
Hydroxylpropyl-β-cyclodextrin as Potential Excipient to Prevent Stress-Induced Aggregation in Liquid Protein Formulations

 ,
,
Abstract
:1. Introduction
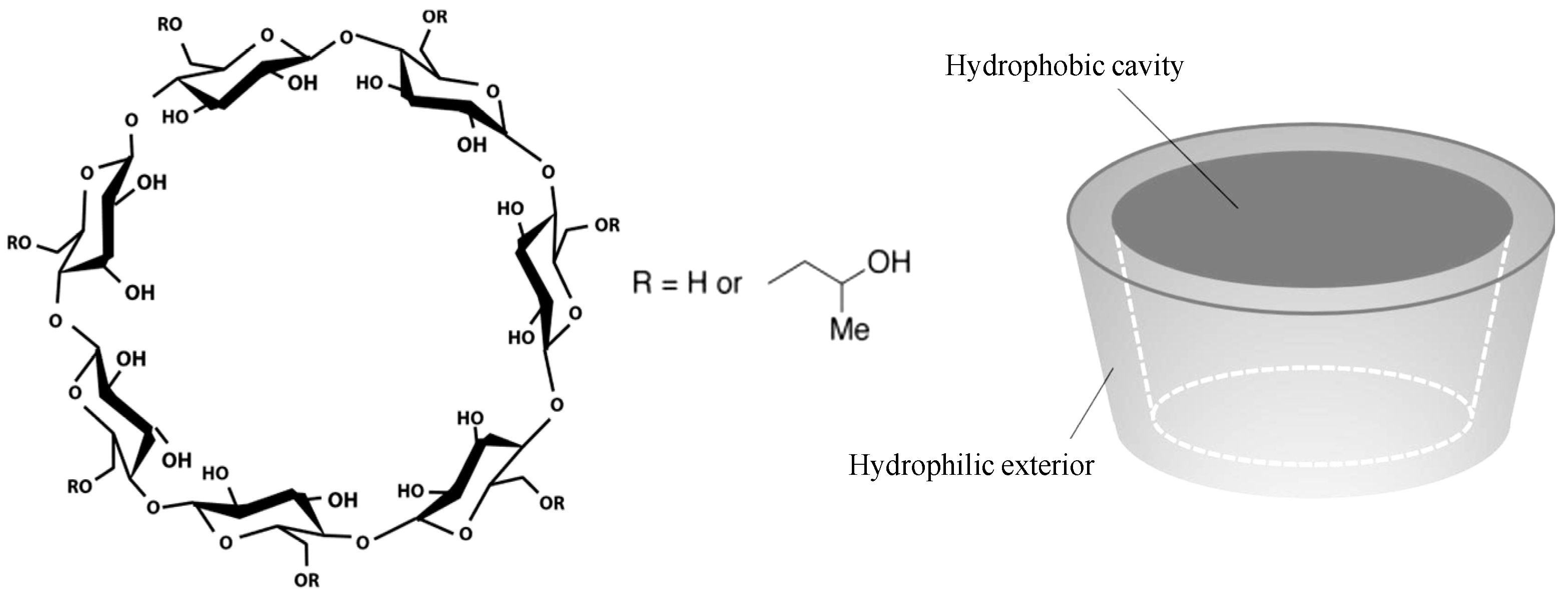
- (i)
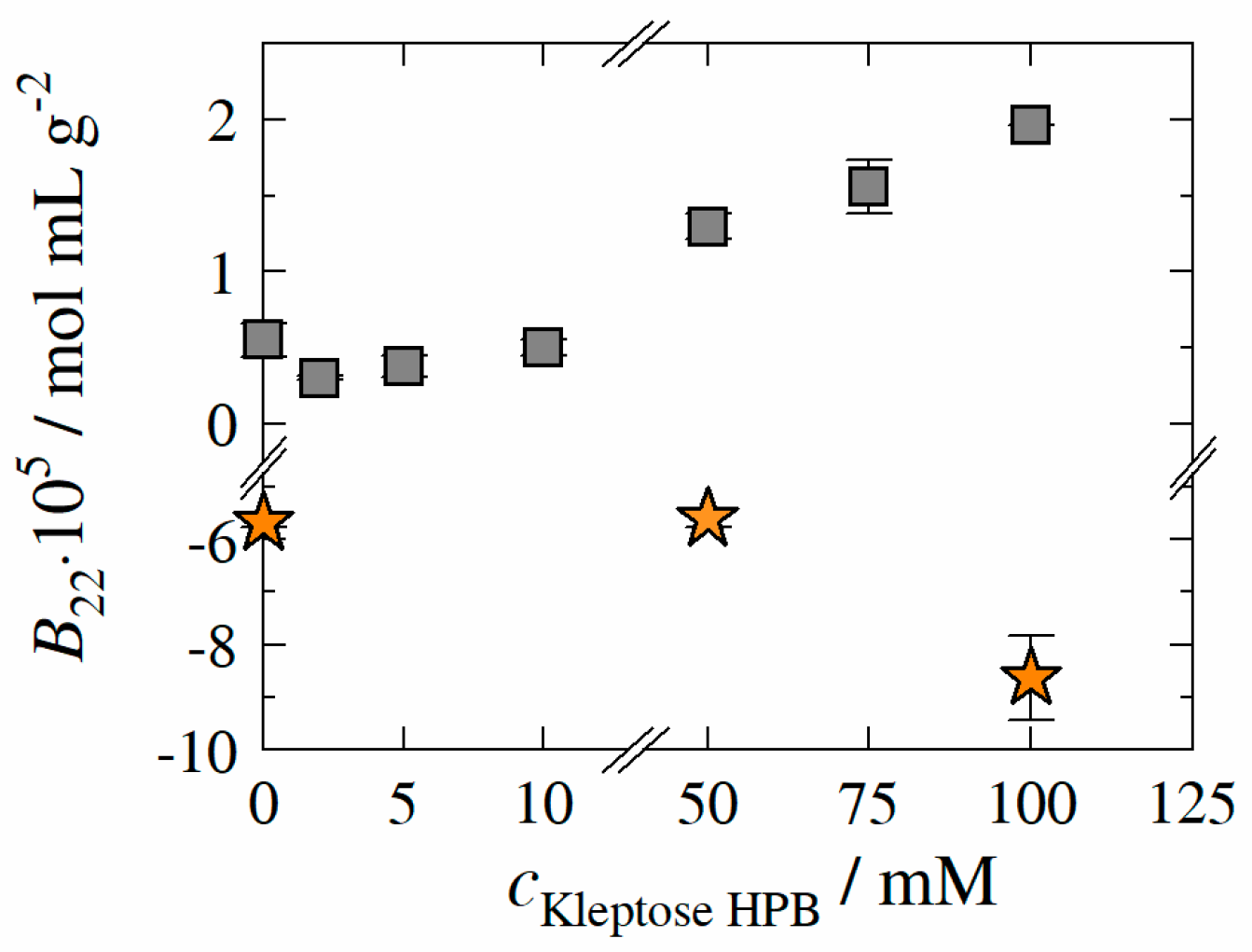
- HP-β-CD is expected to reduce the aggregation propensity of the protein due to a direct interaction/attachment with the protein. This hypothesis was later confirmed by Samra et al. [19] for three different proteins. Härtl et al. [20] further confirmed this stabilization effects (for two different immunoglobulin G type antibodies) by means of static and dynamic light scattering analytics (SLS/DLS—determination of second osmotic virial coefficient B22 or diffusion interaction parameter kD). According to their results, the interaction between HP-β-CD and IgG reduces the apparent hydrophobicity of the protein and thus decreases the adsorption tendency (increase in B22-values with increasing HP-β-CD concentration, indicating rise in net repulsive protein-protein interactions).
- (ii)
- HP-β-CD is expected to reduce the surface tension at the liquid/air interface resulting in a decrease in interface-related instability of IgG (in HCPF). Studies by Sá Couto et al. [21] and Loftsson [22] have already shown a decrease in surface tension due to the presence of HP-β-CD in aqueous solutions. However, the potential of HP-β-CD serving as a surfactant agent and thus stabilizer within HCPF is limited, as shown by the studies according to Serno et al. [18,23,24]. Serno et al. [24] investigated the surface activities of HPB and PS80 by dynamic surface tension analysis and drop profile analysis and identified a lower influence of HP-β-CD (~68 mN m−1—0.35% w/v) on the surface tension than for PS80 (~52 mN m−1—0.004% w/v). Thus, HP-β-CD cannot reduce the IgG aggregation at the liquid/air interface to the same extent as other non-ionic surfactants [24].
