
Modulatory Effect of Diosmin and Diosmetin on Metalloproteinase Activity and Inflammatory Mediators in Human Skin Fibroblasts Treated with Lipopolysaccharide

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
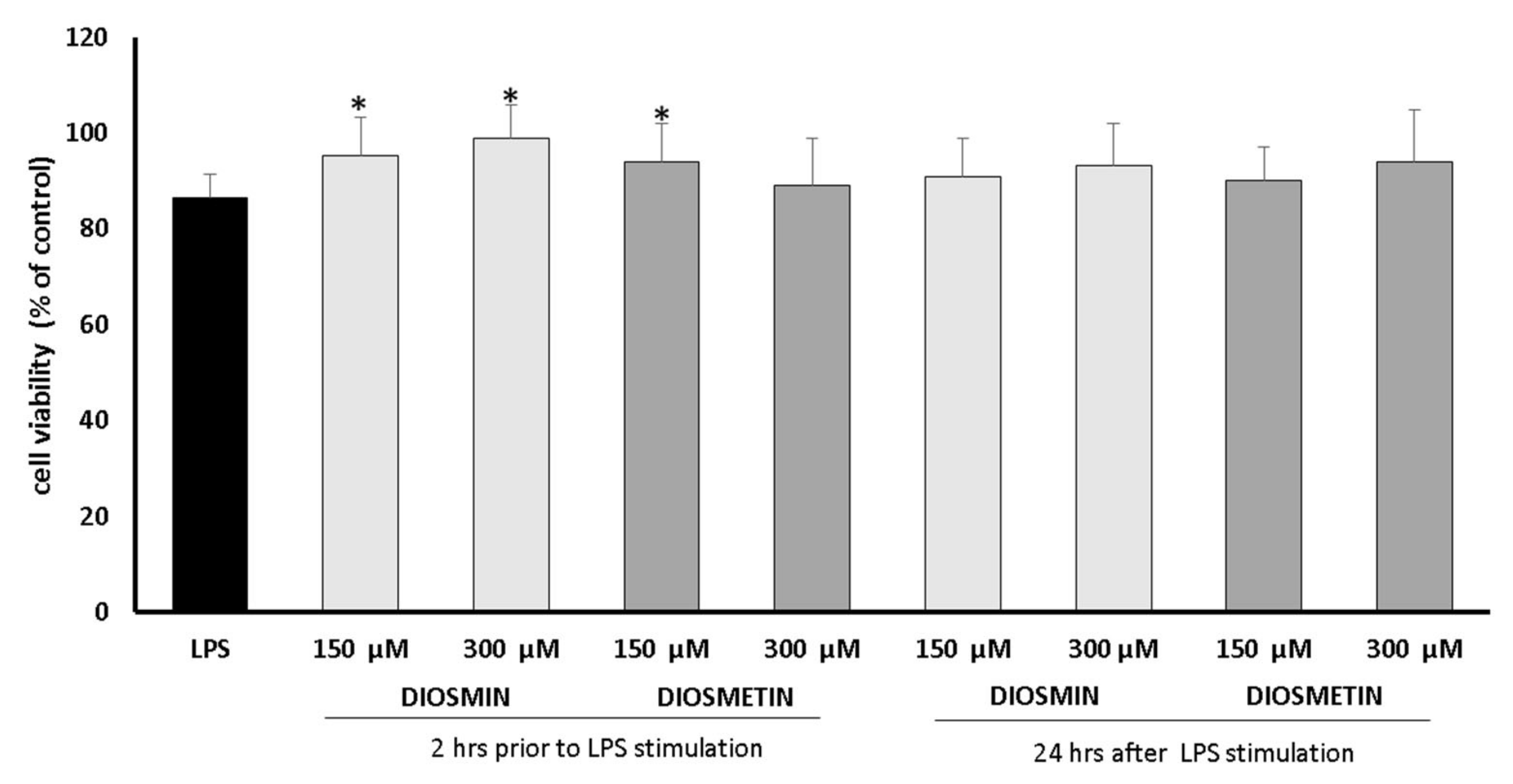
2.1. Cell Viability
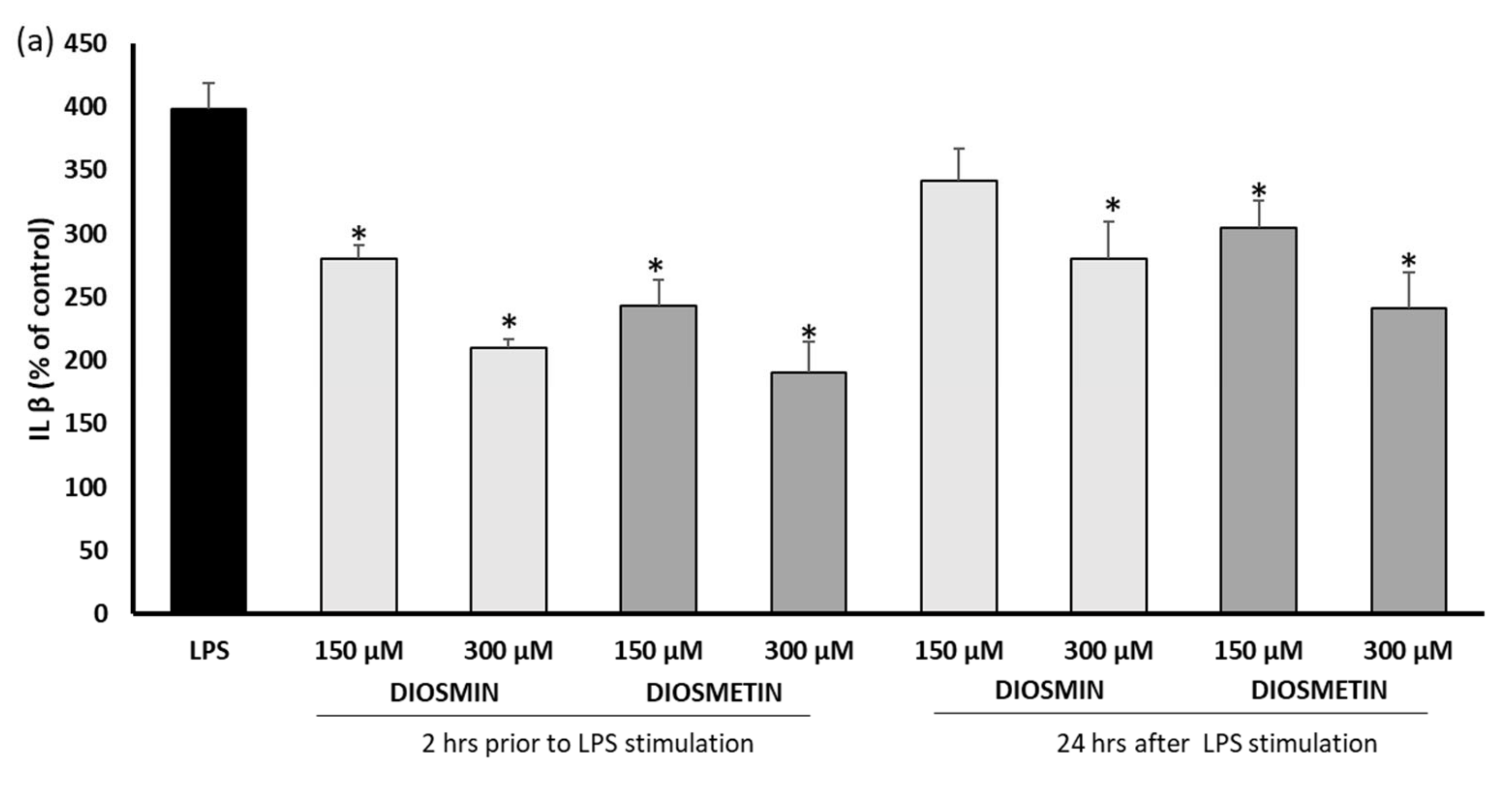
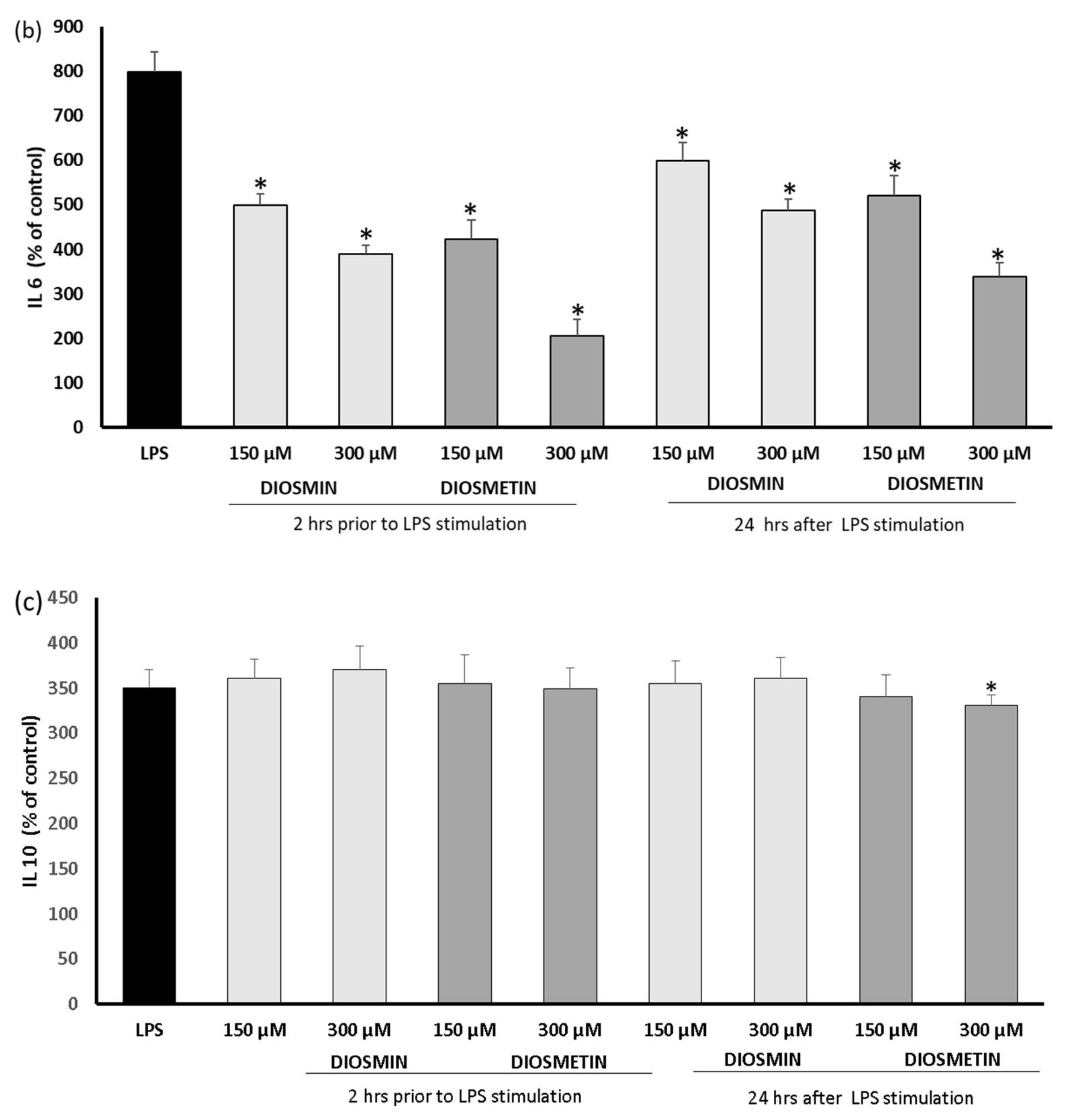
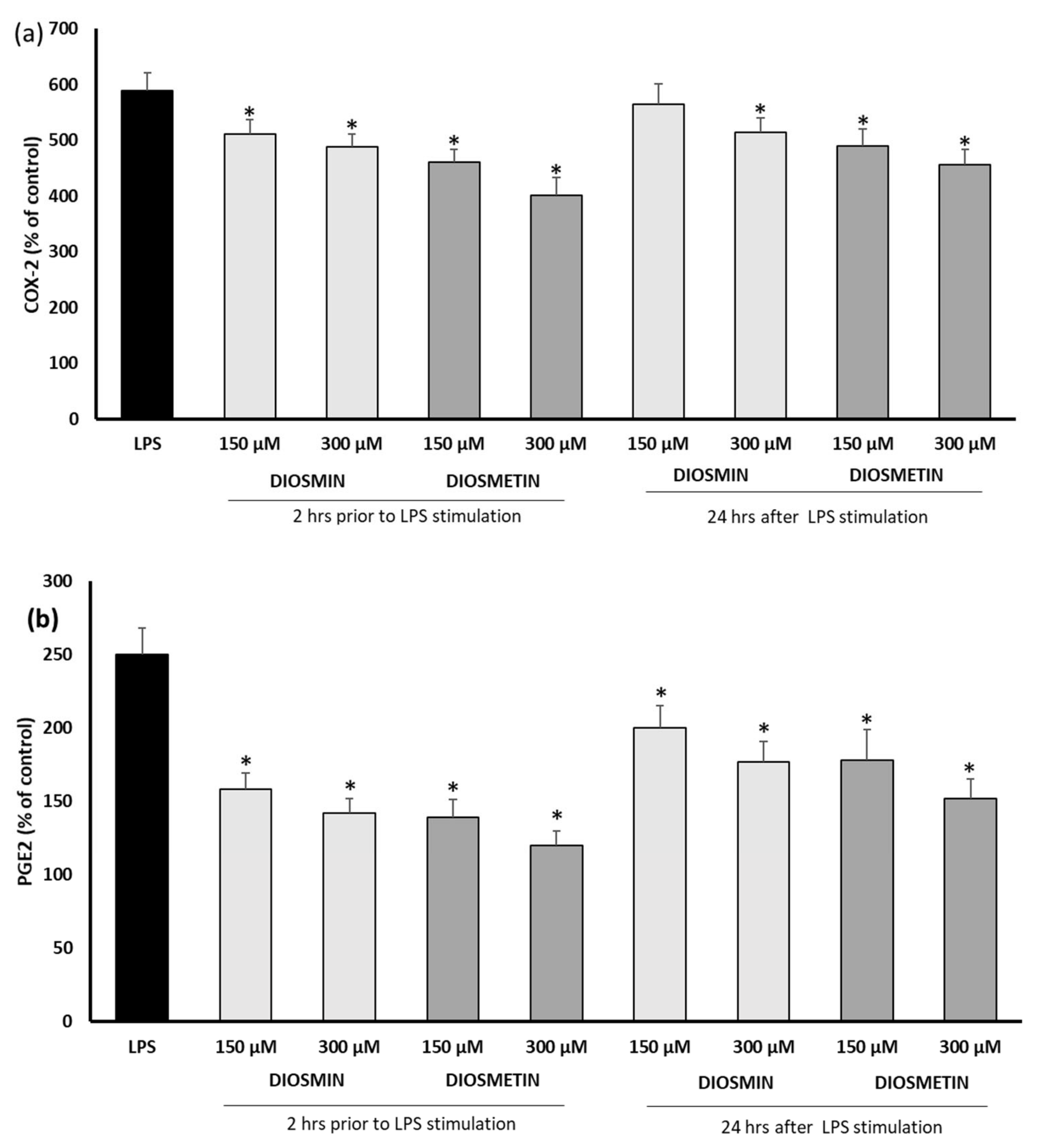
2.2. Effect of Diosmin/Diosmetin on Interleukins, COX-2, and PGE-2 in Human Skin Fibroblast Cells Treated with LPS
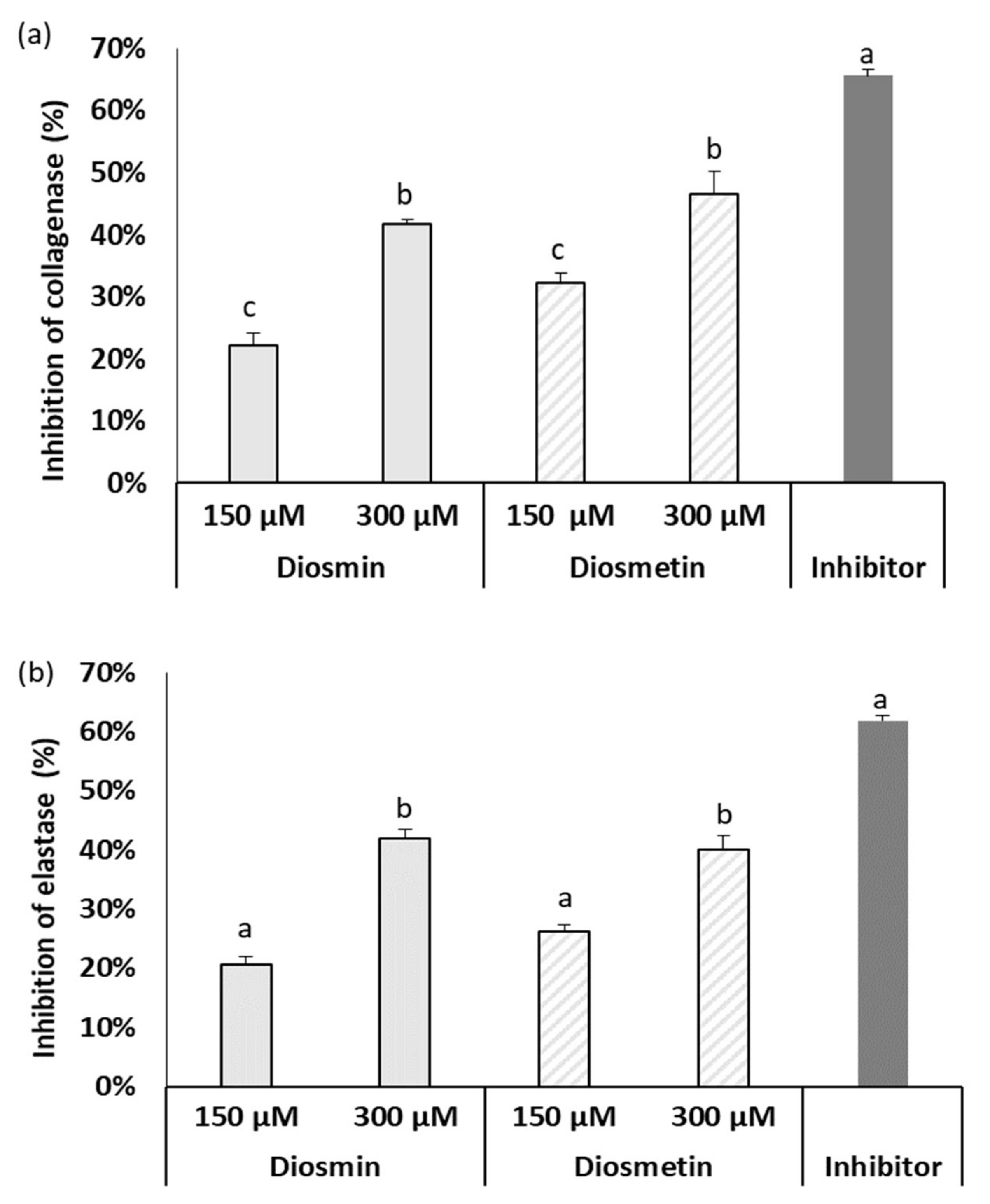
2.3. Effect of Diosmin/Diosmetin on Metalloproteinase Activity
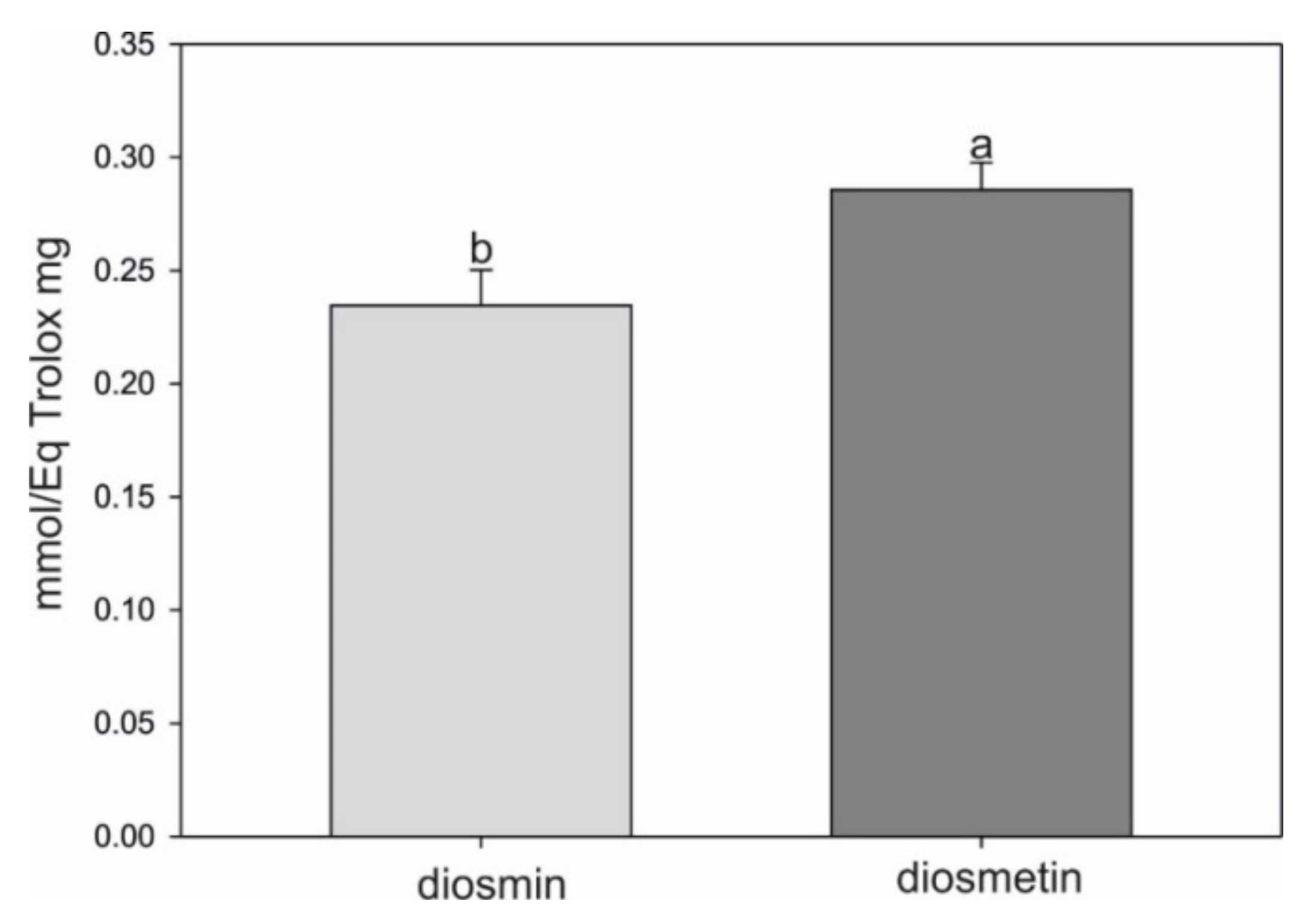
2.4. Antioxidant Assay
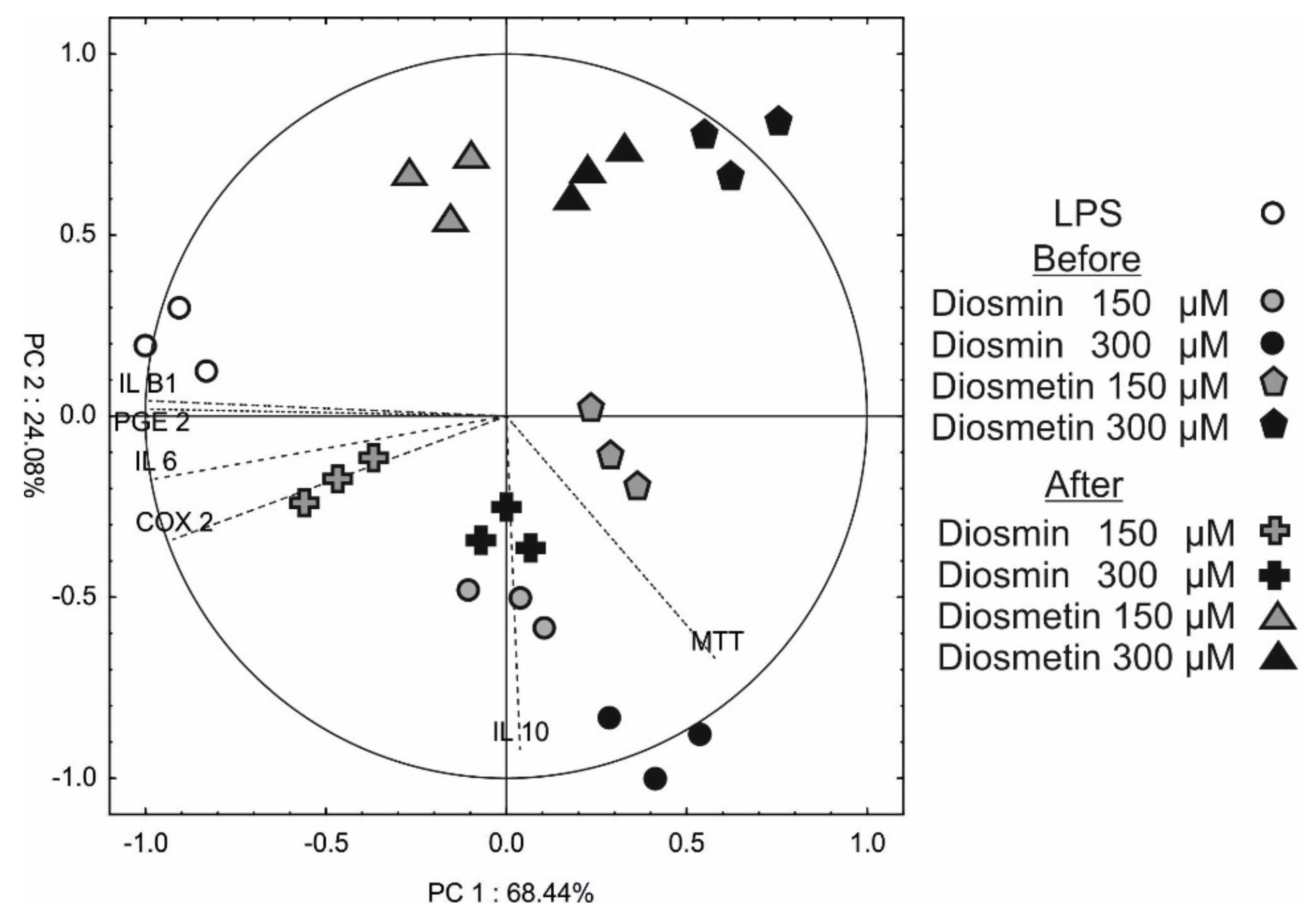
2.5. Principle Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Toxicity Assay
4.2.1. MTT Assay
4.2.2. Neutral Red Uptake Assay
4.3. Experimental Design for Analysis of Interleukins, PEG, and Cox-2 in Cells Treated with LPS
4.4. Metalloproteinase Activity Assay
4.4.1. Anti-Elastase Activity
4.4.2. Anti-Collagenase Activity
4.5. Antioxidant Activity Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bogucka-Kocka, A.; Wozniak, M.; Feldo, M.; Kocki, J.; Szewczyk, K. Diosmin—Isolation Techniques, Determination in Plant Material and Pharmaceutical Formulations, and Clinical Use. Nat. Prod. Commun. 2013, 8, 545–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wan, R.; Yin, G.; Xiong, J.; Hu, Y.; Xing, M.; Cang, X.; Fan, Y.; Xiao, W.; Qiu, L.; et al. Diosmetin ameliorates the severity of cerulein-induced acute pancreatitis in mice by inhibiting the activation of the nuclear factor-κB. Int. J. Clin. Exp. Pathol. 2014, 7, 2133–2142. [Google Scholar] [PubMed]
