
P2X7 Receptors in Astrocytes: A Switch for Ischemic Tolerance



Abstract
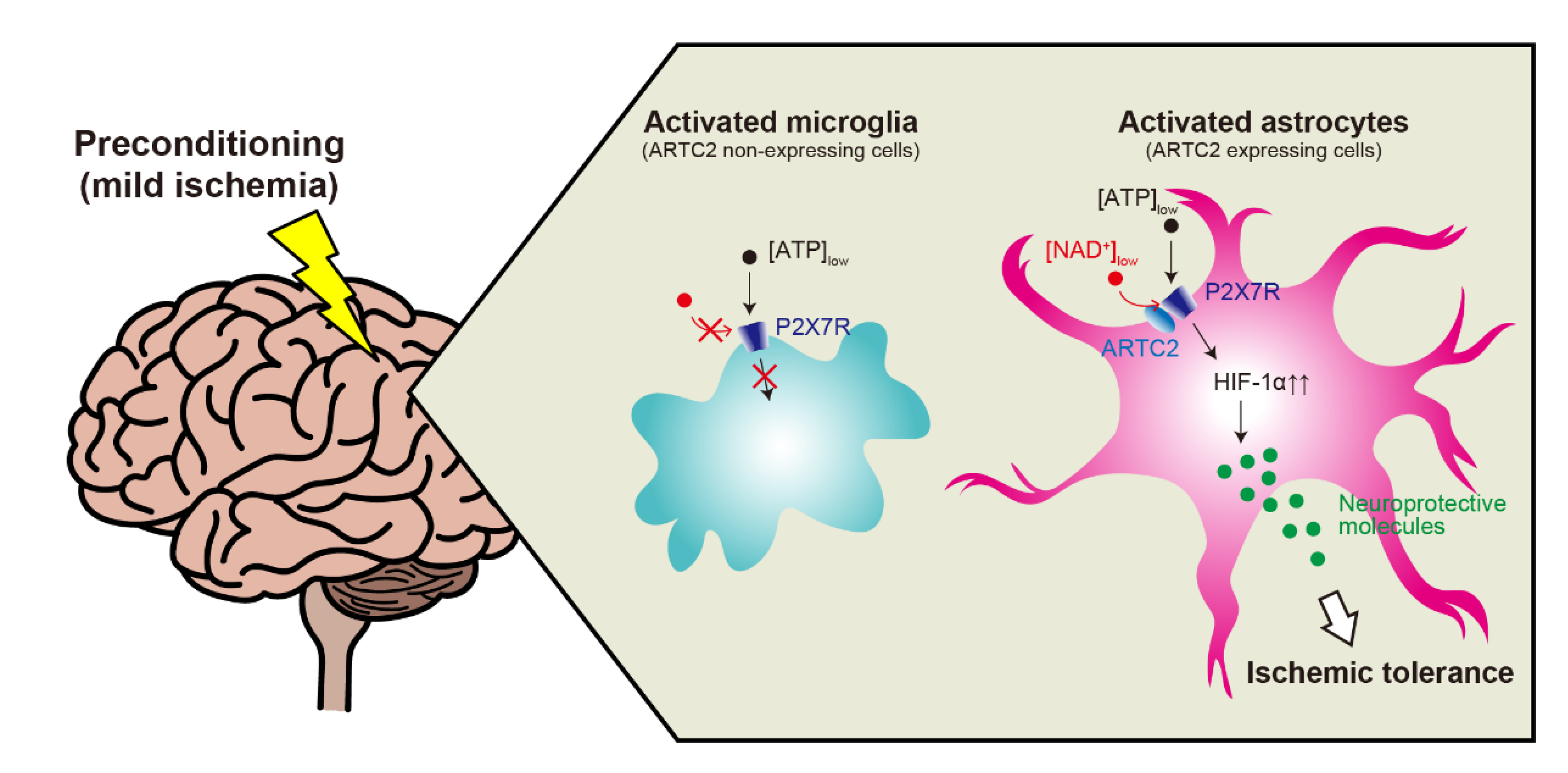
:1. Introduction
2. Localization and Functions of P2X7Rs
3. Astrocytic P2X7R-Mediated Ischemic Tolerance
4. Mechanisms of P2X7R Activation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- O’Collins, V.E.; Macleod, M.R.; Donnan, G.A.; Horky, L.L.; van der Worp, B.H.; Howells, D.W. 1,026 experimental treatments in acute stroke. Ann. Neurol. 2006, 59, 467–477. [Google Scholar] [CrossRef]
- Barres, B.A. The mystery and magic of glia: A perspective on their roles in health and disease. Neuron 2008, 60, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia—A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, E.; Berger, M.; Kussmaul, W.G.; Hirshfeld, J.W.; Herrmann, H.C.; Laskey, W.K. Adaptation to ischemia during percutaneous transluminal coronary angioplasty—Clinical, hemodynamic, and metabolic features. Circulation 1990, 82, 2044–2051. [Google Scholar] [CrossRef] [Green Version]
- Bonventre, J.V. Kidney ischemic preconditioning. Curr. Opin. Nephrol. Hypertens. 2002, 11, 43–48. [Google Scholar] [CrossRef]
- Yadav, S.S.; Sindram, D.; Perry, D.K.; Clavien, P.A. Ischemic preconditioning protects the mouse liver by inhibition of apoptosis through a caspase-dependent pathway. Hepatology 1999, 30, 1223–1231. [Google Scholar] [CrossRef]
- Pang, C.Y.; Yang, R.Z.; Zhong, A.G.; Xu, N.; Boyd, B.; Forrest, C.R. Acute ischemic preconditioning protects against skeletal-muscle infarction in the pig. Cardiovasc. Res. 1995, 29, 782–788. [Google Scholar] [CrossRef]
- Weih, M.; Kallenberg, K.; Bergk, A.; Dirnagl, U.; Harms, L.; Wernecke, K.D.; Einhaupl, K.M. Attenuated stroke severity after prodromal TIA—A role for ischemic tolerance in the brain? Stroke 1999, 30, 1851–1854. [Google Scholar] [CrossRef]
- Kitagawa, K.; Matsumoto, M.; Tagaya, M.; Hata, R.; Ueda, H.; Niinobe, M.; Handa, N.; Fukunaga, R.; Kimura, K.; Mikoshiba, K.; et al. Ischemic tolerance phenomenon found in the brain. Brain Res. 1990, 528, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Dirnagl, U.; Becker, K.; Meisel, A. Preconditioning and tolerance against cerebral ischaemia: From experimental strategies to clinical use. Lancet Neurol. 2009, 8, 398–412. [Google Scholar] [CrossRef] [Green Version]
- Gidday, J.M. Cerebral preconditioning and ischaemic tolerance. Nat. Rev. Neurosci. 2006, 7, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Barres, B. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.Y.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef]
- McDonoug, A.; Weinstein, J.R. The role of microglia in ischemic preconditioning. Glia 2020, 68, 455–471. [Google Scholar] [CrossRef]
- Hirayama, Y.; Ikeda-Matsuo, Y.; Notomi, S.; Enaida, H.; Kinouchi, H.; Koizumi, S. Astrocyte-Mediated Ischemic Tolerance. J. Neurosci. 2015, 35, 3794–3805. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S.; Hirayama, Y.; Morizawa, Y.M. New roles of reactive astrocytes in the brain; an organizer of cerebral ischemia. Neurochem. Int. 2018, 119, 107–114. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Signalling: Therapeutic Developments. Front. Pharmacol. 2017, 8, 661. [Google Scholar] [CrossRef] [Green Version]
