
Genome-Wide Identification and Characterization of G2-Like Transcription Factor Genes in Moso Bamboo (Phyllostachys edulis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Identification of PeGLK Genes
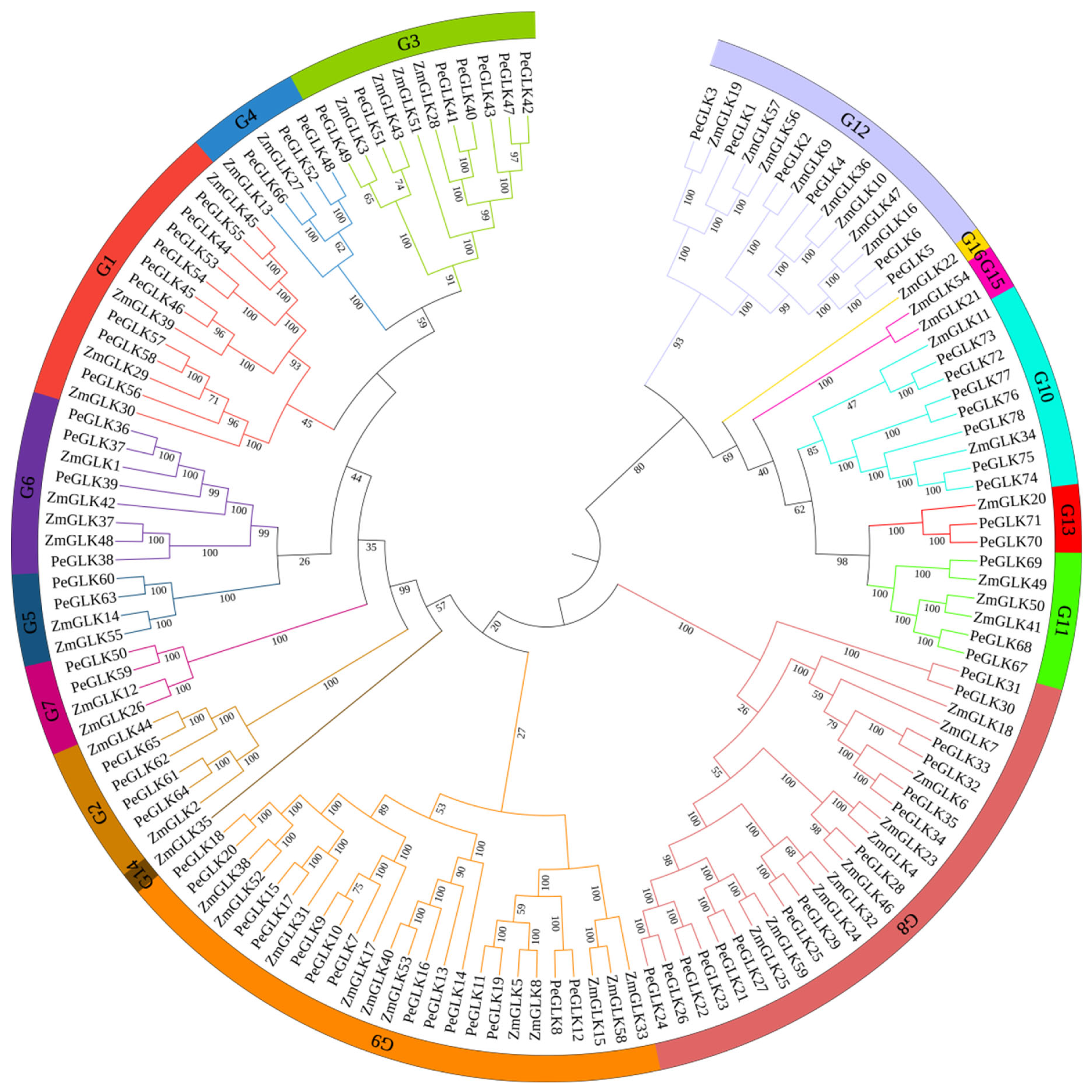
2.3. PeGLK Sequence Alignment and Phylogenetic Analysis
2.4. Gene and Protein Structure Analysis
2.5. Chromosomal Location and Synteny Analysis
2.6. Calculation of Ka/Ks Values
2.7. Putative Promoter Region Analysis of PeGLK Genes
2.8. PeGLK Expression Profiles, RNA Extraction, and qRT-PCR Analysis
3. Results
3.1. Identification of PeGLK Genes in Phyllostachys edulis
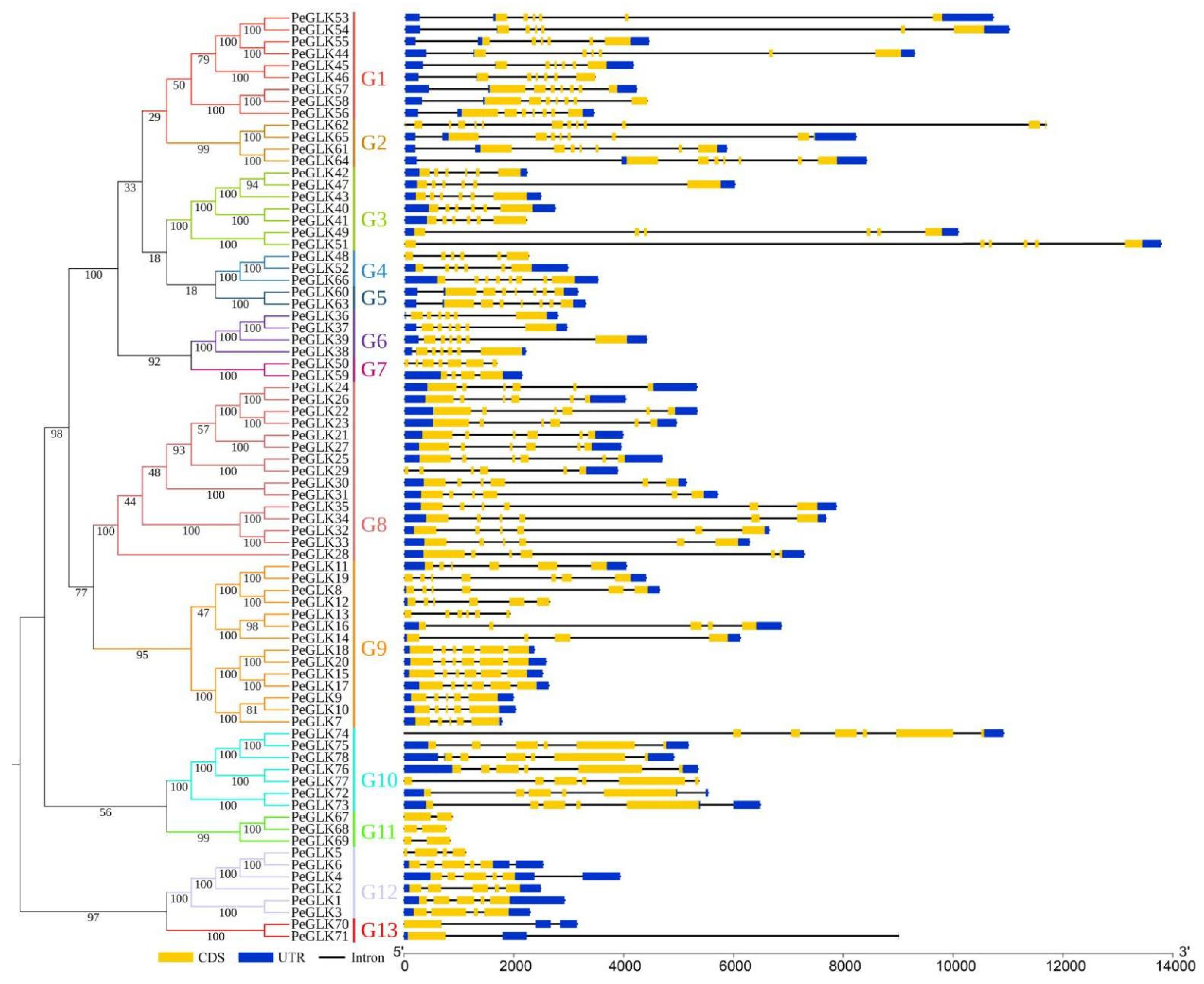
3.2. Phylogenetic and Exon-Intron Structure Analysis of GLK Genes
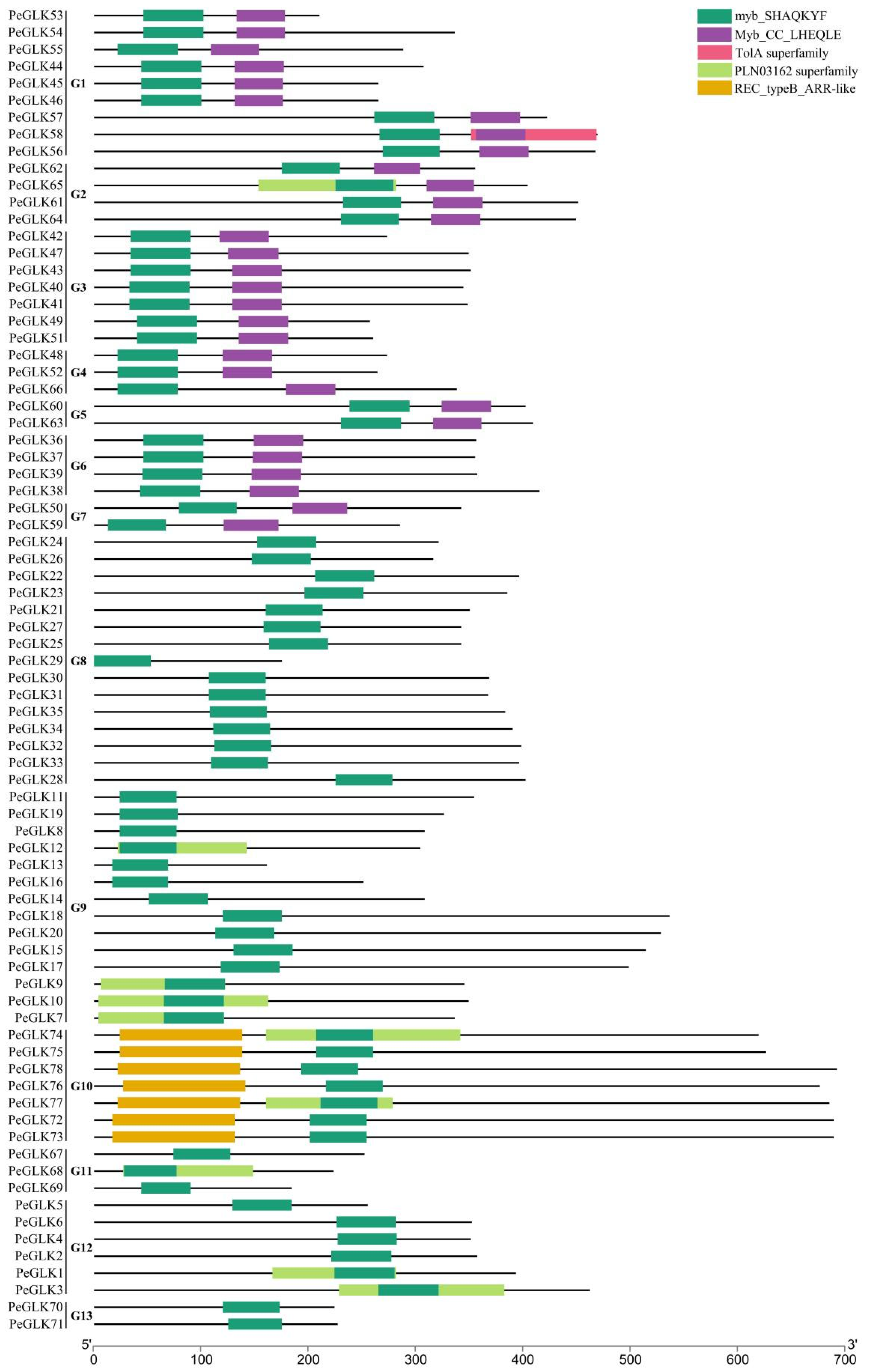
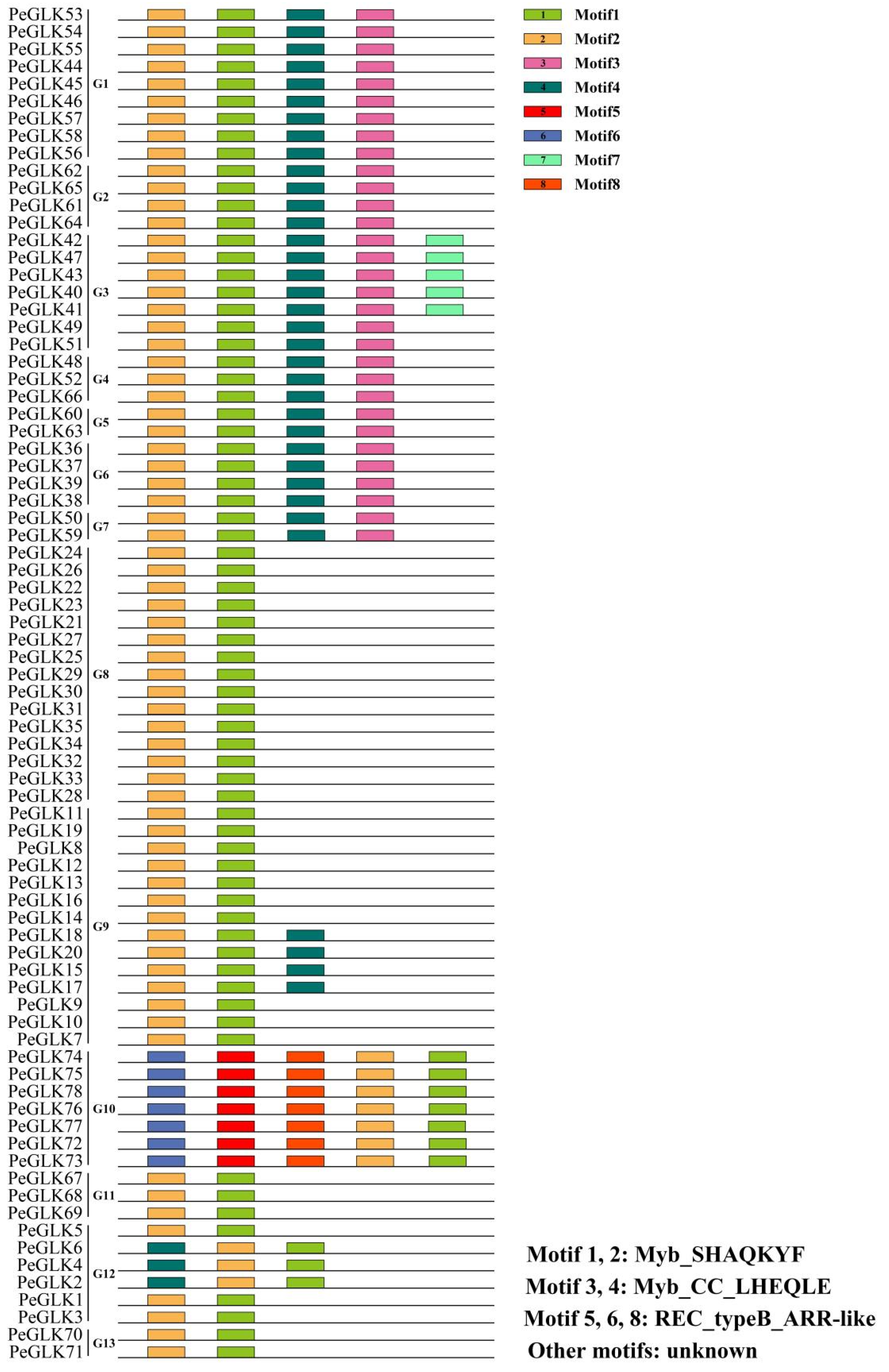
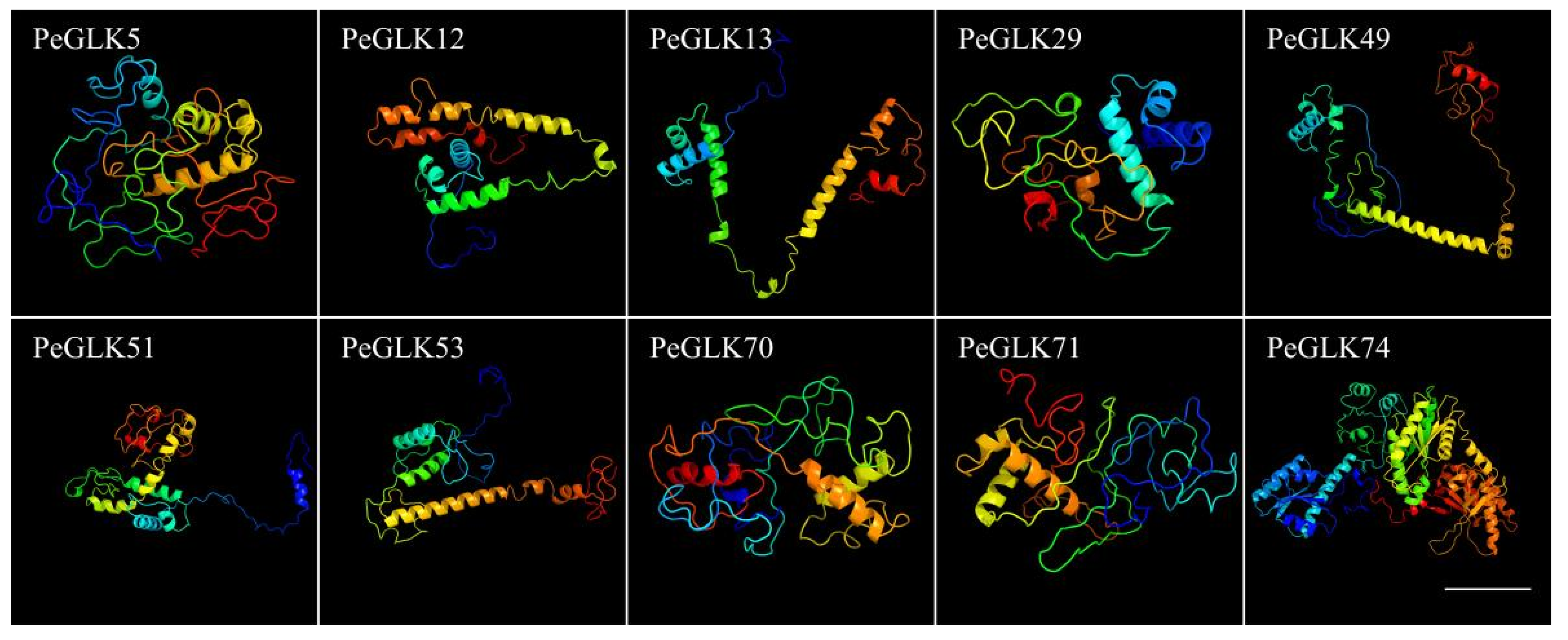
3.3. Identification of Conserved Sequence Motifs and Homology Modeling in PeGLK Genes
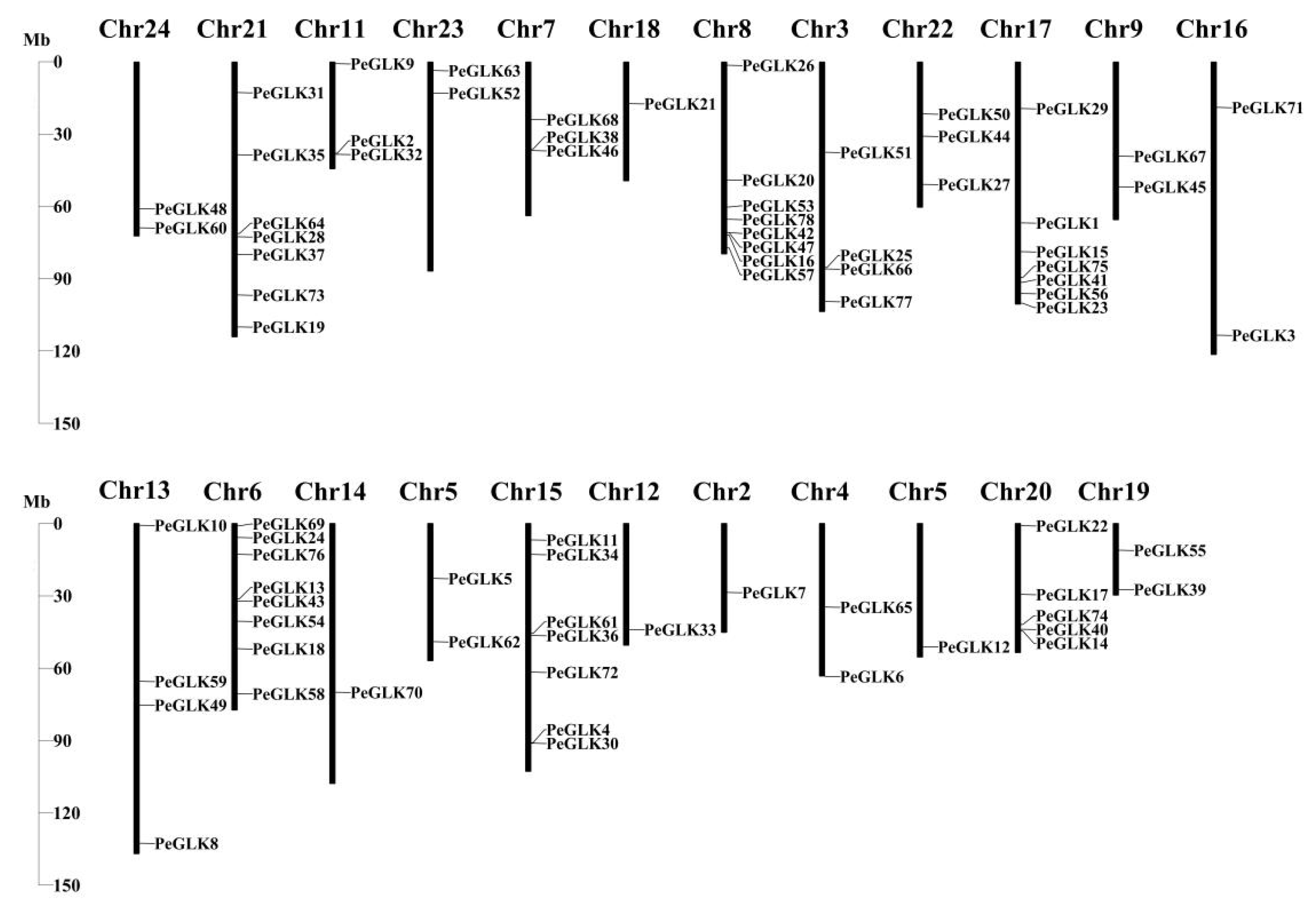
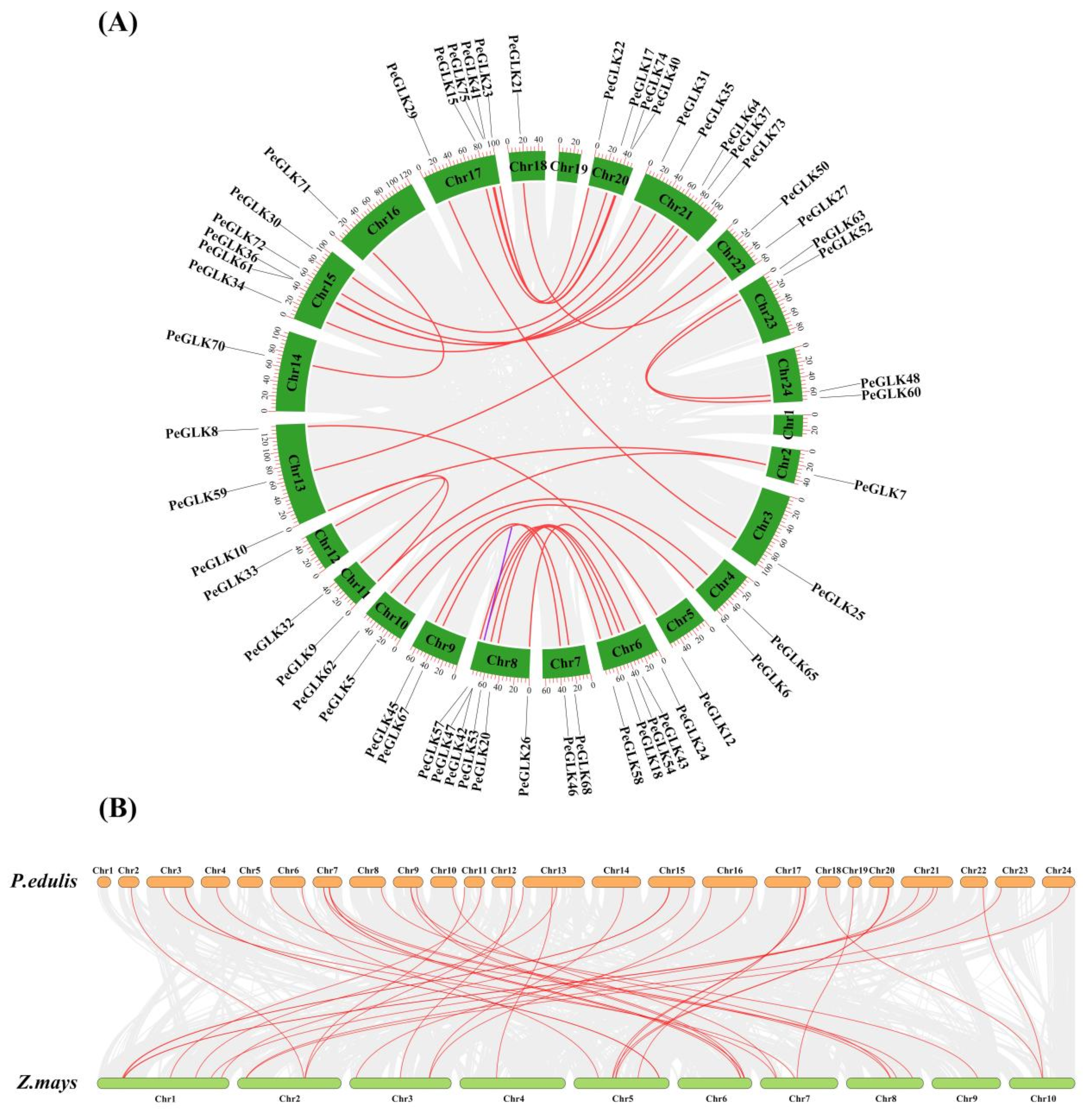
3.4. Physical Locations and Duplication Events of Pelf Genes in Phyllostachys edulis
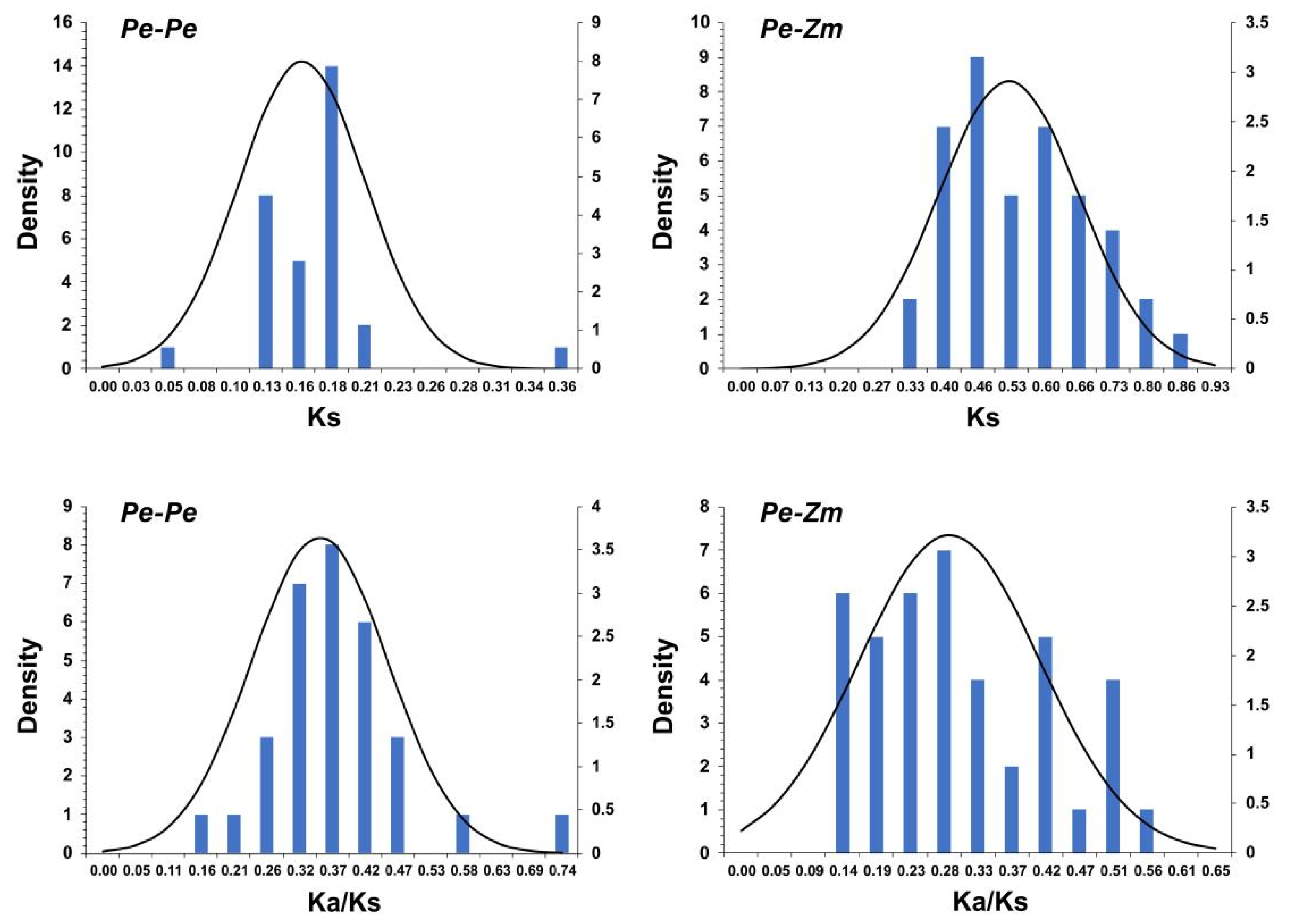
3.5. Evolutionary Patterns and Divergence Times of GLK Genes in Moso Bamboo and Maize
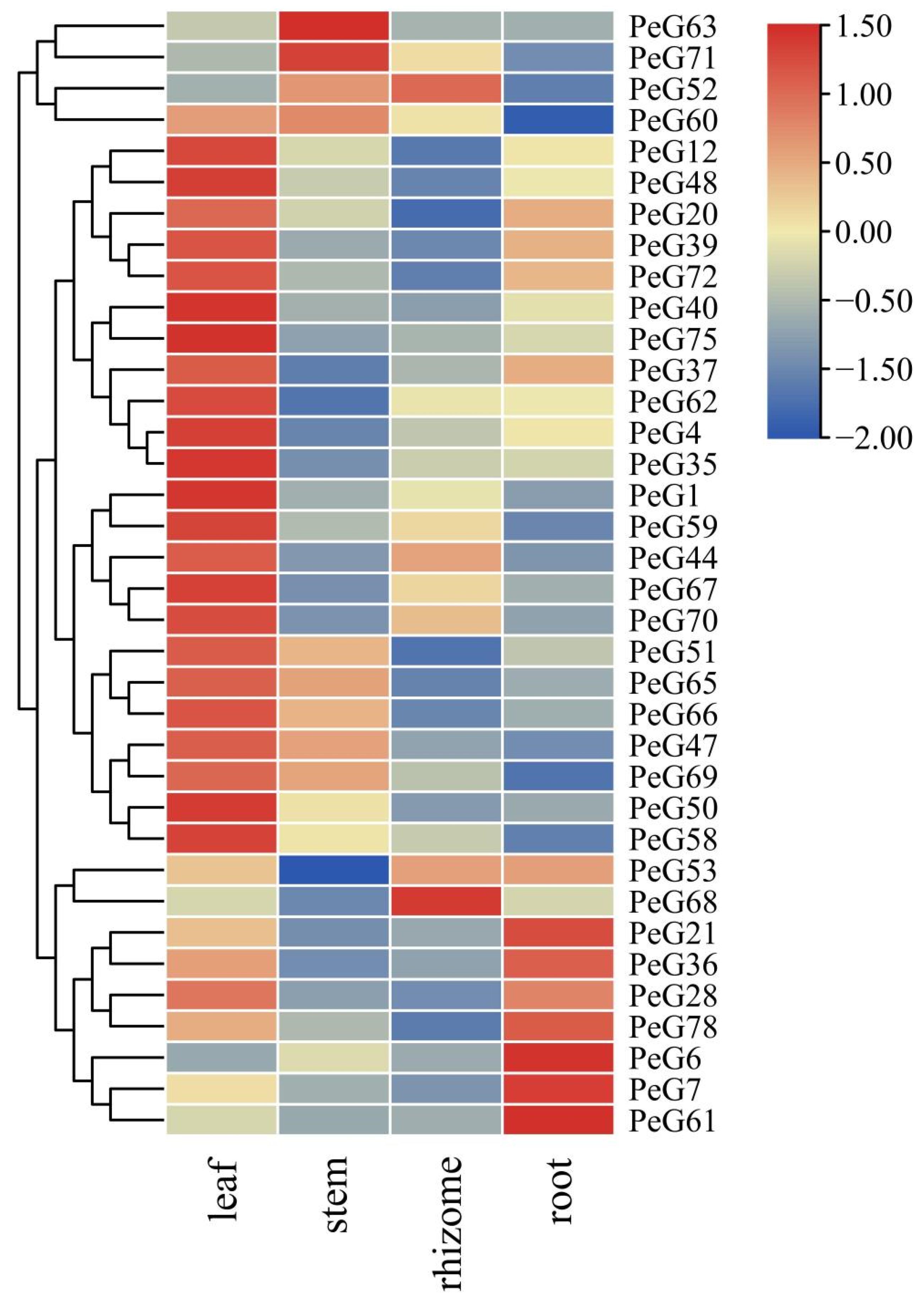
3.6. PeGLK Expression Levels in Different Tissues
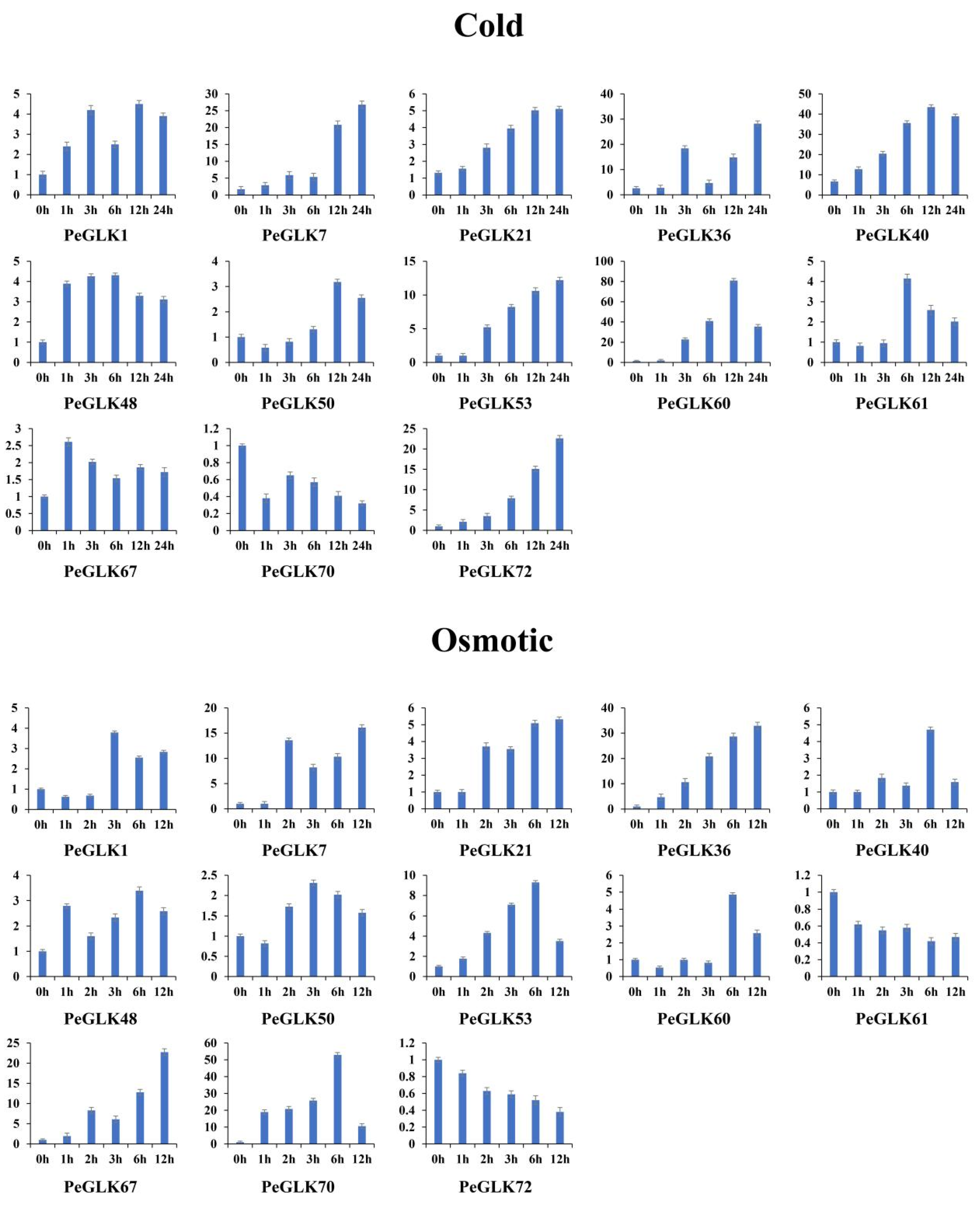
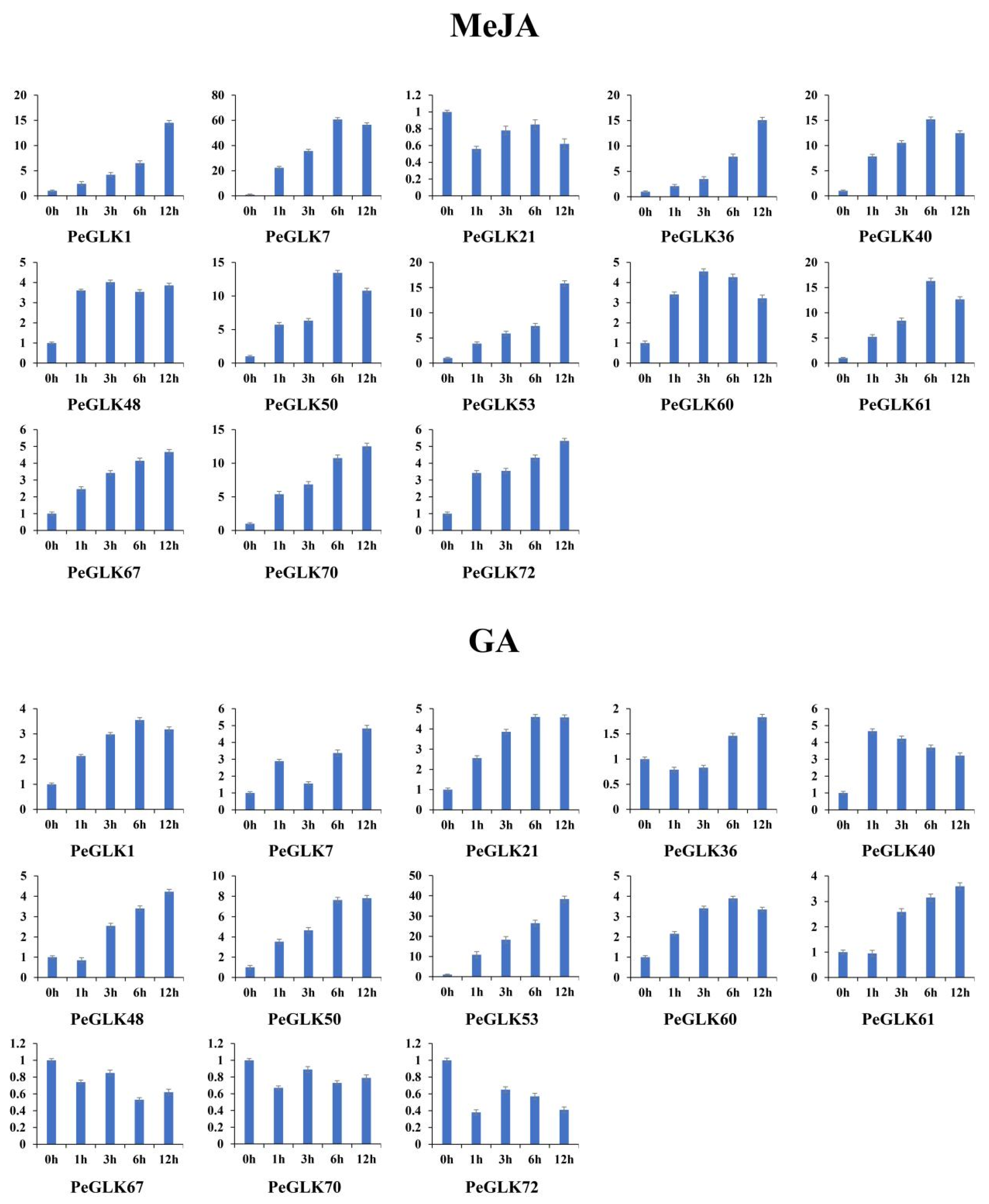
3.7. Expression Profiles of GLK Genes under Abiotic Stress and Phytohormone Treatment
4. Discussion
4.1. GLKs in Phyllostachys edulis
4.2. Physical Locations and Divergence of GLKs in Moso Bamboo and Maize
4.3. Potential Functions of PeGLKs in Moso Bamboo Development and Stress Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| CDD | conserved domain database |
| DBD | Myb DNA-binding domain |
| DRE | drought-responsive element |
| GA | gibberellic acid |
| GLK | Golden2-like |
| LTRE | cold-responsive element |
| MeJA | methyl jasmonate |
| MW | molecular weight |
| MYA | million years ago |
| PEG | polyethylene glycol 6000 |
| pI | isoelectric point |
| SA | silica acid |
| TIP41 | tonoplast intrinsic protein 41 |
References
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef]
- Jarvis, P.; López-Juez, E. Biogenesis and homeostasis of chloroplasts and other plastids. Nat. Rev. Mol. Cell Biol. 2013, 14, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Y.; Li, P.X.; Zou, L.J.; Tan, W.R.; Zheng, T.; Zhang, D.W. GOLDEN2-LIKE transcription factors coordinate the tolerance to Cucumber mosaic virus in Arabidopsis. Biochem. Biophys. Res. Commun. 2016, 477, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.F.S.; Mayor, R.; Kashef, J. Cadherin-11 mediates contact inhibition of locomotion during xenopus neural crest cell migration. PLoS ONE 2013, 8, e85717. [Google Scholar]
- Parente, D.J.; Swint-Kruse, L. Multiple co-evolutionary networks are supported by the common tertiary scaffold of the LacI/GalR proteins. PLoS ONE 2013, 8, e84398. [Google Scholar] [CrossRef]
- Waters, M.T.; Wang, P.; Korkaric, M.; Capper, R.G.; Saunders, N.J.; Langdale, J.A. GLK transcription factors coordinate expression of the photosynthetic apparatus in Arabidopsis. Plant Cell 2009, 21, 1109–1128. [Google Scholar] [CrossRef]
