
Lipase-Catalyzed Synthesis, Antioxidant Activity, Antimicrobial Properties and Molecular Docking Studies of Butyl Dihydrocaffeate

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
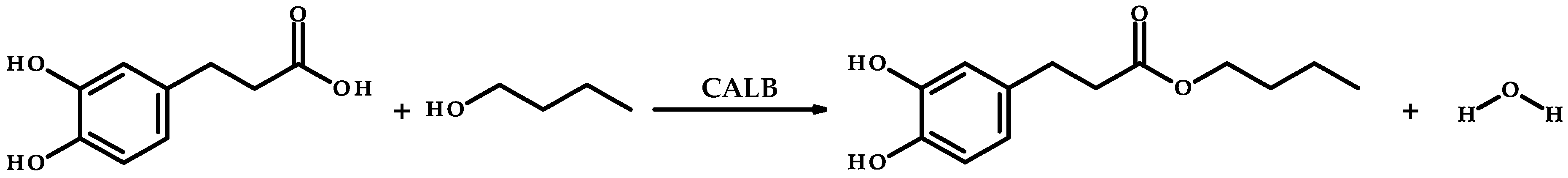
2.1. Enzymatic Synthesis and Evaluation of Antioxidant Activity of Butyl Dihydrocaffeate
2.2. Antimicrobial Properties of Butyl Dihydrocaffeate
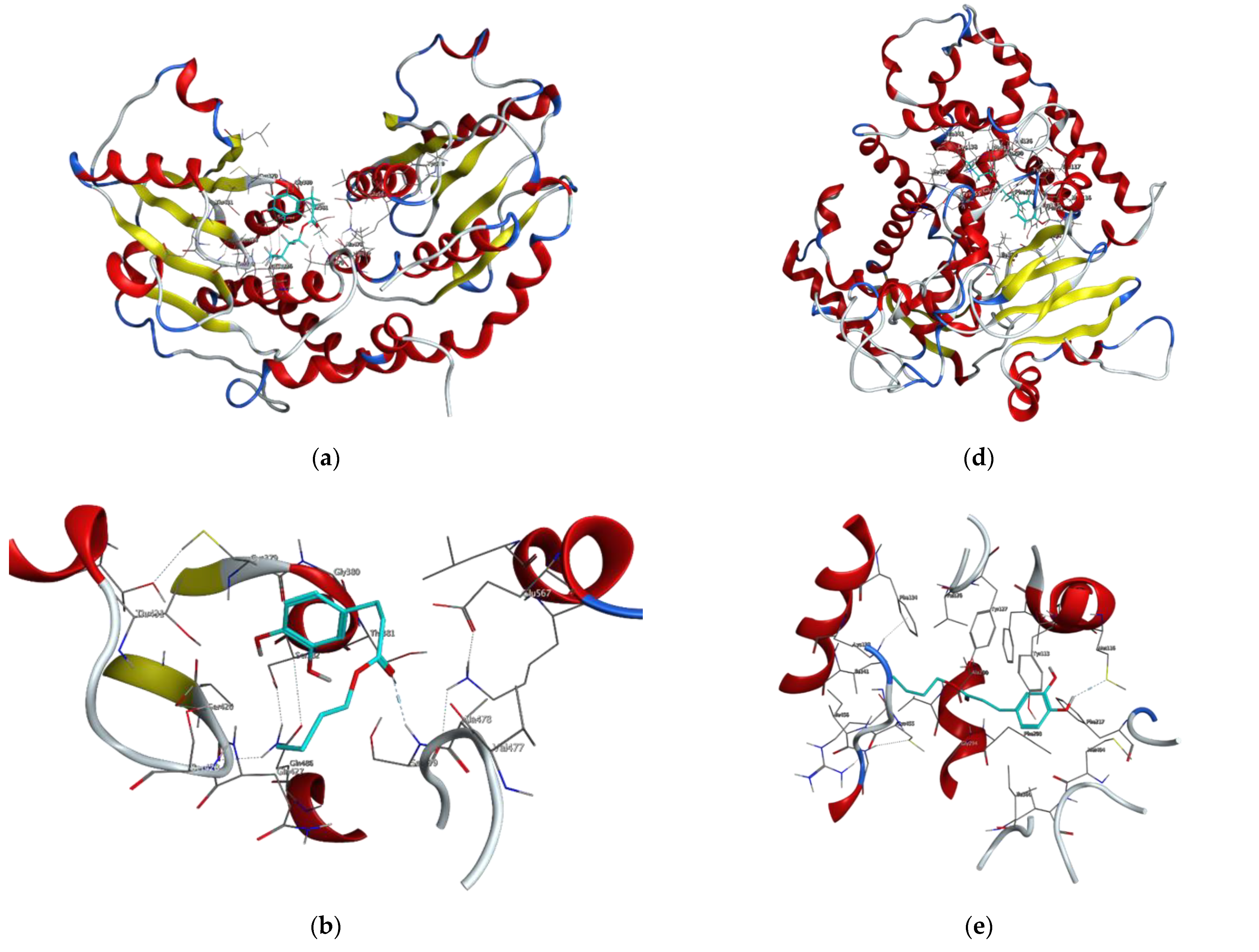
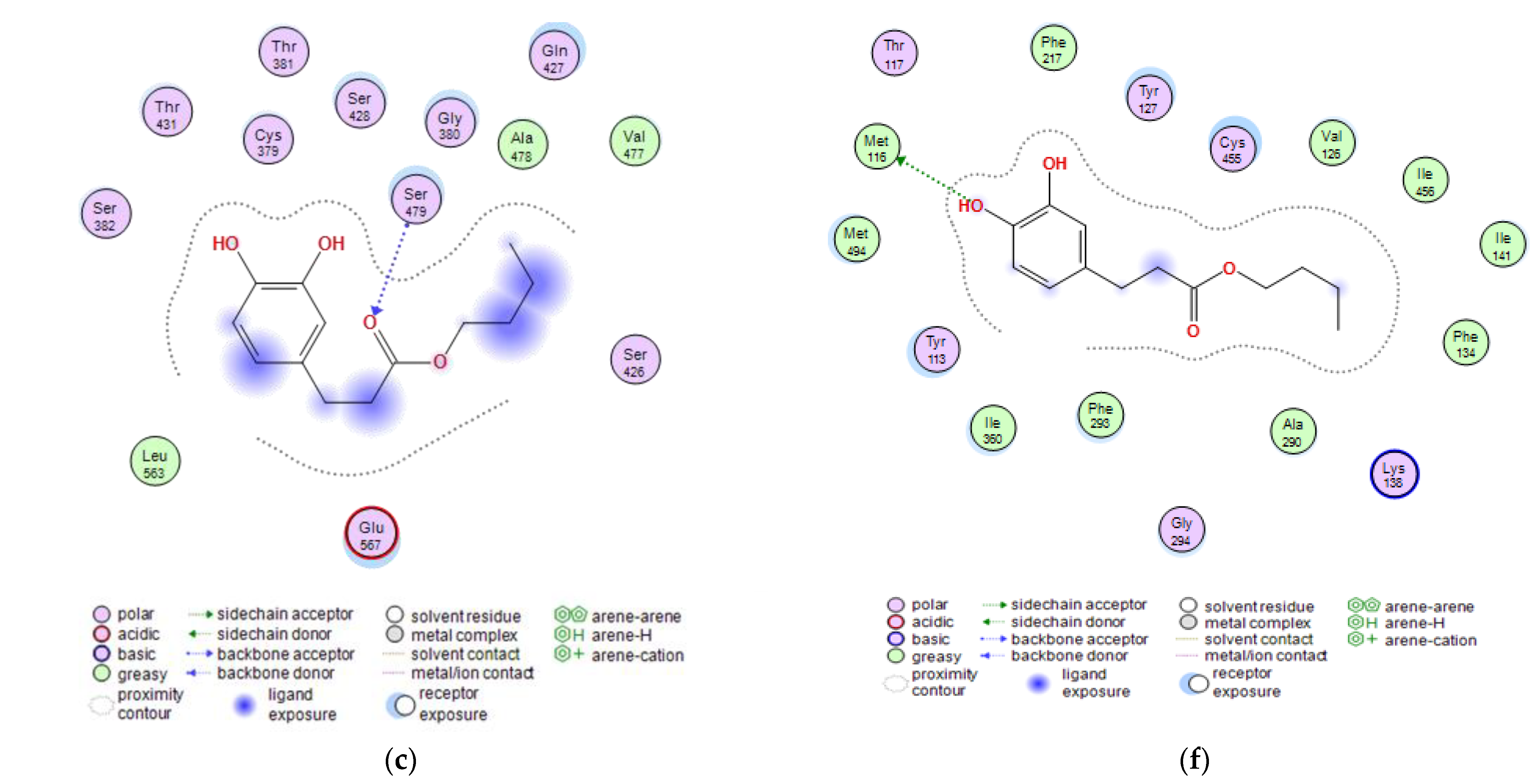
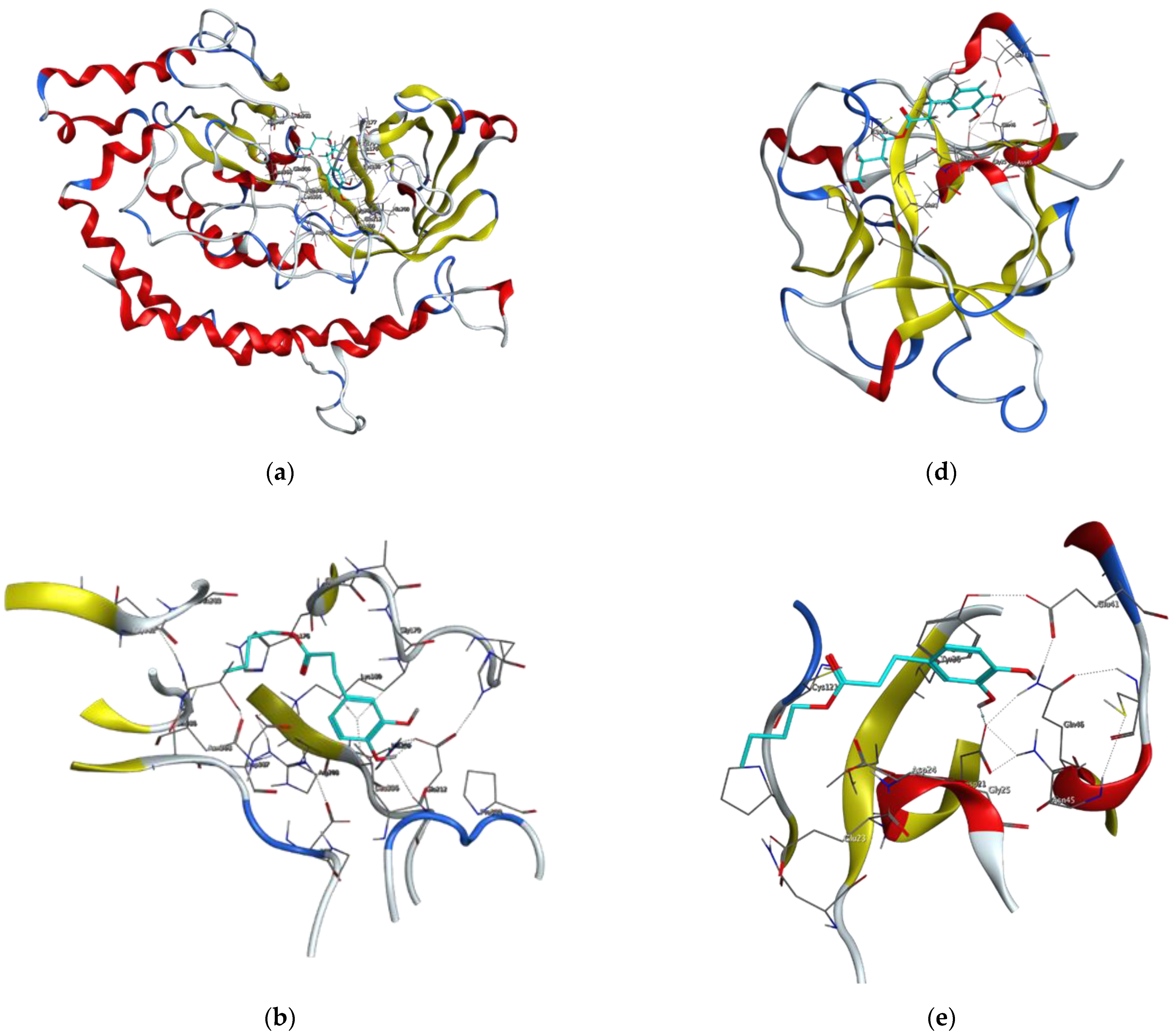
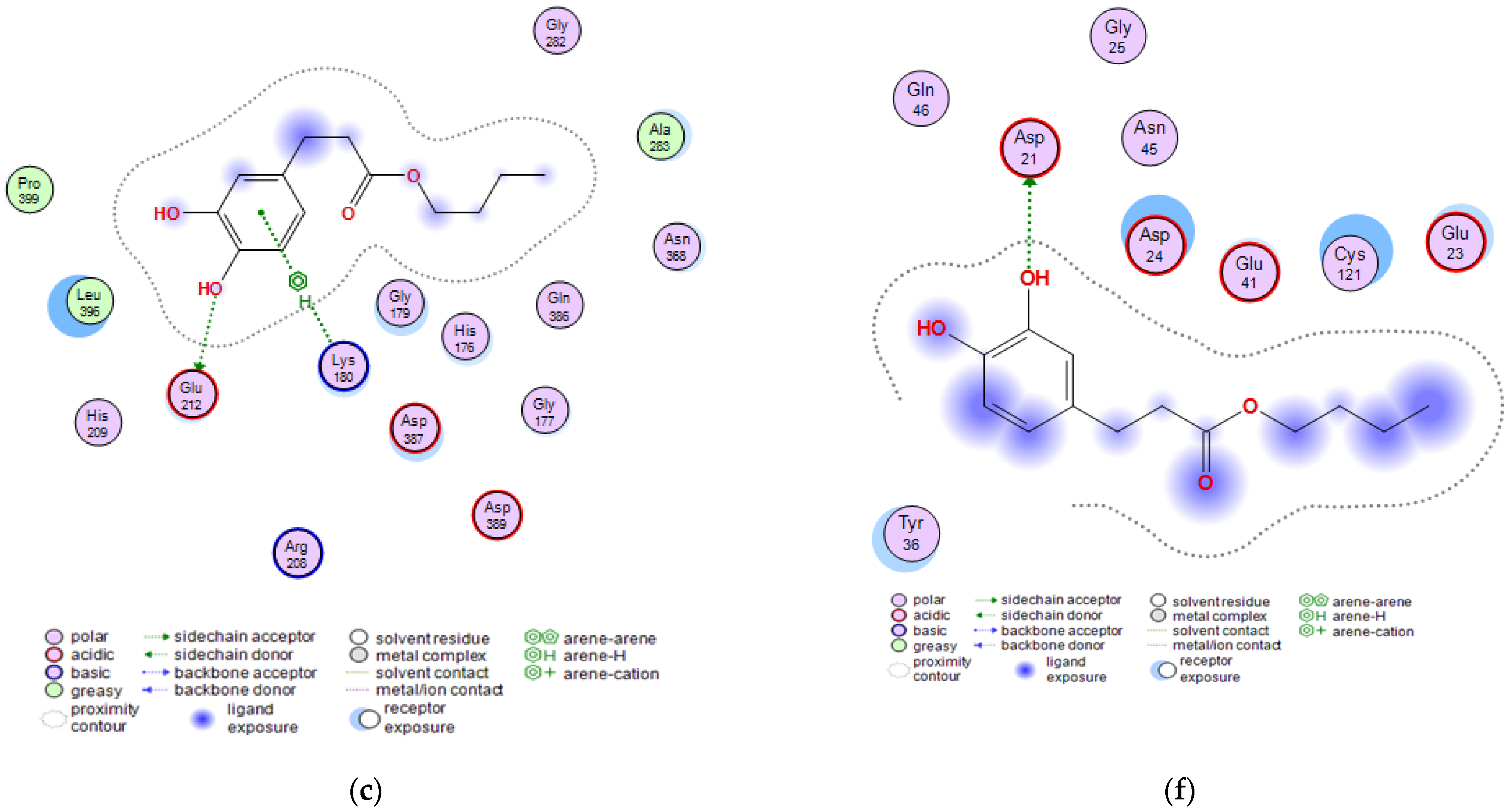
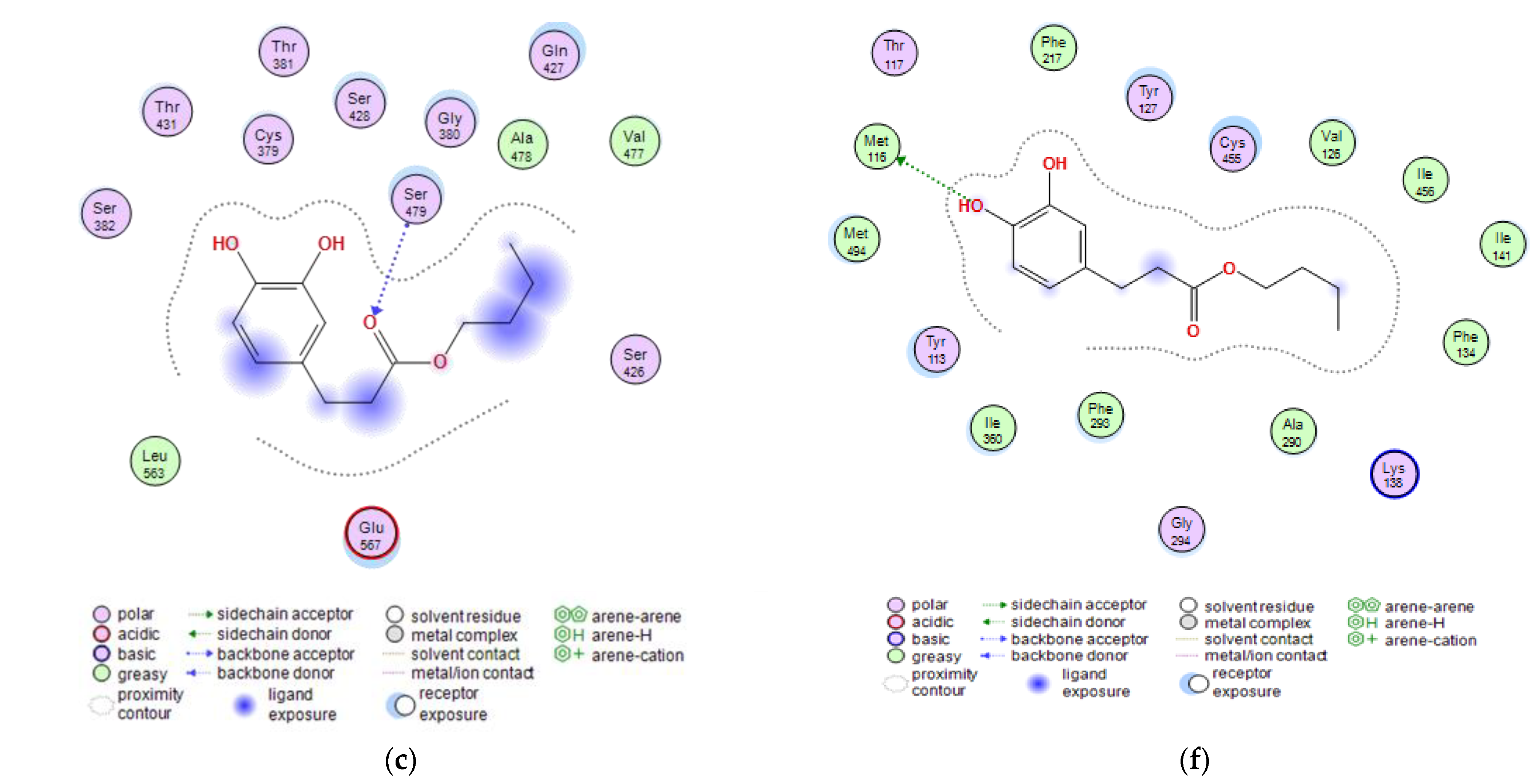
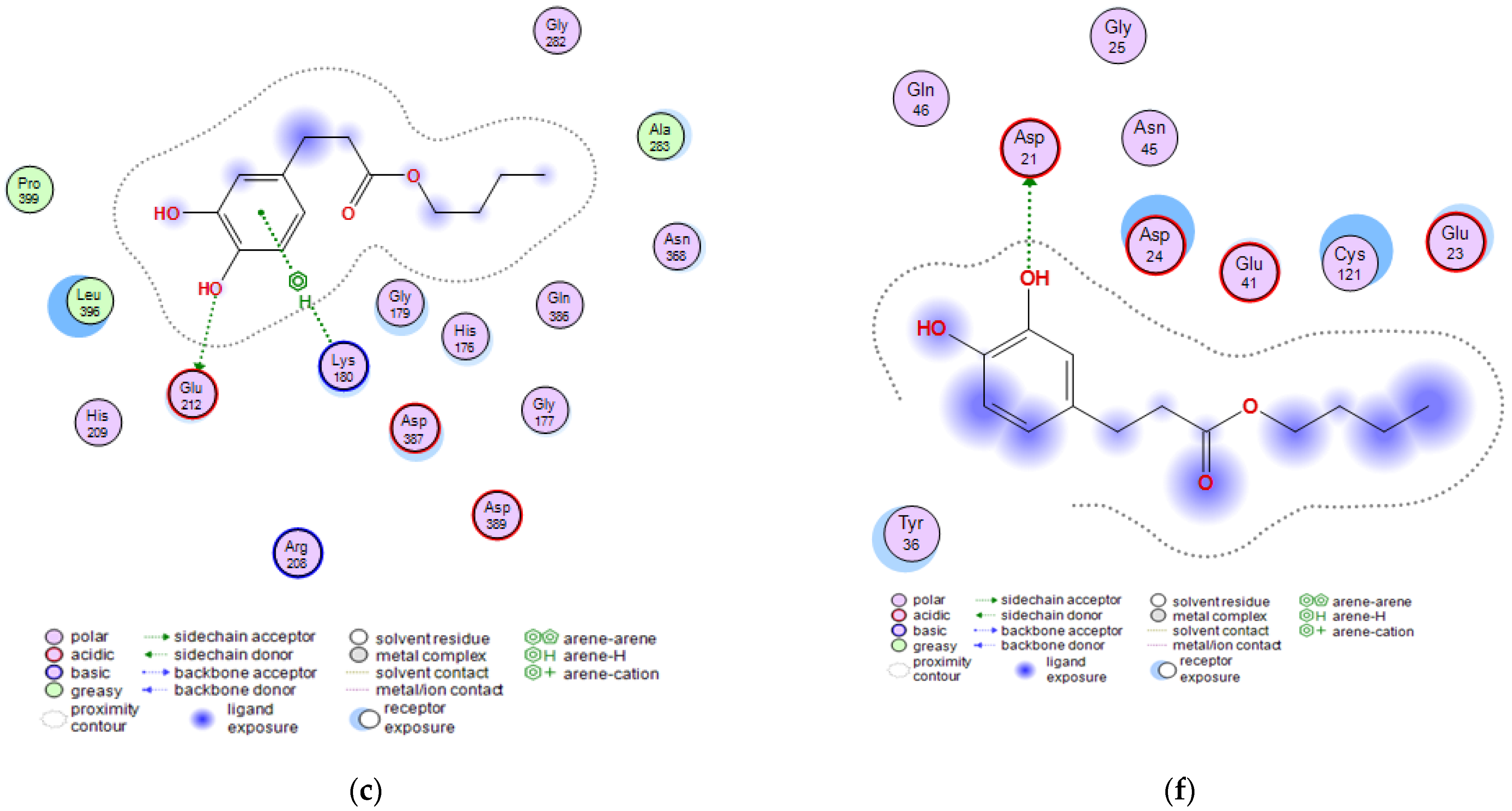
2.3. Molecular Docking Studies of Butyl Dihydrocaffeate
3. Materials and Methods
3.1. Materials
3.2. Microorganisms
3.3. Enzymatic Synthesis of Butyl Dihydrocaffeate
3.4. Evaluation of Antioxidant Activity
3.4.1. The DPPH· Assay
3.4.2. CUPRAC Method
3.5. Evaluation of Antimicrobial Activity
3.5.1. Minimum Inhibitory Concentration (MIC) Determination
3.5.2. Minimum Microbicidal Concentration (MMC) Determination
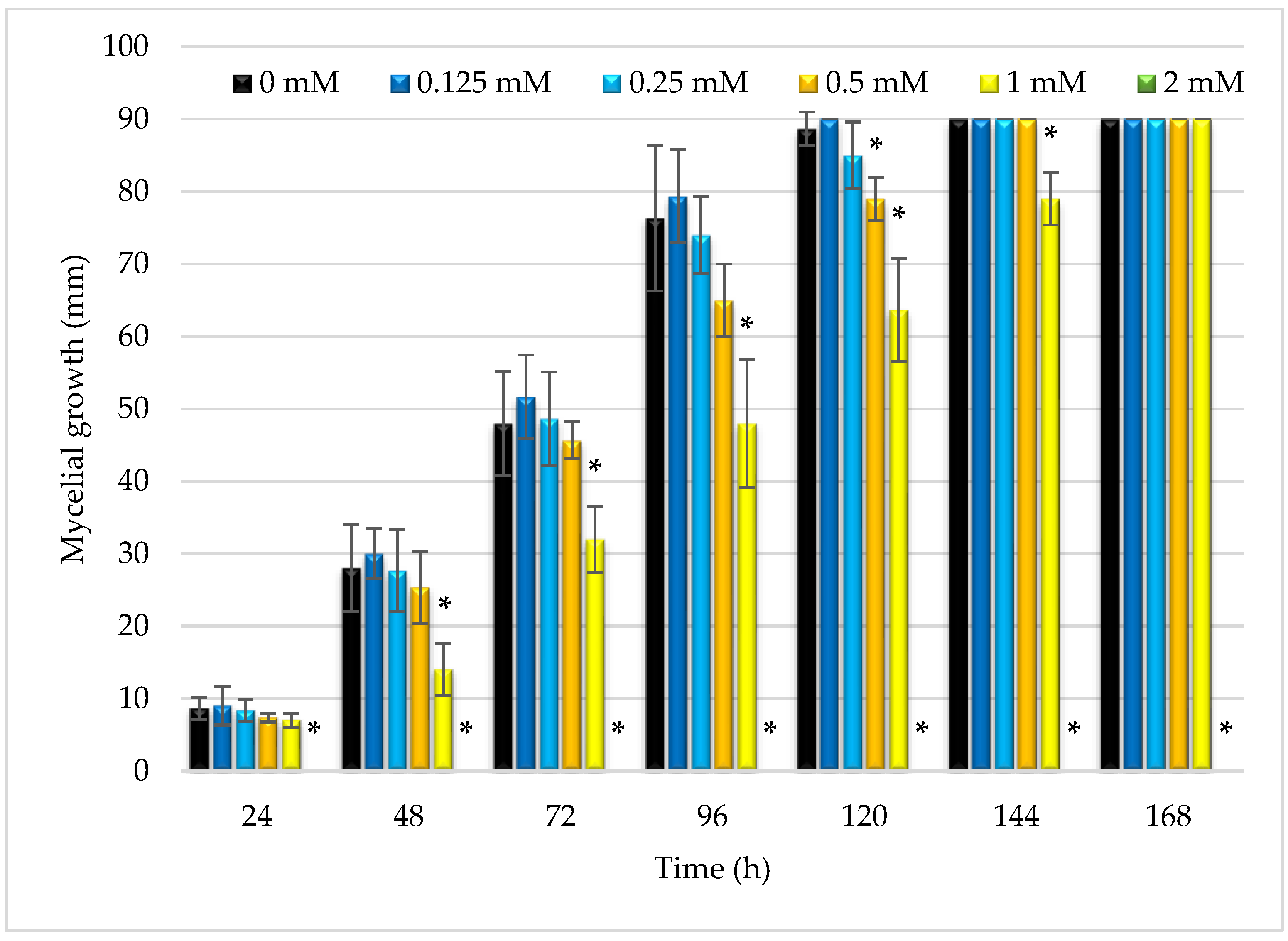
3.5.3. Inhibition of Mycelium Growth of R. oryzae by Butyl Dihydrocaffeate
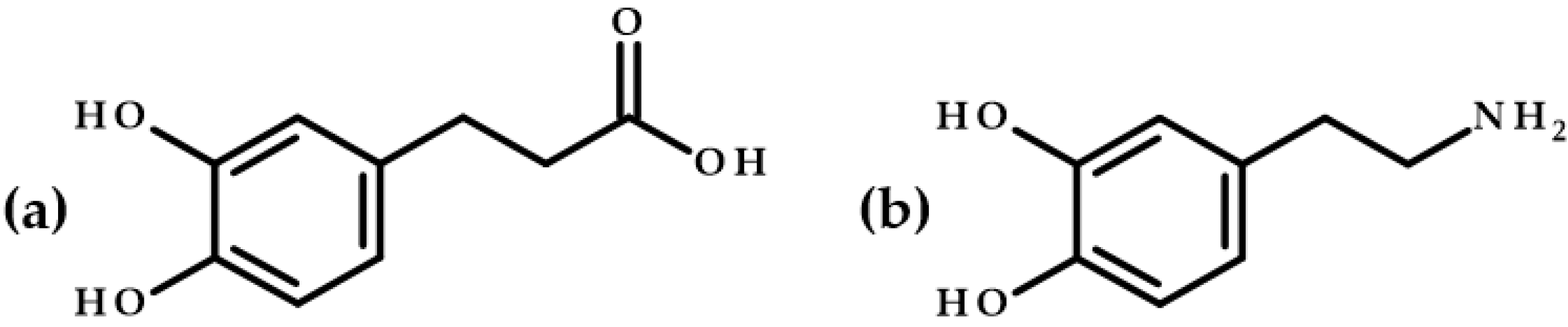
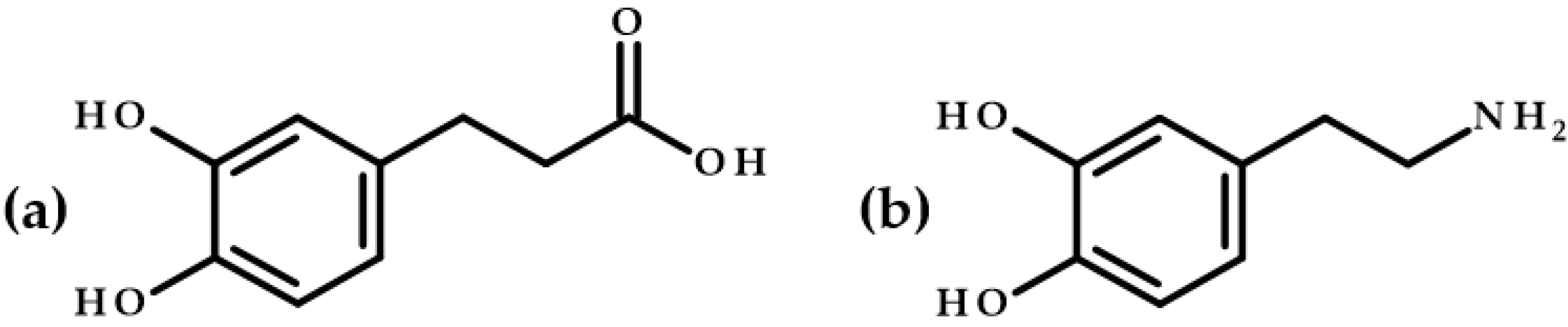
3.6. Structures’ Comparison of Dihydrocaffeic Acid and Its Butyl Ester
3.7. Statistical Analysis
3.8. Target Proteins and Ligands Selection for Molecular Docking Studies
3.9. Protein Modelling and Structure Quality Assessment
3.10. Protein/Ligand Preparation and Docking Simulation
3.10.1. Ligand Selection and Preparation
3.10.2. Protein Target Preparation and Binding/Docking Site Prediction
