
Natural Antioxidant Evaluation: A Review of Detection Methods


 , , , ,
, , , , 
Abstract
1. Introduction
2. Natural Antioxidants
2.1. Carotenoids
2.2. Vitamin E
2.3. Ascorbic Acid
2.4. Phenolic Compounds
Phenolic Compounds Quantification
3. First Issue: Common Antioxidant Activity Methods In Vitro
3.1. DPPH
3.2. TEAC/ABTS
3.3. ORAC
3.4. FRAP and TRAP Assays
3.5. DPPP
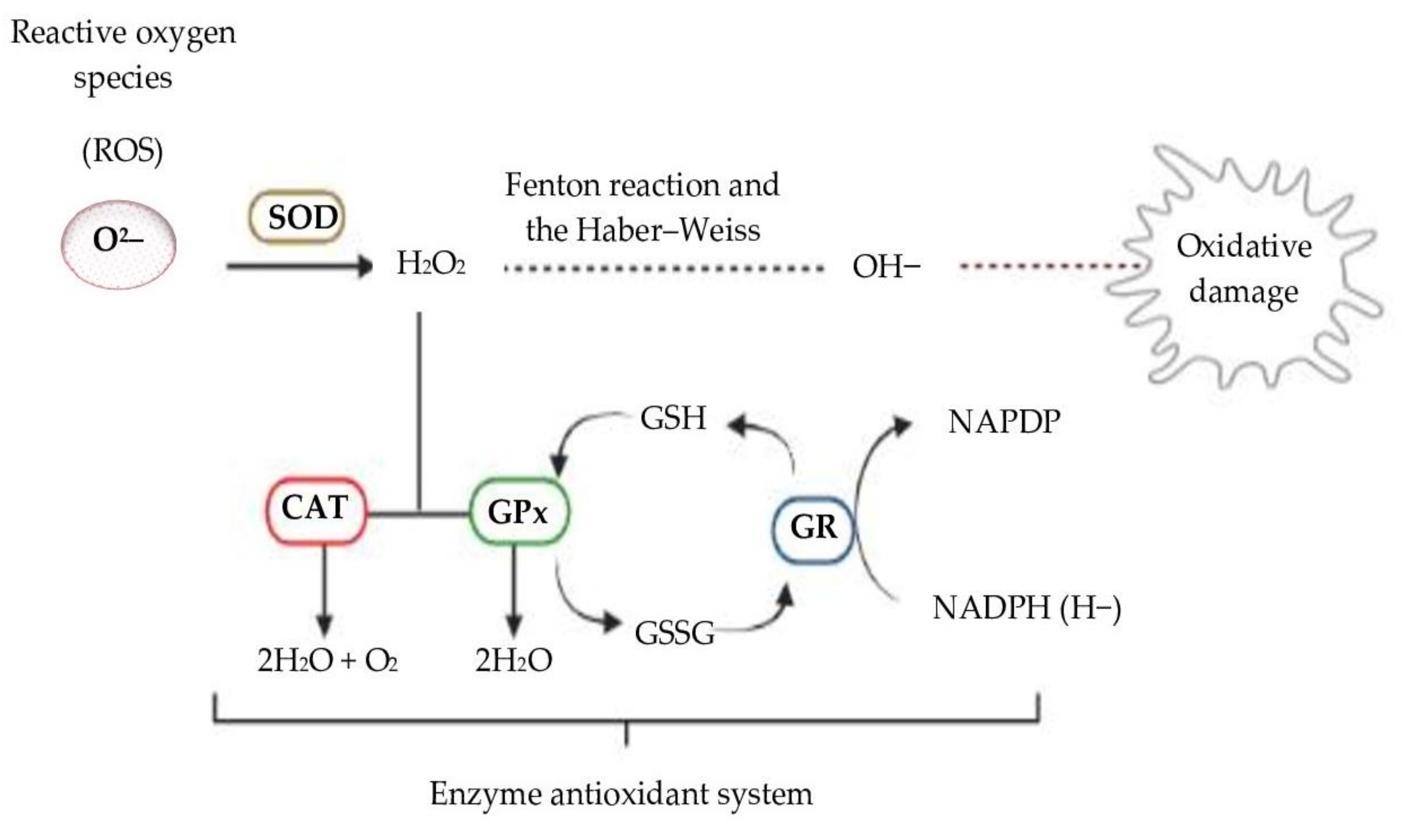
4. Second Issue: Evaluation of Antioxidant Activity in Biological Systems—The Role of Enzymes in Redox Cellular Signaling
4.1. Reduced Glutathione
4.2. Glutathione Peroxidase
4.3. Superoxide Dismutase
4.4. Catalase
4.5. Thiobarbituric Acid-Reactive Substances
5. Third Issue: Bioavailability of Natural Antioxidants and the Role of Their Antioxidant Activity in Biological Systems
5.1. Polyphenols
5.2. Vitamins C and E
5.3. Carotenoids
6. Data Interpretation
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Amorati, R.; Valgimigli, L. Methods to Measure the Antioxidant Activity of Phytochemicals and Plant Extracts. J. Agric. Food Chem. 2018, 66, 3324–3329. [Google Scholar] [CrossRef]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Liaudanskas, M.; Žvikas, V.; Petrikaitė, V. The Potential of Dietary Antioxidants from a Series of Plant Extracts as Anticancer Agents against Melanoma, Glioblastoma, and Breast Cancer. Antioxidants 2021, 10, 1115. [Google Scholar] [CrossRef]
- Lu, Q.Y.; Summanen, P.H.; Lee, R.P.; Huang, J.; Henning, S.M.; Heber, D.; Finegold, S.M.; Li, Z. Prebiotic Potential and Chemical Composition of Seven Culinary Spice Extracts. J. Food Sci. 2017, 82, 1807–1813. [Google Scholar] [CrossRef]
- Fu, L.; Xu, B.T.; Gan, R.Y.; Zhang, Y.; Xu, X.R.; Xia, E.Q.; Li, H.B. Total phenolic contents and antioxidant capacities of herbal and tea infusions. Int. J. Mol. Sci. 2011, 12, 2112–2124. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.F.; Xu, X.R.; Guo, Y.J.; Xia, E.Q.; Li, S.; Wu, S.; Chen, F.; Ling, W.H.; Li, H.B. Determination of antioxidant property and their lipophilic and hydrophilic phenolic contents in cereal grains. J. Funct. Foods 2012, 4, 906–914. [Google Scholar] [CrossRef]
- Cömert, E.D.; Gökmen, V. Evolution of food antioxidants as a core topic of food science for a century. Food Res. Int. 2018, 105, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Harasym, J.; Oledzki, R. Effect of fruit and vegetable antioxidants on total antioxidant capacity of blood plasma. Nutrition 2014, 30, 511–517. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed]
- Sunkara, A.; Raizner, A. Supplemental Vitamins and Minerals for Cardiovascular Disease Prevention and Treatment. Methodist Debakey Cardiovasc. J. 2019, 15, 179–184. [Google Scholar] [CrossRef] [PubMed]
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Wachtel-Galor, S. Increasing the antioxidant content of food: A personal view on whether this is possible or desirable. Int. J. Food Sci. Nutr. 2012, 63, 62–70. [Google Scholar] [CrossRef]
- Guo, Q.; Li, F.; Duan, Y.; Wen, C.; Wang, W.; Zhang, L.; Huang, R.; Yin, Y. Oxidative stress, nutritional antioxidants and beyond. Sci. China Life Sci. 2020, 63, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Abramovič, H.; Grobin, B.; Ulrih, N.P.; Cigić, B. Relevance and Standardization of In vitro Antioxidant Assays: ABTS, DPPH, and Folin–Ciocalteu. J. Chem. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Y. Mass spectrometry for the assessment of the occurrence and biological consequences of DNA adducts. Chem. Soc. Rev. 2015, 44, 7829–7854. [Google Scholar] [CrossRef]
- Apak, R. Current Issues in Antioxidant Measurement. J. Agric. Food Chem. 2019, 67, 9187–9202. [Google Scholar] [CrossRef]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant Activity/Capacity Measurement. 2. Hydrogen Atom Transfer (HAT)-Based, Mixed-Mode (Electron Transfer (ET)/HAT), and Lipid Peroxidation Assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef]
- Fraga, C.G.; Oteiza, P.I.; Galleano, M. In vitro measurements and interpretation of total antioxidant capacity. Biochim Biophys Acta. 2014, 1840, 931–934. [Google Scholar] [CrossRef]
- Istas, G.; Wood, E.; Le Sayec, M.; Rawlings, C.; Yoon, J.; Dandavate, V.; Cera, D.; Rampelli, S.; Costabile, A.; Fromentin, E.; et al. Effects of aronia berry (poly)phenols on vascular function and gut microbiota: A double-blind randomized controlled trial in adult men. Am. J. Clin. Nutr. 2019, 110, 316–329. [Google Scholar] [CrossRef]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J. Bioactive Properties of Carotenoids in Human Health. Nutrients 2019, 11, 2388. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and Chlorophylls as Antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, I. Advances in photosynthesis and respiration. Photosynth. Res. 2012, 32, 327–329. [Google Scholar] [CrossRef][Green Version]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Plant Metab. Synth. Biol. 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Widomska, J.; Gruszecki, W.I.; Subczynski, W.K. Factors Differentiating the Antioxidant Activity of Macular Xanthophylls in the Human Eye Retina. Antioxidants 2021, 10, 601. [Google Scholar] [CrossRef] [PubMed]