- Moreira, D.; Le Guyader, H.; Philippe, H. The origin of red algae and the evolution of chloroplasts. Nature 2000, 405, 69–72. [Google Scholar] [CrossRef]
- Lopez-Juez, E.; Pyke, K.A. Plastids unleashed: Their development and their integration in plant development. Int. J. Dev. Biol. 2005, 49, 557–577. [Google Scholar] [CrossRef]
- Park, J.; Werley, C.A.; Venkatachalam, V.; Kralj, J.M.; Dib-Hajj, S.D.; Waxman, S.G.; Cohen, A.E. Screening fluorescent voltage indicators with spontaneously spiking HEK cells. PLoS ONE 2013, 8, e85221. [Google Scholar] [CrossRef]
- Neuhaus, H.E.; Emes, M.J. Nonphotosynthetic metabsolism in plastid. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 111–140. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Imamura, A.; Hanaki, N.; Nakamura, A.; Suzuki, T.; Taniguchi, M.; Kiba, T.; Ueguchi, C.; Sugiyama, T.; Mizuno, T. Compilation and characterization of Arabiopsis thaliana response regulators implicated in His-Asp phosphorelay signal transduction. Plant Cell Physiol. 1999, 40, 733–742. [Google Scholar] [CrossRef]
- Hosoda, K.; Imamura, A.; Katoh, E.; Hatta, T.; Tachiki, M.; Yamada, H.; Mizuno, T.; Yamazaki, T. Molecular structure of the GARP family of plant Myb-related DNA binding motifs of the Arabidopsis response regulators. Plant Cell 2002, 14, 2015–2029. [Google Scholar] [CrossRef]
- Rossini, L.; Cribb, L.; Martin, D.J.; Langdale, J.A. The maize Golden2 gene defines a novel class of transcriptional regulators in plants. Plant Cell 2001, 13, 1231–1244. [Google Scholar] [CrossRef]
- Waters, M.T.; Moylan, E.C.; Langdale, J.A. GLK transcription factors regulate chloroplast development in a cell-autonomous manner. Plant J. 2008, 56, 432–444. [Google Scholar] [CrossRef]
- Xiao, Y.; You, S.; Kong, W.; Tang, Q.; Bai, W.; Cai, Y.; Zheng, H.; Wang, C.; Jiang, L.; Wang, C.; et al. A GARP transcription factor anther dehiscence defected 1 (OsADD1) regulates rice anther dehiscence. Plant Mol. Biol. 2019, 101, 403–414. [Google Scholar] [CrossRef]
- Fitter, D.W.; Martin, D.J.; Copley, M.J.; Scotland, R.W.; Langdale, J.A. GLK gene pairs regulate chloroplast development in diverse plant species. Plant J. 2002, 31, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Yasumura, Y.; Moylan, E.C.; Langdale, J.A. A conserved transcription factor mediates nuclear control of organelle biogenesis in anciently diverged land plants. Plant Cell 2005, 17, 1894–1907. [Google Scholar] [CrossRef]
- Murmu, J.; Wilton, M.; Allard, G.; Pandeya, R.; Desveaux, D.; Singh, J.; Subramaniam, R. Arabidopsis GOLDEN2-LIKE (GLK) transcription factors activate jasmonic acid (JA)-dependent disease susceptibility to the biotrophic pathogen Hyaloperonospora arabidopsidis, as well as JA-independent plant immunity against the necrotrophic pathogen Botrytis cinerea. Mol. Plant Pathol. 2013, 15, 174–184. [Google Scholar]
- Powell, A.L.T.; Nguyen, C.V.; Hill, T.; Cheng, K.L.; Figueroa-Balderas, R.; Aktas, H.; Ashrafi, H.; Pons, C.; Fernández-Muñoz, R.; Vicente, A.; et al. Uniform ripening encodes a Golden 2-like transcription factor regulating tomato fruit chloroplast development. Science 2012, 336, 1711–1715. [Google Scholar] [CrossRef]