- The aggregation propensity as function of Kleptose®HPB (Roquette) in the absence and presence of a model mAb, that is -globulins from human blood (HB-IgG-A) [hydrodynamic radius rh using dynamic light scattering method (DLS)],
- The concentration-dependent phase behavior of Kleptose®HPB in formulations containing HB-IgG-A (surface tension using pendant drop method),
- The individual influence of Kleptose®HPB on (pre-existing) mAb aggregates, monomers and fragments [mass fractions within formulations using combination of size-exclusion chromatography and multi-angle light scattering (SEC-MALS)],
- The influence of Kleptose®HPB on the conformational and colloidal stability of the two different proteins HB-IgG-A and adalimumab (unfolding temperature Tunfold and second osmotic virial coefficient B22 using nano differential scanning fluorimetry (nanoDSF) and static light scattering method (SLS), respectively).
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Dynamic Light Scattering
2.2.2. Pendant Drop
2.2.3. Nano Differential Scanning Fluorimetry
2.2.4. Composition-Gradient Multi-Angle Light Scattering
2.2.5. Combination of Size-Exclusion Chromatography and Multi-Angle Light Scattering
2.2.6. Long-Term Samples and Storage Conditions
3. Results
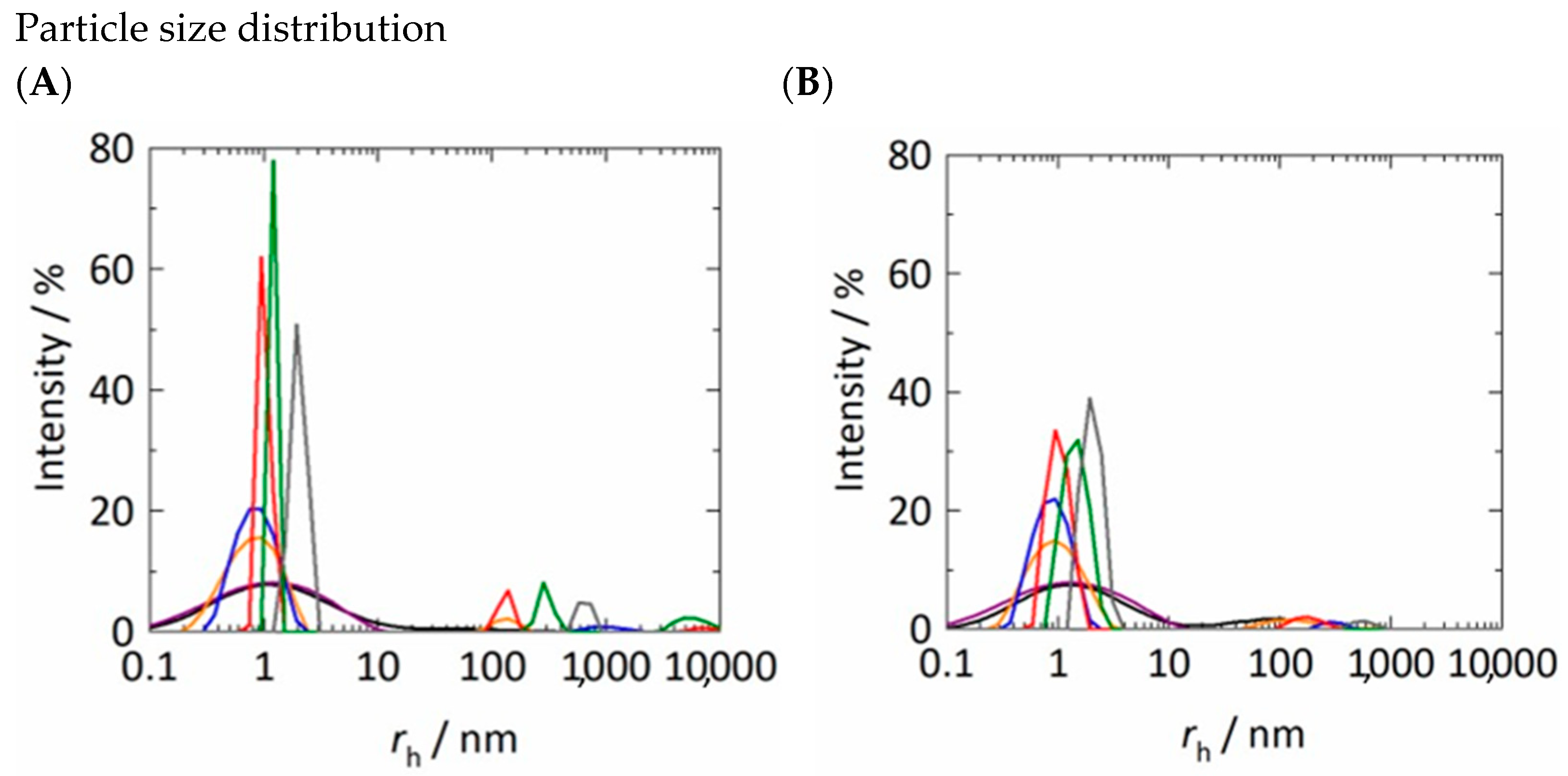
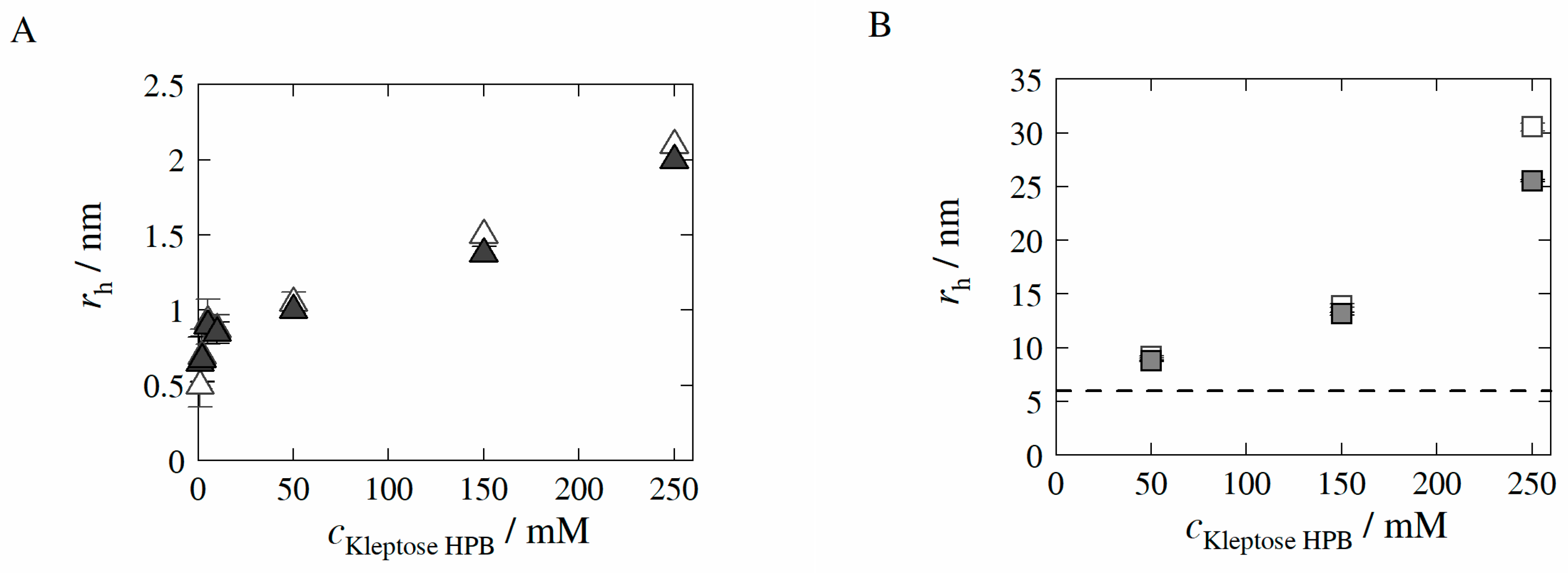
3.1. Hydrodynamic Radii
3.2. Surface Tension
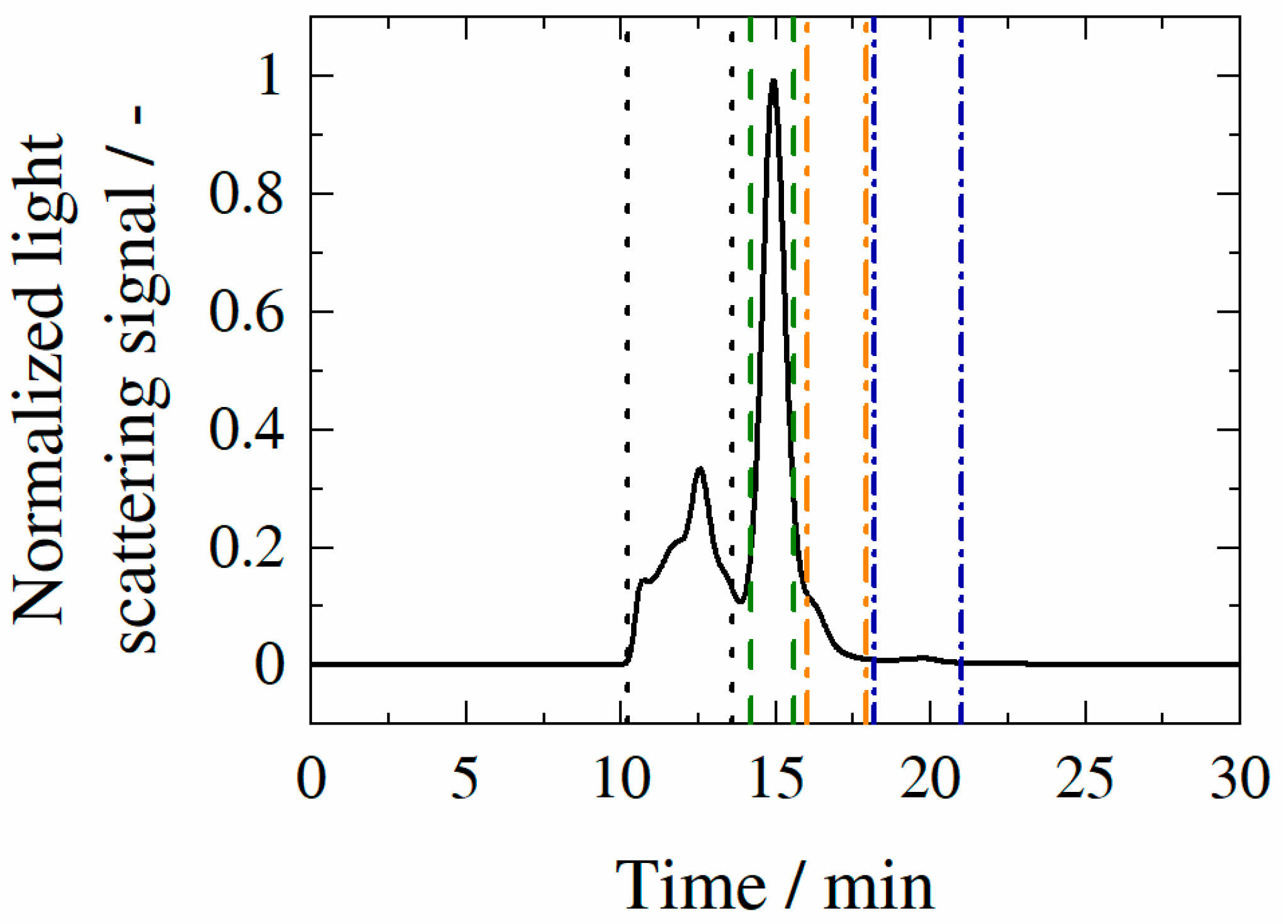
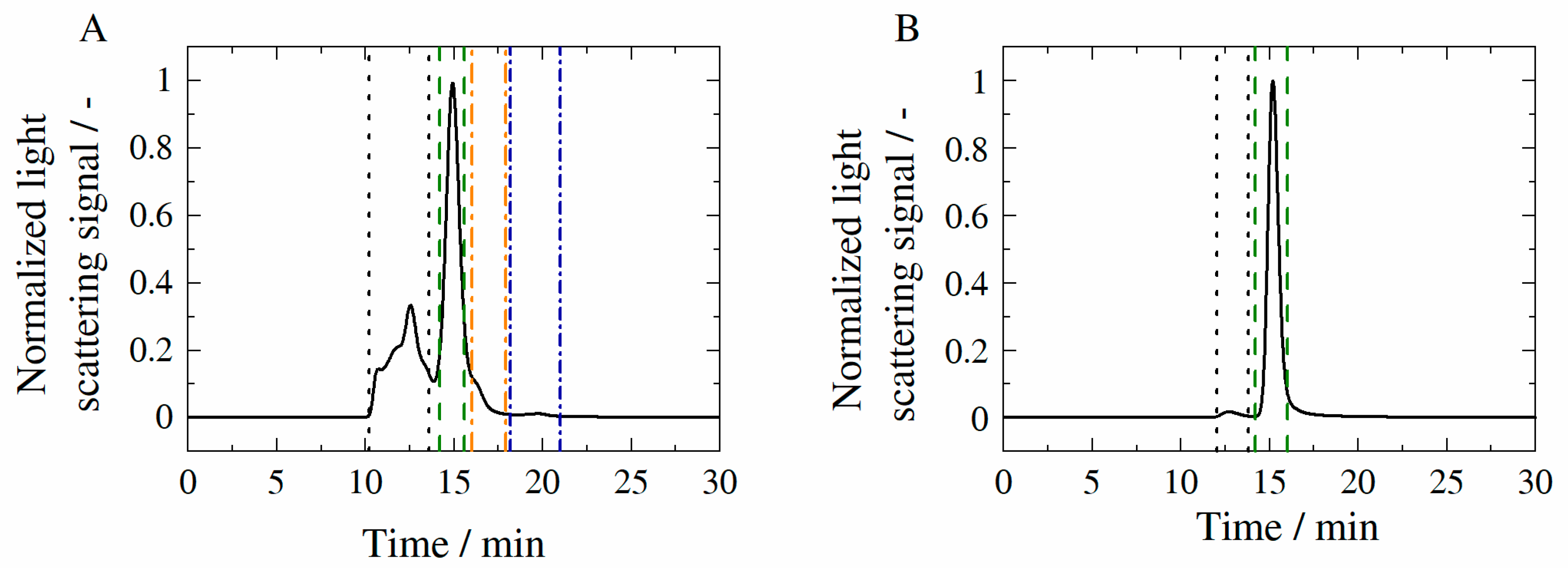
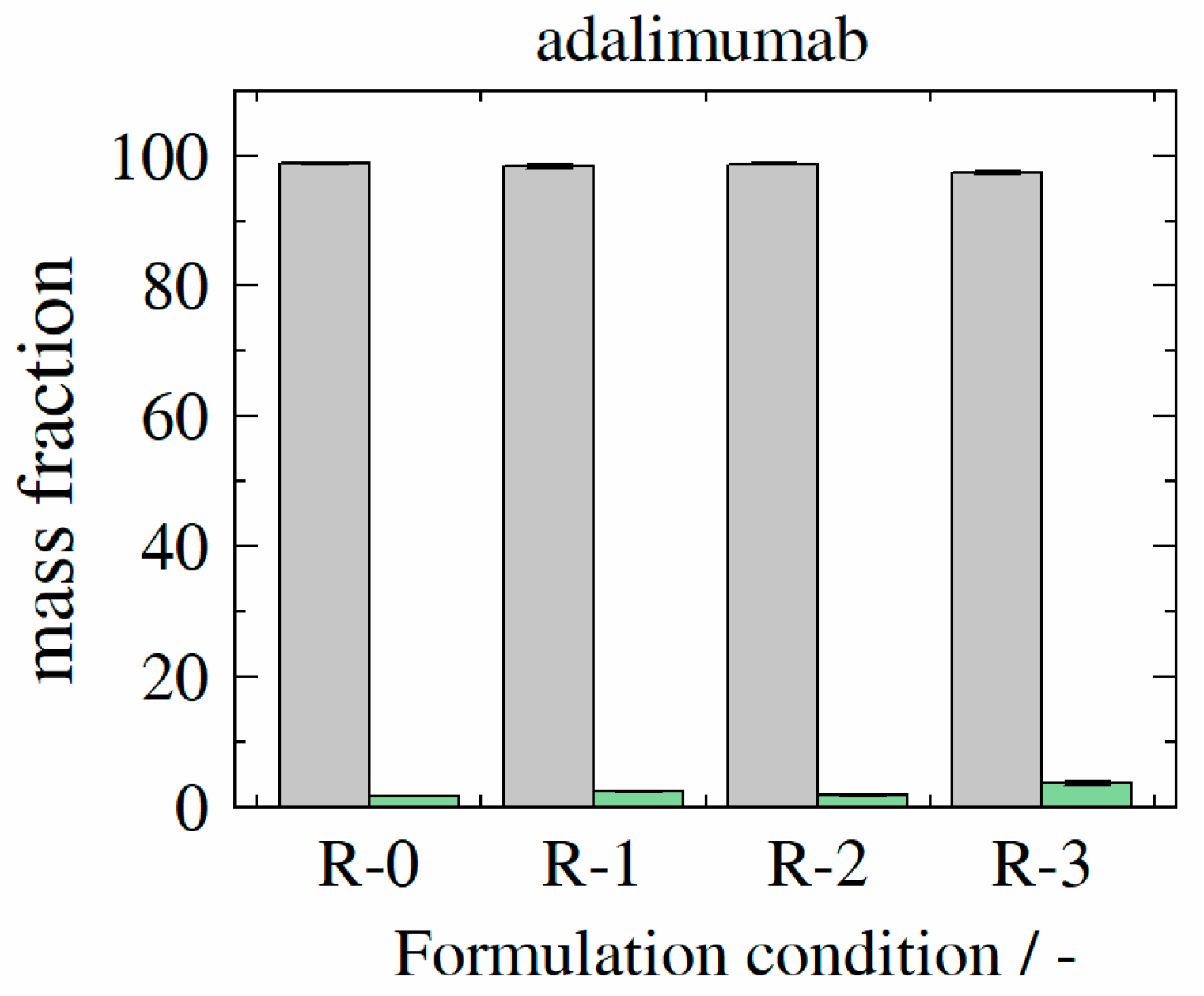
3.3. Aggregate Formation
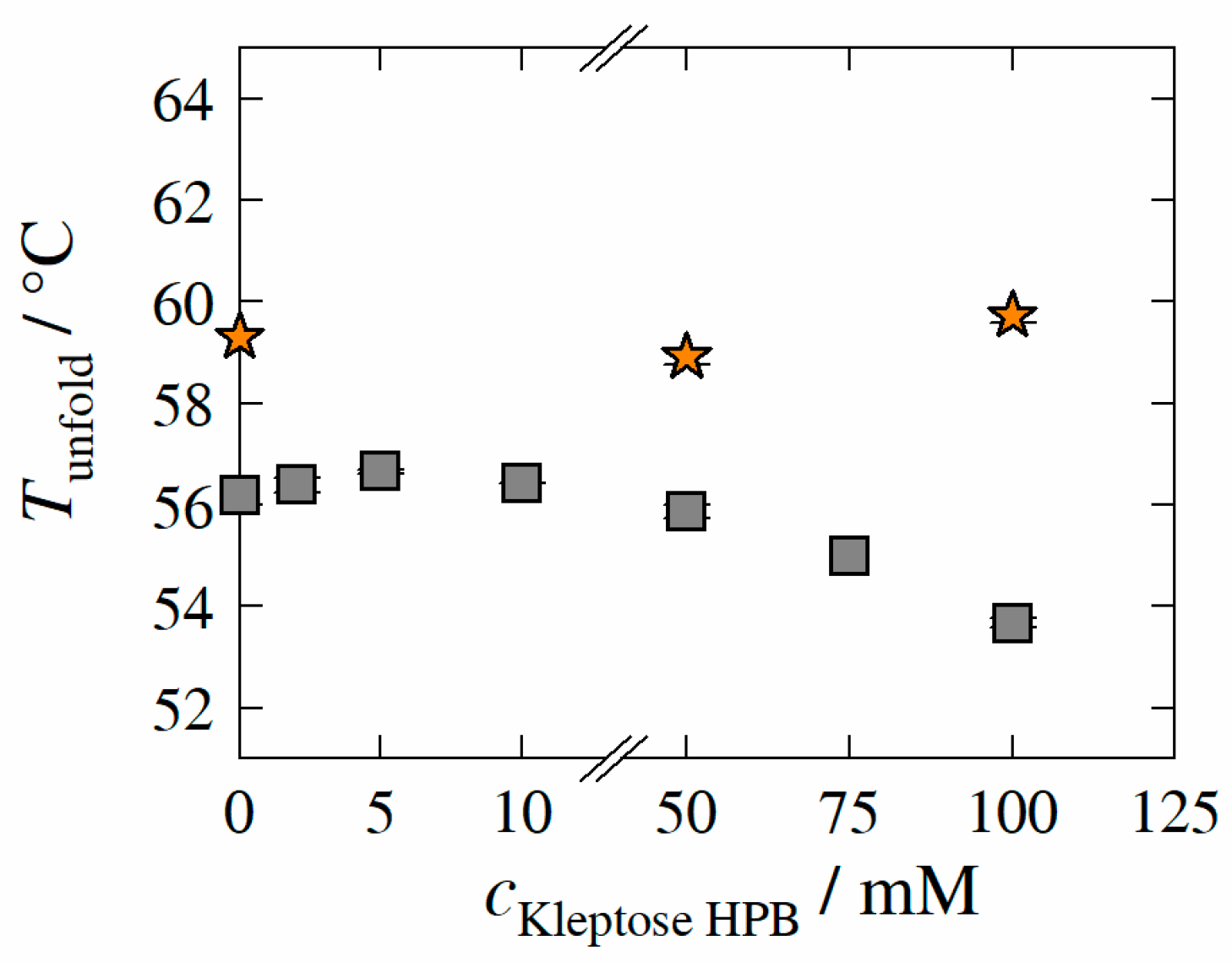
3.4. Unfolding Temperature
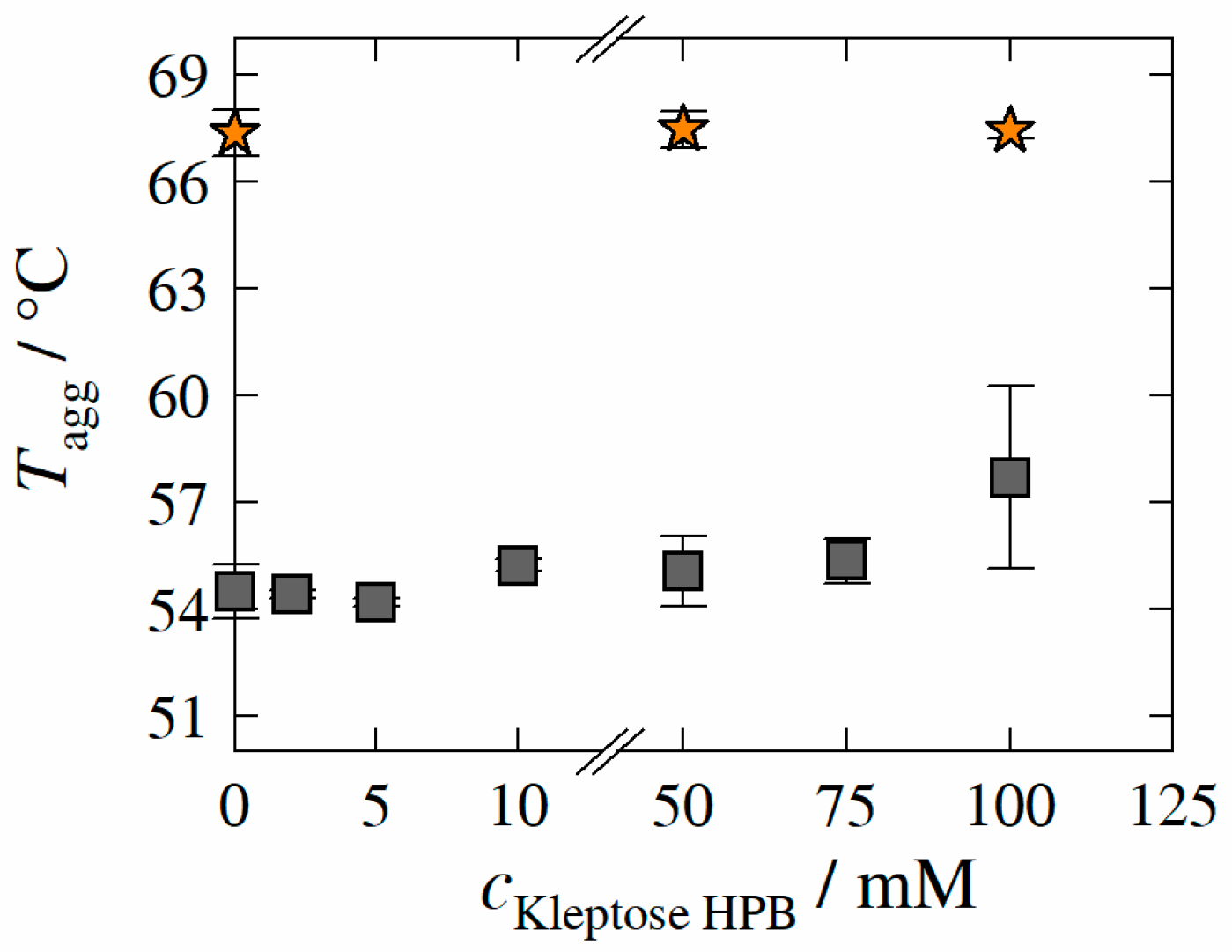
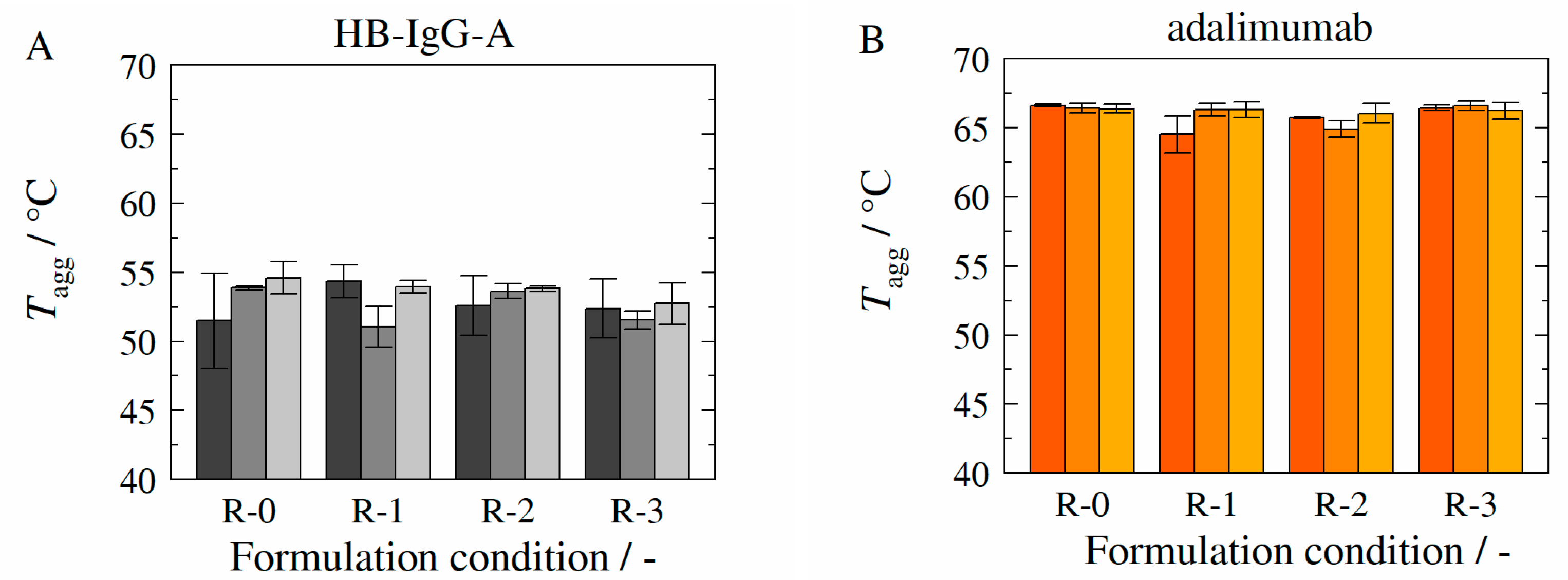
3.5. Aggregation Temperature
3.6. Protein–Protein Interactions
3.7. Formulation Stability (Adalimumab/HB-IgG-A) upon Storage at 4 °C
3.8. Formulation Stability (Adalimumab/HB-IgG-A) upon Storage at 40 °C
4. Discussion
4.1. Auto-Association and Aggregation Propensity of Kleptose®HPB
4.2. Surface Activity Induced by Kleptose®HPB
4.3. Interaction Characteristics between Kleptose®HPB and Proteins (HB-IgG-A & Adalimumab)
- Kleptose®HPB concentration (bellow/above );
- Protein type and protein concentration;
- Quality of protein in solution (aggregate fraction/amount & fragment fraction).
4.4. Potential of Kleptose®HPB as Excipient (Long-Term Stability Studies)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | /mM | /mg mL−1 | (25 °C) /kg m−3 |
|---|---|---|---|