- Duda, M.; Cygan, K.; Wisniewska-Becker, A. Effects of Curcumin on Lipid Membranes: An EPR Spin-label Study. Cell Biochem. Biophys. 2020, 78, 139–147. [Google Scholar] [CrossRef]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxid. Med. Cell Longev. 2019, 8267234. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative stress and Parkinson’s disease. Front. Neuroanat. 2015, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1842, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Amaya, D.B. Status of carotenoid analytical methods and in vitro assays for the assessment of food quality and health effects. Curr. Opin. Food Sci. 2015, 1, 56–63. [Google Scholar]
- Arunkumar, R.; Gorusupudi, A.; Bernstein, P.S. The macular carotenoids: A biochemical overview. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158617. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Chantrell, S.J.; McAuliffe, C.A.; Munn, R.W.; Pratt, A.C.; Land, E.J. Excited states of protoporphyrin IX dimethyl ester: Reaction on the triplet with carotenoids. J. Chem. Soc. Faraday Trans. Phys. Chem. Condens. Phases 1977, 73, 858–865. [Google Scholar] [CrossRef]
- Jeong, Y.; Lim, J.W.; Kim, H. Lycopene Inhibits Reactive Oxygen Species-Mediated NF-κB Signaling and Induces Apoptosis in Pancreatic Cancer Cells. Nutrients 2019, 11, 762. [Google Scholar] [CrossRef]
- Heymann, T.; Heinz, P.; Glomb, M.A. Lycopene inhibits the isomerization of β-carotene during quenching of singlet oxygen and free radicals. J. Agric. Food Chem. 2015, 63, 3279–3287. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Moran, N.A.; Jarvik, T. Lateral transfer of genes from fungi underlies carotenoid production in aphids. Science 2010, 5978, 624–627. [Google Scholar] [CrossRef]
- Saini, R.K.; Nile, S.H.; Park, S.W. Carotenoids from fruits and vegetables: Chemistry, analysis, occurrence, bioavailability and biological activities (Part 3). Food Res. Int. 2015, 76, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 1–28. [Google Scholar] [CrossRef]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Alternative and efficient extraction methods for marine-derived compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Ahmad, S.; Ahmad, A. Green extraction methods and environmental applications of carotenoids-a review. RSC Adv. 2015, 77, 62358–62393. [Google Scholar] [CrossRef]
- Singh, D.; Barrow, C.J.; Mathur, A.S.; Tuli, D.K.; Puri, M. Optimization of zeaxanthin and β-carotene extraction from Chlorella saccharophila isolated from New Zealand marine waters. Biocatal. Agric. Biotechnol. 2015, 2, 166–173. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Progress in microbial carotenoids production. Indian J. Microbiol. 2017, 57, 129–130. [Google Scholar] [CrossRef]
- Alfonsi, K.; Colberg, J.; Dunn, P.J.; Fevig, T.; Jennings, S.; Johnson, T.A.; Stefaniak, M. Green chemistry tools to influence a medicinal chemistry and research chemistry based organisation. Green Chem. 2008, 10, 31–36. [Google Scholar] [CrossRef]
- Capello, C.; Fischer, U.; Hungerbühler, K. What is a green solvent? A comprehensive framework for the environmental assessment of solvents. Green Chem. 2007, 9, 927–934. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Casadesús, A.; Arabia, A.; Pujolriu, R.; Munné-Bosch, S. Differential accumulation of tocochromanols in photosynthetic and non-photosynthetic tissues of strawberry plants subjected to reiterated water deficit. Plant Physiol. Biochem. 2020, 155, 868–876. [Google Scholar] [CrossRef]
- Drotleff, A.M.; Bohnsack, C.; Schneider, I.; Hahn, A.; Ternes, W. Human oral bioavailability and pharmacokinetics of tocotrienols from tocotrienol-rich (tocopherol-low) barley oil and palm oil formulations. J. Funct. Foods 2014, 7, 150–160. [Google Scholar] [CrossRef]
- Azzi, A. Tocopherols, tocotrienols and tocomonoenols: Many similar molecules but only one vitamin E. Redox Biol. 2019, 26, 101259. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, P.J.; Weihrauch, J.L. Vitamin E content of foods. J. Am. Diet. Assoc. 1979, 75, 647–665. [Google Scholar] [CrossRef]
- Birringer, M.; Lorkowski, S. Vitamin E: Regulatory role of metabolites. IUBMB Life 2019, 71, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Dietary intake and blood concentrations of antioxidants and the risk of cardiovascular disease, total cancer, and all-cause mortality: A systematic review and dose-response meta-analysis of prospective studies. Am. J. Clin. Nutr. 2018, 108, 1069–1091. [Google Scholar] [CrossRef]
- Mangge, H.; Becker, K.; Fuchs, D.; Gostner, J.M. Antioxidants, inflammation and cardiovascular disease. World J. Cardiol. 2014, 6, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, V.; Roumeliotis, S.; Zarogiannis, S.; Eleftheriadis, T.; Mertens, P.R. Oxidative stress in hemodialysis: Causative mechanisms, clinical implications, and possible therapeutic interventions. Semin. Dial. 2019, 32, 58–71. [Google Scholar] [CrossRef]