- Sarafoff, N.; Byrne, R.A.; Sibbing, D. Clinical Use of Clopidogrel. Curr. Pharm. Des. 2012, 18, 5224–5239. [Google Scholar] [CrossRef]
- Khakh, B.S.; North, R.A. Neuromodulation by Extracellular ATP and P2X Receptors in the CNS. Neuron 2012, 76, 51–69. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.M.; Neary, J.T. P2X(7) receptors: Properties and relevance to CNS function. Glia 2006, 54, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Sperlagh, B.; Illes, P. P2X7 receptor: An emerging target in central nervous system diseases. Trends Pharmacol. Sci. 2014, 35, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Kopp, R.; Krautloher, A.; Ramirez-Fernandez, A.; Nicke, A. P2X7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef]
- Suzuki, T.; Hide, I.; Ido, K.; Kohsaka, S.; Inoue, K.; Nakata, Y. Production and release of neuroprotective tumor necrosis factor by P2X(7) receptor-activated microglia. J. Neurosci. 2004, 24, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pelegrin, P.; Surprenant, A. Pannexin-1 mediates large pore formation and interleukin-1 beta release by the ATP-gated P2X(7) receptor. EMBO J. 2006, 25, 5071–5082. [Google Scholar] [CrossRef] [Green Version]
- Adinolfi, E.; Raffaghello, L.; Giuliani, A.L.; Cavazzini, L.; Capece, M.; Chiozzi, P.; Bianchi, G.; Kroemer, G.; Pistoia, V.; Di Virgilio, F. Expression of P2X7 Receptor Increases In Vivo Tumor Growth. Cancer Res. 2012, 72, 2957–2969. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Arcuino, G.; Takano, T.; Lin, J.; Peng, W.G.; Wan, P.L.; Li, P.J.; Xu, Q.W.; Liu, Q.S.; Goldman, S.A.; et al. P2X7 receptor inhibition improves recovery after spinal cord injury. Nat. Med. 2004, 10, 821–827. [Google Scholar] [CrossRef]
- Leeson, H.C.; Kasherman, M.A.; Chan-Ling, T.; Lovelace, M.D.; Brownlie, J.C.; Toppinen, K.M.; Gu, B.J.; Weible, M.W. P2X7 Receptors Regulate Phagocytosis and Proliferation in Adult Hippocampal and SVZ Neural Progenitor Cells: Implications for Inflammation in Neurogenesis. Stem Cells 2018, 36, 1764–1777. [Google Scholar] [CrossRef] [Green Version]
- Iwata, M.; Ota, K.T.; Li, X.Y.; Sakaue, F.; Li, N.X.; Dutheil, S.; Banasr, M.; Duric, V.; Yamanashi, T.; Kaneko, K.; et al. Psychological Stress Activates the Inflammasome via Release of Adenosine Triphosphate and Stimulation of the Purinergic Type 2X7 Receptor. Biol. Psychiatry 2016, 80, 12–22. [Google Scholar] [CrossRef]