| A | 0 | 30.0 ± 0.4 | 1010.6 ± 0.2 |
| B | 4.9 ± 0.2 | 29.8 ± 0.3 | 1012.4 ± 0.6 |
| C | 9.9 ± 0.3 | 30.4 ± 0.3 | 1014.6 ± 0.5 |
| D | 5.1 ± 0.3 | - | 1011.2 ± 0.6 |
| E | 9.8 ± 0.1 | - | 1011.3 ± 0.5 |
/mmol kg−1 | (25 °C) /mPa s |
|---|---|
| 0.0542 | 1.1446 ± 0.0004 |
| 0.1150 | 1.5066 ± 0.0007 |
| 0.1796 | 2.0760 ± 0.0001 |
| 0.2556 | 2.9833 ± 0.0008 |
| 0.3381 | 4.4709 ± 0.0011 |
| System | /mM | /mg mL−1 | /Da |
|---|---|---|---|
| unfiltered | 50 | - | 1452.4 ± 4.3 |
| 150 | - | 1373.7 ± 8.6 | |
| 250 | - | 1488.1 ± 148.7 | |
| filtered | 50 | - | 1443.2 ± 42.4 |
| 150 | - | 1055.6 ± 3.7 | |
| 250 | - | 927.7 ± 13.3 | |
| unfiltered | 50 | 30 | 158,402.5 ± 2233.8 |
| 150 | 30 | 150,824.2 ± 2517.1 | |
| 250 | 30 | 163,823.2 ± 12,474.5 | |
| filtered | 50 | 30 | 155,512.7 ± 105.7 |
| 150 | 30 | 134,833.6 ± 204.0 | |
| 250 | 30 | 135,965.1 ± 90.3 |
| Mass Fraction/% | |||||
|---|---|---|---|---|---|
| Adalimumab- | |||||
| Weeks | R-0 | R-1 | R-2 | R-3 | |
| Aggregates | 0 | 0.10 ± 0.02 | 0.60 ± 0.28 | 0.30 ± 0.09 | 0.45 ± 0.07 |
| 4 | 0.65 ± 0.21 | 0.90 ± 0.14 | 0.75 ± 0.07 | 0.80 ± 0.28 | |
| 32 | 1.25 ± 0.22 | 1.20 ± 0.24 | 1.45 ± 0.49 | 1.20 ± 0.71 | |
| Monomer | 0 | 99.05 ± 1.06 | 98.85 ± 0.64 | 99.40 ± 0.71 | 98.85 ± 0.21 |
| 4 | 98.30 ± 0.14 | 97.95 ± 0.35 | 98.20 ± 0.28 | 98.10 ± 0.57 | |
| 32 | 97.90 ± 0.28 | 97.80 ± 0.24 | 97.35 ± 0.78 | 97.8 ± 0.85 | |
| /kDa | |||||
|---|---|---|---|---|---|
| Adalimumab- | |||||
| Weeks | R-0 | R-1 | R-2 | R-3 | |
| Aggregates | 0 | 587.5 ± 4.8 | 443.6 ± 84.3 | 508.4 ± 5.3 | 464.0 ± 6.8 |
| 4 | 453.6 ± 45.8 | 391.7 ± 3.3 | 422.2 ± 23.7 | 404.4 ± 39.0 | |
| 32 | 537.8 ± 2.3 | 666.8 ± 43.0 | 468.2 ± 8.3 | 655.9 ± 71.2 | |
| Monomer | 0 | 148.8 ± 0.1 | 147.8 ± 0.1 | 147.6 ± 0.1 | 147.4 ± 0.1 |
| 4 | 147.5 ± 0.3 | 146.7 ± 0.3 | 146.8 ± 0.2 | 146.6 ± 0.3 | |
| 32 | 151.7 ± 0.8 | 150.9 ± 1.1 | 149.85 ± 1.8 | 151.3 ± 0.4 | |
| Mass Fraction/% | |||||
|---|---|---|---|---|---|
| HB-IgG-A | |||||
| Weeks | R-0 | R-1 | R-2 | R-3 | |
| Aggregates | 0 | 25.35 ± 0.92 | 22.70 ± 0.14 | 21.85 ± 0.21 | 21.90 ± 0.14 |
| 4 | 23.45 ± 0.09 | 20.95 ± 0.07 | 21.55 ± 0.22 | 20.50 ± 0.28 | |
| 24 | 18.05 ± 0.21 | 20.30 ± 0.24 | 16.20 ± 0.09 | 18.45 ± 0.21 | |
| Monomer | 0 | 61.30 ± 0.14 | 64.10 ± 0.28 | 65.15 ± 0.07 | 64.60 ± 0.99 |
| 4 | 54.95 ± 2.83 | 62.75 ± 0.17 | 65.60 ± 4.67 | 63.20 ± 0.28 | |
| 24 | 41.75 ± 0.08 | 53.20 ± 0.28 | 50.70 ± 0.28 | 53.15 ± 0.17 | |
| Fragment (1) | 0 | 7.90 ± 0.28 | 7.50 ± 0.14 | 7.60 ± 0.14 | 7.50 ± 0.14 |