- Steinbruch, M.; Nunes, C.; Gama, R.; Kaufman, R.; Gama, G.; Suchmacher Neto, M.; Nigri, R.; Cytrynbaum, N.; Brauer Oliveira, L.; Bertaina, I.; et al. Is nonmicronized diosmin 600 mg as effective as micronized diosmin 900 mg plus hesperidin 100 mg on chronic venous disease symptoms? Results of a noninferiority study. Int. J. Vasc. Med. 2020, 2020, 4237204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldo, M.; Wójciak-Kosior, M.; Sowa, I.; Kocki, J.; Bogucki, J.; Zubilewicz, T.; Kęsik, J.; Bogucka-Kocka, A. Effect of Diosmin Administration in Patients with Chronic Venous Disorders on Selected Factors Affecting Angiogenesis. Molecules 2019, 24, 3316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldo, M.; Woźniak, M.; Wójciak-Kosior, M.; Sowa, I.; Kot-Waśnik, A.; Aszyk, J.; Bogucki, J.; Zubilewicz, T.; Bogucka-Kocka, A. Influence of diosmin treatment on the level of oxidative stress markers in patients with chronic venous insufficiency. Oxid. Med. Cell. Longev. 2018, 2018, 2561705. [Google Scholar] [CrossRef] [Green Version]
- Jantet, G. Chronic venous insufficiency: Worldwide results of the RELIEF study. Reflux assessment and quality of life improvement with micronized flavonoids. Angiology 2002, 53, 245–256. [Google Scholar] [CrossRef]
