
Rhamnose-Containing Compounds: Biosynthesis and Applications

 , , and
, , and
Abstract
1. Introduction
2. Biosynthetic Pathways of Donors of RCCs
2.1. Biosynthetic Pathways of dTDP-Rha
2.2. Biosynthetic Pathway of GDP-Rha
2.3. Biosynthesis Pathway of UDP-Rha
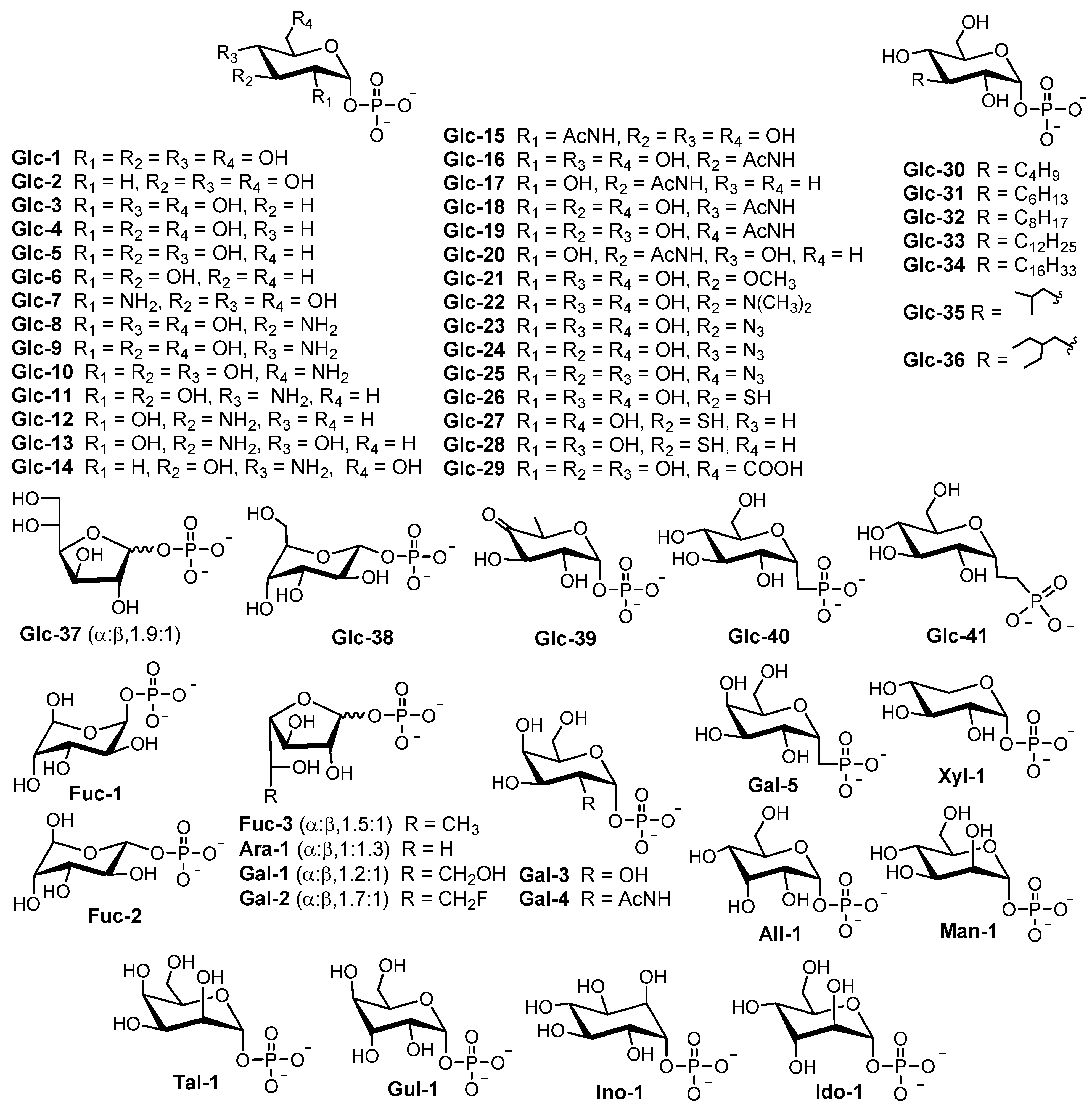
3. Rha-Ts Generating RCCs in Bacteria
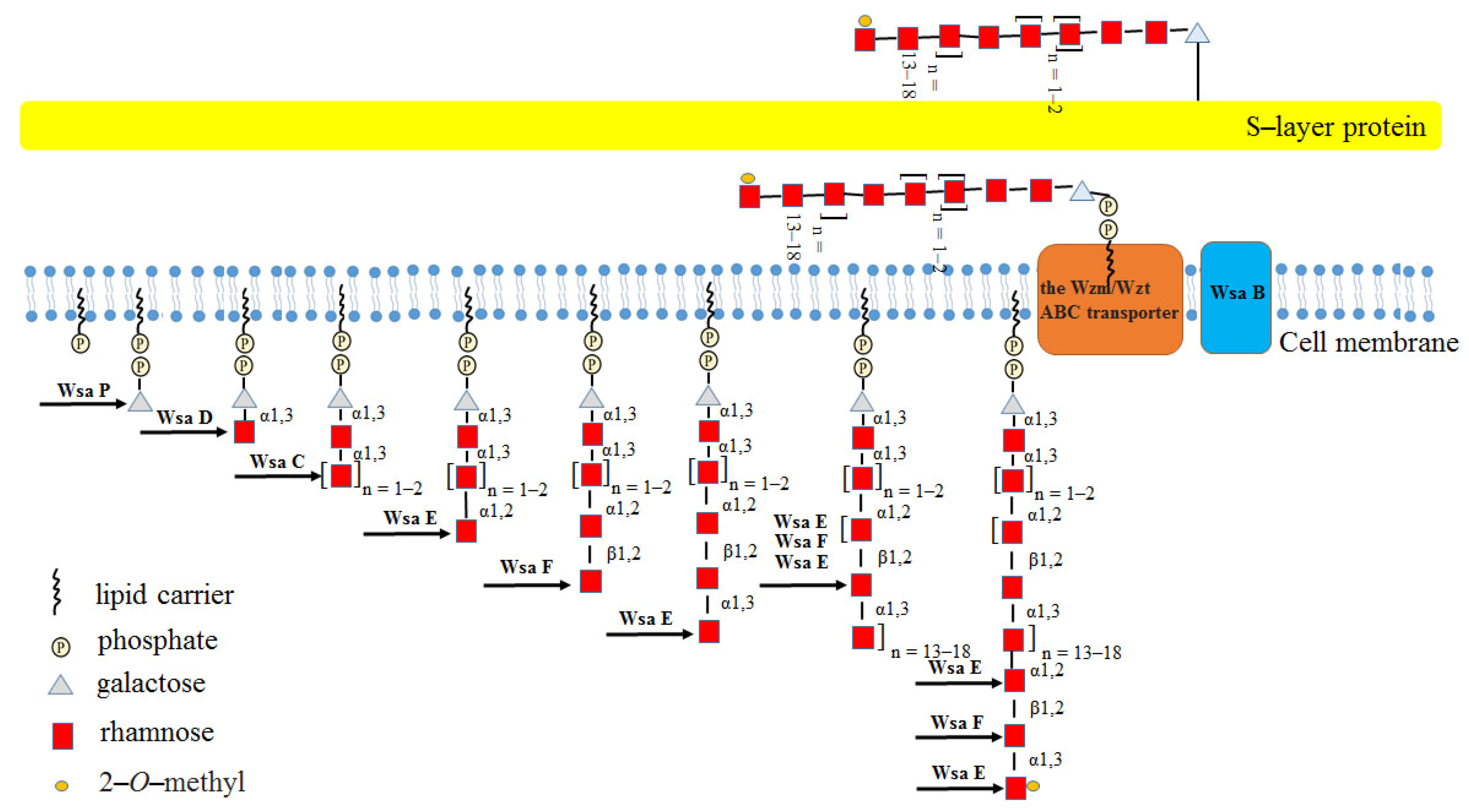
3.1. Rha-Ts from Geobacillus Stearothermophilus
3.2. Spinosyn SpnG Rha-T from Saccharopolyspora Spinosa
3.3. WbbL from Mycobacterium Tuberculosis
3.4. Rha-Ts from Pseudomonas Aeruginosa
3.5. Rha-Ts from Streptococcus Pneumoniae
3.6. Rha-Tss from Other Bacteria
4. Application of RCC in the Research and Development of New Drugs
4.1. Rha Increases the Immunogenicity of Tumour-Associated Carbohydrate Antigen (TACA) Vaccines
4.2. Rha-Containing Tumor-Killing Agents
4.3. Inhibitors of Rha Synthetases as Drug Targets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- McArthur, J.B.; Chen, X. Glycosyltransferase engineering for carbohydrate synthesis. Biochem. Soc. Trans. 2016, 44, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, P.; Lee, K.; Sarkar, S.; Wall, K.A.; Sucheck, S.J. Synthesis of a Liposomal MUC1 Glycopeptide-Based Immunotherapeutic and Evaluation of the Effect of L-Rhamnose Targeting on Cellular Immune Responses. Bioconjug. Chem. 2016, 27, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.X.; Cheng, L.; Pan, M.; Qian, Q.; Zhu, Y.L.; Xu, L.Y.; Ding, Q. D Rhamnose beta-Hederin against human breast cancer by reducing tumor-derived exosomes. Oncol. Lett. 2018, 16, 5172–5178. [Google Scholar] [PubMed]
- Vanacore, A.; Vitiello, G.; Wanke, A.; Cavasso, D.; Clifton, L.A.; Mahdi, L.; Campanero-Rhodes, M.A.; Solis, D.; Wuhrer, M.; Nicolardi, S.; et al. Lipopolysaccharide O-antigen molecular and supramolecular modifications of plant root microbiota are pivotal for host recognition. Carbohydr. Polym. 2022, 277, 118839. [Google Scholar] [CrossRef] [PubMed]
- Concordio-Reis, P.; Alves, V.D.; Moppert, X.; Guezennec, J.; Freitas, F.; Reis, M.A.M. Characterization and Biotechnological Potential of Extracellular Polysaccharides Synthesized by Alteromonas Strains Isolated from French Polynesia Marine Environments. Mar. Drugs 2021, 19, 522. [Google Scholar] [CrossRef]
- Garcia-Vello, P.; Sharma, G.; Speciale, I.; Molinaro, A.; Im, S.H.; De Castro, C. Structural features and immunological perception of the cell surface glycans of Lactobacillus plantarum: A novel rhamnose-rich polysaccharide and teichoic acids. Carbohydr. Polym. 2020, 233, 115857. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhou, P.; Qian, S.; Qin, X.; Fan, Z.; Fu, Q.; Zhan, Z.; Pei, H. Cloning, expression, identification and bioinformatics analysis of Rv3265c gene from Mycobacterium tuberculosis in Escherichia coli. Asian Pac. J. Trop. Med. 2011, 4, 266–270. [Google Scholar] [CrossRef]
- De Castro, C.; Molinaro, A.; Piacente, F.; Gurnon, J.R.; Sturiale, L.; Palmigiano, A.; Lanzetta, R.; Parrilli, M.; Garozzo, D.; Tonetti, M.G.; et al. Structure of N-linked oligosaccharides attached to chlorovirus PBCV-1 major capsid protein reveals unusual class of complex N-glycans. Proc. Natl. Acad. Sci. USA 2013, 110, 13956–13960. [Google Scholar] [CrossRef]
- Martinez, V.; Ingwers, M.; Smith, J.; Glushka, J.; Yang, T.; Bar-Peled, M. Biosynthesis of UDP-4-keto-6-deoxyglucose and UDP-rhamnose in pathogenic fungi Magnaporthe grisea and Botryotinia fuckeliana. J. Biol. Chem. 2012, 287, 879–892. [Google Scholar] [CrossRef]
- Yu, Y.; Wen, Q.; Song, A.; Liu, Y.; Wang, F.; Jiang, B. Isolation and immune activity of a new acidic Cordyceps militaris exopolysaccharide. Int. J. Biol. Macromol. 2022, 194, 706–714. [Google Scholar] [CrossRef]
- Allen, S.; Richardson, J.M.; Mehlert, A.; Ferguson, M.A. Structure of a complex phosphoglycan epitope from gp72 of Trypanosoma cruzi. J. Biol. Chem. 2013, 288, 11093–11105. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Vartak, A.; Sucheck, S.J.; Wall, K.A. Synthesis and Immunological Evaluation of a Single Molecular Construct MUC1 Vaccine Containing L-Rhamnose Repeating Units. Molecules 2020, 25, 3137. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, B.A.; Zorzoli, A.; Dorfmueller, H.C. NDP-rhamnose biosynthesis and rhamnosyltransferases: Building diverse glycoconjugates in nature. Biochem. J. 2021, 478, 685–701. [Google Scholar] [CrossRef]
- Oh, J.; Lee, S.G.; Kim, B.G.; Sohng, J.K.; Liou, K.; Lee, H.C. One-pot enzymatic production of dTDP-4-keto-6-deoxy-D-glucose from dTMP and glucose-1-phosphate. Biotechnol. Bioeng. 2003, 84, 452–458. [Google Scholar] [CrossRef]
- Glaser, L. The synthesis of thymidine-linked sugars. III. On the mechanism of thymidine diphosphate-L-rhamnose formation. Biochim. Biophys. Acta 1961, 51, 169–171. [Google Scholar] [CrossRef]