- Nicod, N.; Parker, R.S. Vitamin E Secretion by Caco-2 Monolayers to APOA1, but Not to HDL, Is Vitamer Selective. J. Nutr. 2013, 143, 1565–1572. [Google Scholar] [CrossRef]
- Szewczyk, K.; Chojnacka, A.; Górnicka, M. Tocopherols and Tocotrienols-Bioactive Dietary Compounds; What Is Certain, What Is Doubt? Int. J. Mol. Sci. 2021, 22, 6222. [Google Scholar] [CrossRef]
- Wong, W.Y.; Ward, L.C.; Fong, C.W.; Yap, W.N.; Brown, L. Anti-inflammatory γ- and δ-tocotrienols improve cardiovascular, liver and metabolic function in diet-induced obese rats. Eur. J. Nutr. 2017, 56, 133–150. [Google Scholar] [CrossRef]
- Wong, S.Y.; Teo, J.S.M.; Chai, S.F.; Yeap, S.L.; Lau, A.J. Vitamin E analogues differentially inhibit human cytochrome P450 3A (CYP3A)-mediated oxidative metabolism of lithocholic acid: Impact of δ-tocotrienol on lithocholic acid cytotoxicity. Toxicology 2019, 423, 62–74. [Google Scholar] [CrossRef]
- Arrozi, A.P.; Shukri, S.N.S.; Ngah, W.Z.W.; Yusof, Y.A.M.; Damanhuri, M.H.A.; Jaafar, F.; Makpol, S. Comparative effects of alpha-and gamma-tocopherol on mitochondrial functions in Alzheimer’s Disease in vitro model. Sci. Rep. 2020, 10, 1–20. [Google Scholar]
- Pingret, D.; Fabiano-Tixier, A.-S.; Chemat, F. Degradation during application of ultrasound in food processing: A review. Food Control 2013, 2, 593–606. [Google Scholar] [CrossRef]
- Han, R.M.; Zhang, J.P.; Skibsted, L.H. Reaction Dynamics of Flavonoids and Carotenoids as Antioxidants. Molecules 2012, 17, 2140–2160. [Google Scholar] [CrossRef] [PubMed]
- Zerbinati, C.; Caponecchia, L.; Fiori, C.; Sebastianelli, A.; Salacone, P.; Ciacciarelli, M.; Iuliano, L. Alpha- and gamma-tocopherol levels in human semen and their potential functional implications. Andrologia 2020, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- De Andrade Lima, M.; Kestekoglou, I.; Charalampopoulos, D.; Chatzifragkou, A. Supercritical Fluid Extraction of Carotenoids from Vegetable Waste Matrices. Molecules 2019, 24, 466. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.; Palou, A. Chromatographic determination of carotenoids in foods. J. Chromatogr. A 2000, 881, 543–555. [Google Scholar] [CrossRef]
- Xu, Z. Comparison of extraction methods for quantifying vitamin E from animal tissues. Bioresour. Technol. 2008, 18, 1–5. [Google Scholar] [CrossRef]
- Pawlowska, E.; Szczepanska, J.; Blasiak, J. Pro- and Antioxidant Effects of Vitamin C in Cancer in correspondence to Its Dietary and Pharmacological Concentrations. Oxid. Med. Cell Longev. 2019, 7286737. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, Y.; Li, J.; Subirade, M.; Song, Y.; Liang, L. Effect of resveratrol or ascorbic acid on the stability of α-tocopherol in O/W emulsions stabilized by whey protein isolate: Simultaneous encapsulation of the vitamin and the protective antioxidant. Food Chem. 2016, 196, 466–474. [Google Scholar] [CrossRef]
- May, J.M. Vitamin C transport and its role in the central nervous system. Subcell Biochem. 2012, 56, 85–103. [Google Scholar]
- Spector, R.; Johanson, C.E. The nexus of vitamin homeostasis and DNA synthesis and modification in mammalian brain. Mol. Brain 2014, 7, 1–14. [Google Scholar] [CrossRef]
- Kaufman, S. Coenzymes and hydroxylases: Ascorbate and dopamine-beta-hydroxylase; tetrahydropteridines and phenylalanine and tyrosine hydroxylases. Pharmacol. Rev. 1966, 18, 61–69. [Google Scholar] [PubMed]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two Faces of Vitamin C-Antioxidative and Pro-Oxidative Agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta 2012, 826, 443–457. [Google Scholar] [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D. Vitamin C in Human Health and Disease. Nutrients 2021, 13, 1595. [Google Scholar] [CrossRef]
- Salaj, N.; Kladar, N.; Čonić, B.S.; Jeremić, K.; Hitl, M.; Gavarić, N.; Božin, B. Traditional multi-herbal formula in diabetes therapy—Antihyperglycemic and antioxidant potential. Arab. J. Chem. 2021, 14, 10–103347. [Google Scholar] [CrossRef]
- Farag, R.S.; Abdel-Latif, M.S.; Abd El Baky, H.H.; Tawfeek, L.S. Phytochemical screening and antioxidant activity of some medicinal plants’ crude juices. Biotechnol. Rep. 2020, 28, e00536. [Google Scholar] [CrossRef]
- Sarmiento-Salinas, F.L.; Perez-Gonzalez, A.; Acosta-Casique, A.; Ix-Ballote, A.; Diaz, A.; Treviño, S.; Rosas-Murrieta, N.H.; Millán-Perez-Peña, L.; Maycotte, P. Reactive oxygen species: Role in carcinogenesis, cancer cell signaling and tumor progression. Life Sci. 2021, 284, 119942. [Google Scholar] [CrossRef] [PubMed]
- Ghafoor, K.; Juhaimi, F.A.M.; Özcan, M.; Uslu, N.; Babiker, E.E.; Ahmed, I.A.M. Total phenolics, total carotenoids, individual phenolics and antioxidant activity of ginger (Zingiber officinale) rhizome as affected by drying methods. LWT 2020, 126, 109354. [Google Scholar] [CrossRef]