3.10.3. Docking Simulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oliveira, M.M.; Ratti, B.A.; Dare, R.G.; Silva, S.O.; Truiti, M.C.T.; Ueda-Nakamura, T.; Auzely-Velty, R.; Nakamura, C.V. Dihydrocaffeic Acid Prevents UVB-Induced Oxidative Stress Leading to the Inhibition of Apoptosis and MMP-1 Expression via p38 Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 2419096. [Google Scholar] [CrossRef] [PubMed]
- Poquet, L.; Clifford, M.N.; Williamson, G. Investigation of the metabolic fate of dihydrocaffeic acid. Biochem. Pharmacol. 2008, 75, 1218–1229. [Google Scholar] [CrossRef]
- Moon, J.H.; Terao, J. Antioxidant Activity of Caffeic Acid and Dihydrocaffeic Acid in Lard and Human Low-Density Lipoprotein. J. Agric. Food Chem. 1998, 46, 5062–5065. [Google Scholar] [CrossRef]
- Baeza, G.; Sarria, B.; Mateos, R.; Bravo, L. Dihydrocaffeic acid, a major microbial metabolite of chlorogenic acids, shows similar protective effect than a yerba mate phenolic extract against oxidative stress in HepG2 cells. Food Res. Int. 2018, 87, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Lee, B.J.; Bu, Y. Protective Effects of Dihydrocaffeic Acid, a Coffee Component Metabolite, on a Focal Cerebral Ischemia Rat Model. Molecules 2015, 20, 11930–11940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esfanjani, A.F.; Assadpour, E.; Jafari, S.M. Improving the bioavailability of phenolic compounds by loading them within lipid-based nanocarriers. Trends Food Sci. Technol. 2018, 76, 56–66. [Google Scholar] [CrossRef]
- Białecka-Florjańczyk, E.; Fabiszewska, A.; Zieniuk, B. Phenolic Acids Derivatives-Biotechnological Methods of Synthesis and Bioactivity. Curr. Pharm. Biotechnol. 2018, 19, 1098–1113. [Google Scholar] [CrossRef]
- Figueroa-Espinoza, M.C.; Villeneuve, P. Phenolic Acids Enzymatic Lipophilization. J. Agric. Food Chem. 2005, 53, 2779–2787. [Google Scholar] [CrossRef]