- Thibodeaux, C.J.; Melancon, C.R.; Liu, H.W. Natural-product sugar biosynthesis and enzymatic glycodiversification. Angew. Chem. Int. Ed. Engl. 2008, 47, 9814–9859. [Google Scholar] [CrossRef]
- Pal, D.; Mukhopadhyay, B. Chemical Synthesis of beta-L-Rhamnose Containing the Pentasaccharide Repeating Unit of the O-Specific Polysaccharide from a Halophilic Bacterium Halomonas ventosae RU5S2EL in the Form of Its 2-Aminoethyl Glycoside. J. Org. Chem. 2021, 86, 8683–8694. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, M.; Prevost, M.J.; Lavoie, S.; Feroldi, T.; Piochon, M.; Groleau, M.C.; Legault, J.; Villaume, S.; Crouzet, J.; Dorey, S.; et al. Total synthesis, isolation, surfactant properties, and biological evaluation of ananatosides and related macrodilactone-containing rhamnolipids. Chem. Sci. 2021, 12, 7533–7546. [Google Scholar] [CrossRef]
- Jordan, D.S.; Daubenspeck, J.M.; Dybvig, K. Rhamnose biosynthesis in mycoplasmas requires precursor glycans larger than monosaccharide. Mol. Microbiol. 2013, 89, 918–928. [Google Scholar] [CrossRef]
- Madduri, K.; Waldron, C.; Merlo, D.J. Rhamnose biosynthesis pathway supplies precursors for primary and secondary metabolism in Saccharopolyspora spinosa. J. Bacteriol. 2001, 183, 5632–5638. [Google Scholar] [CrossRef]
- Yang, S.; An, X.; Gu, G.; Yan, Z.; Jiang, X.; Xu, L.; Xiao, M. Novel dTDP-L-Rhamnose Synthetic Enzymes (RmlABCD) From Saccharothrix syringae CGMCC 4.1716 for One-Pot Four-Enzyme Synthesis of dTDP-L-Rhamnose. Front. Microbiol. 2021, 12, 772839. [Google Scholar] [CrossRef] [PubMed]
- Li, S. Studies and Application of Rhamnosyltransferase and Related Enzymes from Streptococcus pneumonia Serotype 23F. Ph.D. Thesis, Shandong University, Shandong, China, 2017. [Google Scholar]
- Li, S.; Wang, H.; Ma, J.; Gu, G.; Chen, Z.; Guo, Z. One-pot four-enzyme synthesis of thymidinediphosphate-L-rhamnose. Chem. Commun. 2016, 52, 13995–13998. [Google Scholar] [CrossRef] [PubMed]
- Pazur, J.H.; Anderson, J.S. Thymidine triphosphate: Alpha-D-galactose-L-phosphate thymj-L-phosphate thymidylyltransferase from streptococcus faecalis grown on d-galactose. J. Biol. Chem. 1963, 238, 3155–3160. [Google Scholar] [CrossRef]
- Barton, W.A.; Lesniak, J.; Biggins, J.B.; Jeffrey, P.D.; Jiang, J.; Rajashankar, K.R.; Thorson, J.S.; Nikolov, D.B. Structure, mechanism and engineering of a nucleotidylyltransferase as a first step toward glycorandomization. Nat. Struct. Biol. 2001, 8, 545–551. [Google Scholar] [CrossRef]
- Jiang, J.; Biggins, J.B.; Thorson, J.S. A General Enzymatic Method for the Synthesis of Natural and “Unnatural” UDP- and TDP-Nucleotide Sugars. J. Am. Chem. Soc. 2000, 122, 6803–6804. [Google Scholar] [CrossRef]
- Barton, W.A.; Biggins, J.B.; Jiang, J.; Thorson, J.S.; Nikolov, D.B. Expanding pyrimidine diphosphosugar libraries via structure-based nucleotidylyltransferase engineering. Proc. Natl. Acad. Sci. USA 2002, 99, 13397–13402. [Google Scholar] [CrossRef]
- Moretti, R.; Chang, A.; Peltier-Pain, P.; Bingman, C.A.; Phillips, G.J.; Thorson, J.S. Expanding the nucleotide and sugar 1-phosphate promiscuity of nucleotidyltransferase RmlA via directed evolution. J. Biol. Chem. 2011, 286, 13235–13243. [Google Scholar] [CrossRef]
- Jakeman, D.L.; Young, J.L.; Huestis, M.P.; Peltier, P.; Daniellou, R.; Nugier-Chauvin, C.; Ferrieres, V. Engineering ribonucleoside triphosphate specificity in a thymidylyltransferase. Biochemistry 2008, 47, 8719–8725. [Google Scholar] [CrossRef]
- Jiqing Jiang, J.B.B.A. Expanding the Pyrimidine Diphosphosugar Repertoire: The Chemoenzymatic Synthesis of Amino- and Acetamidoglucopyranosyl Derivatives. Angew. Chem. Int. Ed. Engl. 2001, 8, 1502–1505. [Google Scholar] [CrossRef]
- Timmons, S.C.; Mosher, R.H.; Knowles, S.A.; Jakeman, D.L. Exploiting nucleotidylyltransferases to prepare sugar nucleotides. Org. Lett. 2007, 9, 857–860. [Google Scholar] [CrossRef]
- Forget, S.M.; Smithen, D.A.; Jee, A.; Jakeman, D.L. Mechanistic evaluation of a nucleoside tetraphosphate with a thymidylyltransferase. Biochemistry 2015, 54, 1703–1707. [Google Scholar] [CrossRef] [PubMed]
- Huestis, M.P.; Aish, G.A.; Hui, J.P.; Soo, E.C.; Jakeman, D.L. Lipophilic sugar nucleotide synthesis by structure-based design of nucleotidylyltransferase substrates. Org. Biomol. Chem. 2008, 6, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Timmons, S.C.; Hui, J.P.; Pearson, J.L.; Peltier, P.; Daniellou, R.; Nugier-Chauvin, C.; Soo, E.C.; Syvitski, R.T.; Ferrieres, V.; Jakeman, D.L. Enzyme-catalyzed synthesis of furanosyl nucleotides. Org. Lett. 2008, 10, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Moretti, R.; Thorson, J.S. Enhancing the latent nucleotide triphosphate flexibility of the glucose-1-phosphate thymidylyltransferase RmlA. J. Biol. Chem. 2007, 282, 16942–16947. [Google Scholar] [CrossRef]