- Mwamatope, B.; Tembo, D.; Chikowe, I.; Kampira, E.; Nyirenda, C. Total phenolic contents and antioxidant activity of Senna singueana, Melia azedarach, Moringa oleifera and Lannea discolor herbal plants. Sci. Afr. 2020, 9, e00481. [Google Scholar] [CrossRef]
- Yu, M.; Gouvinhas, I.; Rocha, J.; Barros, A.I.R.N.A. Phytochemical and antioxidant analysis of medicinal and food plants towards bioactive food and pharmaceutical resources. Sci. Rep. 2021, 11, 10041. [Google Scholar] [CrossRef]
- Yousfi, F.; Abrigach, F.; Petrovic, J.D.; Sokovic, M.; Ramdani, M. Phytochemical screening and evaluation of the antioxidant and antibacterial potential of Zingiber officinale extracts. S. Afr. J. Bot. 2021, 142, 433–440. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Liang, Z.; Zhao, C. New lignans and their biological activities. Chem. Biodivers. 2014, 11, 1–54. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J. Phenolic-enriched foods: Sources and processing for enhanced health benefits. Proc. Nutr. Soc. 2017, 76, 163–171. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef]
- Dei Cas, M.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef]
- Sheng, H.; Sun, X.; Yan, Y.; Yuan, Q.; Wang, J.; Shen, X. Metabolic Engineering of Microorganisms for the Production of Flavonoids. Front. Bioeng. Biotechnol. 2020, 8, 589069. [Google Scholar] [CrossRef]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising natural compounds against viral infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef]
- Wang, T.; Li, Q.; Bi, K. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Panche, N.A.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–11. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Kono, M.; Ito, A.; Ito, M. Anthocyanins in perilla plants and dried leaves. Phytochemistry 2018, 147, 158–166. [Google Scholar] [CrossRef]
- Ozcan, T.; Akpinar-Bayizit, A.; Yilmaz-Ersan, L.; Delikanli, B. Phenolics in Human Health. Int. J. Chem. Eng. Appl. 2014, 5, 393–396. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects- A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Gerstenmeyer, E.; Reimer, S.; Berghofer, E.; Schwartz, H.; Sontag, G. Effect of thermal heating on some lignans in flax seeds, sesame seeds and rye. Food Chem. 2013, 138, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, V.; Mateus, N. Protein/Polyphenol Interactions: Past and Present Contributions. Mechanisms of Astringency Perception. Cur. Org. Chem. 2012, 16, 724–746. [Google Scholar] [CrossRef]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, S.; Kroslakova, I.; Janzon, R.; Mayer, I.; Saake, B.; Pichelin, F. Characterization of condensed tannins and carbohydrates in hot water bark extracts of European softwood species. Phytochemistry 2015, 120, 53–61. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 16. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical Methods for Determining Bioavailability and Bioaccessibility of Bioactive Compounds from Fruits and Vegetables: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- Kim, M.Y.; Yoon, N.; Lee, Y.J.; Woo, K.S.; Kim, H.Y.; Lee, J.; Jeong, H.S. Influence of Thermal Processing on Free and Bound Forms of Phenolics and Antioxidant Capacity of Rice Hull (Oryza sativa L.). Prev. Nutr. Food Sci. 2020, 25, 310–318. [Google Scholar] [CrossRef]
- Kwatra, B. A review on potential properties and therapeutic applications of grape seed extract. World J. Pharm. Res. 2020, 9, 2519–2540. [Google Scholar]
- Khoddami, A.; Wilkes, A.M.; Roberts, H.T. Techniques for Analysis of Plant Phenolic Compounds. Molecules 2013, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Folin, O.; Ciocalteu, V. On Tyrosine and Tryptophane Determinations in Proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Sasikumar, J.M.; Erba, O.; Egigu, M.C. In vitro antioxidant activity and polyphenolic content of commonly used spices from Ethiopia. Heliyon 2020, 6, e05027. [Google Scholar] [CrossRef]
- Bunzel, M.; Schendel, R.R. Determination of (total) phenolics and antioxidant capacity in food and ingredients. In Food Analysis; Springer: Cham, Switzerland, 2017; pp. 455–468. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety-Chemistry, Applications, Strengths, and Limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Ionita, P. The Chemistry of DPPH·Free Radical and Congeners. Int. J. Mol. Sci. 2021, 22, 1545. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Sirivibulkovit, K.; Nouanthavong, S.; Sameenoi, Y. Paper-based DPPH Assay for Antioxidant Activity Analysis. Anal. Sci. 2018, 34, 795–800. [Google Scholar] [CrossRef]
- Zorzi, M.; Gai, F.; Medana, C.; Aigotti, R.; Morello, S.; Peiretti, P.G. Bioactive Compounds and Antioxidant Capacity of Small Berries. Foods 2020, 9, 623. [Google Scholar] [CrossRef]
- Ali, Y.M.; Kadir, A.A.; Ahmad, Z.; Yaakub, H.; Zakaria, Z.A.; Abdullah, M.N. Free radical scavenging activity of conjugated linoleic acid as single or mixed isomers. Pharm. Biol. 2012, 50, 712–719. [Google Scholar] [CrossRef]