- Hall, L.N.; Rossini, L.; Cribb, L.; Langdale, J.A. GOLDEN 2: A novel transcriptional regulator of cellular differentiation in the maize Leaf. Plant Cell 1998, 10, 925–936. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Han, G.; Zhou, L.; Ali, A.; Zhu, S.; Li, X. Molecular evolution and genetic variation of G2-like transcription factor genes in maize. PLoS ONE 2016, 11, e0161763. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhao, S.; Liu, J.; Zhao, H.; Sun, X.; Wu, T.; Pei, T.; Wang, Y.; Liu, Q.; Yang, H.; et al. Genome-wide identification of tomato Golden 2-like transcription factors and abiotic stress related members screening. BMC Plant Biol. 2022, 22, 82. [Google Scholar] [CrossRef]
- Qin, M.; Zhang, B.; Gu, G.; Yuan, J.; Yang, X.; Yang, J.; Xie, X. Genome-wide analysis of the G2-like transcription factor genes and their expression in different senescence stages of tobacco (Nicotiana Tabacum L.). Front. Genet. 2021, 12, 787. [Google Scholar] [CrossRef]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, e11. [Google Scholar] [CrossRef]
- Cannon, S.; Mitra, A.; Baumgarten, A.; Young, N.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 10, 4. [Google Scholar]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Lugli, G.A.; Tarracchini, C.; Alessandri, G.; Milani, C.; Mancabelli, L.; Turroni, F.; Neuzil-Bunesova, V.; Ruiz, L.; Margolles, A.; Ventura, M. Decoding the genomic variability among members of the Bifidobacterium dentium species. Microorganisms 2020, 8, 1720. [Google Scholar] [CrossRef]
- Carlini, D.; Stephan, W. In vivo introduction of unpreferred synonymous codons into the drosophila Adh gene results in reduced levels of ADH protein. Genetics 2003, 163, 239–243. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Rozas, J. DNA sequence polymorphism analysis using DnaSP. Methods Mol. Biol. 2009, 537, 337–350. [Google Scholar]
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K.; et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- He, Y.; Mao, S.; Gao, Y.; Zhu, L.; Wu, D.; Cui, Y.; Li, J.; Qian, W. Genome-wide identification and expression analysis of WRKY transcription factors under multiple stresses in Brassica napus. PLoS ONE 2016, 11, e0157558. [Google Scholar] [CrossRef]
- Wu, S.; Wu, M.; Dong, Q.; Jiang, H.; Cai, R.; Xiang, Y. Genome-wide identification, classification and expression analysis of the PHD-finger protein family in Populus trichocarpa. Gene 2016, 575, 75–89. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Fan, C.; Ma, J.; Guo, Q.; Li, X.; Wang, H.; Lu, M. Selection of reference genes for quantitative real-time PCR in bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e56573. [Google Scholar]
- Liu, H.L.; Wu, M.; Li, F.; Gao, Y.M.; Chen, F.; Xiang, Y. TCP transcription factors in moso bamboo (Phyllostachys edulis): Genome-wide identification and expression analysis. Front. Plant Sci. 2018, 9, 1263. [Google Scholar] [CrossRef]
- Soding, J. Protein homology detection by HMM-HMM comparison. Bioinformatics 2004, 21, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.H.; Karlowski, W.M.; Pan, R.; Tzeng, Y.H.; Mayer, K.F.X.; Li, W.H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, R.S.; Lin, H.N.; Childs, K.L.; Hansey, C.N.; Buell, C.R. Genome-wide atlas of transcription during maize development. Plant J. 2011, 66, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, H. Chloroplast ultrastructure in plants. New Phytol. 2019, 223, 565–574. [Google Scholar] [CrossRef] [PubMed]