- Yang, J.; Hoffmeister, D.; Liu, L.; Fu, X.; Thorson, J.S. Natural product glycorandomization. Bioorg. Med. Chem. 2004, 12, 1577–1584. [Google Scholar] [CrossRef]
- Zhang, C.; Moretti, R.; Jiang, J.; Thorson, J.S. The in vitro characterization of polyene glycosyltransferases AmphDI and NysDI. ChemBioChem 2008, 9, 2506–2514. [Google Scholar] [CrossRef]
- Jiang, J.; Albermann, C.; Thorson, J.S. Application of the nucleotidylyltransferase Ep toward the chemoenzymatic synthesis of dTDP-desosamine analogues. ChemBioChem 2003, 4, 443–446. [Google Scholar] [CrossRef]
- Albermann, C.; Soriano, A.; Jiang, J.; Vollmer, H.; Biggins, J.B.; Barton, W.A.; Lesniak, J.; Nikolov, D.B.; Thorson, J.S. Substrate specificity of NovM: Implications for novobiocin biosynthesis and glycorandomization. Org. Lett. 2003, 5, 933–936. [Google Scholar] [CrossRef]
- Williams, G.J.; Gantt, R.W.; Thorson, J.S. The impact of enzyme engineering upon natural product glycodiversification. Curr. Opin. Chem. Biol. 2008, 12, 556–564. [Google Scholar] [CrossRef]
- Thorson, J.S.; Barton, W.A.; Hoffmeister, D.; Albermann, C.; Nikolov, D.B. Structure-based enzyme engineering and its impact on in vitro glycorandomization. ChemBioBhem 2004, 5, 16–25. [Google Scholar] [CrossRef]
- Beaton, S.A.; Huestis, M.P.; Sadeghi-Khomami, A.; Thomas, N.R.; Jakeman, D.L. Enzyme-catalyzed synthesis of isosteric phosphono-analogues of sugar nucleotides. Chem. Commun. 2009, 8, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.; Glaser, L. The nucleotide specificity and feedback control of thymidine diphosphate D-glucose pyrophosphorylase. J. Biol. Chem. 1965, 240, 398–405. [Google Scholar] [CrossRef]
- Blankenfeldt, W.; Giraud, M.F.; Leonard, G.; Rahim, R.; Creuzenet, C.; Lam, J.S.; Naismith, J.H. The purification, crystallization and preliminary structural characterization of glucose-1-phosphate thymidylyltransferase (RmlA), the first enzyme of the dTDP-L-rhamnose synthesis pathway from Pseudomonas aeruginosa. Acta. Crystallogr. D. Biol. Crystallogr. 2000, 56 Pt 11, 1501–1504. [Google Scholar] [CrossRef] [PubMed]
- Sivaraman, J.; Sauve, V.; Matte, A.; Cygler, M. Crystal structure of Escherichia coli glucose-1-phosphate thymidylyltransferase (RffH) complexed with dTTP and Mg2+. J. Biol. Chem. 2002, 277, 44214–44219. [Google Scholar] [CrossRef]
- Allard, S.T.; Beis, K.; Giraud, M.F.; Hegeman, A.D.; Gross, J.W.; Wilmouth, R.C.; Whitfield, C.; Graninger, M.; Messner, P.; Allen, A.G.; et al. Toward a structural understanding of the dehydratase mechanism. Structure 2002, 10, 81–92. [Google Scholar] [CrossRef]
- James, D.B.; Yother, J. Genetic and biochemical characterizations of enzymes involved in Streptococcus pneumoniae serotype 2 capsule synthesis demonstrate that Cps2T (WchF) catalyzes the committed step by addition of beta1-4 rhamnose, the second sugar residue in the repeat unit. J. Bacteriol. 2012, 194, 6479–6489. [Google Scholar] [CrossRef]
- Dong, C.; Major, L.L.; Allen, A.; Blankenfeldt, W.; Maskell, D.; Naismith, J.H. High-resolution structures of RmlC from Streptococcus suis in complex with substrate analogs locate the active site of this class of enzyme. Structure 2003, 11, 715–723. [Google Scholar] [CrossRef]
- Dong, C.; Major, L.L.; Srikannathasan, V.; Errey, J.C.; Giraud, M.F.; Lam, J.S.; Graninger, M.; Messner, P.; McNeil, M.R.; Field, R.A.; et al. RmlC, a C3′ and C5′ carbohydrate epimerase, appears to operate via an intermediate with an unusual twist boat conformation. J. Mol. Biol. 2007, 365, 146–159. [Google Scholar] [CrossRef]
- Giraud, M.F.; Leonard, G.A.; Field, R.A.; Berlind, C.; Naismith, J.H. RmlC, the third enzyme of dTDP-L-rhamnose pathway, is a new class of epimerase. Nat. Struct. Biol. 2000, 7, 398–402. [Google Scholar]
- Giraud, M.F.; Gordon, F.M.; Whitfield, C.; Messner, P.; McMahon, S.A.; Naismith, J.H. Purification, crystallization and preliminary structural studies of dTDP-6-deoxy-D-xylo-4-hexulose 3,5-epimerase (RmlC), the third enzyme of the dTDP-L-rhamnose synthesis pathway, from Salmonella enterica serovar typhimurium. Acta Crystallogr. D Biol. Crystallogr. 1999, 55 Pt 3, 706–708. [Google Scholar] [CrossRef]
- Blankenfeldt, W.; Kerr, I.D.; Giraud, M.F.; McMiken, H.J.; Leonard, G.; Whitfield, C.; Messner, P.; Graninger, M.; Naismith, J.H. Variation on a theme of SDR. dTDP-6-deoxy-L- lyxo-4-hexulose reductase (RmlD) shows a new Mg2+-dependent dimerization mode. Structure 2002, 10, 773–786. [Google Scholar] [CrossRef]
- Graninger, M.; Nidetzky, B.; Heinrichs, D.E.; Whitfield, C.; Messner, P. Characterization of dTDP-4-dehydrorhamnose 3,5-epimerase and dTDP-4-dehydrorhamnose reductase, required for dTDP-L-rhamnose biosynthesis in Salmonella enterica serovar Typhimurium LT2. J. Biol. Chem. 1999, 274, 25069–25077. [Google Scholar] [CrossRef] [PubMed]