- Young, C.N.J.; Gorecki, D.C. P2RX7 Purinoceptor as a Therapeutic Target-The Second Coming? Front. Chem. 2018, 6, 248. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.Y.; Li, A.P. P2X(7) receptors in cerebral ischemia. Neurosci. Bull. 2013, 29, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Solle, M.; Labasi, J.; Perregaux, D.G.; Stam, E.; Petrushova, N.; Koller, B.H.; Griffiths, R.J.; Gabel, C.A. Altered cytokine production in mice lacking P2X(7) receptors. J. Biol. Chem. 2001, 276, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Takahashi, E.; Miyagawa, Y.; Yamanaka, H.; Noguchi, K. Induction of the P2X7 receptor in spinal microglia in a neuropathic pain model. Neurosci. Lett. 2011, 504, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Mahi, N.; Kumar, A.; Jaggi, A.S.; Singh, N.; Dhawan, R. Possible role of pannexin 1/P(2)x(7) purinoceptor in neuroprotective mechanism of ischemic postconditioning in mice. J. Surg. Res. 2015, 196, 190–199. [Google Scholar] [CrossRef]
- Matute, C.; Torre, I.; Perez-Cerda, F.; Perez-Samartin, A.; Alberdi, E.; Etxebarria, E.; Arranz, A.M.; Ravid, R.; Rodriguez-Antiguedad, A.; Saanchez-Goomez, M.V.; et al. P2X(7) receptor blockade prevents ATP excitotoxicity in oligodendrocytes and ameliorates experimental autoimmune encephalomyelitis. J. Neurosci. 2007, 27, 9525–9533. [Google Scholar] [CrossRef]
- Apolloni, S.; Amadio, S.; Parisi, C.; Matteucci, A.; Potenza, R.L.; Armida, M.; Popoli, P.; D’Ambrosi, N.; Volonte, C. Spinal cord pathology is ameliorated by P2X7 antagonism in a SOD1-mutant mouse model of amyotrophic lateral sclerosis. Dis. Model. Mech. 2014, 7, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Carmo, M.R.S.; Menezes, A.P.F.; Nunes, A.C.L.; Pliassova, A.; Rolo, A.P.; Palmeira, C.M.; Cunha, R.A.; Canas, P.M.; Andrade, G.M. The P2X7 receptor antagonist Brilliant Blue G attenuates contralateral rotations in a rat model of Parkinsonism through a combined control of synaptotoxicity, neurotoxicity and gliosis. Neuropharmacology 2014, 81, 142–152. [Google Scholar] [CrossRef]
- Diaz-Hernandez, J.I.; Gomez-Villafuertes, R.; Leon-Otegui, M.; Hontecillas-Prieto, L.; del Puerto, A.; Trejo, J.L.; Lucas, J.J.; Garrido, J.J.; Gualix, J.; Miras-Portugal, M.T.; et al. In vivo P2X7 inhibition reduces amyloid plaques in Alzheimer’s disease through GSK3 beta and secretases. Neurobiol. Aging 2012, 33, 1816–1828. [Google Scholar] [CrossRef] [Green Version]
- Chessell, I.P.; Hatcher, J.P.; Bountra, C.; Michel, A.D.; Hughes, J.P.; Green, P.; Egerton, J.; Murfin, M.; Richardson, J.; Peck, W.L.; et al. Disruption of the P2X(7) purinoceptor gene abolishes chronic inflammatory and neuropathic pain. Pain 2005, 114, 386–396. [Google Scholar] [CrossRef]
- Honore, P.; Donnelly-Roberts, D.; Namovic, M.T.; Hsieh, G.; Zhu, C.Z.; Mikusa, J.P.; Hernandez, G.; Zhong, C.M.; Gauvin, D.M.; Chandran, P.; et al. A-740003 N-(1-{(cyanoimino)(5-quinolinylamino)methyl amino}-2,2-dimethylpropyl)-2-(3,4-dimethoxyphenyl)acetamides, a novel and selective P2X(7) receptor antagonist, dose-dependently reduces neuropathic pain in the rat. J. Pharmacol. Exp. Ther. 2006, 319, 1376–1385. [Google Scholar] [CrossRef] [Green Version]
- Engel, T.; Gomez-Villafuertes, R.; Tanaka, K.; Mesuret, G.; Sanz-Rodriguez, A.; Garcia-Huerta, P.; Miras-Portugal, M.T.; Henshall, D.C.; Diaz-Hernandez, M. Seizure suppression and neuroprotection by targeting the purinergic P2X7 receptor during status epilepticus in mice. FASEB J. 2012, 26, 1616–1628. [Google Scholar] [CrossRef]