- La Marca, M.; Beffy, P.; Pugliese, A.; Longo, V. Fermented wheat powder induces the antioxidant and detoxifying system in primary rat hepatocytes. PLoS ONE 2013, 8, e83538. [Google Scholar] [CrossRef]
- Opanowicz, M.; Vain, P.; Draper, J.; Parker, D.; Doonan, J.H. Brachypodiumdistachyon: Making hay with a wild grass. Trends Plant Sci. 2008, 13, 172–177. [Google Scholar] [CrossRef]
- Gu, Z.; Steinmetz, L.M.; Gu, X.; Scharfe, C.; Davis, R.W.; Li, W.H. Role of duplicate genes in genetic robustness against null mutations. Nature 2003, 421, 63–66. [Google Scholar] [CrossRef]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Nasim, J.; Malviya, N.; Kumar, R.; Yadav, D. Genome-wide bioinformatics analysis of dof transcription factor gene family of chickpea and its comparative phylogenetic assessment with Arabidopsis and Rice. Plant Syst. Evol. 2016, 302, 1009–1026. [Google Scholar] [CrossRef]
- Song, H.; Wang, P.; Lin, J.Y.; Zhao, C.; Bi, Y.; Wang, X. Genome-wide identification and characterization of WRKY gene family in peanut. Front. Plant Sci. 2016, 7, 9. [Google Scholar] [CrossRef]
- Wu, M.; Li, Y.; Chen, D.; Liu, H.; Zhu, D.; Xiang, Y. Genome-wide identification and expression analysis of the IQD gene family in moso bamboo (Phyllostachys edulis). Sci. Rep. 2016, 6, 24520. [Google Scholar] [CrossRef]
- Ahuja, I.; de Vos, R.C.H.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef]
- Amerik, A.Y.; Hochstrasser, M. Mechanism and function of deubiquitinating enzymes. BBA-Mol. Cell Res. 2004, 1695, 189–207. [Google Scholar] [CrossRef] [Green Version]
- March, E.; Farrona, S. Plant deubiquitinases and their role in the control of gene expression through modification of histones. Front. Plant Sci. 2018, 8, 02274. [Google Scholar] [CrossRef]
- Gabaldón, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef]
- Nakamura, H.; Muramatsu, M.; Hakata, M.; Ueno, O.; Nagamura, Y.; Hirochika, H.; Takano, M.; Ichikawa, H. Ectopic overexpression of the transcription factor OsGLK1 induces chloroplast development in non-green rice cells. Plant Cell Physiol. 2009, 50, 1933–1949. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequences ID | Position | MW (Da) | PI | CDS Length (bp) | Size (aa) | Exons |
|---|---|---|---|---|---|---|---|
| PeGLK1 | PH02Gene35128.t1 | 17:66858322-66861246(+) | 42,593.43 | 8.19 | 1182 | 393 | 5 |
| PeGLK2 | PH02Gene01193.t1 | 11:37933106-37935593(+) | 38,720.28 | 7.10 | 1074 | 357 | 5 |
| PeGLK3 | PH02Gene09030.t2 | 16:113462311-113464603(−) | 49,571.46 | 6.08 | 1389 | 462 | 4 |
| PeGLK4 | PH02Gene21002.t1 | 15:91066421-91070352(+) | 38,096.01 | 6.99 | 1056 | 351 | 5 |
| PeGLK5 | PH02Gene35342.t1 | 10:22940475-22941589(+) | 27,158.16 | 8.77 | 768 | 255 | 4 |
| PeGLK6 | PH02Gene25255.t1 | 4:63579741-63582272(−) | 37,844.34 | 5.96 | 1059 | 352 | 5 |
| PeGLK7 | PH02Gene17491.t1 | 2:28739813-28741586(−) | 36,728.56 | 6.79 | 1011 | 336 | 5 |
| PeGLK8 | PH02Gene38026.t1 | 13:132810438-132815080(−) | 34,286.6 | 8.62 | 927 | 308 | 6 |