- Ovod, V.; Rudolph, K.; Knirel, Y.; Krohn, K. Immunochemical characterization of O polysaccharides composing the alpha-D-rhamnose backbone of lipopolysaccharide of Pseudomonas syringae and classification of bacteria into serogroups O1 and O2 with monoclonal antibodies. J. Bacteriol. 1996, 178, 6459–6465. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kneidinger, B.; Graninger, M.; Adam, G.; Puchberger, M.; Kosma, P.; Zayni, S.; Messner, P. Identification of two GDP-6-deoxy-D-lyxo-4-hexulose reductases synthesizing GDP-D-rhamnose in Aneurinibacillus thermoaerophilus L420-91T. J. Biol. Chem. 2001, 276, 5577–5583. [Google Scholar] [CrossRef] [PubMed]
- Webb, N.A.; Mulichak, A.M.; Lam, J.S.; Rocchetta, H.L.; Garavito, R.M. Crystal structure of a tetrameric GDP-D-mannose 4,6-dehydratase from a bacterial GDP-D-rhamnose biosynthetic pathway. Protein Sci. 2004, 13, 529–539. [Google Scholar] [CrossRef]
- Ginsburg, V. Formation of guanosine diphosphate L-fucose from guanosine diphosphate D-mannose. J. Biol. Chem. 1960, 235, 2196–2201. [Google Scholar] [CrossRef]
- Mulichak, A.M.; Bonin, C.P.; Reiter, W.D.; Garavito, R.M. Structure of the MUR1 GDP-mannose 4,6-dehydratase from Arabidopsis thaliana: Implications for ligand binding and specificity. Biochemistry 2002, 41, 15578–15589. [Google Scholar] [CrossRef]
- Reitman, M.L.; Trowbridge, I.S.; Kornfeld, S. Mouse lymphoma cell lines resistant to pea lectin are defective in fucose metabolism. J. Biol. Chem. 1980, 255, 9900–9906. [Google Scholar] [CrossRef]
- Bisso, A.; Sturla, L.; Zanardi, D.; De Flora, A.; Tonetti, M. Structural and enzymatic characterization of human recombinant GDP-D-mannose-4,6-dehydratase. FEBS. Lett. 1999, 456, 370–374. [Google Scholar] [CrossRef]
- Tonetti, M.; Zanardi, D.; Gurnon, J.R.; Fruscione, F.; Armirotti, A.; Damonte, G.; Sturla, L.; De Flora, A.; Van Etten, J.L. Paramecium bursaria Chlorella virus 1 encodes two enzymes involved in the biosynthesis of GDP-L-fucose and GDP-D-rhamnose. J. Biol. Chem. 2003, 278, 21559–21565. [Google Scholar] [CrossRef]
- Somoza, J.R.; Menon, S.; Schmidt, H.; Joseph-McCarthy, D.; Dessen, A.; Stahl, M.L.; Somers, W.S.; Sullivan, F.X. Structural and kinetic analysis of Escherichia coli GDP-mannose 4,6 dehydratase provides insights into the enzyme’s catalytic mechanism and regulation by GDP-fucose. Structure 2000, 8, 123–135. [Google Scholar] [CrossRef]
- Rosano, C.; Zuccotti, S.; Sturla, L.; Fruscione, F.; Tonetti, M.; Bolognesi, M. Quaternary assembly and crystal structure of GDP-D-mannose 4,6 dehydratase from Paramecium bursaria Chlorella virus. Biochem. Biophys. Res. Commun. 2006, 339, 191–195. [Google Scholar] [CrossRef] [PubMed]
- King, J.D.; Poon, K.K.; Webb, N.A.; Anderson, E.M.; McNally, D.J.; Brisson, J.R.; Messner, P.; Garavito, R.M.; Lam, J.S. The structural basis for catalytic function of GMD and RMD, two closely related enzymes from the GDP-D-rhamnose biosynthesis pathway. FEBS J. 2009, 276, 2686–2700. [Google Scholar] [CrossRef]
- Qian, L.; Han, X.; Zhang, L.; Liu, X. Structural and biochemical insights into nucleotide-rhamnose synthase/epimerase-reductase from Arabidopsis thaliana. Biochim. Biophys. Acta 2015, 10 Pt A, 1476–1486. [Google Scholar]
- Oka, T.; Nemoto, T.; Jigami, Y. Functional analysis of Arabidopsis thaliana RHM2/MUM4, a multidomain protein involved in UDP-D-glucose to UDP-L-rhamnose conversion. J. Biol. Chem. 2007, 282, 5389–5403. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, J.J.; Kasimova, A.A.; Sviridova, A.N.; Shpirt, A.M.; Shneider, M.M.; Mikhaylova, Y.V.; Shelenkov, A.A.; Popova, A.V.; Perepelov, A.V.; Shashkov, A.S.; et al. Correlation of Acinetobacter baumannii K144 and K86 capsular polysaccharide structures with genes at the K locus reveals the involvement of a novel multifunctional rhamnosyltransferase for structural synthesis. Int. J. Biol. Macromol. 2021, 193 Pt B, 1294–1300. [Google Scholar] [CrossRef]
- Kenyon, J.J.; Arbatsky, N.P.; Sweeney, E.L.; Zhang, Y.; Senchenkova, S.N.; Popova, A.V.; Shneider, M.M.; Shashkov, A.S.; Liu, B.; Hall, R.M.; et al. Involvement of a multifunctional rhamnosyltransferase in the synthesis of three related Acinetobacter baumannii capsular polysaccharides, K55, K74 and K85. Int. J. Biol. Macromol. 2021, 166, 1230–1237. [Google Scholar] [CrossRef]
- Steiner, K.; Hagelueken, G.; Messner, P.; Schaffer, C.; Naismith, J.H. Structural basis of substrate binding in WsaF, a rhamnosyltransferase from Geobacillus stearothermophilus. J. Mol. Biol. 2010, 397, 436–447. [Google Scholar] [CrossRef]
- Steiner, K.; Novotny, R.; Werz, D.B.; Zarschler, K.; Seeberger, P.H.; Hofinger, A.; Kosma, P.; Schaffer, C.; Messner, P. Molecular basis of S-layer glycoprotein glycan biosynthesis in Geobacillus stearothermophilus. J. Biol. Chem. 2008, 283, 21120–21133. [Google Scholar] [CrossRef]
