang, Y.; Charron, G.; He, B.; Sebastian, C.; Du, J.; Kim, R.; Ge, E.; Mostoslavsky, R.; et al. SIRT6 Regulates TNF-Alpha Secretion through Hydrolysis of Long-Chain Fatty Acyl Lysine. Nature 2013, 496, 110–113. [Google Scholar] [CrossRef]
- Bosch-Presegue, L.; Vaquero, A. The Dual Role of Sirtuins in Cancer. Genes Cancer 2011, 2, 648–662. [Google Scholar] [CrossRef]
- Ehrenberg, A.J.; Khatun, A.; Coomans, E.; Betts, M.J.; Capraro, F.; Thijssen, E.H.; Senkevich, K.; Bharucha, T.; Jafarpour, M.; Young, P.N.E.; et al. Relevance of Biomarkers across Different Neurodegenerative Diseases. Alzheimers Res. Ther. 2020, 12, 56. [Google Scholar] [CrossRef]
- Luo, G.; Jian, Z.; Zhu, Y.; Zhu, Y.; Chen, B.; Ma, R.; Tang, F.; Xiao, Y. Sirt1 Promotes Autophagy and Inhibits Apoptosis to Protect Cardiomyocytes from Hypoxic Stress. Int. J. Mol. Med. 2019, 43, 2033–2043. [Google Scholar] [CrossRef]
- Raynes, R.; Brunquell, J.; Westerheide, S.D. Stress Inducibility of SIRT1 and Its Role in Cytoprotection and Cancer. Genes Cancer 2013, 4, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Oberdoerffer, P.; Michan, S.; McVay, M.; Mostoslavsky, R.; Vann, J.; Park, S.-K.; Hartlerode, A.; Stegmuller, J.; Hafner, A.; Loerch, P.; et al. SIRT1 Redistribution on Chromatin Promotes Genomic Stability but Alters Gene Expression during Aging. Cell 2008, 135, 907–918. [Google Scholar] [CrossRef]
- Bai, X.; Yao, L.; Ma, X.; Xu, X. Small Molecules as SIRT Modulators. Mini-Rev. Med. Chem. 2018, 18, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Pacholec, M.; Bleasdale, J.E.; Chrunyk, B.; Cunningham, D.; Flynn, D.; Garofalo, R.S.; Griffith, D.; Griffor, M.; Loulakis, P.; Pabst, B.; et al. SRT1720, SRT2183, SRT1460, and Resveratrol Are Not Direct Activators of SIRT1. J. Biol. Chem. 2010, 285, 8340–8351. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.; Wang, M.; Zhong, A.; Wang, Y.; Du, J.; Wang, J.; Qi, L.; Bi, Z.; Zhang, P.; Lin, T.; et al. SRT1720 Inhibits the Growth of Bladder Cancer in Organoids and Murine Models through the SIRT1-HIF Axis. Oncogene 2021, 40, 6081–6092. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, R. SRT1720, a SIRT1 Activator, Promotes Tumor Cell Migration, and Lung Metastasis of Breast Cancer in Mice. Oncol. Rep. 2012, 27, 1726–1732. [Google Scholar] [CrossRef][Green Version]
- Zhang, X.; Chen, J.; Sun, L.; Xu, Y. SIRT1 Deacetylates KLF4 to Activate Claudin-5 Transcription in Ovarian Cancer Cells. J. Cell Biochem. 2018, 119, 2418–2426. [Google Scholar] [CrossRef]
- Guida, N.; Laudati, G.; Anzilotti, S.; Secondo, A.; Montuori, P.; di Renzo, G.; Canzoniero, L.M.T.; Formisano, L. Resveratrol via Sirtuin-1 Downregulates RE1-Silencing Transcription Factor (REST) Expression Preventing PCB-95-Induced Neuronal Cell Death. Toxicol. Appl. Pharmacol. 2015, 288, 387–398. [Google Scholar] [CrossRef]
- Liu, T.; Ma, Y.; Zhang, R.; Zhong, H.; Wang, L.; Zhao, J.; Yang, L.; Fan, X. Resveratrol Ameliorates Estrogen Deficiency-Induced Depression- and Anxiety-like Behaviors and Hippocampal Inflammation in Mice. Psychopharmacology 2019, 236, 1385–1399. [Google Scholar] [CrossRef]
- Scuderi, C.; Stecca, C.; Bronzuoli, M.R.; Rotili, D.; Valente, S.; Mai, A.; Steardo, L. Sirtuin Modulators Control Reactive Gliosis in an in Vitro Model of Alzheimer’s Disease. Front. Pharmacol. 2014, 5, 89. [Google Scholar] [CrossRef]
- Kumar, R.; Nigam, L.; Singh, A.P.; Singh, K.; Subbarao, N.; Dey, S. Design, Synthesis of Allosteric Peptide Activator for Human SIRT1 and Its Biological Evaluation in Cellular Model of Alzheimer’s Disease. Eur. J. Med. Chem. 2017, 127, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Cea, M.; Soncini, D.; Fruscione, F.; Raffaghello, L.; Garuti, A.; Emionite, L.; Moran, E.; Magnone, M.; Zoppoli, G.; Reverberi, D.; et al. Synergistic Interactions between HDAC and Sirtuin Inhibitors in Human Leukemia Cells. PLoS ONE 2011, 6, e22739. [Google Scholar] [CrossRef] [PubMed]
- Kalle, A.M.; Mallika, A.; Badiger, J.; Alinakhi; Talukdar, P. Sachchidanand Inhibition of SIRT1 by a Small Molecule Induces Apoptosis in Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2010, 401, 13–19. [Google Scholar] [CrossRef]
- Wang, T.; Li, X.; Sun, S. EX527, a Sirt-1 Inhibitor, Induces Apoptosis in Glioma via Activating the P53 Signaling Pathway. Anticancer Drugs 2020, 31, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cao, L.; Li, Z.; Li, Y. SIRT1 Promotes GLUT1 Expression and Bladder Cancer Progression via Regulation of Glucose Uptake. Hum. Cell 2019, 32, 193–201. [Google Scholar] [CrossRef]
- Asaka, R.; Miyamoto, T.; Yamada, Y.; Ando, H.; Mvunta, D.H.; Kobara, H.; Shiozawa, T. Sirtuin 1 Promotes the Growth and Cisplatin Resistance of Endometrial Carcinoma Cells: A Novel Therapeutic Target. Lab. Investig. 2015, 95, 1363–1373. [Google Scholar] [CrossRef]
- Oon, C.E.; Strell, C.; Yeong, K.Y.; Östman, A.; Prakash, J. SIRT1 Inhibition in Pancreatic Cancer Models: Contrasting Effects in Vitro and in Vivo. Eur. J. Pharmacol. 2015, 757, 59–67. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, B.; Xu, H.; Sun, Y.; Shi, Y.; Luo, Y.; Jia, H.; Wang, F. Suppression of Sirt1 Sensitizes Lung Cancer Cells to WEE1 Inhibitor MK-1775-Induced DNA Damage and Apoptosis. Oncogene 2017, 36, 6863–6872. [Google Scholar] [CrossRef]
- Yousafzai, N.A.; Zhou, Q.; Xu, W.; Shi, Q.; Xu, J.; Feng, L.; Chen, H.; Shin, V.Y.; Jin, H.; Wang, X. SIRT1 Deacetylated and Stabilized XRCC1 to Promote Chemoresistance in Lung Cancer. Cell Death Dis. 2019, 10, 363. [Google Scholar] [CrossRef]
- Peck, B.; Chen, C.-Y.; Ho, K.-K.; di Fruscia, P.; Myatt, S.S.; Coombes, R.C.; Fuchter, M.J.; Hsiao, C.-D.; Lam, E.W.-F. SIRT Inhibitors Induce Cell Death and P53 Acetylation through Targeting Both SIRT1 and SIRT2. Mol. Cancer Ther. 2010, 9, 844–855. [Google Scholar] [CrossRef]
- Kim, H.-B.; Lee, S.-H.; Um, J.-H.; Oh, W.K.; Kim, D.-W.; Kang, C.-D.; Kim, S.-H. Sensitization of Multidrug-Resistant Human Cancer Cells to Hsp90 Inhibitors by down-Regulation of SIRT1. Oncotarget 2015, 6, 36202–36218. [Google Scholar] [CrossRef] [PubMed]
- Alvala, M.; Bhatnagar, S.; Ravi, A.; Jeankumar, V.U.; Manjashetty, T.H.; Yogeeswari, P.; Sriram, D. Novel Acridinedione Derivatives: Design, Synthesis, SIRT1 Enzyme and Tumor Cell Growth Inhibition Studies. Bioorg. Med. Chem. Lett. 2012, 22, 3256–3260. [Google Scholar] [CrossRef]
- Manjulatha, K.; Srinivas, S.; Mulakayala, N.; Rambabu, D.; Prabhakar, M.; Arunasree, K.M.; Alvala, M.; Basaveswara Rao, M.V.; Pal, M. Ethylenediamine Diacetate (EDDA) Mediated Synthesis of Aurones under Ultrasound: Their Evaluation as Inhibitors of SIRT1. Bioorg. Med. Chem. Lett. 2012, 22, 6160–6165. [Google Scholar] [CrossRef]
- Mellini, P.; Kokkola, T.; Suuronen, T.; Salo, H.S.; Tolvanen, L.; Mai, A.; Lahtela-Kakkonen, M.; Jarho, E.M. Screen of Pseudopeptidic Inhibitors of Human Sirtuins 1–3: Two Lead Compounds with Antiproliferative Effects in Cancer Cells. J. Med. Chem. 2013, 56, 6681–6695. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.K.; Cho, K.B.; Hien, T.T.; Kim, T.H.; Kim, H.S.; Dao, T.T.; Han, H.-K.; Kwon, S.-M.; Ahn, S.-G.; Yoon, J.-H.; et al. Amurensin G, a Potent Natural SIRT1 Inhibitor, Rescues Doxorubicin Responsiveness via Down-Regulation of Multidrug Resistance 1. Mol. Pharmacol. 2010, 78, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Muscolini, M.; Castiello, L.; Palermo, E.; Zevini, A.; Ferrari, M.; Olagnier, D.; Hiscott, J. SIRT1 Modulates the Sensitivity of Prostate Cancer Cells to Vesicular Stomatitis Virus Oncolysis. J. Virol. 2019, 93, e00626-19. [Google Scholar] [CrossRef] [PubMed]
- Pulla, V.K.; Sriram, D.S.; Viswanadha, S.; Sriram, D.; Yogeeswari, P. Energy-Based Pharmacophore and Three-Dimensional Quantitative Structure-Activity Relationship (3D-QSAR) Modeling Combined with Virtual Screening to Identify Novel Small-Molecule Inhibitors of Silent Mating-Type Information Regulation 2 Homologue 1 (SIRT1). J. Chem. Inf. Model. 2016, 56, 173–187. [Google Scholar] [CrossRef]
- Zhang, Q.; Zeng, S.X.; Zhang, Y.; Zhang, Y.; Ding, D.; Ye, Q.; Meroueh, S.O.; Lu, H. A Small Molecule Inauhzin Inhibits SIRT1 Activity and Suppresses Tumour Growth through Activation of P53. EMBO Mol. Med. 2012, 4, 298–312. [Google Scholar] [CrossRef]
- Wössner, N.; Alhalabi, Z.; González, J.; Swyter, S.; Gan, J.; Schmidtkunz, K.; Zhang, L.; Vaquero, A.; Ovaa, H.; Einsle, O.; et al. Sirtuin 1 Inhibiting Thiocyanates (S1th)—A New Class of Isotype Selective Inhibitors of NAD+ Dependent Lysine Deacetylases. Front. Oncol. 2020, 10, 657. [Google Scholar] [CrossRef]
- Wan, J.; Deng, L.; Zhang, C.; Yuan, Q.; Liu, J.; Dun, Y.; Zhou, Z.; Zhao, H.; Liu, C.; Yuan, D.; et al. Chikusetsu Saponin V Attenuates H2O2-Induced Oxidative Stress in Human Neuroblastoma SH-SY5Y Cells through Sirt1/PGC-1α/Mn-SOD Signaling Pathways. Can. J. Physiol. Pharmacol. 2016, 94, 919–928. [Google Scholar] [CrossRef]