- Nicolaides, A.N. The Most Severe Stage of Chronic Venous Disease: An Update on the Management of Patients with Venous Leg Ulcers. Adv. Ther. 2020, 37, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Jafferany, M.; Pastolero, P. Psychiatric and Psychological Impact of Chronic Skin Disease. Prim. Care Companion CNS Disord. 2018, 20, 17nr02247. [Google Scholar] [CrossRef]
- Boisnic, S.; Branchet, M.C.; Gouhier-Kodas, C.; Verriere, F.; Jabbour, V. Anti-inflammatory and antiradical effects of a 2% diosmin cream in a human skin organ culture as model. J. Cosmet. Dermatol. 2018, 17, 848–854. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Li, J.-M.; Xi, C.; Li, H.-H.; Liu, Y.-L.; Wang, Y.-P.; Xuan, L.-J. Magnesium lithospermate B protects the endothelium from inflammation-induced dysfunction through activation of Nrf2 pathway. Acta Pharm. Sin. 2019, 40, 867–878. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, D.; Liu, Y.; Yu, J.; Qiao, L.; Lin, S.; Chen, D.; Zhong, G.; Lu, X.; Wang, Y.; et al. Long Noncoding RNA HOXA-AS3 integrates NF-κB signaling to regulate endothelium inflammation. Mol. Cell. Biol. 2019, 39, e00139-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visee, R.; Nagase, H. Matrix metalloproteinase and tissue inhibitors of metalloproteinase: Structure, function and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galis, Z.S.; Khatri, J.J. Matrix metalloproteinases in vascular remodeling and vascular disease. Biochem. Pharmacol. 2002, 90, 251–262. [Google Scholar]