| PeGLK9 | PH02Gene18924.t1 | 11:802821-804806(−) | 37,872.77 | 6.35 | 1038 | 345 | 5 |
| PeGLK10 | PH02Gene39352.t1 | 13:873597-875620(−) | 38,491.68 | 6.98 | 1050 | 349 | 5 |
| PeGLK11 | PH02Gene20517.t1 | 15:6935099-6939136(−) | 37,931.95 | 8.00 | 1065 | 354 | 6 |
| PeGLK12 | PH02Gene29515.t1 | 5:51264001-51266636(−) | 33,988.33 | 8.93 | 915 | 304 | 6 |
| PeGLK13 | PH02Gene34333.t1 | 6:31369720-31371637(−) | 18,081.66 | 10.43 | 486 | 161 | 6 |
| PeGLK14 | PH02Gene30127.t1 | 20:44761394-44767510(−) | 33,657.58 | 7.08 | 927 | 308 | 4 |
| PeGLK15 | PH02Gene48395.t1 | 17:78812464-78814980(−) | 56,840.06 | 6.53 | 1545 | 514 | 6 |
| PeGLK16 | PH02Gene17440.t1 | 8:71935937-71942802(+) | 28,099.34 | 8.37 | 756 | 251 | 5 |
| PeGLK17 | PH02Gene38152.t1 | 20:29572044-29574673(−) | 54,525.46 | 6.22 | 1497 | 498 | 6 |
| PeGLK18 | PH02Gene39325.t1 | 6:52094000-52096363(+) | 58,022.99 | 6.46 | 1611 | 536 | 6 |
| PeGLK19 | PH02Gene00846.t1 | 21:110026992-110031388(+) | 35059.67 | 8.95 | 981 | 326 | 7 |
| PeGLK20 | PH02Gene42607.t1 | 8:49040798-49043378(−) | 57,616.54 | 5.94 | 1587 | 528 | 6 |
| PeGLK21 | PH02Gene02579.t2 | 18:17380009-17383980(+) | 36,881.46 | 7.37 | 1053 | 350 | 6 |
| PeGLK22 | PH02Gene39580.t2 | 20:1052803-1058126(−) | 43,052.21 | 9.02 | 1191 | 396 | 6 |
| PeGLK23 | PH02Gene31338.t2 | 17:100218030-100222977(+) | 41,530.78 | 9.40 | 1158 | 385 | 6 |
| PeGLK24 | PH02Gene17108.t1 | 6:5944617-5949931(+) | 34,923.39 | 9.76 | 966 | 321 | 6 |
| PeGLK25 | PH02Gene03796.t1 | 3:85369708-85374396(+) | 36,685.24 | 7.94 | 1029 | 342 | 6 |
| PeGLK26 | PH02Gene29053.t1 | 8:1462771-1466790(+) | 34,689.98 | 9.70 | 951 | 316 | 6 |
| PeGLK27 | PH02Gene49306.t1 | 22:50874099-50878039(+) | 36,221.83 | 7.77 | 1029 | 342 | 6 |
| PeGLK28 | PH02Gene34821.t1 | 21:72609406-72616684(+) | 41,826.53 | 9.19 | 1209 | 402 | 6 |
| PeGLK29 | PH02Gene32674.t2 | 17:19419163-19423040(−) | 19,477.62 | 5.95 | 528 | 175 | 6 |
| PeGLK30 | PH02Gene20991.t1 | 15:91308193-91313320(+) | 38,773.69 | 6.27 | 1107 | 368 | 6 |
| PeGLK31 | PH02Gene17776.t1 | 21:12781867-12787567(−) | 38,324.37 | 6.26 | 1104 | 367 | 6 |
| PeGLK32 | PH02Gene01179.t1 | 11:38405451-38412089(+) | 42,846.77 | 7.14 | 1197 | 398 | 6 |
| PeGLK33 | PH02Gene15844.t1 | 12:44109361-44115643(+) | 42,734.24 | 7.12 | 1191 | 396 | 6 |
| PeGLK34 | PH02Gene20627.t1 | 15:12983299-12990970(−) | 42,273.57 | 9.54 | 1173 | 390 | 6 |
| PeGLK35 | PH02Gene45340.t1 | 21:38563232-38571091(−) | 41,493.76 | 9.56 | 1152 | 383 | 6 |
| PeGLK36 | PH02Gene46403.t1 | 15:46604970-46607753(−) | 39,682.03 | 8.17 | 1071 | 356 | 6 |
| PeGLK37 | PH02Gene15930.t1 | 21:79933476-79936426(+) | 39,576.87 | 8.17 | 1068 | 355 | 6 |
| PeGLK38 | PH02Gene02408.t1 | 7:36153335-36155537(−) | 46,811.37 | 6.66 | 1248 | 415 | 6 |
| PeGLK39 | PH02Gene44186.t1 | 19:27506582-27510983(−) | 39,621.3 | 8.60 | 1074 | 357 | 6 |
| PeGLK40 | PH02Gene39987.t1 | 20:44024199-44026929(+) | 37,799.07 | 8.54 | 1035 | 344 | 6 |
| PeGLK41 | PH02Gene09101.t1 | 17:91509553-91511761(+) | 38,369.88 | 9.12 | 1047 | 348 | 6 |
| PeGLK42 | PH02Gene31703.t1 | 8:70863226-70865442(+) | 30,479.81 | 8.90 | 822 | 273 | 6 |
| PeGLK43 | PH02Gene33296.t1 | 6:32324747-32327222(−) | 38,932.24 | 6.84 | 1056 | 351 | 6 |