- Zieniuk, B.; Białecka-Florjańczyk, E.; Wierzchowska, K.; Fabiszewska, A. Recent advances in the enzymatic synthesis of lipophilic antioxidant and antimicrobial compounds. World J. Microbiol. Biotechnol. 2022, 38, 11. [Google Scholar] [CrossRef]
- Gholivand, S.; Lasekan, O.; Tan, C.P.; Abas, F.; Wei, L.S. Comparative study of the antioxidant activities of some lipase-catalyzed alkyl dihydrocafeates synthesized in ionic liquid. Food Chem. 2017, 224, 365–371. [Google Scholar] [CrossRef]
- Sørensen, A.D.M.; Nielsen, N.S.; Yang, Z.; Xu, X.; Jacobsen, C. Lipophilization of dihydrocaffeic acid affects its antioxidative properties in fish-oil-enriched emulsions. Eur. J. Lipid Sci. Technol. 2012, 114, 134–145. [Google Scholar] [CrossRef]
- Mansouri, A.; Makris, D.P.; Kefalas, P. Determination of hydrogen peroxide scavenging activity of cinnamic and benzoic acids employing a highly sensitive peroxyoxalate chemiluminescence-based assay: Structure–activity relationships. J. Pharm. Biomed. 2005, 39, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Farhoosh, R.; Johnny, S.; Asnaashari, M.; Molaahmadibahraseman, N.; Sharif, A. Structure-antioxidant activity relationships of o-hydroxyl, o-methoxy, and alkyl ester derivatives of p-hydroxybenzoic acid. Food Chem. 2016, 194, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Pyrzynska, K.; Pękal, A. Application of free radical diphenylpicrylhydrazyl (DPPH) to estimate the antioxidant capacity of food samples. Anal. Methods 2013, 5, 4288. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Wianowska, D.; Olszowy, M. On practical problems in estimation of antioxidant activity of compounds by DPPH method (Problems in estimation of antioxidant activity). Food Chem. 2012, 131, 1037–1043. [Google Scholar] [CrossRef]