- Venturi, M.; Bonavin, L.; Annoni, F.; Colombo, L.; Butera, C.; Peracchia, A.; Mussini, E. Biochemical assay of collagen and elastin in the normal and varicose vein wall. J. Surg. Res. 1996, 60, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Sansilvestri-Morel, P.; Fioretti, F.; Rupin, A.; Senni, K.; Fabiani, J.N.; Godeau, G.; Verbeuren, T.J. Comparison of extracellular matrix in skin and saphenous weins from paients with varocose veins: Does the skin reflects venous matrix changes? Clin. Sci. 2007, 112, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Antonicelli, F.; Bellon, G.; Debelle, L.; Hornebeck, W. Elastin-elastases and inflamm-aging. Curr. Top. Dev. Biol. 2007, 79, 99–155. [Google Scholar]
- Fields, G.B. Interstitial collagen metabolism. J. Biol. Chem. 2013, 288, 8785–8793. [Google Scholar] [CrossRef] [Green Version]
- Ortega, M.A.; Fraile-Martínez, O.; García-Montero, C.; Álvarez-Mon, M.A.; Chaowen, C.; Ruiz-Grande, F.; Pekarek, L.; Monserrat, J.; Asúnsolo, A.; García-Honduvilla, N.; et al. Understanding Chronic Venous Disease: A Critical Overview of Its Pathophysiology and Medical Management. J. Clin. Med. 2021, 10, 3239. [Google Scholar] [CrossRef]
- Bergan, J.J. Chronic venous insufficiency and the therapeutic effect of Daflon 500 mg. Angiology 2005, 56 (Suppl. S1), S21–S24. [Google Scholar] [CrossRef]
- Gerges, S.H.; Wahdan, S.A.; Elsherbiny, D.A.; El-Demerdash, E. Diosmin ameliorates inflammation, insulin resistance, and fibrosis in an experimental model of non-alcoholic steatohepatitis in rats. Toxicol. Appl. Pharmacol. 2020, 401, 115101. [Google Scholar] [CrossRef]