| 4 | 10.30 ± 0.42 | 9.20 ± 0.15 | 9.30 ± 0.13 | 9.40 ± 0.13 | |
| 24 | 14.65 ± 0.07 | 12.50 ± 0.23 | 14.20 ± 0.17 | 13.15 ± 0.07 | |
| Fragment (2) | 0 | 2.75 ± 0.92 | 2.85 ± 0.21 | 2.70 ± 0.13 | 3.15 ± 0.49 |
| 4 | 6.15 ± 1.48 | 4.10 ± 0.16 | 2.80 ± 2.26 | 3.95 ± 0.11 | |
| 24 | 20.95 ± 0.07 | 9.75 ± 0.21 | 14.55 ± 0.21 | 11.05 ± 0.07 | |
| /kDa | |||||
|---|---|---|---|---|---|
| HB-IgG-A | |||||
| Weeks | R-0 | R-1 | R-2 | R-3 | |
| Aggregates | 0 | 399.2 ± 4.2 | 386.5 ± 3.3 | 390.6 ± 7.5 | 384.3 ± 4.1 |
| 4 | 378.1 ± 8.3 | 386.9 ± 2.8 | 374.6 ± 22.8 | 374.4 ± 1.6 | |
| 24 | 419.5 ± 1.27 | 413.5 ± 12.3 | 381.9 ± 2.3 | 387.5 ± 1.2 | |
| Monomer | 0 | 152.1 ± 0.8 | 151.5 ± 1.2 | 151.8 ± 0.2 | 151.8 ± 0.1 |
| 4 | 156.2 ± 2.8 | 153.0 ± 1.1 | 152.9 ± 3.9 | 152.5 ± 0.5 | |
| 24 | 151.0 ± 0.5 | 150.8 ± 0.1 | 151.6 ± 0.2 | 150.7 ± 0.2 | |
| Fragments (1) | 0 | 124.6 ± 3.3 | 127.7 ± 0.1 | 127.5 ± 1.0 | 128.1 ± 2.7 |
| 4 | 125.3 ± 6.2 | 126.2 ± 0.1 | 121.2 ± 0.8 | 122.6 ± 1.8 | |
| 24 | 111.2 ± 0.5 | 114.7 ± 0.7 | 113.9 ± 1.0 | 114.6 ± 1.3 | |
| Fragments (2) | 0 | 117.1 ± 47.8 | 99.0 ± 0.6 | 103.9 ± 2.1 | 86.2 ± 9.0 |
| 4 | 84.0 ± 35.2 | 86.4 ± 4.3 | 88.1 ± 1.6 | 86.0 ± 1.3 | |
| 24 | 54.35 ± 0.8 | 59.1 ± 1.0 | 56.6 ± 2.1 | 58.8 ± 2.1 | |

References
- BCC Publishing. Antibody Drugs: Technologies and Global Markets. 2021. Available online: https://www.bccresearch.com/market-research/biotechnology/antibody-drugs-technologies-global-markets-report.html (accessed on 25 May 2022).
- Viola, M.; Sequeira, J.; Seiça, R.; Veiga, F.; Serra, J.; Santos, A.C.; Ribeiro, A.J. Subcutaneous delivery of monoclonal antibodies: How do we get there? J. Control. Release 2018, 286, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Bittner, B.; Richter, W.; Schmidt, J. Subcutaneous Administration of Biotherapeutics: An Overview of Current Challenges and Opportunities. BioDrugs 2018, 32, 425–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Loftsson, T.; Brewster, M.E. Pharmaceutical applications of cyclodextrins: Basic science and product development. J. Pharm. Pharmacol. 2010, 62, 1607–1621. [Google Scholar] [CrossRef]
- Jambhekar, S.S.; Breen, P. Cyclodextrins in pharmaceutical formulations I: Structure and physico-chemical properties, formation of complexes, and types of complex. Drug Discov. Today 2016, 21, 356–362. [Google Scholar] [CrossRef]