- Isiorho, E.A.; Liu, H.W.; Keatinge-Clay, A.T. Structural studies of the spinosyn rhamnosyltransferase, SpnG. Biochemistry. 2012, 51, 1213–1222. [Google Scholar] [CrossRef]
- Huang, K.X.; Xia, L.; Zhang, Y.; Ding, X.; Zahn, J.A. Recent advances in the biochemistry of spinosyns. Appl. Microbiol. Biotechnol. 2009, 82, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Chen, Y.H.; Lin, Y.C.; Tsai, K.C.; Chiu, H.T. Functional characterization and substrate specificity of spinosyn rhamnosyltransferase by in vitro reconstitution of spinosyn biosynthetic enzymes. J. Biol. Chem. 2009, 284, 7352–7363. [Google Scholar] [CrossRef] [PubMed]
- Grzegorzewicz, A.E.; Ma, Y.; Jones, V.; Crick, D.; Liav, A.; McNeil, M.R. Development of a microtitre plate-based assay for lipid-linked glycosyltransferase products using the mycobacterial cell wall rhamnosyltransferase WbbL. Microbiology 2008, 154 Pt 12, 3724–3730. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sivendran, S.; Jones, V.; Sun, D.; Wang, Y.; Grzegorzewicz, A.E.; Scherman, M.S.; Napper, A.D.; McCammon, J.A.; Lee, R.E.; Diamond, S.L.; et al. Identification of triazinoindol-benzimidazolones as nanomolar inhibitors of the Mycobacterium tuberculosis enzyme TDP-6-deoxy-d-xylo-4-hexopyranosid-4-ulose 3,5-epimerase (RmlC). Bioorg. Med. Chem. 2010, 18, 896–908. [Google Scholar] [CrossRef]
- Mills, J.A.; Motichka, K.; Jucker, M.; Wu, H.P.; Uhlik, B.C.; Stern, R.J.; Scherman, M.S.; Vissa, V.D.; Pan, F.; Kundu, M.; et al. Inactivation of the mycobacterial rhamnosyltransferase, which is needed for the formation of the arabinogalactan-peptidoglycan linker, leads to irreversible loss of viability. J. Biol. Chem. 2004, 279, 43540–43546. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Ramirez, M.; Medina, G.; Gonzalez-Valdez, A.; Grosso-Becerra, V.; Soberon-Chavez, G. The Pseudomonas aeruginosa rmlBDAC operon, encoding dTDP-L-rhamnose biosynthetic enzymes, is regulated by the quorum-sensing transcriptional regulator RhlR and the alternative sigma factor sigmaS. Microbiology 2012, 158 Pt 4, 908–916. [Google Scholar] [CrossRef]
- Poon, K.K.; Westman, E.L.; Vinogradov, E.; Jin, S.; Lam, J.S. Functional characterization of MigA and WapR: Putative rhamnosyltransferases involved in outer core oligosaccharide biosynthesis of Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 1857–1865. [Google Scholar] [CrossRef]
- Rocchetta, H.L.; Burrows, L.L.; Lam, J.S. Genetics of O-antigen biosynthesis in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 1999, 63, 523–553. [Google Scholar] [CrossRef] [PubMed]
- Rocchetta, H.L.; Burrows, L.L.; Pacan, J.C.; Lam, J.S. Three rhamnosyltransferases responsible for assembly of the A-band D-rhamnan polysaccharide in Pseudomonas aeruginosa: A fourth transferase, WbpL, is required for the initiation of both A-band and B-band lipopolysaccharide synthesis. Mol. Microbiol. 1998, 28, 1103–1119. [Google Scholar] [CrossRef]
- Melamed, J.; Kocev, A.; Torgov, V.; Veselovsky, V.; Brockhausen, I. Biosynthesis of the Pseudomonas aeruginosa common polysaccharide antigen by D-Rhamnosyltransferases WbpX and WbpY. Glycoconj. J. 2022, 39, 393–411. [Google Scholar] [CrossRef]
- Zhu, K.; Rock, C.O. RhlA converts beta-hydroxyacyl-acyl carrier protein intermediates in fatty acid synthesis to the beta-hydroxydecanoyl-beta-hydroxydecanoate component of rhamnolipids in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 3147–3154. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Fiechter, A.; Reiser, J. Isolation, characterization, and expression in Escherichia coli of the Pseudomonas aeruginosa rhlAB genes encoding a rhamnosyltransferase involved in rhamnolipid biosurfactant synthesis. J. Biol. Chem. 1994, 269, 19787–19795. [Google Scholar] [CrossRef]
- Rahim, R.; Ochsner, U.A.; Olvera, C.; Graninger, M.; Messner, P.; Lam, J.S.; Soberon-Chavez, G. Cloning and functional characterization of the Pseudomonas aeruginosa rhlC gene that encodes rhamnosyltransferase 2, an enzyme responsible for di-rhamnolipid biosynthesis. Mol. Microbiol. 2001, 40, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Deziel, E.; Lepine, F.; Milot, S.; Villemur, R. rhlA is required for the production of a novel biosurfactant promoting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs), the precursors of rhamnolipids. Microbiology 2003, 149 Pt 8, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.F.; Silva, P.M.; Junqueira, M.; Mariano, D.C.; Nogueira, F.C.; Domont, G.B.; Freire, D.M.; Neves, B.C. Characterization of rhamnolipids produced by wild-type and engineered Burkholderia kururiensis. Appl. Microbiol. Biotechnol. 2013, 97, 1909–1921. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Luo, J.; Li, S. Bacteria-Catalyzed Arginine Glycosylation in Pathogens and Host. Front. Cell. Infect. Microbiol. 2020, 10, 185. [Google Scholar] [CrossRef]