- Kim, H.-D.; Hesterman, J.; Call, T.; Magazu, S.; Keeley, E.; Armenta, K.; Kronman, H.; Neve, R.L.; Nestler, E.J.; Ferguson, D. SIRT1 Mediates Depression-Like Behaviors in the Nucleus Accumbens. J. Neurosci. 2016, 36, 8441–8452. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhang, F.; Guan, X. Baicalin Reverse Depressive-like Behaviors through Regulation SIRT1-NF-kB Signaling Pathway in Olfactory Bulbectomized Rats. Phytother. Res. 2019, 33, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Huang, F.; Jian, C.; Qin, L.; Lu, F.; Wang, Y.; Zhang, Z.; Zhang, Q. Neuroprotective Effect of Salidroside against Central Nervous System Inflammation-Induced Cognitive Deficits: A Pivotal Role of Sirtuin 1-Dependent Nrf-2/HO-1/NF-ΚB Pathway. Phytother. Res. 2019, 33, 1438–1447. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J. Training for the Future. Nurs. Stand. 1988, 2, 32. [Google Scholar]
- Min, S.-W.; Cho, S.-H.; Zhou, Y.; Schroeder, S.; Haroutunian, V.; Seeley, W.W.; Huang, E.J.; Shen, Y.; Masliah, E.; Mukherjee, C.; et al. Acetylation of Tau Inhibits Its Degradation and Contributes to Tauopathy. Neuron 2010, 67, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Mi, M. Resveratrol Attenuates Aβ25–35 Caused Neurotoxicity by Inducing Autophagy Through the TyrRS-PARP1-SIRT1 Signaling Pathway. Neurochem. Res. 2016, 41, 2367–2379. [Google Scholar] [CrossRef]
- Shekhar, S.; Yadav, Y.; Singh, A.P.; Pradhan, R.; Desai, G.R.; Dey, A.B.; Dey, S. Neuroprotection by Ethanolic Extract of Syzygium Aromaticum in Alzheimer’s Disease like Pathology via Maintaining Oxidative Balance through SIRT1 Pathway. Exp. Gerontol. 2018, 110, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Yin, J.-B.; Liu, L.-H.; Guo, J.; Wang, S.-H.; Qu, C.-H.; Wang, C.-X. Protective Role of Dihydromyricetin in Alzheimer’s Disease Rat Model Associated with Activating AMPK/SIRT1 Signaling Pathway. Biosci. Rep. 2019, 39, BSR20180902. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Cuccioloni, M.; Angeletti, M.; Berardi, S.; Scarpona, S.; Rossi, G.; Eleuteri, A.M. SLAB51 Probiotic Formulation Activates SIRT1 Pathway Promoting Antioxidant and Neuroprotective Effects in an AD Mouse Model. Mol. Neurobiol. 2018, 55, 7987–8000. [Google Scholar] [CrossRef]
- Lee, H.R.; Shin, H.K.; Park, S.Y.; Kim, H.Y.; Lee, W.S.; Rhim, B.Y.; Hong, K.W.; Kim, C.D. Cilostazol Suppresses β-Amyloid Production by Activating a Disintegrin and Metalloproteinase 10 via the Upregulation of SIRT1-Coupled Retinoic Acid Receptor-β. J. Neurosci. Res. 2014, 92, 1581–1590. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Lee, H.; Yoo, H.B.; Choi, J.-S.; Jung, H.-Y.; Yoon, E.J.; Kim, H.; Jung, Y.-H.; Lee, H.-Y.; Kim, Y.K. Efficacy of Cilostazol Administration in Alzheimer’s Disease Patients with White Matter Lesions: A Positron-Emission Tomography Study. Neurotherapeutics 2019, 16, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Syed, A.; Lukacsovich, T.; Purcell, J.; Barbaro, B.A.; Worthge, S.A.; Wei, S.R.; Pollio, G.; Magnoni, L.; Scali, C.; et al. A Potent and Selective Sirtuin 1 Inhibitor Alleviates Pathology in Multiple Animal and Cell Models of Huntington’s Disease. Hum. Mol. Genet. 2014, 23, 2995–3007. [Google Scholar] [CrossRef] [PubMed]
- Westerberg, G.; Chiesa, J.A.; Andersen, C.A.; Diamanti, D.; Magnoni, L.; Pollio, G.; Darpo, B.; Zhou, M. Safety, Pharmacokinetics, Pharmacogenomics and QT Concentration−effect Modelling of the SirT1 Inhibitor Selisistat in Healthy Volunteers. Br. J. Clin. Pharmacol. 2015, 79, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Süssmuth, S.D.; Haider, S.; Landwehrmeyer, G.B.; Farmer, R.; Frost, C.; Tripepi, G.; Andersen, C.A.; di Bacco, M.; Lamanna, C.; Diodato, E.; et al. An Exploratory Double-Blind, Randomized Clinical Trial with Selisistat, a SirT1 Inhibitor, in Patients with Huntington’s Disease. Br. J. Clin. Pharmacol. 2015, 79, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.H.; Lim, Y.; Oh, H.J.; Park, S.M.; Lee, S.-K.; Ahnn, J.; Kim, D.H.; Song, W.K.; Kwak, T.H.; Park, W.J. Pharmacological Activation of Sirt1 Ameliorates Polyglutamine-Induced Toxicity through the Regulation of Autophagy. PLoS ONE 2013, 8, e64953. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Ban, J.-J.; Chung, J.-Y.; Im, W.; Kim, M. Amelioration of Huntington’s Disease Phenotypes by Beta-Lapachone Is Associated with Increases in Sirt1 Expression, CREB Phosphorylation and PGC-1α Deacetylation. PLoS ONE 2018, 13, e0195968. [Google Scholar] [CrossRef]
- Chen, C.; Xia, B.; Tang, L.; Wu, W.; Tang, J.; Liang, Y.; Yang, H.; Zhang, Z.; Lu, Y.; Chen, G.; et al. Echinacoside Protects against MPTP/MPP+-Induced Neurotoxicity via Regulating Autophagy Pathway Mediated by Sirt1. Metab. Brain Dis. 2019, 34, 203–212. [Google Scholar] [CrossRef]
- Rao, S.P.; Sharma, N.; Kalivendi, S.V. Embelin Averts MPTP-Induced Dysfunction in Mitochondrial Bioenergetics and Biogenesis via Activation of SIRT1. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2020, 1861, 148157. [Google Scholar] [CrossRef]
- Valle, C.; Salvatori, I.; Gerbino, V.; Rossi, S.; Palamiuc, L.; René, F.; Carrì, M.T. Tissue-Specific Deregulation of Selected HDACs Characterizes ALS Progression in Mouse Models: Pharmacological Characterization of SIRT1 and SIRT2 Pathways. Cell Death Dis. 2014, 5, e1296. [Google Scholar] [CrossRef]
- Prozorovski, T.; Ingwersen, J.; Lukas, D.; Göttle, P.; Koop, B.; Graf, J.; Schneider, R.; Franke, K.; Schumacher, S.; Britsch, S.; et al. Regulation of Sirtuin Expression in Autoimmune Neuroinflammation: Induction of SIRT1 in Oligodendrocyte Progenitor Cells. Neurosci. Lett. 2019, 704, 116–125. [Google Scholar] [CrossRef]
- Lim, H.W.; Kang, S.G.; Ryu, J.K.; Schilling, B.; Fei, M.; Lee, I.S.; Kehasse, A.; Shirakawa, K.; Yokoyama, M.; Schnölzer, M.; et al. SIRT1 Deacetylates RORγt and Enhances Th17 Cell Generation. J. Exp. Med. 2015, 212, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Nayagam, V.M.; Wang, X.; Tan, Y.C.; Poulsen, A.; Goh, K.C.; Ng, T.; Wang, H.; Song, H.Y.; Ni, B.; Entzeroth, M.; et al. SIRT1 Modulating Compounds from High-Throughput Screening as Anti-Inflammatory and Insulin-Sensitizing Agents. J. Biomol. Screen. 2006, 11, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Zhou, P.; Xu, X.; Fang, W.; Jia, S.; Liu, W.; Su, X.; Zhang, J.; Wang, H.; Yu, P.; et al. Benzothiazole Derivatives Upregulate SIRT1 and Relevant Genes in High-Fat Fed C57BL/6J Mice. Med. Chem. Res. 2015, 24, 2454–2460. [Google Scholar] [CrossRef]
- Liu, P.; Feng, T.; Zuo, X.; Wang, X.; Luo, J.; Li, N.; Han, X.; Zhu, N.; Xu, S.; Xu, Y.; et al. A Novel SIRT1 Activator E6155 Improves Insulin Sensitivity in Type 2 Diabetic KKAy Mice. Biochem. Biophys. Res. Commun. 2018, 498, 633–639. [Google Scholar] [CrossRef]
- North, B.J.; Verdin, E. Interphase Nucleo-Cytoplasmic Shuttling and Localization of SIRT2 during Mitosis. PLoS ONE 2007, 2, e784. [Google Scholar] [CrossRef]
- Jing, E.; Gesta, S.; Kahn, C.R. SIRT2 Regulates Adipocyte Differentiation through FoxO1 Acetylation/Deacetylation. Cell Metab. 2007, 6, 105–114. [Google Scholar] [CrossRef]
- Dryden, S.C.; Nahhas, F.A.; Nowak, J.E.; Goustin, A.-S.; Tainsky, M.A. Role for Human SIRT2 NAD-Dependent Deacetylase Activity in Control of Mitotic Exit in the Cell Cycle. Mol. Cell. Biol. 2003, 23, 3173–3185. [Google Scholar] [CrossRef]
- Funato, K.; Hayashi, T.; Echizen, K.; Negishi, L.; Shimizu, N.; Koyama-Nasu, R.; Nasu-Nishimura, Y.; Morishita, Y.; Tabar, V.; Todo, T.; et al. SIRT2-mediated Inactivation of P73 Is Required for Glioblastoma Tumorigenicity. EMBO Rep. 2018, 19, e45587. [Google Scholar] [CrossRef]
- Hong, J.Y.; Fernandez, I.; Anmangandla, A.; Lu, X.; Bai, J.J.; Lin, H. Pharmacological Advantage of SIRT2-Selective versus Pan-SIRT1–3 Inhibitors. ACS Chem. Biol. 2021, 16, 1266–1275. [Google Scholar] [CrossRef]
- Rotili, D.; Tarantino, D.; Nebbioso, A.; Paolini, C.; Huidobro, C.; Lara, E.; Mellini, P.; Lenoci, A.; Pezzi, R.; Botta, G.; et al. Discovery of Salermide-Related Sirtuin Inhibitors: Binding Mode Studies and Antiproliferative Effects in Cancer Cells Including Cancer Stem Cells. J. Med. Chem. 2012, 55, 10937–10947. [Google Scholar] [CrossRef]
- Liu, P.Y.; Xu, N.; Malyukova, A.; Scarlett, C.J.; Sun, Y.T.; Zhang, X.D.; Ling, D.; Su, S.-P.; Nelson, C.; Chang, D.K.; et al. The Histone Deacetylase SIRT2 Stabilizes Myc Oncoproteins. Cell Death Differ. 2013, 20, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Kudo, N.; Ito, A.; Arata, M.; Nakata, A.; Yoshida, M. Identification of a Novel Small Molecule That Inhibits Deacetylase but Not Defatty-Acylase Reaction Catalysed by SIRT2. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170070. [Google Scholar] [CrossRef]
- Farooqi, A.S.; Hong, J.Y.; Cao, J.; Lu, X.; Price, I.R.; Zhao, Q.; Kosciuk, T.; Yang, M.; Bai, J.J.; Lin, H. Novel Lysine-Based Thioureas as Mechanism-Based Inhibitors of Sirtuin 2 (SIRT2) with Anticancer Activity in a Colorectal Cancer Murine Model. J. Med. Chem. 2019, 62, 4131–4141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Au, Q.; Zhang, M.; Barber, J.R.; Ng, S.C.; Zhang, B. Identification of a Small Molecule SIRT2 Inhibitor with Selective Tumor Cytotoxicity. Biochem. Biophys. Res. Commun. 2009, 386, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, N.A.; Price, I.R.; Jing, H.; Wang, M.; Yang, M.; Cao, J.; Hong, J.Y.; Zhang, X.; Aramsangtienchai, P.; Sadhukhan, S.; et al. Direct Comparison of SIRT2 Inhibitors: Potency, Specificity, Activity-Dependent Inhibition, and On-Target Anticancer Activities. ChemMedChem 2018, 13, 1890–1894. [Google Scholar] [CrossRef]