- Ali, T.M.; Abo-Salem, O.M.; El Esawy, B.H.; El Askary, A. The potential protective effects of diosmin on streptozotocin-induced diabetic cardiomyopathy in rats. Am. J. Med. Sci. 2020, 359, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Dholakiya, S.L.; Benzeroual, K.E. Protective effect of diosmin on LPS-induced apoptosis in PC12cells and inhibition of TNFalpha expression. Toxicol. In Vitro 2011, 25, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-B.; Lee, W.S.; Shin, J.-S.; Jang, D.S.; Lee, K.T. Xanthotoxin suppresses LPS—Induced expression of iNOS, COX-2, TNFalpha, and IL-6 via AP-1, NFkbeta, and JAK-STAT inactivation in RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 49, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Berköz, M. Diosmin suppresses the proinflammatory mediators in lipopolysaccharide-induced RAW264.7 macrophages via NF-κB and MAPKs signal pathways. Gen. Physiol. Biophys. 2019, 38, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.-W.; Zhu, B.T. Strong activation of cyclooxygenase I and II catalytic activity by dietary bioflavonoids. J. Lipid Res. 2008, 49, 2557–2570. [Google Scholar] [CrossRef] [Green Version]
- Zaragozá, C.; Villaescusa, L.; Monserrat, J.; Zaragozá, F.; Álvarez-Mon, M. Potential Therapeutic Anti-Inflammatory and Immunomodulatory Effects of Dihydroflavones, Flavones, and Flavonols. Molecules 2020, 25, 1017. [Google Scholar] [CrossRef] [Green Version]
- Carballo-Villalobos, A.I.; González-Trujano, M.E.; Pellicer, F.; Alvarado-Vásquez, N.; López-Muñoz, F.J. Central and peripheral anti-hyperalgesic effects of diosmin in a neuropathic pain model in rats. Biomed. Pharmacother. 2018, 97, 310–320. [Google Scholar] [CrossRef]