- He, C.; Liu, N.; Li, F.; Jia, X.; Peng, H.; Liu, Y.; Xiao, Y. Complex Structure of Pseudomonas aeruginosa Arginine Rhamnosyltransferase EarP with Its Acceptor Elongation Factor P. J. Bacteriol. 2019, 201, e00128-19. [Google Scholar] [CrossRef]
- Lassak, J.; Keilhauer, E.C.; Fürst, M.; Wuichet, K.; Gödeke, J.; Starosta, A.L.; Chen, J.M.; Søgaard-Andersen, L.; Rohr, J.; Wilson, D.N.; et al. Arginine-rhamnosylation as new strategy to activate translation elongation factor P. Nat. Chem. Biol. 2015, 11, 266–270. [Google Scholar] [CrossRef]
- Yanagisawa, T.; Takahashi, H.; Suzuki, T.; Masuda, A.; Dohmae, N.; Yokoyama, S. Neisseria meningitidis Translation Elongation Factor P and Its Active-Site Arginine Residue Are Essential for Cell Viability. PLoS ONE 2016, 11, e0147907. [Google Scholar] [CrossRef]
- Geno, K.A.; Gilbert, G.L.; Song, J.Y.; Skovsted, I.C.; Klugman, K.P.; Jones, C.; Konradsen, H.B.; Nahm, M.H. Pneumococcal Capsules and Their Types: Past, Present, and Future. Clin. Microbiol. Rev. 2015, 28, 871–899. [Google Scholar] [CrossRef]
- Park, S.; Nahm, M.H. L-rhamnose is often an important part of immunodominant epitope for pneumococcal serotype 23F polysaccharide antibodies in human sera immunized with PPV23. PLoS ONE 2013, 8, e83810. [Google Scholar] [CrossRef] [PubMed]
- Legnani, L.; Ronchi, S.; Fallarini, S.; Lombardi, G.; Campo, F.; Panza, L.; Lay, L.; Poletti, L.; Toma, L.; Ronchetti, F.; et al. Synthesis, molecular dynamics simulations, and biology of a carba-analogue of the trisaccharide repeating unit of Streptococcus pneumoniae 19F capsular polysaccharide. Org. Biomol. Chem. 2009, 7, 4428–4436. [Google Scholar] [CrossRef] [PubMed]
- Aanensen, D.M.; Mavroidi, A.; Bentley, S.D.; Reeves, P.R.; Spratt, B.G. Predicted functions and linkage specificities of the products of the Streptococcus pneumoniae capsular biosynthetic loci. J. Bacteriol. 2007, 189, 7856–7876. [Google Scholar] [CrossRef]
- James, D.B.; Gupta, K.; Hauser, J.R.; Yother, J. Biochemical activities of Streptococcus pneumoniae serotype 2 capsular glycosyltransferases and significance of suppressor mutations affecting the initiating glycosyltransferase Cps2E. J. Bacteriol. 2013, 195, 5469–5478. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, S.; Xiong, C.; Jin, G.; Chen, Z.; Gu, G.; Guo, Z. Biochemical studies of a beta-1,4-rhamnoslytransferase from Streptococcus pneumonia serotype 23F. Org. Biomol. Chem. 2019, 17, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Ardissone, S.; Noel, K.D.; Klement, M.; Broughton, W.J.; Deakin, W.J. Synthesis of the flavonoid-induced lipopolysaccharide of Rhizobium Sp. strain NGR234 requires rhamnosyl transferases encoded by genes rgpF and wbgA. Mol. Plant-Microbe Interact. 2011, 24, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Sugano, Y.; Shoda, M. Novel glycosyltransferase genes involved in the acetan biosynthesis of Acetobacter xylinum. Biochem. Biophys. Res. Commun. 2002, 295, 230–235. [Google Scholar] [CrossRef]
- Dong, S.; McPherson, S.A.; Wang, Y.; Li, M.; Wang, P.; Turnbough, C.J.; Pritchard, D.G. Characterization of the enzymes encoded by the anthrose biosynthetic operon of Bacillus anthracis. J. Bacteriol. 2010, 192, 5053–5062. [Google Scholar] [CrossRef]
- Zorzoli, A.; Meyer, B.H.; Adair, E.; Torgov, V.I.; Veselovsky, V.V.; Danilov, L.L.; Uhrin, D.; Dorfmueller, H.C. Group A, B, C, and G Streptococcus Lancefield antigen biosynthesis is initiated by a conserved α-d-GlcNAc-β-1,4-L-rhamnosyltransferase. J. Biol. Chem. 2019, 294, 15237–15256. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Mukai, T.; Naka, T.; Fujiwara, N.; Maeda, Y.; Kai, M.; Mizuno, S.; Yano, I.; Makino, M. Novel rhamnosyltransferase involved in biosynthesis of serovar 4-specific glycopeptidolipid from Mycobacterium avium complex. J. Bacteriol. 2010, 192, 5700–5708. [Google Scholar] [CrossRef]
- Tsunashima, H.; Miyake, K.; Motono, M.; Iijima, S. Organization of the capsule biosynthesis gene locus of the oral streptococcus Streptococcus anginosus. J. Biosci. Bioeng. 2012, 113, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Gong, W.; Sun, C.; Zhao, J.; Wang, H.; Chen, Z.; Xiao, M.; Gu, G. Sequential One-Pot Three-Enzyme Synthesis of the Tetrasaccharide Repeating Unit of Group B Streptococcus Serotype VIII Capsular Polysaccharide. Chin. J. Chem. 2022, 40, 1039–1044. [Google Scholar] [CrossRef]
- Li, Q.; Hobbs, M.; Reeves, P.R. The variation of dTDP-L-rhamnose pathway genes in Vibrio cholerae. Microbiology 2003, 149 Pt 9, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Chen, R.; Xie, K.; Chen, D.; Guo, B.; Liu, X.; Liu, J.; Zhang, M.; Dai, J. A regiospecific rhamnosyltransferase from Epimedium pseudowushanense catalyzes the 3-O-rhamnosylation of prenylflavonols. Org. Biomol. Chem. 2018, 16, 452–458. [Google Scholar] [CrossRef]