- Triantis, T.M.; Yannakopoulou, E.; Nikokavoura, A.; Dimotikali, D.; Papadopoulos, K. Chemiluminescent studies on the antioxidant activity of amino acids. Anal. Chim. Acta 2007, 591, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Koch, W.; Kukuła-Koch, W.; Czop, M.; Helon, P.; Gumbarewicz, E. The Role of Extracting Solvents in the Recovery of Polyphenols from Green Tea and Its Antiradical Activity Supported by Principal Component Analysis. Molecules 2020, 25, 2173. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.A.; Jumbri, K.; Ramli, A.; Abd Ghani, N.; Ahmad, H.; Lim, J.W. A Kinetic Approach of DPPH Free Radical Assay of Ferulate-Based Protic Ionic Liquids (PILs). Molecules 2018, 23, 3201. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Oldoni, T.L.C.; Silva, R.C.; Carpes, S.T.; Massarioli, A.P.; Alencar, S.M. Antioxidant activity and development of one chromatographic method to determine the phenolic compounds from Agroindustrial Pomace. An. Acad. Bras. Cienc. 2020, 92, e20181068. [Google Scholar] [CrossRef] [PubMed]
- Elgndi, M.A.; Filip, S.; Pavlíc, B.; Stanojkovíc, T.; Zizak, Z.; Zecovic, Z. Antioxidative and cytotoxic activity of essential oils and extracts of Satureja montana L. Coriandrum sativum L. and Ocimum basilicum L. obtained by supercritical fluid extraction. J. Supercrit. Fluids 2017, 128, 128–137. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Lahouar, L.; El Arem, A.; Ghrairi, F.; Chahdoura, H.; Ben Salem, H.; El Felah, M.; Achour, L. Phytochemical content and antioxidant properties of diverse varieties of whole barley (Hordeum vulgare L.) grown in Tunisia. Food Chem. 2014, 145, 578–583. [Google Scholar] [CrossRef]
- Jorge, N.; Silva, A.C.; Aranha, C.P. Antioxidant activity of oils extracted from orange (Citrus sinensis) seeds. An. Acad. Bras. Cienc. 2016, 88, 951–958. [Google Scholar] [CrossRef]
- Olech, M.; Nowak, R.; Nowacka, N.; Pecio, Ł.; Oleszek, W.; Los, R.; Malm, A.; Rzymowska, J. Evaluation of rose roots, a post-harvest plantation residue as a source of phytochemicals with radical scavenging, cytotoxic, and antimicrobial activity. Ind. Crops Prod. 2015, 69, 129–136. [Google Scholar] [CrossRef]
- Nowak, D.; Gośliński, M.; Wojtowicz, E.; Przygoński, K. Antioxidant Properties and Phenolic Compounds of Vitamin C-Rich Juices. J. Food Sci. 2018, 83, 2237–2246. [Google Scholar] [CrossRef]
- Lee, S.G.; Wang, T.; Vance, T.M.; Hubert, P.; Kim, D.O.; Koo, S.I.; Chun, O.K. Validation of Analytical Methods for Plasma Total Antioxidant Capacity by Comparing with Urinary 8-Isoprostane Level. J. Microbiol. Biotechnol. 2017, 27, 388–394. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [PubMed]
- Miilošević, M.D.; Marinković, A.D.; Petrović, P.; Klaus, A.; Nikolić, M.G.; Prlainović, N.Ž.; Cvijetić, I.N. Synthesis, characterization and SAR studies of bis(imino)pyridines as antioxidants, acetylcholinesterase inhibitors and antimicrobial agents. Bioorganic Chem. 2020, 102, 104073. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, O.H.; Moreira, T.F.M.; De Oliveira, A.; Bracht, L.; Ineu, R.P.; Leimann, F.V. Antioxidant Activity of Encapsulated Extracts and Bioactives from Natural Sources. Curr. Pharm. Des. 2020, 26, 3847–3861. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.O.; Dommes, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Prior, R. Oxygen radical absorbance capacity (ORAC): New horizons in relating dietary antioxidants/bioactives and health benefits. J. Funct. Foods 2015, 18, 795–810. [Google Scholar] [CrossRef]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Ortiz, R.; Antilén, M.; Speisky, H.; Aliaga, M.E.; López-Alarcón, C.; Baugh, S. Application of a microplate-based ORAC-pyrogallol red assay for the estimation ofantioxidant capacity: First Action. J. AOAC Int. 2012, 95, 1558–1561. [Google Scholar] [CrossRef]
- Ortiz, R.; Antilén, M.; Speisky, H.; Aliaga, M.E.; López-Alarcón, C. Analytical parameters of the microplate-based ORAC-pyrogallol red assay. J. AOAC Int. 2011, 94, 1562–1566. [Google Scholar] [CrossRef]
- Ou, B.; Chang, T.; Huang, D.; Prior, R.L. Determination of total antioxidant capacity by oxygen radical absorbance capacity (ORAC) using fluorescein as the fluorescence probe: First Action 2012.23. J. AOAC Int. 2013, 96, 6–1372. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, G.; Prior, R.L. Total Antioxidant Capacity of Fruits. J. Agric. Food Chem. 1996, 44, 701–705. [Google Scholar] [CrossRef]
- Cao, G.; Prior, R.L. Measurement of oxygen radical absorbance capacity in biological samples. Methods Enzymol. 1999, 299, 50–62. [Google Scholar]
- Yongsheng, C.; Gu, C.; Xiong, F.; Rui-Hai, L. Phytochemical Profiles and Antioxidant Activity of Different Varieties of Adinandra Tea (Adinandra Jack). J. Agric. Food Chem. 2015, 63, 169–176. [Google Scholar]
- ORAC Database. Available online: http://oracdatabase.com/ (accessed on 3 June 2018).