- Herouy, Y.; Pornschlegel, G.; Stetter, C.; Grenz, H.; Schöpf, E.; Norgauer, J.; Vanscheidt, W.; May, A.E.; Preissner, K.T. Lipodermatosclerosis is characterised by elevated expression and activation of matrix metalloproteinase: Implication for venous ulcer formation. J. Investig. Dermatol. 1998, 111, 822–827. [Google Scholar] [CrossRef]
- Kowalewski, R.; Sobolewski, K.; Wolanska, M.; Gacko, M. Matrix metalloproteinases in the tein vall. Int. Angiol. 2004, 23, 164–169. [Google Scholar]
- López-Posada Sayer, G.; Smith, P.D. Immunocytochemical characterisation of the inflammatory cell infiltrate of varicose veins. Eur. J. Vasc. Endovasc. Surg. 2010, 40, 754–765. [Google Scholar]
- Chu, H.B.; Yan, F.; Zhao, J.H.; Xu, Y.B.; Wang, T.; Guo, W.J. Assessment of the infiltration of inflammatory cells in the walls of thrombotic varicose veins. Angiology 2012, 64, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Badier –Commander, C.; Couvelard, A.; Henin, D.; Verbeuren, T.; Michel, J.B.; Jacob, M.P. Smooth muscle cell modulation and cytokine overproduction in varocose veins. An in situ study. J. Pathol. 2001, 193, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, M.M.; Khallil, R.A. Matrix metalloproteinase inhibitors as investigative tools in the pathogenesis amnagement of vascular disease. EXS 2012, 103, 209–279. [Google Scholar] [PubMed] [Green Version]
- Herouy, Y.; Mellios, P.; Bandemir, E.; Dichmann, S.; Nockowski, P.; Schöpf, E.; Norgauer, J. Inflammation in stasis dermatitis upregulates MMP-1 MMP-2 and MMP-13 expression. J. Dermatol. Sci. 2001, 25, 198–205. [Google Scholar] [CrossRef]