- Diaz-Hernandez, M.; Diez-Zaera, M.; Sanchez-Nogueiro, J.; Gomez-Villafuertes, R.; Canals, J.M.; Alberch, J.; Miras-Portugal, M.T.; Lucas, J.J. Altered P2X7-receptor level and function in mouse models of Huntington’s disease and therapeutic efficacy of antagonist administration. FASEB J. 2009, 23, 1893–1906. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Masuch, A.; Shieh, C.H.; van Rooijen, N.; van Calker, D.; Biber, K. Mechanism of Microglia Neuroprotection: Involvement of P2X7, TNF alpha, and Valproic Acid. Glia 2016, 64, 76–89. [Google Scholar] [CrossRef]
- Ortega, F.; Perez-Sen, R.; Delicado, E.G.; Miras-Portugal, M.T. ERK1/2 activation is involved in the neuroprotective action of P2Y(13) and P2X7 receptors against glutamate excitotoxicity in cerebellar granule neurons. Neuropharmacology 2011, 61, 1210–1221. [Google Scholar] [CrossRef]
- Ortega, F.; Perez-Sen, R.; Morente, V.; Delicado, E.G.; Miras-Portugal, M.T. P2X7, NMDA and BDNF receptors converge on GSK3 phosphorylation and cooperate to promote survival in cerebellar granule neurons. Cell. Mol. Life Sci. 2010, 67, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- Ikeda-Matsuo, Y.; Ota, A.; Fukada, T.; Uematsu, S.; Akira, S.; Sasaki, Y. Microsomal prostaglandin E synthase-1 is a critical factor of stroke-reperfusion injury. Proc. Natl. Acad. Sci. USA 2006, 103, 11790–11795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, J.L.; Xu, L.J.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic Analysis of Reactive Astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trendelenburg, G.; Dirnagl, U. Neuroprotective role of astrocytes in cerebral ischemia: Focus on ischemic preconditioning. Glia 2005, 50, 307–320. [Google Scholar] [CrossRef]
- Hoshi, A.; Yamamoto, T.; Shimizu, K.; Sugiura, Y.; Ugawa, Y. Chemical preconditioning-induced reactive astrocytosis contributes to the reduction of post-ischemic edema through aquaporin-4 downregulation. Exp. Neurol. 2011, 227, 89–95. [Google Scholar] [CrossRef]
- Hassel, B.; Paulsen, R.E.; Johnsen, A.; Fonnum, F. Selective-Inhibition of glial-cell metabolism invivo by fluorocitrate. Brain Res. 1992, 576, 120–124. [Google Scholar] [CrossRef]
- Franke, H.; Illes, P. Involvement of P2 receptors in the growth and survival of neurons in the CNS. Pharmacol. Ther. 2006, 109, 297–324. [Google Scholar] [CrossRef]
- Le Feuvre, R.; Brough, D.; Rothwell, N. Extracellular ATP and P2X7 receptors in neurodegeneration. Eur. J. Pharmacol. 2002, 447, 261–269. [Google Scholar] [CrossRef]