- Jing, H.; Hu, J.; He, B.; Negrón Abril, Y.L.; Stupinski, J.; Weiser, K.; Carbonaro, M.; Chiang, Y.-L.; Southard, T.; Giannakakou, P.; et al. A SIRT2-Selective Inhibitor Promotes c-Myc Oncoprotein Degradation and Exhibits Broad Anticancer Activity. Cancer Cell 2016, 29, 767–768. [Google Scholar] [CrossRef]
- Eren, G.; Bruno, A.; Guntekin-Ergun, S.; Cetin-Atalay, R.; Ozgencil, F.; Ozkan, Y.; Gozelle, M.; Kaya, S.G.; Costantino, G. Pharmacophore Modeling and Virtual Screening Studies to Identify Novel Selective SIRT2 Inhibitors. J. Mol. Graph. Model. 2019, 89, 60–73. [Google Scholar] [CrossRef]
- Seifert, T.; Malo, M.; Kokkola, T.; Engen, K.; Fridén-Saxin, M.; Wallén, E.A.A.; Lahtela-Kakkonen, M.; Jarho, E.M.; Luthman, K. Chroman-4-One- and Chromone-Based Sirtuin 2 Inhibitors with Antiproliferative Properties in Cancer Cells. J. Med. Chem. 2014, 57, 9870–9888. [Google Scholar] [CrossRef]
- Yang, L.-L.; Wang, H.-L.; Zhong, L.; Yuan, C.; Liu, S.-Y.; Yu, Z.-J.; Liu, S.; Yan, Y.-H.; Wu, C.; Wang, Y.; et al. X-Ray Crystal Structure Guided Discovery of New Selective, Substrate-Mimicking Sirtuin 2 Inhibitors That Exhibit Activities against Non-Small Cell Lung Cancer Cells. Eur. J. Med. Chem. 2018, 155, 806–823. [Google Scholar] [CrossRef]
- Moniot, S.; Forgione, M.; Lucidi, A.; Hailu, G.S.; Nebbioso, A.; Carafa, V.; Baratta, F.; Altucci, L.; Giacché, N.; Passeri, D.; et al. Development of 1,2,4-Oxadiazoles as Potent and Selective Inhibitors of the Human Deacetylase Sirtuin 2: Structure–Activity Relationship, X-Ray Crystal Structure, and Anticancer Activity. J. Med. Chem. 2017, 60, 2344–2360. [Google Scholar] [CrossRef]
- Kozako, T.; Mellini, P.; Ohsugi, T.; Aikawa, A.; Uchida, Y.-I.; Honda, S.-I.; Suzuki, T. Novel Small Molecule SIRT2 Inhibitors Induce Cell Death in Leukemic Cell Lines. BMC Cancer 2018, 18, 791. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.S.; Scian, M.; Sripathy, S.; Posakony, J.; Lao, U.; Loe, T.K.; Leko, V.; Thalhofer, A.; Schuler, A.D.; Bedalov, A.; et al. Development of Pyrazolone and Isoxazol-5-One Cambinol Analogues as Sirtuin Inhibitors. J. Med. Chem. 2014, 57, 3283–3294. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Sripathy, S.; Webster, A.; Park, A.; Lao, U.; Hsu, J.H.; Loe, T.; Bedalov, A.; Simon, J.A. Discovery of Selective SIRT2 Inhibitors as Therapeutic Agents in B-Cell Lymphoma and Other Malignancies. Molecules 2020, 25, 455. [Google Scholar] [CrossRef]
- Shah, A.A.; Ito, A.; Nakata, A.; Yoshida, M. Identification of a Selective SIRT2 Inhibitor and Its Anti-Breast Cancer Activity. Biol. Pharm. Bull. 2016, 39, 1739–1742. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, R.C.; Uchiechowska, U.; Meier, R.; Hruby, H.; Valkov, V.; Verdin, E.; Sippl, W.; Jung, M. Structure–Activity Studies on Splitomicin Derivatives as Sirtuin Inhibitors and Computational Prediction of Binding Mode. J. Med. Chem. 2008, 51, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Yeong, K.Y.; Khaw, K.Y.; Takahashi, Y.; Itoh, Y.; Murugaiyah, V.; Suzuki, T. Discovery of Gamma-Mangostin from Garcinia Mangostana as a Potent and Selective Natural SIRT2 Inhibitor. Bioorg. Chem. 2020, 94, 103403. [Google Scholar] [CrossRef]
- McCarthy, A.R.; Sachweh, M.C.C.; Higgins, M.; Campbell, J.; Drummond, C.J.; van Leeuwen, I.M.M.; Pirrie, L.; Ladds, M.J.G.W.; Westwood, N.J.; Laín, S. Tenovin-D3, a Novel Small-Molecule Inhibitor of Sirtuin SirT2, Increases P21 (CDKN1A) Expression in a P53-Independent Manner. Mol. Cancer Ther. 2013, 12, 352–360. [Google Scholar] [CrossRef]
- Yang, H.; Chen, Y.; Jiang, Y.; Wang, D.; Yan, J.; Zhou, Z. TP53 Mutation Influences the Efficacy of Treatment of Colorectal Cancer Cell Lines with a Combination of Sirtuin Inhibitors and Chemotherapeutic Agents. Exp. Ther. Med. 2020, 20, 1415–1422. [Google Scholar] [CrossRef]
- Outeiro, T.F.; Kontopoulos, E.; Altmann, S.M.; Kufareva, I.; Strathearn, K.E.; Amore, A.M.; Volk, C.B.; Maxwell, M.M.; Rochet, J.-C.; McLean, P.J.; et al. Sirtuin 2 Inhibitors Rescue Alpha-Synuclein-Mediated Toxicity in Models of Parkinson’s Disease. Science 2007, 317, 516–519. [Google Scholar] [CrossRef]
- Tatum, P.R.; Sawada, H.; Ota, Y.; Itoh, Y.; Zhan, P.; Ieda, N.; Nakagawa, H.; Miyata, N.; Suzuki, T. Identification of Novel SIRT2-Selective Inhibitors Using a Click Chemistry Approach. Bioorg. Med. Chem. Lett. 2014, 24, 1871–1874. [Google Scholar] [CrossRef]
- Taylor, D.M.; Balabadra, U.; Xiang, Z.; Woodman, B.; Meade, S.; Amore, A.; Maxwell, M.M.; Reeves, S.; Bates, G.P.; Luthi-Carter, R.; et al. A Brain-Permeable Small Molecule Reduces Neuronal Cholesterol by Inhibiting Activity of Sirtuin 2 Deacetylase. ACS Chem. Biol. 2011, 6, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, T.; Schiedel, M.; Karaman, B.; Roessler, C.; North, B.J.; Lehotzky, A.; Oláh, J.; Ladwein, K.I.; Schmidtkunz, K.; Gajer, M.; et al. Selective Sirt2 Inhibition by Ligand-Induced Rearrangement of the Active Site. Nat. Commun. 2015, 6, 6263. [Google Scholar] [CrossRef] [PubMed]
- Lain, S.; Hollick, J.J.; Campbell, J.; Staples, O.D.; Higgins, M.; Aoubala, M.; McCarthy, A.; Appleyard, V.; Murray, K.E.; Baker, L.; et al. Discovery, in Vivo Activity, and Mechanism of Action of a Small-Molecule P53 Activator. Cancer Cell 2008, 13, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Harting, K.; Knöll, B. SIRT2-Mediated Protein Deacetylation: An Emerging Key Regulator in Brain Physiology and Pathology. Eur. J. Cell Biol. 2010, 89, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, Y.; Cao, W.; Wei, X.; Chen, J.; Ying, W. SIRT2 Plays Significant Roles in Lipopolysaccharides-Induced Neuroinflammation and Brain Injury in Mice. Neurochem. Res. 2016, 41, 2490–2500. [Google Scholar] [CrossRef]
- Spires-Jones, T.L.; Fox, L.M.; Rozkalne, A.; Pitstick, R.; Carlson, G.A.; Kazantsev, A.G. Inhibition of Sirtuin 2 with Sulfobenzoic Acid Derivative AK1 Is Non-Toxic and Potentially Neuroprotective in a Mouse Model of Frontotemporal Dementia. Front. Pharmacol. 2012, 3, 42. [Google Scholar] [CrossRef]
- Silva, D.F.; Esteves, A.R.; Oliveira, C.R.; Cardoso, S.M. Mitochondrial Metabolism Power SIRT2-Dependent Deficient Traffic Causing Alzheimer’s-Disease Related Pathology. Mol. Neurobiol. 2017, 54, 4021–4040. [Google Scholar] [CrossRef]
- Diaz-Perdigon, T.; Belloch, F.B.; Ricobaraza, A.; Elboray, E.E.; Suzuki, T.; Tordera, R.M.; Puerta, E. Early Sirtuin 2 Inhibition Prevents Age-Related Cognitive Decline in a Senescence-Accelerated Mouse Model. Neuropsychopharmacology 2020, 45, 347–357. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, J.Q.; Hong, T.T.; Sun, Y.H.; Huang, H.L.; Chen, F.; Chen, X.J.; Chen, H.Y.; Dong, S.S.; Cui, L.L.; et al. RTN4B-Mediated Suppression of Sirtuin 2 Activity Ameliorates β-Amyloid Pathology and Cognitive Impairment in Alzheimer’s Disease Mouse Model. Aging Cell 2020, 19, e13194. [Google Scholar] [CrossRef]
- Esteves, A.R.; Palma, A.M.; Gomes, R.; Santos, D.; Silva, D.F.; Cardoso, S.M. Acetylation as a Major Determinant to Microtubule-Dependent Autophagy: Relevance to Alzheimer’s and Parkinson Disease Pathology. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2008–2023. [Google Scholar] [CrossRef]
- Biella, G.; Fusco, F.; Nardo, E.; Bernocchi, O.; Colombo, A.; Lichtenthaler, S.F.; Forloni, G.; Albani, D. Sirtuin 2 Inhibition Improves Cognitive Performance and Acts on Amyloid-β Protein Precursor Processing in Two Alzheimer’s Disease Mouse Models. J. Alzheimers Dis. 2016, 53, 1193–1207. [Google Scholar] [CrossRef] [PubMed]
- Quinti, L.; Casale, M.; Moniot, S.; Pais, T.F.; van Kanegan, M.J.; Kaltenbach, L.S.; Pallos, J.; Lim, R.G.; Naidu, S.D.; Runne, H.; et al. SIRT2- and NRF2-Targeting Thiazole-Containing Compound with Therapeutic Activity in Huntington’s Disease Models. Cell Chem. Biol. 2016, 23, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Luthi-Carter, R.; Taylor, D.M.; Pallos, J.; Lambert, E.; Amore, A.; Parker, A.; Moffitt, H.; Smith, D.L.; Runne, H.; Gokce, O.; et al. SIRT2 Inhibition Achieves Neuroprotection by Decreasing Sterol Biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 7927–7932. [Google Scholar] [CrossRef] [PubMed]
- Chopra, V.; Quinti, L.; Kim, J.; Vollor, L.; Narayanan, K.L.; Edgerly, C.; Cipicchio, P.M.; Lauver, M.A.; Choi, S.H.; Silverman, R.B.; et al. The Sirtuin 2 Inhibitor AK-7 Is Neuroprotective in Huntington’s Disease Mouse Models. Cell Rep. 2012, 2, 1492–1497. [Google Scholar] [CrossRef]
- Ai, T.; Wilson, D.J.; More, S.S.; Xie, J.; Chen, L. 5-((3-Amidobenzyl)Oxy)Nicotinamides as Sirtuin 2 Inhibitors. J. Med. Chem. 2016, 59, 2928–2941. [Google Scholar] [CrossRef]
- Garske, A.L.; Smith, B.C.; Denu, J.M. Linking SIRT2 to Parkinson’s Disease. ACS Chem. Biol. 2007, 2, 529–532. [Google Scholar] [CrossRef]
- Harrison, I.F.; Smith, A.D.; Dexter, D.T. Pathological Histone Acetylation in Parkinson’s Disease: Neuroprotection and Inhibition of Microglial Activation through SIRT 2 Inhibition. Neurosci. Lett. 2018, 666, 48–57. [Google Scholar] [CrossRef]
- Wang, X.; Guan, Q.; Wang, M.; Yang, L.; Bai, J.; Yan, Z.; Zhang, Y.; Liu, Z. Aging-Related Rotenone-Induced Neurochemical and Behavioral Deficits: Role of SIRT2 and Redox Imbalance, and Neuroprotection by AK-7. Drug Des. Dev. Ther. 2015, 9, 2553–2563. [Google Scholar] [CrossRef]