- Zong, G.; Fei, S.; Liu, X.; Li, J.; Gao, Y.; Yang, X.; Wang, X.; Shen, Y. Crystal structures of rhamnosyltransferase UGT89C1 from Arabidopsis thaliana reveal the molecular basis of sugar donor specificity for UDP-β-L-rhamnose and rhamnosylation mechanism. Plant J. 2019, 99, 257–269. [Google Scholar]
- Takenaka, Y.; Kato, K.; Ogawa-Ohnishi, M.; Tsuruhama, K.; Kajiura, H.; Yagyu, K.; Takeda, A.; Takeda, Y.; Kunieda, T.; Hara-Nishimura, I.; et al. Pectin RG-I rhamnosyltransferases represent a novel plant-specific glycosyltransferase family. Nat. Plants 2018, 4, 669–676. [Google Scholar] [CrossRef]
- Lu, C.; Dong, Y.; Ke, K.; Zou, K.; Wang, Z.; Xiao, W.; Pei, J.; Zhao, L. Modification to increase the thermostability and catalytic efficiency of α-L-rhamnosidase from Bacteroides thetaiotaomicron and high-level expression. Enzym. Microb. Technol. 2022, 158, 110040. [Google Scholar] [CrossRef]
- Xu, L.; Liu, X.; Li, Y.; Yin, Z.; Jin, L.; Lu, L.; Qu, J.; Xiao, M. Enzymatic rhamnosylation of anticancer drugs by an α-L-rhamnosidase from Alternaria sp. L1 for cancer-targeting and enzyme-activated prodrug therapy. Appl. Microbiol. Biotechnol. 2019, 103, 7997–8008. [Google Scholar] [CrossRef]
- Hakomori, S. Tumor-associated carbohydrate antigens defining tumor malignancy: Basis for development of anti-cancer vaccines. Adv. Exp. Med. Biol. 2001, 491, 369–402. [Google Scholar]
- Hakomori, S. Aberrant glycosylation in cancer cell membranes as focused on glycolipids: Overview and perspectives. Cancer Res. 1985, 45, 2405–2414. [Google Scholar]
- Guo, Z.; Wang, Q. Recent development in carbohydrate-based cancer vaccines. Curr. Opin. Chem. Biol. 2009, 13, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.M.; Danishefsky, S.J. A Vision for Vaccines Built from Fully Synthetic Tumor-Associated Antigens: From the Laboratory to the Clinic. J. Am. Chem. Soc. 2013, 135, 14462–14472. [Google Scholar] [CrossRef]
- Grabenstein, J.D.; Klugman, K.P. A century of pneumococcal vaccination research in humans. Clin. Microbiol. Infect. 2012, 18 (Suppl. 5), 15–24. [Google Scholar] [CrossRef]
- Holmberg, L.A.; Sandmaier, B.M. Vaccination with Theratope (STn-KLH) as treatment for breast cancer. Expert. Rev. Vaccines 2004, 3, 655–663. [Google Scholar] [CrossRef]
- Franco, A. Glycoconjugates as vaccines for cancer immunotherapy: Clinical trials and future directions. Anticancer Agents Med. Chem. 2008, 8, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, Z.; Tang, S.; Guo, Z. Carbohydrate-monophosphoryl lipid a conjugates are fully synthetic self-adjuvanting cancer vaccines eliciting robust immune responses in the mouse. ACS Chem. Biol. 2012, 7, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wang, Q.; Zhang, J.; Wu, Q.; Guo, Z. Synthesis and Evaluation of Protein Conjugates of GM3 Derivatives Carrying Modified Sialic Acids as Highly Immunogenic Cancer Vaccine Candidates. Medchemcomm 2011, 2, 524–530. [Google Scholar] [CrossRef][Green Version]
- Oyelaran, O.; McShane, L.M.; Dodd, L.; Gildersleeve, J.C. Profiling human serum antibodies with a carbohydrate antigen microarray. J. Proteome Res. 2009, 8, 4301–4310. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, B.; Ma, Z.; Wei, M.; Liu, J.; Li, D.; Zhang, H.; Wang, P.G.; Chen, M. L-Rhamnose Enhances the Immunogenicity of Melanoma-Associated Antigen A3 for Stimulating Antitumor Immune Responses. Bioconjug. Chem. 2016, 27, 1112–1118. [Google Scholar] [CrossRef]
- Sarkar, S.; Lombardo, S.A.; Herner, D.N.; Talan, R.S.; Wall, K.A.; Sucheck, S.J. Synthesis of a single-molecule L-rhamnose-containing three-component vaccine and evaluation of antigenicity in the presence of anti-L-rhamnose antibodies. J. Am. Chem. Soc. 2010, 132, 17236–17246. [Google Scholar] [CrossRef]
- Sarkar, S.; Salyer, A.C.; Wall, K.A.; Sucheck, S.J. Synthesis and immunological evaluation of a MUC1 glycopeptide incorporated into L-rhamnose displaying liposomes. Bioconjug. Chem. 2013, 24, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Rao, X.; Cai, L.; Liu, X.; Wang, H.; Wu, W.; Zhu, C.; Chen, M.; Wang, P.G.; Yi, W. Targeting Tumor Cells by Natural Anti-Carbohydrate Antibodies Using Rhamnose-Functionalized Liposomes. ACS Chem. Biol. 2016, 11, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Vartak, A.; Karmakar, P.; Sucheck, S.J.; Wall, K.A. Augmenting Vaccine Immunogenicity through the Use of Natural Human Anti-rhamnose Antibodies. ACS Chem. Biol. 2018, 13, 2130–2142. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Hong, H.; Wang, J.; Li, C.; Zhou, Z.; Wu, Z. Rhamnose modified bovine serum albumin as a carrier protein promotes the immune response against sTn antigen. Chem. Commun. 2020, 56, 13959–13962. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gu, L.; Zhang, W.; Motari, E.; Cai, L.; Styslinger, T.J.; Wang, P.G. L-rhamnose antigen: A promising alternative to alpha-gal for cancer immunotherapies. ACS Chem. Biol. 2011, 6, 185–191. [Google Scholar] [CrossRef]