- Le Feuvre, R.A.; Brough, D.; Touzani, O.; Rothwell, N.J. Role of P2X(7) receptors in ischemic and excitotoxic brain injury in vivo. J. Cereb. Blood Flow Metab. 2003, 23, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Dirnagl, U.; Simon, R.P.; Hallenbeck, J.M. Ischemic tolerance and endogenous neuroprotection. Trends Neurosci. 2003, 26, 248–254. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-Inducible Factor 1 and Cardiovascular Disease. Annu. Rev. Physiol. 2014, 76, 39–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prass, K.; Scharff, A.; Ruscher, K.; Lowl, D.; Muselmann, C.; Victorov, I.; Kapinya, K.; Dirnagl, U.; Meisel, A. Hypoxia-induced stroke tolerance in the mouse is mediated by erythropoietin. Stroke 2003, 34, 1981–1986. [Google Scholar] [CrossRef] [Green Version]
- Digicaylioglu, M.; Lipton, S.A. Erythropoietin-mediated neuroprotection involves cross-talk between Jak2 and NF-kappa B signalling cascades. Nature 2001, 412, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, F.; Falzoni, S.; Adinolfi, E.; Ferrari, D.; Di Virgilio, F. The P2X7 receptor is a key modulator of aerobic glycolysis. Cell Death Dis. 2012, 3, e370. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, Y.; Koizumi, S. Hypoxia-Independent Mechanisms of HIF-1 alpha Expression in Astrocytes after Ischemic Preconditioning. Glia 2017, 65, 523–530. [Google Scholar] [CrossRef]
- Tikka, T.M.; Koistinaho, J.E. Minocycline provides neuroprotection against N-methyl-D-aspartate neurotoxicity by inhibiting microglia. J. Immunol. 2001, 166, 7527–7533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, Y.; Anzai, N.; Koizumi, S. Mechanisms underlying sensitization of P2X7 receptors in astrocytes for induction of ischemic tolerance. Glia 2021, 69, 2100–2110. [Google Scholar] [CrossRef]
- Hong, S.Y.; Schwarz, N.; Brass, A.; Seman, M.; Haag, F.; Koch-Nolte, F.; Schilling, W.P.; Dubyak, G.R. Differential Regulation of P2X(7) Receptor Activation by Extracellular Nicotinamide Adenine Dinucleotide and Ecto-ADP-Ribosyltransferases in Murine Macrophages and T Cells. J. Immunol. 2009, 183, 578–592. [Google Scholar] [CrossRef] [Green Version]
- Seman, M.; Adriouch, S.; Scheuplein, F.; Krebs, C.; Freese, D.; Glowacki, G.; Deterre, P.; Haag, F.; Koch-Nolte, F. NAD-induced T cell death: ADP-ribosylation of cell surface proteins by ART2 activates the cytolytic P2X7 purinoceptor. Immunity 2003, 19, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Rissiek, B.; Haag, F.; Boyer, O.; Koch-Nolte, F.; Adriouch, S. P2X7 on mouse T cells: One channel, many functions. Front. Immunol. 2015, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Bannas, P.; Well, L.; Lenz, A.; Rissiek, B.; Haag, F.; Schmid, J.; Hochgrafe, K.; Trepel, M.; Adam, G.; Ittrich, H.; et al. In vivo near-infrared fluorescence targeting of T cells: Comparison of nanobodies and conventional monoclonal antibodies. Contrast Media Mol. Imaging 2014, 9, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahl, S.; Nissen, M.; Girisch, R.; Duffy, T.; Leiter, E.H.; Haag, F.; Koch-Nolte, F. Metalloprotease-mediated shedding of enzymatically active mouse ecto-ADP-ribosyltransferase ART2.2 upon T cell activation. J. Immunol. 2000, 165, 4463–4469. [Google Scholar] [CrossRef] [PubMed]