- Speisky, H.; López-Alarcón, C.; Gómez, M.; Fuentes, J.; Sandoval-Acuña, C. First web-based database on total phenolics and oxygen radical absorbance capacity (ORAC) of fruits produced and consumed within the south Andes region of South America. J. Agric. Food Chem. 2012, 60, 8851–8859. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 15. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.A. Approach to optimization of FRAP methodology for studies based on selected monoterpenes. Molecules 2020, 25, 5267. [Google Scholar] [CrossRef] [PubMed]
- Berker, K.I.; Demirata, B.; Apak, R. Determination of total antioxidant capacity of lipophilic and hydrophilic antioxidants in the same solution by using ferric-ferricyanide assay. Food Anal. Methods 2012, 5, 9. [Google Scholar] [CrossRef]
- Berker, K.I.; Güçlük, K.; Tor, I.; Demirata, B.; Apak, R. Total antioxidant capacity assay using optimized ferricyanide/Prussian blue method. Food Anal. Methods 2010, 3, 14. [Google Scholar]
- Maciejczyk, M.; Szulimowska, J.; Taranta-Janusz, K.; Werbel, K.; Wasilewska, A.; Zalewska, A. Salivary FRAP as a marker of chronic kidney disease progression in children. Antioxidants 2019, 8, 409. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.K.; Jaiswal, A.; Sharma, P.; Kumar, P.; Singh, A.N. Oxidative stress and biomarker of TNF-α, MDA and FRAP in hypertension. J. Med. Life 2019, 12, 7. [Google Scholar] [CrossRef]
- Tomandlova, M.; Parenica, J.; Lokaj, P.; Ondrus, T.; Kala, P.; Miklikova, M.; Helanova, K.; Helan, M.; Malaska, J.; Benesova, K.; et al. Prognostic value of oxidative stress in patients with acute myocardial infarction complicated by cardiogenic shock: A prospective cohort study. Free Radic. Biol. Med. 2021, 174, 7. [Google Scholar] [CrossRef]
- Wayner, D.D.M.; Burton, G.W.; Ingold, K.U.; Locke, S. Quantitative measurement of the total peroxyl radical-trapping antioxidantcapability of human blood plasma by controlled peroxidation. FEBS Lett. 1985, 187, 5. [Google Scholar] [CrossRef]
- Lissi, E.; Salim-Hanna, M.; Pascual, C.; DelCastillo, M.D. Evaluation od total antioxidant potential (TRAP) and total antioxidant reactivity from luminol-enhanced chemiluminescence measurements. Free Radic. Biol. Med. 1995, 18, 6. [Google Scholar] [CrossRef]
- Dresch, M.T.K.; Rossato, S.B.; Kappel, V.D.; Biegelmeyer, R.; Hoff, M.L.M.; Mayorga, P.; Zuanazzi, J.A.S.; Henriques, A.T.; Moreira, J.C.F. Optimization and validation of an alternative method to evaluate total reactive antioxidant potential. Anal. Biochem. 2009, 385, 15. [Google Scholar] [CrossRef]
- Praud, D.; Parpinel, M.; Serafini, M.; Belloco, R.; Tavani, A.; Lagiou, P.; LaVecchia, C.; Rossi, M. Non-enzymatic antioxidant capacity and risk of gastric cancer. Cancer Epidemiol. 2015, 39, 6. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, C.J.; Shui, Y.B. Intraocular oxygen and antioxidant status: New insights on the effect of vitrectomy and glaucoma pathogenesis. Am. J. Ophthalmol. 2019, 203, 13. [Google Scholar] [CrossRef] [PubMed]
- Okimoto, Y.; Watanabe, A.; Niki, E.; Yamashita, T.; Noguchi, N. A novel fluorescent probe diphenyl-1-pyrenylphosphine to follow lipid peroxidation in cell membranes. FEBS Lett. 2000, 474, 137–140. [Google Scholar] [CrossRef]
- Takahashi, M.; Shibata, M.; Niki, E. Estimation of lipid peroxidation od live cells using a fluorescent probe, Diphenyl-1-pyrenylphosphine. Free Radic. Biol. Med. 2001, 31, 164–174. [Google Scholar] [CrossRef]
- Weinstein, D.S.; Liu, W.; Ngu, K.; Langevine, C.; Combs, D.W.; Zhuang, S.; Chen, C.; Madsen, C.S.; Harper, T.W.; Robl, J.A. Discovery od selective imidazole-based inhibitors of mammalian 15-lipoxygenase: Highly potent against human enzyme within a cellular environment. Bioorg. Med. Chem. Lett. 2007, 17, 5115–5120. [Google Scholar] [CrossRef]
- Dahlström, M.; Forsström, D.; Johannesson, M.; Huque-Andersson, Y.; Björk, M.; Silfverplatz, E.; Sanin, A.; Schaal, W.; Pelcman, B.; Forsell, P.K.A. Development of a fluorescent intensity assay amenable for high throughput screening for determining 15-lipoxygenase activity. J. Biomol. Screen. 2010, 15, 671–679. [Google Scholar] [CrossRef]
- Dobrian, A.D.; Lieb, D.C.; Cole, B.K.; Taylor-Fishwick, D.A.; Chakrabarti, S.K.; Nadler, J.L. Functional and pathological roles of the 12- and 15-lipoxygenases. Prog. Lipid Res. 2011, 50, 115–131. [Google Scholar] [CrossRef]
- Cropotova, J.; Rustad, T. A new fluorimetric method for simultaneous determination of lipid and protein hydroperoxides in muscle foods with the use of diphenyl-1-pyrenylphosphine (DPPP). LTW Food Sci. Technol. 2020, 128, 109467. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Bou, R.; Chen, B.; Guardiola, F.; Codony, R.; Decker, E.A. Determination of lipid and protein hydroperoxides using the fluorescent probe diphenyl-1-pyrenylphosphine. Food Chem. 2010, 123, 892–900. [Google Scholar] [CrossRef]