- Häusler, O.; Müller-Goymann, C.C. Properties and Structure of Aqueous Solutions of Hydroxypropyl-beta-Cyclodextrin. Starch 1993, 45, 183–187. [Google Scholar] [CrossRef]
- Loftsson, T.; Jarho, P.; Másson, M.; Järvinen, T. Cyclodextrins in drug delivery. Expert Opin. Drug Deliv. 2005, 2, 335–351. [Google Scholar] [CrossRef]
- Gidwani, B.; Vyas, A. A Comprehensive Review on Cyclodextrin-Based Carriers for Delivery of Chemotherapeutic Cytotoxic Anticancer Drugs. Biomed. Res. Int. 2015, 2015, 198268. [Google Scholar] [CrossRef] [Green Version]
- Werner, M.B.W.; Brauns, U. Pharmaceutical Compositions Containing Drugs Which Are Instable or Sparingly Soluble in Water and Methods for Their Preparation. U.S. Patent No. 6,407,079, 18 June 2002. [Google Scholar]
- Pitha, J. Pharmaceutical Preparations Containing Cyclodextrin Derivatives. U.S. Patent No. 4,727,064, 29 May 1985. [Google Scholar]
- Cesari, A.; Uccello Barretta, G.; Kirschner, K.N.; Pappalardo, M.; Basile, L.; Guccione, S.; Russotto, C.; Lauro, M.R.; Cavaliere, F.; Balzano, F. Interaction of natural flavonoid eriocitrin with β-cyclodextrin and hydroxypropyl-β-cyclodextrin: An NMR and molecular dynamics investigation. New J. Chem. 2020, 44, 16431–16441. [Google Scholar] [CrossRef]
- Wang, Z.; Landy, D.; Sizun, C.; Cézard, C.; Solgadi, A.; Przybylski, C.; de Chaisemartin, L.; Herfindal, L.; Barratt, G.; Legrand, F.-X. Cyclodextrin complexation studies as the first step for repurposing of chlorpromazine. Int. J. Pharm. 2020, 584, 119391. [Google Scholar] [CrossRef]
- Jiang, Y.; Jiang, X.; Law, K.; Chen, Y.; Gu, J.; Zhang, W.; Xin, H.; Sha, X.; Fang, X. Enhanced anti-tumor effect of 9-nitro-camptothecin complexed by hydroxypropyl-β-cyclodextrin and safety eval-uation. Int. J. Pharm. 2011, 415, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Lahiani-Skiba, M.; Bounoure, F.; Fessi, H.; Skiba, M. Effect of cyclodextrins on lonidamine release and in-vitro cytotoxicity. J. Incl. Phenom. Macrocycl. Chem. 2011, 69, 481–485. [Google Scholar] [CrossRef]
- Wu, H.H.; Garidel, P.; Michaela, B. HP-β-CD for the formulation of IgG and Ig-based biotherapeutics. Int. J. Pharm. 2021, 601, 120531. [Google Scholar] [CrossRef]
- Gokhale, R. Hydroxylpropyl beta-cyclodextrin: A promising excipient for protein stabilization. Presented at the Drug Delivery & Formulation Summit, Berlin, Germany, 11–13 March 2019. [Google Scholar]
- Serno, T.; Geidobler, R.; Winter, G. Protein stabilization by cyclodextrins in the liquid and dried state. Adv. Drug Deliv. Rev. 2011, 63, 1086–1106. [Google Scholar] [CrossRef] [PubMed]