- Wołosiak, R.; Drużyńska, B.; Derewiaka, D.; Piecyk, M.; Majewska, E.; Ciecierska, M.; Worobiej, E.; Pakosz, P. Verification of the Conditions for Determination of Antioxidant Activity by ABTS and DPPH Assays—A Practical Approach. Molecules 2022, 27, 50. [Google Scholar] [CrossRef]
- Çelik, S.E.; Özyürek, M.; Güçlü, K.; Apak, R. Solvent effects on the antioxidant capacity of lipophilic and hydrophilic antioxidants measured by CUPRAC, ABTS/persulphate and FRAP methods. Talanta 2010, 81, 1300–1309. [Google Scholar] [CrossRef]
- Zieniuk, B.; Groborz, K.; Wołoszynowska, M.; Ratusz, K.; Białecka-Florjańczyk, E.; Fabiszewska, A. Enzymatic Synthesis of Lipophilic Esters of Phenolic Compounds, Evaluation of Their Antioxidant Activity and Effect on the Oxidative Stability of Selected Oils. Biomolecules 2021, 11, 314. [Google Scholar] [CrossRef]
- Roleira, F.M.F.; Siquet, C.; Orru, E.; Garrido, E.M.; Garrido, J.; Milhazes, N.; Podda, G.; Paiva-Martins, F.; Reis, S.; Carvalho, R.A.; et al. Lipophilic phenolic antioxidants: Correlation between antioxidant profile, partition coefficients and redox properties. Bioorg. Med. Chem. 2010, 18, 5816–5825. [Google Scholar] [CrossRef]
- Gaspar, A.; Martins, M.; Silva, P.; Garrido, E.M.; Garrido, J.; Firuzi, O.; Miri, R.; Saso, L.; Borges, F. Dietary Phenolic Acids and Derivatives. Evaluation of the Antioxidant Activity of Sinapic Acid and Its Alkyl Esters. J. Agric. Food Chem. 2010, 58, 11273–11280. [Google Scholar] [CrossRef]
- Załuski, D.; Cieśla, Ł.; Janeczko, Z. Chapter 7-The Structure–Activity Relationships of Plant Secondary Metabolites with Antimicrobial, Free Radical Scavenging and Inhibitory Activity toward Selected Enzymes. Stud. Nat. Prod. Chem. 2015, 45, 217–249. [Google Scholar] [CrossRef]