- Guan, Q.; Wang, M.; Chen, H.; Yang, L.; Yan, Z.; Wang, X. Aging-Related 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Neurochemial and Behavioral Deficits and Redox Dysfunction: Improvement by AK-7. Exp. Gerontol. 2016, 82, 19–29. [Google Scholar] [CrossRef]
- di Fruscia, P.; Zacharioudakis, E.; Liu, C.; Moniot, S.; Laohasinnarong, S.; Khongkow, M.; Harrison, I.F.; Koltsida, K.; Reynolds, C.R.; Schmidtkunz, K.; et al. The Discovery of a Highly Selective 5,6,7,8-Tetrahydrobenzo [4,5]Thieno [2,3-d]Pyrimidin-4(3H)-One SIRT2 Inhibitor That Is Neuroprotective in an in Vitro Parkinson’s Disease Model. ChemMedChem 2015, 10, 69–82. [Google Scholar] [CrossRef]
- Chen, X.; Wales, P.; Quinti, L.; Zuo, F.; Moniot, S.; Herisson, F.; Rauf, N.A.; Wang, H.; Silverman, R.B.; Ayata, C.; et al. The Sirtuin-2 Inhibitor AK7 Is Neuroprotective in Models of Parkinson’s Disease but Not Amyotrophic Lateral Sclerosis and Cerebral Ischemia. PLoS ONE 2015, 10, e0116919. [Google Scholar] [CrossRef]
- Song, J.; Yang, B.; Jia, X.; Li, M.; Tan, W.; Ma, S.; Shi, X.; Feng, L. Distinctive Roles of Sirtuins on Diabetes, Protective or Detrimental? Front Endocrinol. 2018, 9, 724. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Dey, C.S. SIRT2 Negatively Regulates Insulin Resistance in C2C12 Skeletal Muscle Cells. Biochim. Biophys. Acta 2014, 1842, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Onyango, P.; Celic, I.; McCaffery, J.M.; Boeke, J.D.; Feinberg, A.P. SIRT3, a Human SIR2 Homologue, Is an NAD-Dependent Deacetylase Localized to Mitochondria. Proc. Natl. Acad. Sci. USA 2002, 99, 13653–13658. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.-H.; Kim, H.-S.; Song, S.; Lee, I.H.; Liu, J.; Vassilopoulos, A.; Deng, C.-X.; Finkel, T. A Role for the Mitochondrial Deacetylase Sirt3 in Regulating Energy Homeostasis. Proc. Natl. Acad. Sci. USA 2008, 105, 14447–14452. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.L.; Emerling, B.M.; Ricoult, S.J.H.; Guarente, L. SirT3 Suppresses Hypoxia Inducible Factor 1α and Tumor Growth by Inhibiting Mitochondrial ROS Production. Oncogene 2011, 30, 2986–2996. [Google Scholar] [CrossRef]
- Tao, R.; Coleman, M.C.; Pennington, J.D.; Ozden, O.; Park, S.-H.; Jiang, H.; Kim, H.-S.; Flynn, C.R.; Hill, S.; Hayes McDonald, W.; et al. Sirt3-Mediated Deacetylation of Evolutionarily Conserved Lysine 122 Regulates MnSOD Activity in Response to Stress. Mol. Cell 2010, 40, 893–904. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, L.L.; Wen, X.; Wang, X.Y.; Liu, J.; Cheng, Y.; Huang, J. Sirtuin-3 (SIRT3), a Therapeutic Target with Oncogenic and Tumor-Suppressive Function in Cancer. Cell Death Dis. 2014, 5, e1047. [Google Scholar] [CrossRef]
- Alhazzazi, T.Y.; Kamarajan, P.; Xu, Y.; Ai, T.; Chen, L.; Verdin, E.; Kapila, Y.L. A Novel Sirtuin-3 Inhibitor, LC-0296, Inhibits Cell Survival and Proliferation, and Promotes Apoptosis of Head and Neck Cancer Cells. Anticancer Res. 2016, 36, 49–60. [Google Scholar]
- Ma, J.; Liu, B.; Yu, D.; Zuo, Y.; Cai, R.; Yang, J.; Cheng, J. SIRT3 Deacetylase Activity Confers Chemoresistance in AML via Regulation of Mitochondrial Oxidative Phosphorylation. Br. J. Haematol. 2019, 187, 49–64. [Google Scholar] [CrossRef]
- Li, M.; Chiang, Y.-L.; Lyssiotis, C.A.; Teater, M.R.; Hong, J.Y.; Shen, H.; Wang, L.; Hu, J.; Jing, H.; Chen, Z.; et al. Non-Oncogene Addiction to SIRT3 Plays a Critical Role in Lymphomagenesis. Cancer Cell 2019, 35, 916–931.e9. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, Q.; Li, Z.; Yang, X.; Xia, W.; Chen, Z. Adjudin Synergizes with Paclitaxel and Inhibits Cell Growth and Metastasis by Regulating the Sirtuin 3-Forkhead Box O3a Axis in Human Small-Cell Lung Cancer. Thorac. Cancer 2019, 10, 642–658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zou, L.; Shi, D.; Liu, J.; Zhang, J.; Zhao, R.; Wang, G.; Zhang, L.; Ouyang, L.; Liu, B. Structure-Guided Design of a Small-Molecule Activator of Sirtuin-3 That Modulates Autophagy in Triple Negative Breast Cancer. J. Med. Chem. 2021, 64, 14192–14216. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-J.; Liu, X.-Y.; Li, J.-H.; Guo, J.; Li, F.; Gui, Y.; Li, X.-H.; Yang, L.; Wu, C.-Y.; Yuan, Y.; et al. Gastrodin Attenuates Microglia Activation through Renin-Angiotensin System and Sirtuin3 Pathway. Neurochem. Int. 2018, 120, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Burgess, J.D.; Faroqi, A.H.; DeMeo, N.N.; Fiesel, F.C.; Springer, W.; Delenclos, M.; McLean, P.J. Alpha-Synuclein-Induced Mitochondrial Dysfunction Is Mediated via a Sirtuin 3-Dependent Pathway. Mol. Neurodegener. 2020, 15, 5. [Google Scholar] [CrossRef]
- Ramesh, S.; Govindarajulu, M.; Lynd, T.; Briggs, G.; Adamek, D.; Jones, E.; Heiner, J.; Majrashi, M.; Moore, T.; Amin, R.; et al. SIRT3 Activator Honokiol Attenuates β-Amyloid by Modulating Amyloidogenic Pathway. PLoS ONE 2018, 13, e0190350. [Google Scholar] [CrossRef]
- Lee, A.Y.; Christensen, S.M.; Duong, N.; Tran, Q.-A.; Xiong, H.M.; Huang, J.; James, S.; Vallabh, D.; Talbott, G.; Rose, M.; et al. Sirt3 Pharmacologically Promotes Insulin Sensitivity through PI3/AKT/MTOR and Their Downstream Pathway in Adipocytes. Int. J. Mol. Sci. 2022, 23, 3740. [Google Scholar] [CrossRef]
- Huang, J.-Y.; Hirschey, M.D.; Shimazu, T.; Ho, L.; Verdin, E. Mitochondrial Sirtuins. Biochim. Biophys. Acta 2010, 1804, 1645–1651. [Google Scholar] [CrossRef]
- Jeong, S.M.; Hwang, S.; Seong, R.H. SIRT4 Regulates Cancer Cell Survival and Growth after Stress. Biochem. Biophys. Res. Commun. 2016, 470, 251–256. [Google Scholar] [CrossRef]
- He, Q.; Chen, K.; Ye, R.; Dai, N.; Guo, P.; Wang, L. Associations of Sirtuins with Clinicopathological Variables and Prognosis in Human Ovarian Cancer. Oncol. Lett. 2020, 19, 3278–3288. [Google Scholar] [CrossRef] [PubMed]
- Igci, M.; Kalender, M.E.; Borazan, E.; Bozgeyik, I.; Bayraktar, R.; Bozgeyik, E.; Camci, C.; Arslan, A. High-Throughput Screening of Sirtuin Family of Genes in Breast Cancer. Gene 2016, 586, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, Y.; Gao, J.; Yuan, X. Tumor-Suppressive Function of SIRT4 in Neuroblastoma through Mitochondrial Damage. Cancer Manag. Res. 2018, 10, 5591–5603. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ding, X.; Wu, Q.; Qi, J.; Zhu, M.; Miao, C. Monomethyltransferase SET8 Facilitates Hepatocellular Carcinoma Growth by Enhancing Aerobic Glycolysis. Cell Death Dis. 2019, 10, 312. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Qin, Y.; Ji, S.; Xu, W.; Liu, W.; Sun, Q.; Zhang, Z.; Liu, M.; Ni, Q.; Yu, X.; et al. UHRF1 Promotes Aerobic Glycolysis and Proliferation via Suppression of SIRT4 in Pancreatic Cancer. Cancer Lett. 2019, 452, 226–236. [Google Scholar] [CrossRef]
- Shih, J.; Liu, L.; Mason, A.; Higashimori, H.; Donmez, G. Loss of SIRT4 Decreases GLT-1-Dependent Glutamate Uptake and Increases Sensitivity to Kainic Acid. J. Neurochem. 2014, 131, 573–581. [Google Scholar] [CrossRef]
- Buck, E.; Bayer, H.; Lindenberg, K.S.; Hanselmann, J.; Pasquarelli, N.; Ludolph, A.C.; Weydt, P.; Witting, A. Comparison of Sirtuin 3 Levels in ALS and Huntington’s Disease-Differential Effects in Human Tissue Samples vs. Transgenic Mouse Models. Front. Mol. Neurosci. 2017, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Mostoslavsky, R.; Haigis, K.M.; Fahie, K.; Christodoulou, D.C.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; Karow, M.; Blander, G.; et al. SIRT4 Inhibits Glutamate Dehydrogenase and Opposes the Effects of Calorie Restriction in Pancreatic Beta Cells. Cell 2006, 126, 941–954. [Google Scholar] [CrossRef]
- Hutton, J.C.; Sener, A.; Malaisse, W.J. Interaction of Branched Chain Amino Acids and Keto Acids upon Pancreatic Islet Metabolism and Insulin Secretion. J. Biol. Chem. 1980, 255, 7340–7346. [Google Scholar] [CrossRef]
- Huynh, F.K.; Hu, X.; Lin, Z.; Johnson, J.D.; Hirschey, M.D. Loss of Sirtuin 4 Leads to Elevated Glucose- and Leucine-Stimulated Insulin Levels and Accelerated Age-Induced Insulin Resistance in Multiple Murine Genetic Backgrounds. J. Inherit. Metab. Dis. 2018, 41, 59–72. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Zha, X. Overview of SIRT5 as a Potential Therapeutic Target: Structure, Function and Inhibitors. Eur. J. Med. Chem. 2022, 236, 114363. [Google Scholar] [CrossRef]
- Abril, Y.L.N.; Fernandez, I.R.; Hong, J.Y.; Chiang, Y.-L.; Kutateladze, D.A.; Zhao, Q.; Yang, M.; Hu, J.; Sadhukhan, S.; Li, B.; et al. Pharmacological and Genetic Perturbation Establish SIRT5 as a Promising Target in Breast Cancer. Oncogene 2021, 40, 1644–1658. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Franzini, A.; Pomicter, A.D.; Halverson, B.J.; Antelope, O.; Mason, C.C.; Ahmann, J.M.; Senina, A.V.; Vellore, N.A.; Jones, C.L.; et al. SIRT5 is a druggable metabolic vulnerability in acute myeloid leukemia. Blood Cancer Discov. 2021, 2, 266–287. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Shukla, S.K.; Vernucci, E.; He, C.; Wang, D.; King, R.J.; Jha, K.; Siddhanta, K.; Mullen, N.J.; Attri, K.S.; et al. Metabolic Rewiring by Loss of Sirt5 Promotes Kras-Induced Pancreatic Cancer Progression. Gastroenterology 2021, 161, 1584–1600. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Peritore, C.; Ginsberg, J.; Shih, J.; Arun, S.; Donmez, G. Protective Role of SIRT5 against Motor Deficit and Dopaminergic Degeneration in MPTP-Induced Mice Model of Parkinson’s Disease. Behav. Brain Res. 2015, 281, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wei, Y.; Li, J.; Bai, Y.; Yin, P.; Wang, S. SIRT5 Represses Neurotrophic Pathways and Aβ Production in Alzheimer’s Disease by Targeting Autophagy. ACS Chem. Neurosci. 2021, 12, 4428–4437. [Google Scholar] [CrossRef]