| PeGLK44 | PH02Gene40793.t1 | 22:30906449-30915724(−) | 33,752.45 | 5.95 | 924 | 307 | 6 |
| PeGLK45 | PH02Gene05591.t1 | 9:51866782-51870935(−) | 29,354.19 | 6.67 | 798 | 265 | 6 |
| PeGLK46 | PH02Gene02376.t1 | 7:36864743-36868199(−) | 29,442.24 | 7.10 | 798 | 265 | 6 |
| PeGLK47 | PH02Gene31704.t1 | 8:70867912-70873914(+) | 38,717.97 | 6.90 | 1050 | 349 | 6 |
| PeGLK48 | PH02Gene10199.t1 | 24:60967409-60969661(+) | 30,026.13 | 8.33 | 822 | 273 | 6 |
| PeGLK49 | PH02Gene09178.t1 | 13:75454095-75464169(−) | 28,140.14 | 7.78 | 774 | 257 | 6 |
| PeGLK50 | PH02Gene04584.t1 | 22:21670037-21671709(−) | 37,153.86 | 6.76 | 1029 | 342 | 7 |
| PeGLK51 | PH02Gene45566.t1 | 3:37548923-37570928(−) | 28,160.04 | 8.90 | 783 | 260 | 6 |
| PeGLK52 | PH02Gene02264.t1 | 23:13009053-13012018(−) | 29,036.04 | 9.10 | 795 | 264 | 6 |
| PeGLK53 | PH02Gene13954.t1 | 8:60262961-60273665(+) | 23,380.35 | 6.60 | 633 | 210 | 6 |
| PeGLK54 | PH02Gene40923.t1 | 6:40681881-40692878(−) | 36,241.28 | 6.60 | 1011 | 336 | 6 |
| PeGLK55 | PH02Gene42198.t1 | 19:11230444-11234879(−) | 31,628.95 | 6.02 | 867 | 288 | 6 |
| PeGLK56 | PH02Gene29378.t1 | 17:96169688-96173122(+) | 51,782.72 | 6.71 | 1404 | 467 | 7 |
| PeGLK57 | PH02Gene02799.t1 | 8:77249489-77253699(+) | 46,249.31 | 5.51 | 1269 | 422 | 7 |
| PeGLK58 | PH02Gene29763.t1 | 6:70768706-70773109(+) | 51,370.07 | 5.75 | 1410 | 469 | 7 |
| PeGLK59 | PH02Gene45947.t2 | 13:65634449-65636584(−) | 31,526.73 | 5.57 | 858 | 285 | 4 |
| PeGLK60 | PH02Gene00387.t1 | 24:68886977-68890124(−) | 44,369.49 | 6.73 | 1209 | 402 | 7 |
| PeGLK61 | PH02Gene19293.t1 | 15:45733490-45739347(−) | 49,605.87 | 5.85 | 1356 | 451 | 7 |
| PeGLK62 | PH02Gene11518.t1 | 10:49175750-49187421(+) | 38,777.43 | 5.61 | 1068 | 355 | 13 |
| PeGLK63 | PH02Gene20571.t1 | 23:3653518-3656801(+) | 45,464.34 | 6.74 | 1230 | 409 | 7 |
| PeGLK64 | PH02Gene12389.t1 | 21:71256608-71265008(−) | 49,101.24 | 5.98 | 1350 | 449 | 7 |
| PeGLK65 | PH02Gene36110.t1 | 4:34815482-34823691(−) | 43,616.22 | 5.02 | 1215 | 404 | 8 |
| PeGLK66 | PH02Gene03773.t2 | 3:85969044-85972558(+) | 38,144.82 | 9.58 | 1017 | 338 | 8 |
| PeGLK67 | PH02Gene21337.t1 | 9:39183574-39184446(−) | 24,896.89 | 9.51 | 759 | 252 | 2 |
| PeGLK68 | PH02Gene22746.t1 | 7:23963609-23964367(−) | 22,731.56 | 9.04 | 672 | 223 | 2 |
| PeGLK69 | PH02Gene10071.t1 | 6:1102829-1103662(+) | 20,032.6 | 6.06 | 555 | 184 | 2 |
| PeGLK70 | PH02Gene11430.t1 | 14:70153233-70156386(+) | 23,995.12 | 6.61 | 675 | 224 | 1 |
| PeGLK71 | PH02Gene20674.t1 | 16:18948494-18957497(−) | 24,222.27 | 6.25 | 684 | 227 | 1 |
| PeGLK72 | PH02Gene11768.t1 | 15:61838091-61843627(−) | 74,154.2 | 5.79 | 2070 | 689 | 5 |
| PeGLK73 | PH02Gene26606.t1 | 21:96791235-96797716(−) | 74,480.85 | 5.86 | 2070 | 689 | 5 |
| PeGLK74 | PH02Gene30734.t1 | 20:42027130-42038041(+) | 67,709.51 | 6.06 | 1860 | 619 | 6 |
| PeGLK75 | PH02Gene04797.t1 | 17:89520790-89525967(+) | 68,432.23 | 6.11 | 1881 | 626 | 6 |
| PeGLK76 | PH02Gene11687.t1 | 6:12955225-12960572(+) | 74,366.97 | 5.98 | 2031 | 676 | 6 |
| PeGLK77 | PH02Gene21205.t1 | 3:99493453-99498815(−) | 74,410.85 | 6.04 | 2058 | 685 | 6 |
| PeGLK78 | PH02Gene33414.t1 | 8:65390201-65395109(+) | 74,995.96 | 5.59 | 2079 | 692 | 6 |