- Shi, Y.G.; Zhu, Y.J.; Shao, S.Y.; Zhang, R.R.; Wu, Y.; Zhu, C.M.; Liang, X.R.; Cai, W.Q. Alkyl Ferulate Esters as Multifunctional Food Additives: Antibacterial Activity and Mode of Action against Escherichia coli in Vitro. J. Agric. Food Chem. 2018, 66, 12088–12101. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.G.; Bian, L.Q.; Zhu, Y.J.; Zhang, R.R.; Shao, S.Y.; Wu, Y.; Chen, Y.W.; Dang, Y.L.; Ding, Y.; Sun, H. Multifunctional alkyl ferulate esters as potential food additives: Antibacterial activity and mode of action against Listeria monocytogenes and its application on American sturgeon caviar preservation. Food Control 2019, 96, 390–402. [Google Scholar] [CrossRef]
- Shi, Y.G.; Zhang, R.R.; Zhu, C.M.; Liang, X.R.; Ettelaie, R.; Jiang, L.; Lin, S. On the mechanism behind enhanced antibacterial activity of alkyl gallate esters against foodborne pathogens and its application in Chinese icefish preservation. Food Microbiol. 2021, 99, 103817. [Google Scholar] [CrossRef]
- Xu, J.-W.; Liao, C.-C.; Hung, K.-C.; Wang, Z.-Y.; Tung, Y.-T.; Wu, J.-H. Proteomics Reveals Octyl Gallate as an Environmentally Friendly Wood Preservative Leading to Reactive Oxygen Species-Driven Metabolic Inflexibility and Growth Inhibition in White-Rot Fungi (Lenzites betulina and Trametes versicolor). J. Fungi 2021, 7, 145. [Google Scholar] [CrossRef] [PubMed]
- Bozzini, T.; Botta, G.; Delfino, M.; Onofri, S.; Saladino, R.; Amatore, D.; Sgarbanti, R.; Nencioni, L.; Palamara, A.T. Tyrosinase and Layer-by-Layer supported tyrosinases in the synthesis of lipophilic catechols with antiinfluenza activity. Bioorg. Med. Chem. 2013, 21, 7699–7708. [Google Scholar] [CrossRef] [PubMed]