- Wang, G.; Meyer, J.G.; Cai, W.; Softic, S.; Li, M.E.; Verdin, E.; Newgard, C.; Schilling, B.; Kahn, C.R. Regulation of UCP1 and Mitochondrial Metabolism in Brown Adipose Tissue by Reversible Succinylation. Mol. Cell 2019, 74, 844–857.e7. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Fei, X. SIRT5 Regulates Pancreatic β-Cell Proliferation and Insulin Secretion in Type 2 Diabetes. Exp. Ther. Med. 2018, 16, 1417–1425. [Google Scholar] [CrossRef]
- You, W.; Rotili, D.; Li, T.-M.; Kambach, C.; Meleshin, M.; Schutkowski, M.; Chua, K.F.; Mai, A.; Steegborn, C. Structural Basis of Sirtuin 6 Activation by Synthetic Small Molecules. Angew. Chem. Int. Ed. Engl. 2017, 56, 1007–1011. [Google Scholar] [CrossRef]
- Iachettini, S.; Trisciuoglio, D.; Rotili, D.; Lucidi, A.; Salvati, E.; Zizza, P.; di Leo, L.; del Bufalo, D.; Ciriolo, M.R.; Leonetti, C.; et al. Pharmacological Activation of SIRT6 Triggers Lethal Autophagy in Human Cancer Cells. Cell Death Dis. 2018, 9, 996. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, J.; Deng, W.; Chen, Y.; Shang, J.; Song, K.; Zhang, L.; Wang, C.; Lu, S.; Yang, X.; et al. Identification of a Cellularly Active SIRT6 Allosteric Activator. Nat. Chem. Biol. 2018, 14, 1118–1126. [Google Scholar] [CrossRef]
- Shang, J.-L.; Ning, S.-B.; Chen, Y.-Y.; Chen, T.-X.; Zhang, J. MDL-800, an Allosteric Activator of SIRT6, Suppresses Proliferation and Enhances EGFR-TKIs Therapy in Non-Small Cell Lung Cancer. Acta Pharmacol. Sin. 2021, 42, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Zhu, Z.; Chen, Y.; Song, J.; Huang, Y.; Song, K.; Zhong, J.; Xu, X.; Wei, J.; Wang, C.; et al. Small-Molecule Activating SIRT6 Elicits Therapeutic Effects and Synergistically Promotes Anti-Tumor Activity of Vitamin D3 in Colorectal Cancer. Theranostics 2020, 10, 5845–5864. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sun, W.; Huang, S.; Zhang, H.; Lin, G.; Li, H.; Qiao, J.; Li, L.; Yang, S. Discovery of Potent Small-Molecule SIRT6 Activators: Structure-Activity Relationship and Anti-Pancreatic Ductal Adenocarcinoma Activity. J. Med. Chem. 2020, 63, 10474–10495. [Google Scholar] [CrossRef] [PubMed]
- Sociali, G.; Galeno, L.; Parenti, M.D.; Grozio, A.; Bauer, I.; Passalacqua, M.; Boero, S.; Donadini, A.; Millo, E.; Bellotti, M.; et al. Quinazolinedione SIRT6 Inhibitors Sensitize Cancer Cells to Chemotherapeutics. Eur. J. Med. Chem. 2015, 102, 530–539. [Google Scholar] [CrossRef]
- Damonte, P.; Sociali, G.; Parenti, M.D.; Soncini, D.; Bauer, I.; Boero, S.; Grozio, A.; von Holtey, M.; Piacente, F.; Becherini, P.; et al. SIRT6 Inhibitors with Salicylate-like Structure Show Immunosuppressive and Chemosensitizing Effects. Bioorg. Med. Chem. 2017, 25, 5849–5858. [Google Scholar] [CrossRef]
- Cagnetta, A.; Soncini, D.; Orecchioni, S.; Talarico, G.; Minetto, P.; Guolo, F.; Retali, V.; Colombo, N.; Carminati, E.; Clavio, M.; et al. Depletion of SIRT6 Enzymatic Activity Increases Acute Myeloid Leukemia Cells’ Vulnerability to DNA-Damaging Agents. Haematologica 2018, 103, 80–90. [Google Scholar] [CrossRef]
- Nicholatos, J.W.; Francisco, A.B.; Bender, C.A.; Yeh, T.; Lugay, F.J.; Salazar, J.E.; Glorioso, C.; Libert, S. Nicotine Promotes Neuron Survival and Partially Protects from Parkinson’s Disease by Suppressing SIRT6. Acta Neuropathol. Commun. 2018, 6, 120. [Google Scholar] [CrossRef]
- Ferrara, G.; Benzi, A.; Sturla, L.; Marubbi, D.; Frumento, D.; Spinelli, S.; Abbotto, E.; Ivaldi, F.; von Holtey, M.; Murone, M.; et al. Sirt6 Inhibition Delays the Onset of Experimental Autoimmune Encephalomyelitis by Reducing Dendritic Cell Migration. J Neuroinflamm. 2020, 17, 228. [Google Scholar] [CrossRef]
- Parenti, M.D.; Grozio, A.; Bauer, I.; Galeno, L.; Damonte, P.; Millo, E.; Sociali, G.; Franceschi, C.; Ballestrero, A.; Bruzzone, S.; et al. Discovery of Novel and Selective SIRT6 Inhibitors. J. Med. Chem. 2014, 57, 4796–4804. [Google Scholar] [CrossRef]
- Sociali, G.; Magnone, M.; Ravera, S.; Damonte, P.; Vigliarolo, T.; von Holtey, M.; Vellone, V.G.; Millo, E.; Caffa, I.; Cea, M.; et al. Pharmacological Sirt6 Inhibition Improves Glucose Tolerance in a Type 2 Diabetes Mouse Model. FASEB J. 2017, 31, 3138–3149. [Google Scholar] [CrossRef]
- Sun, W.; Chen, X.; Huang, S.; Li, W.; Tian, C.; Yang, S.; Li, L. Discovery of 5-(4-Methylpiperazin-1-Yl)-2-Nitroaniline Derivatives as a New Class of SIRT6 Inhibitors. Bioorg. Med. Chem. Lett. 2020, 30, 127215. [Google Scholar] [CrossRef] [PubMed]
- Grob, A.; Roussel, P.; Wright, J.E.; McStay, B.; Hernandez-Verdun, D.; Sirri, V. Involvement of SIRT7 in Resumption of RDNA Transcription at the Exit from Mitosis. J. Cell Sci. 2009, 122, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Ford, E.; Voit, R.; Liszt, G.; Magin, C.; Grummt, I.; Guarente, L. Mammalian Sir2 Homolog SIRT7 Is an Activator of RNA Polymerase I Transcription. Genes Dev. 2006, 20, 1075–1080. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Liu, B.; Ning, C.; Li, Y.; Wang, Y.; Li, Z. Discovery of SIRT7 Inhibitor as New Therapeutic Options Against Liver Cancer. Front. Cell Dev. Biol. 2021, 9, 813233. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Kim, D.; Cho, S.J.; Jung, K.-Y.; Kim, J.-H.; Lee, J.M.; Jung, H.J.; Kim, K.R. Identification of a Novel SIRT7 Inhibitor as Anticancer Drug Candidate. Biochem. Biophys. Res. Commun. 2019, 508, 451–457. [Google Scholar] [CrossRef]
- Vazquez, B.N.; Thackray, J.K.; Serrano, L. Sirtuins and DNA Damage Repair: SIRT7 Comes to Play. Nucleus 2017, 8, 107–115. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Lu, L.; Huang, T.; Hou, W.; Wang, F.; Yu, L.; Wu, F.; Qi, J.; Chen, X.; et al. Sirt7 Associates with ELK1 to Participate in Hyperglycemia Memory and Diabetic Nephropathy via Modulation of DAPK3 Expression and Endothelial Inflammation. Transl. Res. 2022, 247, 99–116. [Google Scholar] [CrossRef]
- Heltweg, B.; Gatbonton, T.; Schuler, A.D.; Posakony, J.; Li, H.; Goehle, S.; Kollipara, R.; Depinho, R.A.; Gu, Y.; Simon, J.A.; et al. Antitumor Activity of a Small-Molecule Inhibitor of Human Silent Information Regulator 2 Enzymes. Cancer Res. 2006, 66, 4368–4377. [Google Scholar] [CrossRef]
- Lu, B.; Zhang, D.; Wang, X.; Lin, D.; Chen, Y.; Xu, X. Targeting SIRT1 to Inhibit the Proliferation of Multiple Myeloma Cells. Oncol. Lett. 2021, 21, 306. [Google Scholar] [CrossRef]
- Portmann, S.; Fahrner, R.; Lechleiter, A.; Keogh, A.; Overney, S.; Laemmle, A.; Mikami, K.; Montani, M.; Tschan, M.P.; Candinas, D.; et al. Antitumor Effect of SIRT1 Inhibition in Human HCC Tumor Models in Vitro and in Vivo. Mol. Cancer Ther. 2013, 12, 499–508. [Google Scholar] [CrossRef]
- Marshall, G.M.; Liu, P.Y.; Gherardi, S.; Scarlett, C.J.; Bedalov, A.; Xu, N.; Iraci, N.; Valli, E.; Ling, D.; Thomas, W.; et al. SIRT1 Promotes N-Myc Oncogenesis through a Positive Feedback Loop Involving the Effects of MKP3 and ERK on N-Myc Protein Stability. PLoS Genet 2011, 7, e1002135. [Google Scholar] [CrossRef] [PubMed]
- Holloway, K.R.; Barbieri, A.; Malyarchuk, S.; Saxena, M.; Nedeljkovic-Kurepa, A.; Cameron Mehl, M.; Wang, A.; Gu, X.; Pruitt, K. SIRT1 Positively Regulates Breast Cancer Associated Human Aromatase (CYP19A1) Expression. Mol. Endocrinol. 2013, 27, 480–490. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.R.; Pirrie, L.; Hollick, J.J.; Ronseaux, S.; Campbell, J.; Higgins, M.; Staples, O.D.; Tran, F.; Slawin, A.M.Z.; Lain, S.; et al. Synthesis and Biological Characterisation of Sirtuin Inhibitors Based on the Tenovins. Bioorg. Med. Chem. 2012, 20, 1779–1793. [Google Scholar] [CrossRef]
- Ke, X.; Qin, Q.; Deng, T.; Liao, Y.; Gao, S.-J. Heterogeneous Responses of Gastric Cancer Cell Lines to Tenovin-6 and Synergistic Effect with Chloroquine. Cancers 2020, 12, 365. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.B.; Kim, Y.; Kim, D.; Cho, E.Y.; Han, J.; Kim, H.K.; Shim, Y.M.; Kim, D.-H. Metformin and Tenovin-6 Synergistically Induces Apoptosis through LKB1-Independent SIRT1 down-Regulation in Non-Small Cell Lung Cancer Cells. J. Cell Mol. Med. 2019, 23, 2872–2889. [Google Scholar] [CrossRef]
- Yuan, H.; Wang, Z.; Li, L.; Zhang, H.; Modi, H.; Horne, D.; Bhatia, R.; Chen, W. Activation of Stress Response Gene SIRT1 by BCR-ABL Promotes Leukemogenesis. Blood 2012, 119, 1904–1914. [Google Scholar] [CrossRef]
- Schnekenburger, M.; Goffin, E.; Lee, J.-Y.; Jang, J.Y.; Mazumder, A.; Ji, S.; Rogister, B.; Bouider, N.; Lefranc, F.; Miklos, W.; et al. Discovery and Characterization of R/S-N-3-Cyanophenyl-N’-(6-Tert-Butoxycarbonylamino-3,4-Dihydro-2,2-Dimethyl-2H-1-Benzopyran-4-Yl)Urea, a New Histone Deacetylase Class III Inhibitor Exerting Antiproliferative Activity against Cancer Cell Lines. J. Med. Chem. 2017, 60, 4714–4733. [Google Scholar] [CrossRef]