- Samra, H.S.; He, F.; Bhambhani, A.; Pipkin, J.D.; Zimmerer, R.; Joshi, S.B.; Middaugh, C.R. The effects of substituted cyclodextrins on the colloidal and conformational stability of selected proteins. J. Pharm. Sci. 2010, 99, 2800–2818. [Google Scholar] [CrossRef] [PubMed]
- Härtl, E.; Dixit, N.; Besheer, A.; Kalonia, D.; Winter, G. Weak antibody-cyclodextrin interactions determined by quartz crystal microbalance and dynamic/static light scattering. Eur. J. Pharm. Biopharm. 2013, 85, 781–789. [Google Scholar] [CrossRef]
- Sá Couto, A.R.; Ryzhakov, A.; Loftsson, T. 2-Hydroxypropyl-β-Cyclodextrin Aggregates: Identification and Development of Analytical Techniques. Materials 2018, 11, 1971. [Google Scholar] [CrossRef] [Green Version]
- Loftsson, T. Drug permeation through biomembranes: Cyclodextrins and the unstirred water layer. Pharmazie 2012, 67, 363–370. [Google Scholar]
- Serno, T.; Carpenter, J.F.; Randolph, T.W.; Winter, G. Inhibition of agitation-induced aggregation of an IgG-antibody by hydroxypropyl-beta-cyclodextrin. J. Pharm. Sci. 2010, 99, 1193–1206. [Google Scholar] [CrossRef]
- Serno, T.; Härtl, E.; Besheer, A.; Miller, R.; Winter, G. The role of polysorbate 80 and HPβCD at the air-water interface of IgG solutions. Pharm. Res. 2013, 30, 117–130. [Google Scholar] [CrossRef]
- Podzimek, S. Light Scattering, Size Exclusion Chromatography and Asymmetric Flow Field Flow Fractionation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Wyatt Technology Corporation. Dynamic Light Scattering (DLS): User Manual, Santa Barbara (CA, USA). 2017. Available online: http://www.wyatt.com/files/events/Solutions-Guide-LR.pdf (accessed on 15 May 2022).
- Rotenberg, Y.; Boruvka, L.; Neumann, A. Determination of surface tension and contact angle from the shapes of axisymmetric fluid interfaces. J. Colloid Interface Sci. 1983, 93, 169–183. [Google Scholar] [CrossRef]
- Gonalves, F.A.; Kestin, J.; Sengers, J.V. Surface-tension effects in suspended-level capillary viscometers. Int. J. Thermophys. 1991, 12, 1013–1028. [Google Scholar] [CrossRef]
- McClure, S.M.; Ahl, P.L.; Blue, J.T. High Throughput Differential Scanning Fluorimetry (DSF) Formulation Screening with Complementary Dyes to Assess Protein Unfolding and Aggregation in Presence of Surfactants. Pharm. Res. 2018, 35, 81. [Google Scholar] [CrossRef] [PubMed]