- Botta, G.; Bizzarri, B.M.; Garozzo, A.; Timpanaro, R.; Bisignano, B.; Amatore, D.; Palamara, A.T.; Nencioni, L.; Saladino, R. Carbon nanotubes supported tyrosinase in the synthesis of lipophilic hydroxytyrosol and dihydrocaffeoyl catechols with antiviral activity against DNA and RNA viruses. Bioorg. Med. Chem. 2015, 23, 5345–5351. [Google Scholar] [CrossRef]
- Zaki, S.M.; Elkholy, I.M.; Elkady, N.A.; Abdel-Ghany, K. Mucormycosis in Cairo, Egypt: Review of 10 reported cases. Med. Mycol. 2014, 52, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Madanagopal, P.; Ramprabhu, N.; Jagadeesan, R. In silico prediction and structure-based multitargeted molecular docking analysis of selected bioactive compounds against mucormycosis. Bull. Natl. Res. Cent. 2022, 46, 24. [Google Scholar] [CrossRef]
- Ghosh, B.; Rani Ray, R. Current Commercial Perspective of Rhizopus oryzae: A Review. J. Appl. Sci. 2011, 11, 2470–2486. [Google Scholar] [CrossRef] [Green Version]
- Cantabrana, I.; Perise, R.; Hernandez, I. Uses of Rhizopus oryzae in the kitchen. Int. J. Gastron. Food Sci. 2015, 2, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Prajapati, J.; Rao, P.; Poojara, L.; Goswami, D.; Acharya, D.; Patel, S.K.; Rawal, R.M. Unravelling the antifungal mode of action of curcumin by potential inhibition of CYP51B: A computational study validated in vitro on mucormycosis agent, Rhizopus oryzae. Arch. Biochem. Biophys. 2021, 15, 109048. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular Properties that Influence the Oral Bioavailability of Drug Candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Gupta, U.; Gupta, S.; Wadhwa, G.; Gabrani, R.; Sharma, S.K.; Jain, C.K. Molecular docking of glucosamine-6-phosphate synthase in Rhizopus oryzae. Bioinformation 2011, 7, 285–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [Green Version]