- Laaroussi, H.; Ding, Y.; Teng, Y.; Deschamps, P.; Vidal, M.; Yu, P.; Broussy, S. Synthesis of Indole Inhibitors of Silent Information Regulator 1 (SIRT1), and Their Evaluation as Cytotoxic Agents. Eur. J. Med. Chem. 2020, 202, 112561. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Chao, E.D.; Blackwell, H.E.; Moazed, D.; Schreiber, S.L. Identification of a Class of Small Molecule Inhibitors of the Sirtuin Family of NAD-Dependent Deacetylases by Phenotypic Screening. J. Biol. Chem. 2001, 276, 38837–38843. [Google Scholar] [CrossRef]
- Ota, H.; Tokunaga, E.; Chang, K.; Hikasa, M.; Iijima, K.; Eto, M.; Kozaki, K.; Akishita, M.; Ouchi, Y.; Kaneki, M. Sirt1 Inhibitor, Sirtinol, Induces Senescence-like Growth Arrest with Attenuated Ras-MAPK Signaling in Human Cancer Cells. Oncogene 2006, 25, 176–185. [Google Scholar] [CrossRef]
- Zhou, W.; Ni, T.K.; Wronski, A.; Glass, B.; Skibinski, A.; Beck, A.; Kuperwasser, C. The SIRT2 Deacetylase Stabilizes Slug to Control Malignancy of Basal-like Breast Cancer. Cell Rep. 2016, 17, 1302–1317. [Google Scholar] [CrossRef]
- Kojima, K.; Ohhashi, R.; Fujita, Y.; Hamada, N.; Akao, Y.; Nozawa, Y.; Deguchi, T.; Ito, M. A Role for SIRT1 in Cell Growth and Chemoresistance in Prostate Cancer PC3 and DU145 Cells. Biochem. Biophys. Res. Commun. 2008, 373, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Lara, E.; Mai, A.; Calvanese, V.; Altucci, L.; Lopez-Nieva, P.; Martinez-Chantar, M.L.; Varela-Rey, M.; Rotili, D.; Nebbioso, A.; Ropero, S.; et al. Salermide, a Sirtuin Inhibitor with a Strong Cancer-Specific Proapoptotic Effect. Oncogene 2009, 28, 781–791. [Google Scholar] [CrossRef]
- Liu, G.; Su, L.; Hao, X.; Zhong, N.; Zhong, D.; Singhal, S.; Liu, X. Salermide Up-Regulates Death Receptor 5 Expression through the ATF4-ATF3-CHOP Axis and Leads to Apoptosis in Human Cancer Cells. J. Cell Mol. Med. 2012, 16, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.J.; Lee, Y.T.; Yeong, K.Y.; Petersen, S.H.; Kono, K.; Tan, S.C.; Oon, C.E. Anticancer Activities of a Benzimidazole Compound through Sirtuin Inhibition in Colorectal Cancer. Future Med. Chem. 2018, 10, 2039–2057. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, N.A.; Hong, J.Y.; Hu, J.; Jing, H.; Wang, M.; Price, I.R.; Cao, J.; Yang, M.; Zhang, X.; Lin, H. A Small-Molecule SIRT2 Inhibitor That Promotes K-Ras4a Lysine Fatty-Acylation. ChemMedChem 2019, 14, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.T.; Gertz, M.; Steegborn, C. Crystal Structures of Sirt3 Complexes with 4’-Bromo-Resveratrol Reveal Binding Sites and Inhibition Mechanism. Chem. Biol. 2013, 20, 1375–1385. [Google Scholar] [CrossRef]
- George, J.; Nihal, M.; Singh, C.K.; Ahmad, N. 4’-Bromo-Resveratrol, a Dual Sirtuin-1 and Sirtuin-3 Inhibitor, Inhibits Melanoma Cell Growth through Mitochondrial Metabolic Reprogramming. Mol. Carcinog. 2019, 58, 1876–1885. [Google Scholar] [CrossRef]
- Hui, Q.; Li, X.; Fan, W.; Gao, C.; Zhang, L.; Qin, H.; Wei, L.; Zhang, L. Discovery of 2-(4-Acrylamidophenyl)-Quinoline-4-Carboxylic Acid Derivatives as Potent SIRT3 Inhibitors. Front. Chem. 2022, 10, 880067. [Google Scholar] [CrossRef]
- Audrito, V.; Vaisitti, T.; Rossi, D.; Gottardi, D.; D’Arena, G.; Laurenti, L.; Gaidano, G.; Malavasi, F.; Deaglio, S. Nicotinamide Blocks Proliferation and Induces Apoptosis of Chronic Lymphocytic Leukemia Cells through Activation of the P53/MiR-34a/SIRT1 Tumor Suppressor Network. Cancer Res. 2011, 71, 4473–4483. [Google Scholar] [CrossRef]
- Alhazzazi, T.Y.; Kamarajan, P.; Joo, N.; Huang, J.-Y.; Verdin, E.; D’Silva, N.J.; Kapila, Y.L. Sirtuin-3 (SIRT3), a Novel Potential Therapeutic Target for Oral Cancer. Cancer 2011, 117, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Contoyiannis, P.A.; Draginis, E.; Adamopoulos, D.A.; Triantafyllou, G. Letter: Levodopa in Coma Due to Fulminant Hepatitis. Br. Med. J. 1975, 1, 272. [Google Scholar] [CrossRef] [PubMed]
- Jung-Hynes, B.; Nihal, M.; Zhong, W.; Ahmad, N. Role of Sirtuin Histone Deacetylase SIRT1 in Prostate Cancer. A Target for Prostate Cancer Management via Its Inhibition? J. Biol. Chem. 2009, 284, 3823–3832. [Google Scholar] [CrossRef]
- Liu, D.; Pitta, M.; Jiang, H.; Lee, J.-H.; Zhang, G.; Chen, X.; Kawamoto, E.M.; Mattson, M.P. Nicotinamide Forestalls Pathology and Cognitive Decline in Alzheimer Mice: Evidence for Improved Neuronal Bioenergetics and Autophagy Procession. Neurobiol. Aging 2013, 34, 1564–1580. [Google Scholar] [CrossRef] [PubMed]
- Green, K.N.; Steffan, J.S.; Martinez-Coria, H.; Sun, X.; Schreiber, S.S.; Thompson, L.M.; LaFerla, F.M. Nicotinamide Restores Cognition in Alzheimer’s Disease Transgenic Mice via a Mechanism Involving Sirtuin Inhibition and Selective Reduction of Thr231-Phosphotau. J. Neurosci. 2008, 28, 11500–11510. [Google Scholar] [CrossRef] [PubMed]
- Hathorn, T.; Snyder-Keller, A.; Messer, A. Nicotinamide Improves Motor Deficits and Upregulates PGC-1α and BDNF Gene Expression in a Mouse Model of Huntington’s Disease. Neurobiol. Dis. 2011, 41, 43–50. [Google Scholar] [CrossRef]
- Lee, S.-J.; Choi, S.-E.; Jung, I.-R.; Lee, K.-W.; Kang, Y. Protective Effect of Nicotinamide on High Glucose/Palmitate-Induced Glucolipotoxicity to INS-1 Beta Cells Is Attributed to Its Inhibitory Activity to Sirtuins. Arch. Biochem. Biophys. 2013, 535, 187–196. [Google Scholar] [CrossRef]
- PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 14 April 2022).
- Document Search-Web of Science Core Collection. Available online: https://www.webofscience.com/wos/woscc/basic-search (accessed on 14 April 2022).
- Huhtiniemi, T.; Suuronen, T.; Rinne, V.M.; Wittekindt, C.; Lahtela-Kakkonen, M.; Jarho, E.; Wallén, E.A.A.; Salminen, A.; Poso, A.; Leppänen, J. Oxadiazole-Carbonylaminothioureas as SIRT1 and SIRT2 Inhibitors. J. Med. Chem. 2008, 51, 4377–4380. [Google Scholar] [CrossRef]
- Huhtiniemi, T.; Wittekindt, C.; Laitinen, T.; Leppänen, J.; Salminen, A.; Poso, A.; Lahtela-Kakkonen, M. Comparative and Pharmacophore Model for Deacetylase SIRT1. J. Comput. Aided Mol. Des. 2006, 20, 589–599. [Google Scholar] [CrossRef]
- Sakkiah, S.; Krishnamoorthy, N.; Gajendrarao, P.; Thangapandian, S.; Lee, Y.; Kim, S.; Suh, J.K.; Kim, H.H.; Lee, K.W. Pharmacophore Mapping and Virtual Screening for SIRT1 Activators. Bull. Korean Chem. Soc. 2009, 30, 1152–1156. [Google Scholar]
- Maybridge Library | Thermo Fisher Scientific-IT. Available online: https://www.thermofisher.com/it/en/home/global/forms/lab-solutions/maybridge-library.html (accessed on 13 May 2022).
- Sakkiah, S.; Arooj, M.; Lee, K.W.; Torres, J.Z. Theoretical Approaches to Identify the Potent Scaffold for Human Sirtuin1 Activator: Bayesian Modeling and Density Functional Theory. Med. Chem. Res. 2014, 23, 3998–4010. [Google Scholar] [CrossRef]
- Vyas, V.K.; Goel, A.; Ghate, M.; Patel, P. Ligand and Structure-Based Approaches for the Identification of SIRT1 Activators. Chem. Biol. Interact. 2015, 228, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Shoichet, B.K. ZINC-A Free Database of Commercially Available Compounds for Virtual Screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Azminah, A.; Erlina, L.; Radji, M.; Mun’im, A.; Syahdi, R.R.; Yanuar, A. In Silico and in Vitro Identification of Candidate SIRT1 Activators from Indonesian Medicinal Plants Compounds Database. Comput. Biol. Chem. 2019, 83, 107096. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Case, A.W.; Riera, T.V.; Considine, T.; Lee, J.E.; Hamuro, Y.; Zhao, H.; Jiang, Y.; Sweitzer, S.M.; Pietrak, B.; et al. Crystallographic Structure of a Small Molecule SIRT1 Activator-Enzyme Complex. Nat. Commun. 2015, 6, 7645. [Google Scholar] [CrossRef]
- Basis Data Tanaman Obat Indonesia. Available online: http://herbaldb.farmasi.ui.ac.id/v3/ (accessed on 13 May 2022).
- Zhao, X.; Allison, D.; Condon, B.; Zhang, F.; Gheyi, T.; Zhang, A.; Ashok, S.; Russell, M.; MacEwan, I.; Qian, Y.; et al. The 2.5 Å Crystal Structure of the SIRT1 Catalytic Domain Bound to Nicotinamide Adenine Dinucleotide (NAD+) and an Indole (EX527 Analogue) Reveals a Novel Mechanism of Histone Deacetylase Inhibition. J. Med. Chem. 2013, 56, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Asinex.Com-Home. Available online: https://www.asinex.com/ (accessed on 13 May 2022).
- Pulla, V.K.; Alvala, M.; Sriram, D.S.; Viswanadha, S.; Sriram, D.; Yogeeswari, P. Structure-Based Drug Design of Small Molecule SIRT1 Modulators to Treat Cancer and Metabolic Disorders. J. Mol. Graph. Model. 2014, 52, 46–56. [Google Scholar] [CrossRef]
- Autiero, I.; Costantini, S.; Colonna, G. Human Sirt-1: Molecular Modeling and Structure-Function Relationships of an Unordered Protein. PLoS ONE 2009, 4, e7350. [Google Scholar] [CrossRef]
- Padmanabhan, B.; Ramu, M.; Mathur, S.; Unni, S.; Thiyagarajan, S. Identification of New Inhibitors for Human SIRT1: An in-Silico Approach. Med. Chem. 2016, 12, 347–361. [Google Scholar] [CrossRef]
- Davenport, A.M.; Huber, F.M.; Hoelz, A. Structural and Functional Analysis of Human SIRT1. J. Mol. Biol. 2014, 426, 526–541. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Shrivastava, S.; Hassanali, M.; Stothard, P.; Chang, Z.; Woolsey, J. DrugBank: A Comprehensive Resource for in Silico Drug Discovery and Exploration. Nucleic Acids Res. 2006, 34, D668–D672. [Google Scholar] [CrossRef] [PubMed]
- Princeton BioMolecular Research, Inc. Available online: https://princetonbio.com/ (accessed on 13 May 2022).