- Rissiek, B.; Stabernack, J.; Cordes, M.; Duan, Y.H.; Behr, S.; Menzel, S.; Magnus, T.; Koch-Nolte, F. Astrocytes and Microglia Are Resistant to NAD(+)-Mediated Cell Death Along the ARTC2/P2X7 Axis. Front. Mol. Neurosci. 2020, 12, 330. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, N.; Drouot, L.; Nicke, A.; Fliegert, R.; Boyer, O.; Guse, A.H.; Haag, F.; Adriouch, S.; Koch-Nolte, F. Alternative Splicing of the N-Terminal Cytosolic and Transmembrane Domains of P2X7 Controls Gating of the Ion Channel by ADP-Ribosylation. PLoS ONE 2012, 7, e41269. [Google Scholar] [CrossRef]
- Kido, Y.; Kawahara, C.; Terai, Y.; Ohishi, A.; Kobayashi, S.; Hayakawa, M.; Kamatsuka, Y.; Nishida, K.; Nagasawa, K. Regulation of Activity of P2X7 Receptor by Its Splice Variants in Cultured Mouse Astrocytes. Glia 2014, 62, 440–451. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Roles | Pathology (In Vivo Model) | Findings | Ref. |
|---|---|---|---|
| Protective | Cerebral ischemic tolerance by preconditioning (MCAO) | Cerebral ischemic tolerance is abolished in P2X7R knock-out mice | [15] |
| Cerebral ischemic tolerance by postconditioning (BCAO) | Ischemic postconditioning-induced neuroprotective effects are abolished by pretreatment of pannexin 1/P2X7R antagonist mefloquine | [33] | |
| Harmful | Multiple sclerosis (EAE) | BBG or oATP ameliorates chronic EAE by reducing demyelination | [34] |
| ALS (SOD1-G93A mice) | BBG attenuates motor neuron loss in SOD1-G93A mice | [35] | |
| Parkinson’s disease (6-OHDA) | BBG attenuates the 6-OHDA-induced neurotoxicity | [36] | |
| Alzheimer’s disease (hAPP-J20 mice) | BBG prevents the development of amyloid plaques in hAPP-J20 mice | [37] | |
| Neuropathic pain (SNI, PSL, and SNL) | P2X7R antagonist A-438079 suppresses the development of mechanical hypersensitivity in SNI model | [32] | |
| Development of both thermal and mechanical hypersensitivity after PSL is absent in P2X7R knock-out mice | [38] | ||
| P2X7R antagonist A-740003 reduces SNL-induced mechanical allodynia | [39] | ||
| Status epilepticus (KA) | BBG or P2X7R antagonist A438079 protects against KA-induced neuronal death | [40] | |
| Huntington’s disease (R6/1 mice) | Administration of BBG to R6/1 mice attenuates their motor-coordination deficit | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirayama, Y.; Anzai, N.; Kinouchi, H.; Koizumi, S. P2X7 Receptors in Astrocytes: A Switch for Ischemic Tolerance. Molecules 2022, 27, 3655. https://doi.org/10.3390/molecules27123655
Hirayama Y, Anzai N, Kinouchi H, Koizumi S. P2X7 Receptors in Astrocytes: A Switch for Ischemic Tolerance. Molecules. 2022; 27(12):3655. https://doi.org/10.3390/molecules27123655
Chicago/Turabian StyleHirayama, Yuri, Naohiko Anzai, Hiroyuki Kinouchi, and Schuichi Koizumi. 2022. "P2X7 Receptors in Astrocytes: A Switch for Ischemic Tolerance" Molecules 27, no. 12: 3655. https://doi.org/10.3390/molecules27123655
APA StyleHirayama, Y., Anzai, N., Kinouchi, H., & Koizumi, S. (2022). P2X7 Receptors in Astrocytes: A Switch for Ischemic Tolerance. Molecules, 27(12), 3655. https://doi.org/10.3390/molecules27123655