- Skiles, J.W.; Gonnella, N.C.; Jeng, A.Y. The design, stucture and therapeutic application of matrix metalloproteinase inhibitors. Curr. Med. Chem. 2001, 8, 425–474. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, J.-K.; Choi, J.; Jang, H.; Seol, J.W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Anti-inflammatory effects of quercetin and galangin in LPS-stimulated RAW246. 7 macrophages and DNCB-induced atopic dermatitis animal model. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar]
- Dymarska, M.; Janeczko, T.; Kostrzewa-Susłow, E. Glycosylation of methoxylated flavonoids in the cultures of Isaria fumosorosea KCH J2. Molecules 2018, 23, 2578. [Google Scholar] [CrossRef] [Green Version]
- Cassidy, A.; Minihane, A.-H. The role of metabolism (and microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nurt. 2017, 105, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Grossman, R.M.; Krueger, J.; Yourish, D.; Granelli-Piperno, A.; Murphy, D.P.; May, L.T.; Kupper, T.S.; Sehgal, P.B.; Gottlieb, A.B. Interleukin 6 is expressed in high levels in psoriatic skin and stimulates proliferation of cultured human keratinocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 6367–6371. [Google Scholar] [CrossRef] [Green Version]
- Gallucci, R.M.; Simeonova, P.P.; Matheson, J.M.; Kommineni, C.; Guriel, J.L.; Sugawara, T.; Luster, M.I. Impaired cutaneous wound healing in interleukin-6-deficient and immunosuppressed mice. FASEB 2000, 14, 