- Gebremariam, T.; Liu, M.; Luo, G.; Bruno, V.; Phan, Q.T.; Waring, A.J.; Edwards, J.E., Jr.; Filler, S.G.; Yeaman, M.R.; Ibrahim, A.S. CotH3 mediates fungal invasion of host cells during mucormycosis. J. Clin. Investig. 2014, 124, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Soliman, S.M.S.; Baldin, C.; Gu, Y.; Singh, S.; Geremariam, T.; Swidergall, M.; Alqarihi, A.; Youssef, E.G.; Alkhazraji, S.; Pikoulas, A.; et al. Mucoricin is a ricin-like toxin that is critical for the pathogenesis of mucormycosis. Nat. Microbiol. 2021, 6, 313–326. [Google Scholar] [CrossRef]
- Pokharkar, O.; Lakshmanan, H.; Zyryanov, G.; Tsurkan, M. In Silico Evaluation of Antifungal Compounds from Marine Sponges against COVID-19-Associated Mucormycosis. Mar. Drugs 2022, 20, 215. [Google Scholar] [CrossRef]
- Zanetti, M.; Carniel, T.K.; Valerio, A.; de Oliveira, J.V.; de Oliveira, D.; de Araujo, P.H.H.; Riella, H.G.; Fiori, M.A. Synthesis of geranyl cinnamate by lipase-catalyzed reaction and its evaluation as an antimicrobial agent. J. Chem. Technol. Biotechnol. 2017, 92, 115–121. [Google Scholar] [CrossRef]
- Özyürek, M.; Güçlü, K.; Apak, R. The main and modified CUPRAC methods of antioxidant measurement. Trends Analyt. Chem. 2011, 30, 652–664. [Google Scholar] [CrossRef]
- ISO 20776–1; Clinical Laboratory Testing and In Vitro Diagnostic Test Systems–Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices–Part 1: Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases. International Organization for Standards: Geneva, Switzerland, 2006.