- Zhou, D.; Shao, L.; Spitz, D.R. Reactive oxygen species in normal and tumor stem cells. Adv. Cancer Res. 2014, 122, 1–67. [Google Scholar]
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Dontha, S. A review on antioxidant methods. Asian J. Pharm. Clin. Res. 2016, 9, 14–32. [Google Scholar]
- Rees, K.R.; Sinha, K.P. Blood enzymes in liver injury. J. Pathol. Bacteriol. 1960, 80, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Townsend, D.M.; Tew, K.D.; Tapiero, H. The importance of glutathione in human disease. Biomed. Pharm. 2003, 57, 145–155. [Google Scholar] [CrossRef]
- Flohé, L. The fairytale of the GSSG/GSH redox potential. Cell. Funct. Glutathione 2013, 1830, 3139–3142. [Google Scholar] [CrossRef] [PubMed]
- Alli, J.A.; Kehinde, A.O.; Kosoko, A.M.; Ademowo, O.G. Oxidative Stress and Reduced Vitamins C and E Levels Are Associated with Multi-Drug Resistant Tuberculosis. J. Tuberc. Res. 2014, 02, 52. [Google Scholar] [CrossRef]
- Apostolova, N.; Victor, V.M. Molecular strategies for targeting antioxidants to mitochondria: Therapeutic implications. Antioxid. Redox Signal. 2015, 22, 686–729. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A. Reduced glutathione: A radioprotector or a modulator of DNA-repair activity. Nutrients 2013, 5, 525–542. [Google Scholar] [CrossRef]
- Polefka, T.G.; Meyer, T.A.; Agin, P.P.; Bianchini, R.J. Cutaneous oxidative stress. J. Cosmet. Dermatol. 2012, 11, 55–64. [Google Scholar] [CrossRef]
- Meng, D.; Zhang, P.; Zhang, L.; Wang, H.; Ho, C.T.; Li, S.; Shahidi, F.; Zhao, H. Detection of cellular redox reactions and antioxidant activity assays. J. Funct. Foods 2017, 37, 467–479. [Google Scholar] [CrossRef]
- Brehe, J.E.; Burch, H.B. Enzymatic assay for glutathione. Anal. Biochem. 1976, 74, 189–197. [Google Scholar] [CrossRef]
- Liu, J.; Jia, L.; Kan, J.; Jin, C. In vitro and in vivo antioxidant activity of ethanolic extract of white button mushroom (Agaricus bisporus). Food Chem. Toxicol. 2013, 51, 310–316. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Cell. Funct. Glutathione 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Ye, Z.W.; Zhang, J.; Townsend, D.M.; Tew, K.D. Oxidative stress, redox regulation and diseases of cellular differentiation. Redox Regul. Differ. 2015, 1850, 1607–1621. [Google Scholar] [CrossRef]
- Flohe, L.; Günzler, W.A.; Schock, H.H. Glutathione peroxidase: A selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef]
- Thomson, C.D. Assessment of requirements for selenium and adequacy of selenium status: A review. Eur. J. Clin. Nutr. 2004, 58, 391. [Google Scholar] [CrossRef]
- Catarino, M.D.; Amarante, S.J.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Brown Algae Phlorotannins: A Marine Alternative to Break the Oxidative Stress, Inflammation and Cancer Network. Foods 2021, 10, 1478. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, S.; Guo, Y.; Jiao, Y.; Zhao, Y. Compositional characterization of soluble apple polysaccharides, and their antioxidant and hepatoprotective effects on acute CCl4-caused liver damage in mice. Food Chem. 2013, 138, 1256–1264. [Google Scholar] [CrossRef]
- Strauss, R.G.; Snyder, E.L.; Wallace, P.D.; Rosenberger, T.G. Oxygen-detoxifying enzymes in neutrophils of infants and their mothers. J. Lab. Clin. Med. 1980, 95, 897–904. [Google Scholar]
- Limón-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Oxidative Stress Mech. Environ. Toxic. 2009, 674, 137–147. [Google Scholar] [CrossRef]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Alterations in Concentration/Activity of Superoxide Dismutases in Context of Obesity and Selected Single Nucleotide Polymorphisms in Genes: SOD1, SOD2, SOD3. Int. J. Mol. Sci. 2020, 21, 5069. [Google Scholar] [CrossRef] [PubMed]
- Marklund, S.L.; Holme, E.; Hellner, L. Superoxide dismutase in extracellular fluids. Clin. Chim. Acta Int. J. Clin. Chem. 1982, 126, 41–51. [Google Scholar] [CrossRef]
- Yasui, K.; Kobayashi, N.; Yamazaki, T.; Agematsu, K.; Matsuzaki, S.; Ito, S.; Nakata, S.; Baba, A.; Koike, K. Superoxide dismutase (SOD) as a potential inhibitory mediator of inflammation via neutrophil apoptosis. Free Radic. Res. 2005, 39, 755–762. [Google Scholar] [CrossRef]
- Rondanelli, M.; Miraglia, N.; Putignano, P.; Castagliuolo, I.; Brun, P.; Dall’Acqua, S.; Peroni, G.; Faliva, M.A.; Naso, M.; Nichetti, M.; et al. Effects of 60-day Saccharomyces boulardii and Superoxide Dismutase Supplementation on Body Composition, Hunger Sensation, Pro/Antioxidant Ratio, Inflammation and Hormonal Lipo-Metabolic Biomarkers in Obese Adultos: A Double-Blind, Placebo-Controlled Trial. Nutrients 2021, 13, 2512. [Google Scholar]