- Ou, C.; Prabhu, S.K.; Zhang, X.; Zong, G.; Yang, Q.; Wang, L.X. Synthetic Antibody-Rhamnose Cluster Conjugates Show Potent Complement-Dependent Cell Killing by Recruiting Natural Antibodies. Chemistry 2022, 28, e202200146. [Google Scholar] [CrossRef]
- Zhou, K.; Hong, H.; Lin, H.; Gong, L.; Li, D.; Shi, J.; Zhou, Z.; Xu, F.; Wu, Z. Chemical Synthesis of Antibody-Hapten Conjugates Capable of Recruiting the Endogenous Antibody to Magnify the Fc Effector Immunity of Antibody for Cancer Immunotherapy. J. Med. Chem. 2022, 65, 323–332. [Google Scholar] [CrossRef]
- De Coen, R.; Nuhn, L.; Perera, C.; Arista-Romero, M.; Risseeuw, M.D.P.; Freyn, A.; Nachbagauer, R.; Albertazzi, L.; Van Calenbergh, S.; Spiegel, D.A.; et al. Synthetic Rhamnose Glycopolymer Cell-Surface Receptor for Endogenous Antibody Recruitment. Biomacromolecules 2020, 21, 793–802. [Google Scholar] [CrossRef]
- Hilinski, M.K.; Mrozowski, R.M.; Clark, D.E.; Lannigan, D.A. Analogs of the RSK inhibitor SL0101: Optimization of in vitro biological stability. Bioorg. Med. Chem. Lett. 2012, 22, 3244–3247. [Google Scholar] [CrossRef]
- Chun, J.; Ha, I.J.; Kim, Y.S. Antiproliferative and apoptotic activities of triterpenoid saponins from the roots of Platycodon grandiflorum and their structure-activity relationships. Planta Med. 2013, 79, 639–645. [Google Scholar] [CrossRef]
- Ha, I.J.; Kang, M.; Na, Y.C.; Park, Y.; Kim, Y.S. Preparative separation of minor saponins from Platycodi Radix by high-speed counter-current chromatography. J. Sep. Sci. 2011, 34, 2559–2565. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, J.; Gu, G.; Li, G.; Cui, C.; Sun, B.; Lou, H. In situ RBL receptor visualization and its mediated anticancer activity for solasodine rhamnosides. Chembiochem 2011, 12, 2418–2420. [Google Scholar] [CrossRef] [PubMed]
- Garnier, P.; Wang, X.T.; Robinson, M.A.; van Kasteren, S.; Perkins, A.C.; Frier, M.; Fairbanks, A.J.; Davis, B.G. Lectin-directed enzyme activated prodrug therapy (LEAPT): Synthesis and evaluation of rhamnose-capped prodrugs. J. Drug Target. 2010, 18, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Tomsik, P.; Stoklasova, A.; Micuda, S.; Niang, M.; Suba, P.; Knizek, J.; Rezacova, M. Evaluation of the antineoplastic activity of L-rhamnose in vitro. A comparison with 2-deoxyglucose. Acta Med. 2008, 51, 113–119. [Google Scholar]
- Ma, Y.; Pan, F.; McNeil, M. Formation of dTDP-rhamnose is essential for growth of mycobacteria. J. Bacteriol. 2002, 184, 3392–3395. [Google Scholar] [CrossRef]
- Xiao, G.; Alphey, M.S.; Tran, F.; Pirrie, L.; Milbeo, P.; Zhou, Y.; Bickel, J.K.; Kempf, O.; Kempf, K.; Naismith, J.H.; et al. Next generation Glucose-1-phosphate thymidylyltransferase (RmlA) inhibitors: An extended SAR study to direct future design. Bioorg. Med. Chem. 2021, 50, 116477. [Google Scholar] [CrossRef]
- Alphey, M.S.; Pirrie, L.; Torrie, L.S.; Boulkeroua, W.A.; Gardiner, M.; Sarkar, A.; Maringer, M.; Oehlmann, W.; Brenk, R.; Scherman, M.S.; et al. Allosteric competitive inhibitors of the glucose-1-phosphate thymidylyltransferase (RmlA) from Pseudomonas aeruginosa. ACS Chem. Biol. 2013, 8, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Loranger, M.W.; Forget, S.M.; McCormick, N.E.; Syvitski, R.T.; Jakeman, D.L. Synthesis and evaluation of L-rhamnose 1C-phosphonates as nucleotidylyltransferase inhibitors. J. Org. Chem. 2013, 78, 9822–9833. [Google Scholar] [CrossRef]
- Kantardjieff, K.A.; Kim, C.Y.; Naranjo, C.; Waldo, G.S.; Lekin, T.; Segelke, B.W.; Zemla, A.; Park, M.S.; Terwilliger, T.C.; Rupp, B. Mycobacterium tuberculosis RmlC epimerase (Rv3465): A promising drug-target structure in the rhamnose pathway. Acta Crystallogr. D Biol. Crystallogr. 2004, 60 Pt 5, 895–902. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Chen, F.; Li, Y.; Wang, L.; Li, H.; Gu, G.; Li, E. Rhamnose-Containing Compounds: Biosynthesis and Applications. Molecules 2022, 27, 5315. https://doi.org/10.3390/molecules27165315
Li S, Chen F, Li Y, Wang L, Li H, Gu G, Li E. Rhamnose-Containing Compounds: Biosynthesis and Applications. Molecules. 2022; 27(16):5315. https://doi.org/10.3390/molecules27165315
Chicago/Turabian StyleLi, Siqiang, Fujia Chen, Yun Li, Lizhen Wang, Hongyan Li, Guofeng Gu, and Enzhong Li. 2022. "Rhamnose-Containing Compounds: Biosynthesis and Applications" Molecules 27, no. 16: 5315. https://doi.org/10.3390/molecules27165315
APA StyleLi, S., Chen, F., Li, Y., Wang, L., Li, H., Gu, G., & Li, E. (2022). Rhamnose-Containing Compounds: Biosynthesis and Applications. Molecules, 27(16), 5315. https://doi.org/10.3390/molecules27165315