- Ertan-Bolelli, T.; Bolelli, K. In Silico Design of Novel Sirtuin 1 Enzyme Activators for the Treatment of Age-Related Diseases and Life Span. Curr. Comput. Aided Drug. Des. 2021, 17, 412–420. [Google Scholar] [CrossRef]
- Wang, F.; Liu, D.; Wang, H.; Luo, C.; Zheng, M.; Liu, H.; Zhu, W.; Luo, X.; Zhang, J.; Jiang, H. Computational Screening for Active Compounds Targeting Protein Sequences: Methodology and Experimental Validation. J. Chem. Inf. Model. 2011, 51, 2821–2828. [Google Scholar] [CrossRef] [PubMed]
- Vapnik, V. The Nature of Statistical Learning Theory; Springer: New York, NY, USA, 1999. [Google Scholar]
- Zheng, M.; Liu, Z.; Xue, C.; Zhu, W.; Chen, K.; Luo, X.; Jiang, H. Mutagenic Probability Estimation of Chemical Compounds by a Novel Molecular Electrophilicity Vector and Support Vector Machine. Bioinformatics 2006, 22, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Gertrudes, J.C.; Maltarollo, V.G.; Silva, R.A.; Oliveira, P.R.; Honório, K.M.; da Silva, A.B.F. Machine Learning Techniques and Drug Design. Curr. Med. Chem. 2012, 19, 4289–4297. [Google Scholar] [CrossRef] [PubMed]
- Specs-Compound Management Services and Research Compounds for the Life Science Industry. Available online: https://specs.net/ (accessed on 13 May 2022).
- Sun, Y.; Zhou, H.; Zhu, H.; Leung, S. Ligand-Based Virtual Screening and Inductive Learning for Identification of SIRT1 Inhibitors in Natural Products. Sci. Rep. 2016, 6, 19312. [Google Scholar] [CrossRef]
- Sanderson, K. Databases Aim to Bridge the East-West Divide of Drug Discovery. Nat. Med. 2011, 17, 1531. [Google Scholar] [CrossRef]
- Xue, R.; Fang, Z.; Zhang, M.; Yi, Z.; Wen, C.; Shi, T. TCMID: Traditional Chinese Medicine Integrative Database for Herb Molecular Mechanism Analysis. Nucleic Acids Res. 2013, 41, D1089–D1095. [Google Scholar] [CrossRef]
- Chiba, S.; Ohue, M.; Gryniukova, A.; Borysko, P.; Zozulya, S.; Yasuo, N.; Yoshino, R.; Ikeda, K.; Shin, W.H.; Kihara, D.; et al. A Prospective Compound Screening Contest Identified Broader Inhibitors for Sirtuin 1. Sci. Rep. 2019, 9, 19585. [Google Scholar] [CrossRef]
- Uciechowska, U.; Schemies, J.; Neugebauer, R.C.; Huda, E.-M.; Schmitt, M.L.; Meier, R.; Verdin, E.; Jung, M.; Sippl, W. Thiobarbiturates as Sirtuin Inhibitors: Virtual Screening, Free-Energy Calculations, and Biological Testing. ChemMedChem 2008, 3, 1965–1976. [Google Scholar] [CrossRef]
- ChemBridge | Home. Available online: https://www.chembridge.com/ (accessed on 13 May 2022).
- Uciechowska, U.; Schemies, J.; Scharfe, M.; Lawson, M.; Wichapong, K.; Jung, M.; Sippl, W. Binding Free Energy Calculations and Biological Testing of Novel Thiobarbiturates as Inhibitors of the Human NAD+ Dependent Histone Deacetylase Sirt2. MedChemComm 2012, 3, 167–173. [Google Scholar] [CrossRef]
- Sivaraman, P.; Mattegunta, S.; Subbaraju, G.V.; Satyanarayana, C.; Padmanabhan, B. Design of a Novel Nucleoside Analog as Potent Inhibitor of the NAD+ Dependent Deacetylase, SIRT2. Syst. Synth. Biol. 2010, 4, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Home-NCI Wiki. Available online: https://wiki.nci.nih.gov/dashboard.action (accessed on 13 May 2022).
- Sacconnay, L.; Ryckewaert, L.; Randazzo, G.M.; Petit, C.; Passos, C.D.S.; Jachno, J.; Michailovienė, V.; Zubrienė, A.; Matulis, D.; Carrupt, P.-A.; et al. 5-Benzylidene-Hydantoin Is a New Scaffold for SIRT Inhibition: From Virtual Screening to Activity Assays. Eur. J. Pharm. Sci. 2016, 85, 59–67. [Google Scholar] [CrossRef]
- Moniot, S.; Schutkowski, M.; Steegborn, C. Crystal Structure Analysis of Human Sirt2 and Its ADP-Ribose Complex. J. Struct. Biol. 2013, 182, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Tervo, A.J.; Kyrylenko, S.; Niskanen, P.; Salminen, A.; Leppänen, J.; Nyrönen, T.H.; Järvinen, T.; Poso, A. An In Silico Approach to Discovering Novel Inhibitors of Human Sirtuin Type 2. J. Med. Chem. 2004, 47, 6292–6298. [Google Scholar] [CrossRef] [PubMed]
- Tervo, A.J.; Suuronen, T.; Kyrylenko, S.; Kuusisto, E.; Kiviranta, P.H.; Salminen, A.; Leppanen, J.; Poso, A. Discovering Inhibitors of Human Sirtuin Type 2: Novel Structural Scaffolds. J. Med. Chem. 2006, 49, 7239–7241. [Google Scholar] [CrossRef] [PubMed]
- Sakkiah, S.; Thangapandian, S.; Park, C.; Son, M.; Lee, K.W. Molecular Docking and Dynamics Simulation, Receptor-Based Hypothesis: Application to Identify Novel Sirtuin 2 Inhibitors. Chem. Biol. Drug Des. 2012, 80, 315–327. [Google Scholar] [CrossRef]
- CHEMDIV INC-FULLY INTEGRATED TARGET-TO-CLINIC CONTRACT RESEARCH ORGANIZATION (CRO). Available online: https://www.chemdiv.com/ (accessed on 13 May 2022).
- Sakkiah, S.; Baek, A.; Lee, K.W. Pharmacophore Modeling and Molecular Dynamics Simulation to Identify the Critical Chemical Features against Human Sirtuin 2 Inhibitors. J. Mol. Struct. 2012, 1011, 66–75. [Google Scholar] [CrossRef]
- Schiedel, M.; Rumpf, T.; Karaman, B.; Lehotzky, A.; Oláh, J.; Gerhardt, S.; Ovádi, J.; Sippl, W.; Einsle, O.; Jung, M. Aminothiazoles as Potent and Selective Sirt2 Inhibitors: A Structure–Activity Relationship Study. J. Med. Chem. 2016, 59, 1599–1612. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 2. Enrichment Factors in Database Screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Kouznetsova, V.L.; Tsigelny, I.F. Targeting Epigenetic Regulators Using Machine Learning: Potential Sirtuin 2 Inhibitors. J. Comput. Biophys. Chem. 2021, 20, 841–851. [Google Scholar] [CrossRef]
- Khanfar, M.A.; Alqtaishat, S. Discovery of Potent Natural-Product-Derived SIRT2 Inhibitors Using Structure- Based Exploration of SIRT2 Pharmacophoric Space Coupled With QSAR Analyses. Anticancer Agents Med. Chem. 2021, 21, 2278–2286. [Google Scholar] [CrossRef] [PubMed]
- Triballeau, N.; Acher, F.; Brabet, I.; Pin, J.P.; Bertrand, H.O. Virtual Screening Workflow Development Guided by the “Receiver Operating Characteristic” Curve Approach. Application to High-Throughput Docking on Metabotropic Glutamate Receptor Subtype 4. J. Med. Chem. 2005, 48, 2534–2547. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.; Hopfinger, A.J. Application of Genetic Function Approximation to Quantitative Structure-Activity Relationships and Quantitative Structure-Property Relationships. J. Chem. Inf. Comput. Sci. 1994, 34, 854–866. [Google Scholar] [CrossRef]
- Taha, M.O.; Bustanji, Y.; Al-Ghussein, M.A.S.; Mohammad, M.; Zalloum, H.; Al-Masri, I.M.; Atallah, N. Pharmacophore Modeling, Quantitative Structure–Activity Relationship Analysis, and in Silico Screening Reveal Potent Glycogen Synthase Kinase-3β Inhibitory Activities for Cimetidine, Hydroxychloroquine, and Gemifloxacin. J. Med. Chem. 2008, 51, 2062–2077. [Google Scholar] [CrossRef] [PubMed]
- Khanfar, M.A.; Banat, F.; Alabed, S.; Alqtaishat, S. Discovery of Potent NEK2 Inhibitors as Potential Anticancer Agents Using Structure-Based Exploration of NEK2 Pharmacophoric Space Coupled with QSAR Analyses. Mol. Divers. 2017, 21, 187–200. [Google Scholar] [CrossRef]
- Khanfar, M.A.; Alqtaishat, S. Discovery of Potent IRAK-4 Inhibitors as Potential Anti-Inflammatory and Anticancer Agents Using Structure-Based Exploration of IRAK-4 Pharmacophoric Space Coupled with QSAR Analyses. Comput. Biol. Chem. 2019, 79, 147–154. [Google Scholar] [CrossRef]
- MEGx Purified Natural Product Screening Compounds-AnalytiCon Discovery. Available online: https://ac-discovery.com/purified-natural-product-screening-compounds/ (accessed on 13 May 2022).
- Kokkonen, P.; Kokkola, T.; Suuronen, T.; Poso, A.; Jarho, E.; Lahtela-Kakkonen, M. Virtual Screening Approach of Sirtuin Inhibitors Results in Two New Scaffolds. Eur. J. Pharm. Sci. 2015, 76, 27–32. [Google Scholar] [CrossRef]
- Salo, H.S.; Laitinen, T.; Poso, A.; Jarho, E.; Lahtela-Kakkonen, M. Identification of Novel SIRT3 Inhibitor Scaffolds by Virtual Screening. Bioorg. Med. Chem. Lett. 2013, 23, 2990–2995. [Google Scholar] [CrossRef]
- Choubey, S.K.; Prabhu, D.; Nachiappan, M.; Biswal, J.; Jeyakanthan, J. Molecular Modeling, Dynamics Studies and Density Functional Theory Approaches to Identify Potential Inhibitors of SIRT4 Protein from Homo Sapiens: A Novel Target for the Treatment of Type 2 Diabetes. J. Biomol. Struct. Dyn. 2017, 35, 3316–3329. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ji, S.; Yu, Z.-J.; Wang, H.-L.; Cheng, X.; Li, W.-J.; Jing, L.; Yu, Y.; Chen, Q.; Yang, L.-L.; et al. Structure-Based Discovery of New Selective Small-Molecule Sirtuin 5 Inhibitors. Chem. Biol. Drug Des. 2018, 91, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Tenhunen, J.; Kučera, T.; Huovinen, M.; Küblbeck, J.; Bisenieks, E.; Vigante, B.; Ogle, Z.; Duburs, G.; Doležal, M.; Moaddel, R.; et al. Screening of SIRT6 Inhibitors and Activators: A Novel Activator Has an Impact on Breast Cancer Cells. Biomed. Pharmacother. 2021, 138, 111452. [Google Scholar] [CrossRef] [PubMed]
- Schlicker, C.; Boanca, G.; Lakshminarasimhan, M.; Steegborn, C. Structure-Based Development of Novel Sirtuin Inhibitors. Aging 2011, 3, 852–872. [Google Scholar] [CrossRef]
- Karaman, B.; Alhalabi, Z.; Swyter, S.; Mihigo, S.; Andrae-Marobela, K.; Jung, M.; Sippl, W.; Ntie-Kang, F. Identification of Bichalcones as Sirtuin Inhibitors by Virtual Screening and In Vitro Testing. Molecules 2018, 23, 416. [Google Scholar] [CrossRef]
- Sinha, S.; Patel, S.; Athar, M.; Vora, J.; Chhabria, M.T.; Jha, P.C.; Shrivastava, N. Structure-Based Identification of Novel Sirtuin Inhibitors against Triple Negative Breast Cancer: An in Silico and in Vitro Study. Int. J. Biol. Macromol. 2019, 140, 454–468. [Google Scholar] [CrossRef]
- SARI, S.; AVCI, A.; KOÇAK ASLAN, E. In Silico Repurposing of Drugs for Pan-HDAC and Pan-SIRT Inhibitors: Consensus Structure-Based Virtual Screening and Pharmacophore Modeling Investigations. Turk. J. Pharm. Sci. 2021, 18, 730–737. [Google Scholar] [CrossRef]
- Kadam, R.U.; Tavares, J.; Kiran, V.M.; Cordeiro, A.; Ouaissi, A.; Roy, N. Structure Function Analysis of Leishmania Sirtuin: An Ensemble of In Silico and Biochemical Studies. Chem. Biol. Drug Des. 2008, 71, 501–506. [Google Scholar] [CrossRef]
- Soares, M.B.P.; Silva, C.V.; Bastos, T.M.; Guimarães, E.T.; Figueira, C.P.; Smirlis, D.; Azevedo, W.F. Anti-Trypanosoma Cruzi Activity of Nicotinamide. Acta Trop. 2012, 122, 224–229. [Google Scholar] [CrossRef]
- Sacconnay, L.; Angleviel, M.; Randazzo, G.M.; Marçal Ferreira Queiroz, M.; Ferreira Queiroz, E.; Wolfender, J.-L.; Carrupt, P.-A.; Nurisso, A. Computational Studies on Sirtuins from Trypanosoma Cruzi: Structures, Conformations and Interactions with Phytochemicals. PLoS Negl. Trop. Dis. 2014, 8, e2689. [Google Scholar] [CrossRef]
- Singh, R.; Singh, S.; Pandey, P.N. In Silico Analysis of Sirt2 from Schistosoma Mansoni: Structures, Conformations, and Interactions with Inhibitors. J. Biomol. Struct. Dyn. 2016, 34, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Huhtiniemi, T.; Salo, H.S.; Suuronen, T.; Poso, A.; Salminen, A.; Leppänen, J.; Jarho, E.; Lahtela-Kakkonen, M. Structure-Based Design of Pseudopeptidic Inhibitors for SIRT1 and SIRT2. J. Med. Chem. 2011, 54, 6456–6468. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Wei, W.; Jiang, Y.; Peng, H.; Cai, J.; Mao, C.; Dai, H.; Choy, W.; Bemis, J.E.; Jirousek, M.R.; et al. Crystal Structures of Human SIRT3 Displaying Substrate-Induced Conformational Changes. J. Biol. Chem. 2009, 284, 24394–24405. [Google Scholar] [CrossRef]
- Halgren, T. New Method for Fast and Accurate Binding-Site Identification and Analysis. Chem. Biol. Drug Des. 2007, 69, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Szczepankiewicz, B.G.; Dai, H.; Koppetsch, K.J.; Qian, D.; Jiang, F.; Mao, C.; Perni, R.B. Synthesis of Carba-NAD and the Structures of Its Ternary Complexes with SIRT3 and SIRT5. J. Org. Chem. 2012, 77, 7319–7329. [Google Scholar] [CrossRef]
- QM-Polarized Ligand Docking Protocol. Available online: http://gohom.win/ManualHom/Schrodinger/Schrodinger_2015-2_docs/qpld/qpld_user_manual.pdf (accessed on 29 July 2022).