| Paralogous | Ka | Ks | Ka/Ks | Selection Pressure | Duplicate Type |
|---|---|---|---|---|---|
| PeGLK5/6 | 0.04 | 0.12 | 0.31 | Purifying selection | Segmental duplication |
| PeGLK7/10 | 0.07 | 0.19 | 0.37 | Purifying selection | Segmental duplication |
| PeGLK7/9 | 0.08 | 0.18 | 0.46 | Purifying selection | Segmental duplication |
| PeGLK9/10 | 0.07 | 0.17 | 0.39 | Purifying selection | Segmental duplication |
| PeGLK8/12 | 0.05 | 0.12 | 0.45 | Purifying selection | Segmental duplication |
| PeGLK15/17 | 0.09 | 0.12 | 0.72 | Purifying selection | Segmental duplication |
| PeGLK18/20 | 0.08 | 0.20 | 0.39 | Purifying selection | Segmental duplication |
| PeGLK21/27 | 0.06 | 0.18 | 0.30 | Purifying selection | Segmental duplication |
| PeGLK22/23 | 0.04 | 0.12 | 0.33 | Purifying selection | Segmental duplication |
| PeGLK24/26 | 0.04 | 0.18 | 0.25 | Purifying selection | Segmental duplication |
| PeGLK25/29 | 0.03 | 0.18 | 0.14 | Purifying selection | Segmental duplication |
| PeGLK30/31 | 0.05 | 0.14 | 0.35 | Purifying selection | Segmental duplication |
| PeGLK32/33 | 0.06 | 0.17 | 0.37 | Purifying selection | Segmental duplication |
| PeGLK34/35 | 0.04 | 0.12 | 0.36 | Purifying selection | Segmental duplication |
| PeGLK36/37 | 0.03 | 0.18 | 0.19 | Purifying selection | Segmental duplication |
| PeGLK40/41 | 0.03 | 0.13 | 0.24 | Purifying selection | Segmental duplication |
| PeGLK42/43 | 0.05 | 0.18 | 0.27 | Purifying selection | Segmental duplication |
| PeGLK43/47 | 0.05 | 0.17 | 0.28 | Purifying selection | Segmental duplication |
| PeGLK45/46 | 0.04 | 0.12 | 0.31 | Purifying selection | Segmental duplication |
| PeGLK48/52 | 0.05 | 0.17 | 0.30 | Purifying selection | Segmental duplication |
| PeGLK50/59 | 0.06 | 0.17 | 0.34 | Purifying selection | Segmental duplication |
| PeGLK53/54 | 0.05 | 0.14 | 0.33 | Purifying selection | Segmental duplication |
| PeGLK57/58 | 0.06 | 0.17 | 0.34 | Purifying selection | Segmental duplication |
| PeGLK60/63 | 0.06 | 0.18 | 0.34 | Purifying selection | Segmental duplication |
| PeGLK61/64 | 0.05 | 0.13 | 0.40 | Purifying selection | Segmental duplication |
| PeGLK62/65 | 0.20 | 0.36 | 0.57 | Purifying selection | Segmental duplication |
| PeGLK67/68 | 0.03 | 0.12 | 0.24 | Purifying selection | Segmental duplication |
| PeGLK70/72 | 0.08 | 0.18 | 0.46 | Purifying selection | Segmental duplication |
| PeGLK71/73 | 0.04 | 0.12 | 0.33 | Purifying selection | Segmental duplication |
| PeGLK74/75 | 0.04 | 0.13 | 0.27 | Purifying selection | Segmental duplication |
| PeGLK42/47 | 0.02 | 0.05 | 0.40 | Purifying selection | Tandem duplication |
| Paralogous | Ka | Ks | Ka/Ks | Selection Pressure |
|---|---|---|---|---|
| ZmGLK27/PeGLK66 | 0.09 | 0.79 | 0.11 | Purifying selection |
| ZmGLK48/PeGLK38 | 0.10 | 0.56 | 0.17 | Purifying selection |
| ZmGLK9/PeGLK2 | 0.14 | 0.59 | 0.23 | Purifying selection |
| ZmGLK2/PeGLK61 | 0.16 | 0.41 | 0.38 | Purifying selection |
| ZmGLK2/PeGLK64 | 0.17 | 0.42 | 0.40 | Purifying selection |
| ZmGLK1/PeGLK36 | 0.08 | 0.65 | 0.13 | Purifying selection |
| ZmGLK1/PeGLK37 | 0.08 | 0.71 | 0.12 | Purifying selection |
| ZmGLK6/PeGLK35 | 0.25 | 0.69 | 0.36 | Purifying selection |