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo-distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ononamadu, C.J.; Abdalla, M.; Ihegboro, G.O.; Li, J.; Owolarafe, T.A.; John, T.D.; Tian, Q. In silico identification and study of potential anti-mosquito juvenile hormone binding protein (MJHBP) compounds as candidates for dengue virus-Vector insecticides. Biochem. Biophys. Rep. 2021, 28, 101178. [Google Scholar] [CrossRef] [PubMed]
- Galli, C.L.; Sensi, C.; Fumagalli, A.; Parravicini, C.; Marinovich, M.; Eberini, I. A Computational Approach to Evaluate the Androgenic Affinity of Iprodione, Procymidone, Vinclozolin and Their Metabolites. PLoS ONE 2014, 9, e104822. [Google Scholar] [CrossRef] [PubMed]
- Abin-Carriquiry, J.A.; Zunini, M.P.; Cassels, B.K.; Wonnacott, S.; Dajas, F. In silico characterization of cytisinoids docked into an acetylcholine binding protein. Bioorg. Med. Chem. Lett. 2010, 20, 3683–3687. [Google Scholar] [CrossRef]
- Corbeil, C.R.; Williams, C.I.; Labute, P. Variability in docking success rates due to dataset preparation. J. Comput. Aided. Mol. Des. 2012, 26, 775–786. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPPH· | CUPRAC | |||
|---|---|---|---|---|
| Compound | Methanol (mM) | Ethyl Acetate (mM) | Chloroform (mM) | (TEAC) |
| BDHC | 0.16 ± 0.01 Ca | 0.18 ± 0.01 Ba | 0.22 ± 0.01 Bb | 3.50 ± 0.03 A |
| DHCA | 0.12 ± 0.01 Aa | 0.14 ± 0.01 Ab | 0.15 ± 0.01 Ab | 2.73 ± 0.08 B |
| LAA | 0.28 ± 0.04 Da | 0.23 ± 0.02 Ca | 3.25 ± 0.08 b | 1.56 ± 0.01 D |
| BHT | 0.58 ± 0.03 Ea | 21.34 ± 1.55 c | 16.60 ± 1.79 b | 1.92 ± 0.01 C |
| GA | 0.09 ± 0.01 Aa | 0.13 ± 0.1 Ab | 0.17 ± 0.01 Ac | 3.37 ± 0.06 A |
| CA | 0.14 ± 0.01 Ba | 0.13 ± 0.01 Aa | 0.17 ± 0.01 Ab | 3.35 ± 0.09 A |
| BDHC | DHCA | 1-Butanol | ||||
|---|---|---|---|---|---|---|
| MIC (mM) | MMC (mM) | MIC (mM) | MMC (mM) | MIC (mM) | MMC (mM) | |
| E. coli PCM 2057 | 16 | 32 | 16 | 32 | 32 | 64 |
| E. cloacae PCM 2848 | 8 | 16 | 2 | 4 | 16 | 32 |
| S. marcescens PCM 549 | 4 | 8 | 16 | 32 | 16 | 32 |
| B. subtilis PCM 486 | 8 | 16 | 2 | >64 | 16 | >64 |
| L. monocytogenes PCM 2191 | 8 | 16 | 8 | 16 | 16 | 64 |
| S. aureus PCM 2054 | 4 | 8 | 2 | 4 | 16 | >64 |
| R. oryzae DSM 2199 | 1 | 2 | 32 | >64 | 32 | >64 |
| DHCA | BDHC | Reference Value * | |
|---|---|---|---|
| Molecular weight (g/mol) | 182.17 | 238.28 | <500 |
| Hydrogen bond donors | 3 | 2 | <5 |
| Hydrogen bond acceptors | 4 | 4 | <10 |
| Rotatable bonds | 3 | 7 | <10 |
| Molar refractivity | 46.48 | 65.58 | |
| TPSA (Ų) | 77.76 | 66.76 | ≤140 |
| LogP | 0.63 | 2.44 | <5 |
| LogS | −0.62 (Very soluble) | −4.42 (Moderately soluble) | |
| Rule of five violations | 0 | 0 | |
| Gastrointestinal absorption | High | High | |
| The blood–brain barrier permeant | No | Yes |
| Target Protein | Compound | Binding Energy (kcal/mol) | Interacting Amino Acid Residues |
|---|---|---|---|
| Glutamine-fructose-6-phosphate transaminase (GFAT) | Posaconazole | −7.5116 | a Glu567, b Ser428, c Ser382 |
| Isavuconazole | −5.8663 | a Glu567, b Ser428 | |
| 12,28-Oxamanzamine A | −6.1423 | b Glu567, b Thr381 | |
| DHCA | −4.1152 | b Ser479 | |
| BDHC | −5.1152 | b Ser479 | |
| 14-α sterol demethylase B | Posaconazole | −9.7030 | a Cys455, c Tyr133, c Phe222 |
| Isavuconazole | −6.1767 | a His453, b Gly294, c Val291, c Cys455 | |
| 12,28-Oxamanzamine A | −4.0297 | a Met494 | |
| DHCA | −4.5334 | a Met494 | |
| BDHC | −6.1416 | a Met116 | |
| Invasin CotH3 | Posaconazole | −8.9723 | a Ala303, c Lys180 |
| Isavuconazole | −7.3442 | c Gly179, c Lys180 | |
| 12,28-Oxamanzamine A | −7.3644 | a Asn368, a Asp387, e His176, c Thr367, d Asp387 | |
| DHCA | −4.9011 | b Asn368, b Gln386 | |
| BDHC | −6.3490 | a Glu212, c Lys180 | |
| Mucoricin | Posaconazole | −6.5630 | a Glu23 |
| Isavuconazole | −5.4709 | a Glu41, a Asp21 | |
| 12,28-Oxamanzamine A | −5.5611 | aGlu41 | |
| DHCA | −4.3442 | a Asp21, b Lys59 | |
| BDHC | −4.6642 | a Asp21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieniuk, B.; Ononamadu, C.J.; Jasińska, K.; Wierzchowska, K.; Fabiszewska, A. Lipase-Catalyzed Synthesis, Antioxidant Activity, Antimicrobial Properties and Molecular Docking Studies of Butyl Dihydrocaffeate. Molecules 2022, 27, 5024. https://doi.org/10.3390/molecules27155024
Zieniuk B, Ononamadu CJ, Jasińska K, Wierzchowska K, Fabiszewska A. Lipase-Catalyzed Synthesis, Antioxidant Activity, Antimicrobial Properties and Molecular Docking Studies of Butyl Dihydrocaffeate. Molecules. 2022; 27(15):5024. https://doi.org/10.3390/molecules27155024
Chicago/Turabian StyleZieniuk, Bartłomiej, Chimaobi James Ononamadu, Karina Jasińska, Katarzyna Wierzchowska, and Agata Fabiszewska. 2022. "Lipase-Catalyzed Synthesis, Antioxidant Activity, Antimicrobial Properties and Molecular Docking Studies of Butyl Dihydrocaffeate" Molecules 27, no. 15: 5024. https://doi.org/10.3390/molecules27155024
APA StyleZieniuk, B., Ononamadu, C. J., Jasińska, K., Wierzchowska, K., & Fabiszewska, A. (2022). Lipase-Catalyzed Synthesis, Antioxidant Activity, Antimicrobial Properties and Molecular Docking Studies of Butyl Dihydrocaffeate. Molecules, 27(15), 5024. https://doi.org/10.3390/molecules27155024