- Garidel, P.; Hegyi, M.; Bassarab, S.; Weichel, M. A rapid, sensitive and economical assessment of monoclonal antibody conformational stability by intrinsic tryptophan fluorescence spectroscopy. Bio-Technol. J. 2008, 3, 1201–1211. [Google Scholar] [CrossRef]
- Chattopadhyay, G.; Varadarajan, R. Facile measurement of protein stability and folding kinetics using a nano differential scanning fluorimeter. Protein Sci. 2019, 28, 1127–1134. [Google Scholar] [CrossRef]
- Malik, K.; Matejtschuk, P.; Thelwell, C.; Burns, C.J. Differential scanning fluorimetry: Rapid screening of formulations that promote the stability of reference preparations. J. Pharm. Biomed. Anal. 2013, 77, 163–166. [Google Scholar] [CrossRef]
- Boland, C.; Olatunji, S.; Bailey, J.; Howe, N.; Weichert, D.; Fetics, S.K.; Yu, X.; Merino-Gracia, J.; Delsaut, C.; Caffrey, M. Membrane (and Soluble) Protein Stability and Binding Measurements in the Lipid Cubic Phase Using Label-Free Differential Scanning Fluorimetry. Anal. Chem. 2018, 90, 12152–12160. [Google Scholar] [CrossRef]
- Kim, S.H.; Yoo, H.J.; Park, E.J.; Na, D.H. Nano Differential Scanning Fluorimetry-Based Thermal Stability Screening and Optimal Buffer Selection for Immunoglobulin G. Pharmaceuticals 2021, 15, 29. [Google Scholar] [CrossRef]
- Garidel, P. Right angle light scattering protein thermostability screening: From research to development. Spectrosc. Eur. 2012, 24, 13–18. [Google Scholar]
- Garidel, P.; Karow, A.R.; Blech, M. Orthogonal spectroscopic techniques for the early developability assessment of therapeutic protein candidates. Spectrosc. Eur. 2014, 26, 9–13. [Google Scholar]
- Pagano, B.; Iaccarino, N.; Di Porzio, A.; Randazzo, A.; Amato, J. Screening of DNA G-quadruplex stabilizing ligands by nano differential scanning fluorimetry. Analyst 2019, 144, 6512–6516. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Maschberger, M.; Breitsprecher, D. Thermal Unfolding of Antibodies with the Prometheus NT.48. BioTechniques 2015, 59, 251. [Google Scholar] [CrossRef] [Green Version]
- Blech, M.; Garidel, P. High Precision Thermal Stability and Aggregation Profiling by nanoDSF. Poster 2016. [Google Scholar] [CrossRef]
- Söltl, F.; Derix, J.; Garidel, P.; Blech, M.; Breitsprecher, D. Unfolding and Aggregation of mAbs: Analysis of Formulation-Dependent Colloidal and Conformational Stability of Monoclonal Antibodies. Application Note NT-PR-005. 2015. Available online: https://resources.nanotempertech.com/application-notes/application-note-nt-pr-005-mab-aggregation?locale=en_us (accessed on 15 May 2022).