2525–2531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dresler, S.; Rutkowska, E.; Bednarek, W.; Stanisławski, G.; Kubrak, T.; Bogucka-Kocka, A.; Wójcik, M. Selected secondary metabolites in Echium vulgare L. populations from nonmetalliferous and metalliferous areas. Phytochemistry 2017, 133, 4–14. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feldo, M.; Wójciak, M.; Ziemlewska, A.; Dresler, S.; Sowa, I. Modulatory Effect of Diosmin and Diosmetin on Metalloproteinase Activity and Inflammatory Mediators in Human Skin Fibroblasts Treated with Lipopolysaccharide. Molecules 2022, 27, 4264. https://doi.org/10.3390/molecules27134264
Feldo M, Wójciak M, Ziemlewska A, Dresler S, Sowa I. Modulatory Effect of Diosmin and Diosmetin on Metalloproteinase Activity and Inflammatory Mediators in Human Skin Fibroblasts Treated with Lipopolysaccharide. Molecules. 2022; 27(13):4264. https://doi.org/10.3390/molecules27134264
Chicago/Turabian StyleFeldo, Marcin, Magdalena Wójciak, Aleksandra Ziemlewska, Sławomir Dresler, and Ireneusz Sowa. 2022. "Modulatory Effect of Diosmin and Diosmetin on Metalloproteinase Activity and Inflammatory Mediators in Human Skin Fibroblasts Treated with Lipopolysaccharide" Molecules 27, no. 13: 4264. https://doi.org/10.3390/molecules27134264
APA StyleFeldo, M., Wójciak, M., Ziemlewska, A., Dresler, S., & Sowa, I. (2022). Modulatory Effect of Diosmin and Diosmetin on Metalloproteinase Activity and Inflammatory Mediators in Human Skin Fibroblasts Treated with Lipopolysaccharide. Molecules, 27(13), 4264. https://doi.org/10.3390/molecules27134264