- Zhou, Y.; Zhang, H.; He, B.; Du, J.; Lin, H.; Cerione, R.A.; Hao, Q. The Bicyclic Intermediate Structure Provides Insights into the Desuccinylation Mechanism of Human Sirtuin 5 (SIRT5). J. Biol. Chem. 2012, 287, 28307–28314. [Google Scholar] [CrossRef]
- Wang, H.-L.; Liu, S.; Yu, Z.-J.; Wu, C.; Cheng, L.; Wang, Y.; Chen, K.; Zhou, S.; Chen, Q.; Yu, Y.; et al. Interactions between Sirtuins and Fluorogenic Small-Molecule Substrates Offer Insights into Inhibitor Design. RSC Adv. 2017, 7, 36214–36222. [Google Scholar] [CrossRef]
- Pan, P.W.; Feldman, J.L.; Devries, M.K.; Dong, A.; Edwards, A.M.; Denu, J.M. Structure and Biochemical Functions of SIRT6. J. Biol. Chem. 2011, 286, 14575–14587. [Google Scholar] [CrossRef]
- del Rio, A.; Barbosa, A.J.M.; Caporuscio, F.; Mangiatordi, G.F. CoCoCo: A Free Suite of Multiconformational Chemical Databases for High-Throughput Virtual Screening Purposes. Mol. Biosyst. 2010, 6, 2122. [Google Scholar] [CrossRef]
- Home-Enamine. Available online: https://enamine.net/ (accessed on 13 May 2022).
- Mai, A.; Valente, S.; Meade, S.; Carafa, V.; Tardugno, M.; Nebbioso, A.; Galmozzi, A.; Mitro, N.; de Fabiani, E.; Altucci, L.; et al. Study of 1,4-Dihydropyridine Structural Scaffold: Discovery of Novel Sirtuin Activators and Inhibitors. J. Med. Chem. 2009, 52, 5496–5504. [Google Scholar] [CrossRef]
- Valente, S.; Mellini, P.; Spallotta, F.; Carafa, V.; Nebbioso, A.; Polletta, L.; Carnevale, I.; Saladini, S.; Trisciuoglio, D.; Gabellini, C.; et al. 1,4-Dihydropyridines Active on the SIRT1/AMPK Pathway Ameliorate Skin Repair and Mitochondrial Function and Exhibit Inhibition of Proliferation in Cancer Cells. J. Med. Chem. 2016, 59, 1471–1491. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Lu, S.; Huang, Z.; Liu, X.; Mou, L.; Luo, Y.; Zhao, Y.; Liu, Y.; Chen, Z.; Hou, T.; et al. Allosite: A Method for Predicting Allosteric Sites. Bioinformatics 2013, 29, 2357–2359. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, A.; Solanki, V.; Parkesh, R.; Thakur, K.G. Crystal Structure of the N-Terminal Domain of Human SIRT7 Reveals a Three-Helical Domain Architecture. Proteins Struct. Funct. Bioinform. 2016, 84, 1558–1563. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://Zhanggroup.Org/I-TASSER/ (accessed on 29 July 2022).
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein Structure and Function Prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. I-TASSER Server: New Development for Protein Structure and Function Predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, A.; Min, J.; Antoshenko, T.; Wang, C.L.; Allali-Hassani, A.; Dong, A.; Loppnau, P.; Vedadi, M.; Bochkarev, A.; Sternglanz, R.; et al. Structural Basis of Inhibition of the Human NAD+-Dependent Deacetylase SIRT5 by Suramin. Structure 2007, 15, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J.E.; Clark-Garvey, S.; Kalac, M.; Scotto, L.; Marchi, E.; Neylon, E.; Johannet, P.; Wei, Y.; Zain, J.; O’Connor, O.A. Sirtuin and Pan-Class I/II Deacetylase (DAC) Inhibition Is Synergistic in Preclinical Models and Clinical Studies of Lymphoma. Blood 2013, 122, 2104–2113. [Google Scholar] [CrossRef]
- Park, M.A.; Mitchell, C.; Zhang, G.; Yacoub, A.; Allegood, J.; Häussinger, D.; Reinehr, R.; Larner, A.; Spiegel, S.; Fisher, P.B.; et al. Vorinostat and Sorafenib Increase CD95 Activation in Gastrointestinal Tumor Cells through a Ca2+ De Novo Ceramide-PP2A-Reactive Oxygen Species–Dependent Signaling Pathway. Cancer Res. 2010, 70, 6313–6324. [Google Scholar] [CrossRef]
- Ntie-Kang, F.; Amoa Onguéné, P.; Fotso, G.W.; Andrae-Marobela, K.; Bezabih, M.; Ndom, J.C.; Ngadjui, B.T.; Ogundaini, A.O.; Abegaz, B.M.; Meva’a, L.M. Virtualizing the P-ANAPL Library: A Step towards Drug Discovery from African Medicinal Plants. PLoS ONE 2014, 9, e90655. [Google Scholar] [CrossRef]
- Baell, J.B.; Holloway, G.A. New Substructure Filters for Removal of Pan Assay Interference Compounds (PAINS) from Screening Libraries and for Their Exclusion in Bioassays. J. Med. Chem. 2010, 53, 2719–2740. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.-B.; Jing, H.; Aramsangtienchai, P.; He, B.; Khan, S.; Hu, J.; Lin, H.; Hao, Q. Efficient Demyristoylase Activity of SIRT2 Revealed by Kinetic and Structural Studies. Sci. Rep. 2015, 5, 8529. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Goto, Y.; Nishimasu, H.; Morimoto, J.; Ishitani, R.; Dohmae, N.; Takeda, N.; Nagai, R.; Komuro, I.; Suga, H.; et al. Structural Basis for Potent Inhibition of SIRT2 Deacetylase by a Macrocyclic Peptide Inducing Dynamic Structural Change. Structure 2014, 22, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, T.; Gerhardt, S.; Einsle, O.; Jung, M. Seeding for Sirtuins: Microseed Matrix Seeding to Obtain Crystals of Human Sirt3 and Sirt2 Suitable for Soaking. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 1498–1510. [Google Scholar] [CrossRef]
- Gertz, M.; Fischer, F.; Nguyen, G.T.T.; Lakshminarasimhan, M.; Schutkowski, M.; Weyand, M.; Steegborn, C. Ex-527 Inhibits Sirtuins by Exploiting Their Unique NAD-Dependent Deacetylation Mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, E2772–E2781. [Google Scholar] [CrossRef]
- You, W.; Steegborn, C. Structural Basis of Sirtuin 6 Inhibition by the Hydroxamate Trichostatin A: Implications for Protein Deacylase Drug Development. J. Med. Chem. 2018, 61, 10922–10928. [Google Scholar] [CrossRef]
- McGann, M. FRED and HYBRID Docking Performance on Standardized Datasets. J. Comput. Aided Mol. Des. 2012, 26, 897–906. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef]
- RCSB PDB-2B4Y: Crystal Structure of Human Sirtuin Homolog 5. Available online: https://www.rcsb.org/structure/2B4Y (accessed on 13 April 2022).
- Gaspar, L.; Coron, R.P.; KongThoo Lin, P.; Costa, D.M.; Perez-Cabezas, B.; Tavares, J.; Roura-Ferrer, M.; Ramos, I.; Ronin, C.; Major, L.L.; et al. Inhibitors of Trypanosoma Cruzi Sir2 Related Protein 1 as Potential Drugs against Chagas Disease. PLoS Negl. Trop. Dis. 2018, 12, e0006180. [Google Scholar] [CrossRef]
- Ghazy, E.; Abdelsalam, M.; Robaa, D.; Pierce, R.J.; Sippl, W. Histone Deacetylase (HDAC) Inhibitors for the Treatment of Schistosomiasis. Pharmaceuticals 2022, 15, 80. [Google Scholar] [CrossRef]
- Fioravanti, R.; Mautone, N.; Rovere, A.; Rotili, D.; Mai, A. Targeting Histone Acetylation/Deacetylation in Parasites: An Update (2017–2020). Curr. Opin. Chem. Biol. 2020, 57, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ouaissi, M.; Ouaissi, A. Histone Deacetylase Enzymes as Potential Drug Targets in Cancer and Parasitic Diseases. J. Biomed. Biotechnol. 2006, 2006, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lancelot, J.; Cabezas-Cruz, A.; Caby, S.; Marek, M.; Schultz, J.; Romier, C.; Sippl, W.; Jung, M.; Pierce, R.J. Schistosome Sirtuins as Drug Targets. Future Med. Chem. 2015, 7, 765–782. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W. Sirtuins as Emerging Anti-Parasitic Targets. Eur. J. Med. Chem. 2013, 59, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Religa, A.A.; Waters, A.P. Sirtuins of Parasitic Protozoa: In Search of Function(s). Mol. Biochem. Parasitol. 2012, 185, 71–88. [Google Scholar] [CrossRef]
- Ihlenfeldt, W.-D.; Voigt, J.H.; Bienfait, B.; Oellien, F.; Nicklaus, M.C. Enhanced CACTVS Browser of the Open NCI Database. J. Chem. Inf. Comput. Sci. 2002, 42, 46–57. [Google Scholar] [CrossRef]
- Kadam, R.U.; Kiran, V.M.; Roy, N. Comparative Protein Modeling and Surface Analysis of Leishmania Sirtuin: A Potential Target for Antileishmanial Drug Discovery. Bioorg. Med. Chem. Lett. 2006, 16, 6013–6018. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).