- Gupta, I.; Dharadhar, S.; Sixma, T.; Khan, S. Biochemical characterization of TRIM72 E3 ligase and its interaction with the insulin receptor substrate 1. Biochem. Biophys. Rep. 2020, 21, 100729. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.S.; Gabriel, D.A. Laser Light Scattering; Dover: New York, NY, USA, 1994; ISBN 9780486683287. [Google Scholar]
- Schleinitz, M.; Teschner, D.; Sadowski, G.; Brandenbusch, C. Second osmotic virial coefficients of therapeutic proteins in the presence of excipient-mixtures can be predicted to aid an efficient formulation design. J. Mol. Liq. 2019, 283, 575–583. [Google Scholar] [CrossRef]
- Wessner, M.; Bommarius, B.; Brandenbusch, C.; Bommarius, A.S. Purification of chimeric amine dehydrogenase using a tailor-made aqueous two-phase system—A case study. J. Mol. Liq. 2021, 323, 114991. [Google Scholar] [CrossRef]
- Stolzke, T.; Brandenbusch, C. Simplified choice of suitable excipients within biologics formulation design using protein-protein interaction- and water activity-maps. Eur. J. Pharm. Biopharm. 2022, 176, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Kuzman, D.; Bunc, M.; Ravnik, M.; Reiter, F.; Žagar, L.; Bončina, M. Long-term stability predictions of therapeutic monoclonal antibodies in solution using Arrhenius-based kinetics. Sci. Rep. 2021, 11, 20534. [Google Scholar] [CrossRef]
- Thiagarajan, G.; Semple, A.; James, J.K.; Cheung, J.K.; Shameem, M. A comparison of biophysical characterization techniques in predicting monoclonal antibody stability. MAbs 2016, 8, 1088–1097. [Google Scholar] [CrossRef] [Green Version]
- Svilenov, H.; Winter, G. The ReFOLD assay for protein formulation studies and prediction of protein aggregation during long-term storage. Eur. J. Pharm. Biopharm. 2019, 137, 131–139. [Google Scholar] [CrossRef]
- Do, T.T.; van Hooghten, R.; van den Mooter, G. A study of the aggregation of cyclodextrins: Determination of the critical aggregation concentration, size of aggregates and thermodynamics using isodesmic and K2-K models. Int. J. Pharm. 2017, 521, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, V.; Kajwe, A.; Murahari, M.; Pujar, G.V.; Inturi, B.K.; Sherje, A.P. Inclusion Complexes of Nateglinide with HP-β-CD and L-Arginine for Solubility and Dissolution Enhancement: Preparation, Characterization, and Molecular Docking Study. J. Pharm. Innov. 2017, 12, 168–181. [Google Scholar] [CrossRef]
- Sherje, A.P.; Patel, F.; Murahari, M.; Suvarna, V.; Patel, K. Study on effect of l-arginine on solubility and dissolution of Zaltoprofen: Preparation and characterization of binary and ternary cyclodextrin inclusion complexes. Chem. Phys. Lett. 2018, 694, 120–128. [Google Scholar] [CrossRef]
- Lee, J.C.; Timasheff, S.N. The stabilization of proteins by sucrose. J. Biol. Chem. 1981, 256, 7193–7201. [Google Scholar] [CrossRef]
- Sapir, L.; Harries, D. Linking trehalose self-association with binary aqueous solution equation of state. J. Phys. Chem. B 2011, 115, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Winther, L.R.; Qvist, J.; Halle, B. Hydration and mobility of trehalose in aqueous solution. J. Phys. Chem. B 2012, 116, 9196–9207. [Google Scholar] [CrossRef]

| Component | CAS-No | Formula | Purity | Supplier |
|---|---|---|---|---|
| HB-IgG-A | 9007-83-4 | - | ≥99.0% | Sigma Aldrich, Schnelldorf, Germany |
| HB-IgG-B | NA (LOT#BS19201051) | - | 98.0% | BOC Science, Shirley, NY, USA |
| Adalimumab-solution | - | - | - | In-house preparation Roquette Asia Pacific Pte. Ltd., Singapore |
| Kleptose®HPB | 128446-35-5 | ≥97.0% | Roquette Freres, Lestrem, France | |
| l-arginine HCl | 1119-34-2 | C6H14N4O2·HCl | ≥98.0% | Sigma Aldrich, Schnelldorf, Germany |
| Dipotassium hydrogen phosphate | 7758-11-4 | K2HPO4 | ≥99.8% | BDH Prolabo, VWR, Langenfeld, Germany |
| Sodium dihydrogen phosphate | 7558-80-7 | NaH2PO4 | ≥98.0% | BDH Prolabo, VWR, Langenfeld, Germany |
| System | /mM | /mg mL−1 | /mN m−1 |
|---|---|---|---|
| Water | - | - | 71.97 [28] |
| Kleptose®HPB in K2HPO4-NaH2PO4 solution | 5.1 ± 0.3 | - | 61.49 ± 0.23 |
| 9.8 ± 0.1 | - | 60.27 ± 0.09 | |
| HB-IgG-A in K2HPO4-NaH2PO4 solution | - | 30.0 ± 0.4 | 55.15 ± 0.01 |
| HB-IgG-A + Kleptose®HPB in K2HPO4-NaH2PO4 solution | 4.9 ± 0.2 | 29.8 ± 0.3 | 54.86 ± 0.08 |
| 9.9 ± 0.3 | 30.4 ± 0.3 | 54.71 ± 0.09 |
| System | Aggregates /% | Monomer /% | Fragment (1) /% | Fragment (2) /% |
|---|---|---|---|---|
| HB-IgG-A | 20.3 ± 0.4 | 68.0 ± 0.4 | 5.8 ± 0.1 | 2.7 ± 0.1 |
| HB-IgG-A + 5 mM Kleptose®HPB | 20.2 ± 0.1 | 67.9 ± 1.1 | 5.7 ± 0.1 | 2.5 ± 0.6 |
| HB-IgG-A + 10 mM Kleptose®HPB | 20.9 ± 0.1 | 66.7 ± 0.1 | 5.7 ± 0.1 | 2.8 ± 0.1 |
| HB-IgG-A + 150 mM Kleptose®HPB | 18.8 ± 0.2 | 64.8 ± 0.2 | 4.9 ± 0.4 | 7.0 ± 0.5 |
| HB-IgG-A + 250 mM Kleptose®HPB | 17.5 ± 0.1 | 65.2 ± 0.3 | 5.1 ± 0.1 | 7.7 ± 0.1 |
| HB-IgG-B | 22.8 ± 0.03 | 76.1 ± 0.02 | 0.46 ± 0.005 | 0.58 ± 0.005 |
| HB-IgG-B + 5 mM Kleptose®HPB | 22.7 ± 0.07 | 76.3 ± 0.1 | 0.43 ± 0.02 | 0.56 ± 0.005 |
| HB-IgG-B + 10 mM Kleptose®HPB | 22.5 ± 0.03 | 76.5 ± 0.04 | 0.44 ± 0.025 | 0.57 ± 0.015 |
| HB-IgG-B + 150 mM Kleptose®HPB | 22.1 ± 0.03 | 76.8 ± 0.02 | 0.43 ± 0.025 | 0.58 ± 0.015 |
| HB-IgG-B + 250 mM Kleptose®HPB | 22.4 ± 0.11 | 76.58 ± 0.09 | 0.41 ± 0.015 | 0.55 ± 0.01 |
| System | Aggregates /kDa | Monomer /kDa | Fragments (1) /kDa | Fragments (2) /kDa |
|---|---|---|---|---|
| HB-IgG-A in lean buffer | 343.7 ± 1.6 | 148.9 ± 0.3 | 119.8 ± 0.2 | 80.5 ± 5.3 |
| HB-IgG-A + 5 mM Kleptose®HPB | 351.5 ± 11.2 | 148.9 ± 0.9 | 118.6 ± 4.0 | 77.9 ± 18.2 |
| HB-IgG-A + 10 mM Kleptose®HPB | 345.2 ± 1.3 | 148.2 ± 0.2 | 116.1 ± 0.2 | 65.6 ± 2.8 |
| HB-IgG-A + 150 mM Kleptose®HPB | 398.6 ± 1.9 | 153.5 ± 0.1 | 144.0 ± 0.5 | 39.6 ± 0.2 |
| HB-IgG-A + 250 mM Kleptose®HPB | 377.1 ± 1.8 | 152.8 ± 0.3 | 138.2 ± 2.1 | 43.0 ± 4.0 |
| Formulation Condition | /mmol kg−1 | /mmol kg−1 | /mmol kg−1 |
|---|---|---|---|
| R-0 | 0 | 0 | 0 |
| R-1 | 0 | 55 | 50 |
| R-2 | 55 | 0 | 50 |
| R-3 | 100 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stolzke, T.; Krieg, F.; Peng, T.; Zhang, H.; Häusler, O.; Brandenbusch, C. Hydroxylpropyl-β-cyclodextrin as Potential Excipient to Prevent Stress-Induced Aggregation in Liquid Protein Formulations. Molecules 2022, 27, 5094. https://doi.org/10.3390/molecules27165094
Stolzke T, Krieg F, Peng T, Zhang H, Häusler O, Brandenbusch C. Hydroxylpropyl-β-cyclodextrin as Potential Excipient to Prevent Stress-Induced Aggregation in Liquid Protein Formulations. Molecules. 2022; 27(16):5094. https://doi.org/10.3390/molecules27165094
Chicago/Turabian StyleStolzke, Tanja, Franziska Krieg, Tao Peng, Hailong Zhang, Olaf Häusler, and Christoph Brandenbusch. 2022. "Hydroxylpropyl-β-cyclodextrin as Potential Excipient to Prevent Stress-Induced Aggregation in Liquid Protein Formulations" Molecules 27, no. 16: 5094. https://doi.org/10.3390/molecules27165094
APA StyleStolzke, T., Krieg, F., Peng, T., Zhang, H., Häusler, O., & Brandenbusch, C. (2022). Hydroxylpropyl-β-cyclodextrin as Potential Excipient to Prevent Stress-Induced Aggregation in Liquid Protein Formulations. Molecules, 27(16), 5094. https://doi.org/10.3390/molecules27165094