| ZmGLK28/PeGLK41 | 0.07 | 0.38 | 0.18 | Purifying selection |
| ZmGLK28/PeGLK40 | 0.07 | 0.38 | 0.18 | Purifying selection |
| ZmGLK34/PeGLK75 | 0.14 | 0.59 | 0.24 | Purifying selection |
| ZmGLK34/PeGLK74 | 0.15 | 0.64 | 0.23 | Purifying selection |
| ZmGLK32/PeGLK25 | 0.14 | 0.57 | 0.25 | Purifying selection |
| ZmGLK43/PeGLK51 | 0.08 | 0.53 | 0.14 | Purifying selection |
| ZmGLK49/PeGLK69 | 0.13 | 0.49 | 0.27 | Purifying selection |
| ZmGLK20/PeGLK70 | 0.27 | 0.50 | 0.53 | Purifying selection |
| ZmGLK20/PeGLK71 | 0.23 | 0.45 | 0.51 | Purifying selection |
| ZmGLK51/PeGLK42 | 0.14 | 0.42 | 0.32 | Purifying selection |
| ZmGLK29/PeGLK58 | 0.11 | 0.55 | 0.19 | Purifying selection |
| ZmGLK3/PeGLK49 | 0.11 | 0.41 | 0.26 | Purifying selection |
| ZmGLK39/PeGLK46 | 0.06 | 0.50 | 0.13 | Purifying selection |
| ZmGLK39/PeGLK45 | 0.06 | 0.50 | 0.12 | Purifying selection |
| ZmGLK59/PeGLK21 | 0.12 | 0.55 | 0.21 | Purifying selection |
| ZmGLK59/PeGLK27 | 0.12 | 0.42 | 0.28 | Purifying selection |
| ZmGLK10/PeGLK4 | 0.17 | 0.60 | 0.28 | Purifying selection |
| ZmGLK13/PeGLK52 | 0.16 | 0.41 | 0.40 | Purifying selection |
| ZmGLK13/PeGLK48 | 0.16 | 0.46 | 0.36 | Purifying selection |
| ZmGLK44/PeGLK62 | 0.20 | 0.46 | 0.43 | Purifying selection |
| ZmGLK44/PeGLK65 | 0.12 | 0.38 | 0.32 | Purifying selection |
| ZmGLK26/PeGLK59 | 0.20 | 0.39 | 0.51 | Purifying selection |
| ZmGLK45/PeGLK55 | 0.09 | 0.56 | 0.17 | Purifying selection |
| ZmGLK19/PeGLK3 | 0.13 | 0.35 | 0.39 | Purifying selection |
| ZmGLK18/PeGLK33 | 0.32 | 0.84 | 0.39 | Purifying selection |
| ZmGLK41/PeGLK68 | 0.07 | 0.31 | 0.24 | Purifying selection |
| ZmGLK47/PeGLK67 | 0.07 | 0.37 | 0.20 | Purifying selection |
| ZmGLK50/PeGLK68 | 0.10 | 0.33 | 0.31 | Purifying selection |
| ZmGLK50/PeGLK67 | 0.08 | 0.38 | 0.20 | Purifying selection |
| ZmGLK31/PeGLK15 | 0.25 | 0.79 | 0.32 | Purifying selection |
| ZmGLK37/PeGLK38 | 0.10 | 0.64 | 0.15 | Purifying selection |
| ZmGLK17/PeGLK9 | 0.31 | 0.64 | 0.48 | Purifying selection |
| ZmGLK17/PeGLK10 | 0.34 | 0.70 | 0.49 | Purifying selection |
| ZmGLK17/PeGLK7 | 0.33 | 0.70 | 0.47 | Purifying selection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, R.; Guo, L.; Wang, R.; Zhang, Q.; Yao, H. Genome-Wide Identification and Characterization of G2-Like Transcription Factor Genes in Moso Bamboo (Phyllostachys edulis). Molecules 2022, 27, 5491. https://doi.org/10.3390/molecules27175491
Wu R, Guo L, Wang R, Zhang Q, Yao H. Genome-Wide Identification and Characterization of G2-Like Transcription Factor Genes in Moso Bamboo (Phyllostachys edulis). Molecules. 2022; 27(17):5491. https://doi.org/10.3390/molecules27175491
Chicago/Turabian StyleWu, Ruihua, Lin Guo, Ruoyu Wang, Qian Zhang, and Hongjun Yao. 2022. "Genome-Wide Identification and Characterization of G2-Like Transcription Factor Genes in Moso Bamboo (Phyllostachys edulis)" Molecules 27, no. 17: 5491. https://doi.org/10.3390/molecules27175491
APA StyleWu, R., Guo, L., Wang, R., Zhang, Q., & Yao, H. (2022). Genome-Wide Identification and Characterization of G2-Like Transcription Factor Genes in Moso Bamboo (Phyllostachys edulis). Molecules, 27(17), 5491. https://doi.org/10.3390/molecules27175491