- Vaneev, A.N.; Kost, O.A.; Eremeev, N.L.; Beznos, O.V.; Alova, A.V.; Gorelkin, P.V.; Erofeev, A.S.; Chesnokova, N.B.; Kabanov, A.V.; Klyachko, N.L. Superoxide Dismutase 1 Nanoparticles (Nano-SOD1) como um medicamento potencial para o tratamento de doenças inflamatórias dos olhos. Biomedicines 2021, 9, 396. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.C.; Corsi, D.; Cavi, N.; Bruni, N.; Dosio, F. Superoxide Dismutase Administration: A Review of Proposed Human Uses. Molecules 2021, 26, 1844. [Google Scholar] [CrossRef]
- Sgouros, D.; Katoulis, A.; Rigopoulos, D. Novel topical agent containing superoxide dismutase 100 000 IU and 4% of plant extracts as a mono-therapy for atopic dermatitis. J. Cosmet. Dermatol. 2017, 17, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Bonetta, R. Potential Therapeutic Applications of MnSODs and SOD-Mimetics. Chemistry 2018, 24, 5032–5041. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Zhu, Y.; Tong, Q.; Kosmacek, E.A.; Lichter, E.Z.; Oberley-Deegan, R.E. The Addition of Manganese Porphyrins during Radiation Inhibits Prostate Cancer Growth and Simultaneously Protects Normal Prostate Tissue from Radiation Damage. Antioxidants 2018, 7, 21. [Google Scholar] [CrossRef]
- Heer, C.D.; Davis, A.B.; Riffe, D.B.; Wagner, B.A.; Falls, K.C.; Allen, B.G.; Buettner, G.R.; Beardsley, R.A.; Riley, D.P.; Spitz, D.R. Superoxide Dismutase Mimetic GC4419 Enhances the Oxidation of Pharmacological Ascorbate and Its Anticancer Effects in an H₂O₂-Dependent Manner. Antioxidants 2018, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Coudriet, G.M.; Delmastro-Greenwood, M.M.; Previte, D.M.; Marré, M.L.; O’Connor, E.C.; Novak, E.A.; Vincent, G.; Mollen, K.P.; Lee, S.; Dong, H.H.; et al. Treatment with a Catalytic Superoxide Dismutase (SOD) Mimetic Improves Liver Steatosis, Insulin Sensitivity, and Inflammation in Obesity-Induced Type 2 Diabetes. Antioxidants 2017, 6, 85. [Google Scholar] [CrossRef]
- Cline, J.M.; Dugan, G.; Bourland, J.D.; Perry, D.L.; Stitzel, J.D.; Weaver, A.A.; Jiang, C.; Tovmasyan, A.; Owzar, K.; Spasojevic, I.; et al. Post-Irradiation Treatment with a Superoxide Dismutase Mimic, MnTnHex-2-PyP5+, Mitigates Radiation Injury in the Lungs of Non-Human Primates after Whole-Thorax Exposure to Ionizing Radiation. Antioxidants 2018, 7, 40. [Google Scholar] [CrossRef]
- Shrishrimal, S.; Kosmacek, E.; Chatterjee, A.; Tyson, M.; Oberley-Deegan, R. The SOD Mimic, MnTE-2-PyP, Protects from Chronic Fibrosis and Inflammation in Irradiated Normal Pelvic Tissues. Antioxidantes 2017, 6, 87. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Peng, K.; Fu, Z.-Y.; Tang, J.; Yang, M.J.; Chen, Q.C. Association of the C47T polymorphism in superoxide dismutase gene 2 with noise-induced hearing loss: A meta-analysis. Braz. J. Otorhinolaryngol. 2017, 83, 80–87. [Google Scholar] [CrossRef]
- Abati, E.; Bresolin, N.; Comi, G.; Corti, S. Silence superoxide dismutase 1 (SOD1): A promising therapeutic target for amyotrophic lateral sclerosis (ALS). Expert Opin. Ther. Targets 2020, 24, 295–310. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Flohé, L. Basic principles and emerging concepts in the redox control of transcription factors. Antioxid. Redox Signal. 2011, 15, 2335–2381. [Google Scholar] [CrossRef]
- Mehmet, A.; Hilal, A.; Dilek, O.; Ferruh, A.; Abdullah, G.; Murat, C.; Serhan, Y.; Volkan, H.; Gamze, K.; Işıl, T. Effects of various anesthesia maintenance on serum levels of selenium, copper, zinc, iron and antioxidant capacity. Braz. J. Anesthesiol. 2015, 65, 51–60. [Google Scholar]
- Collins, Y.; Chouchani, E.T.; James, A.M.; Menger, K.E.; Cochemé, H.M.; Murphy, M.P. Mitochondrial redox signalling at a glance. J. Cell Sci. 2012, 125, 801–806. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.P.; Cozzolino, S.F.; Marreiro, D.N.; Caldas, D.C.; Silva, K.G.; Sousa Almondes, K.G.; Neto, J.M.; Pimentel, J.C.; de Carvalho, C.G.; Nogueira, N.N. Mineral status and superoxide dismutase enzyme activity in Alzheimer’s disease. J. Trace Elem. Med. Biol. 2017, 44, 83–87. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Role of free radicals and catalytic metal ions in human disease: An overview. Methods Enzymol. 1990, 186, 1–85. [Google Scholar]
- Griess, B.; Tom, E.; Domann, F.; Teoh-Fitzgerald, M. Extracellular superoxide dismutase and its role in cancer. Free Radic. Biol. Med. 2017, 112, 464–479. [Google Scholar] [CrossRef]
- Quick, K.L.; Hardt, J.I.; Dugan, L.L. Rapid microplate assay for superoxide scavenging efficiency. J. Neurosci. Methods 2000, 97, 139–144. [Google Scholar] [CrossRef]
- Yamashita, N.; Murata, M.; Inoue, S.; Burkitt, M.J.; Milne, L.; Kawanishi, S. Alpha-tocopherol induces oxidative damage to DNA in the presence of copper (II) ions. Chem. Res. Toxicol. 1998, 11, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Chelikani, P.; Fita, I.; Loewen, P.C. Diversity of structures and properties among catalases. Cell. Mol. Life Sci. CMLS 2004, 61, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Gaetani, G.F.; Ferraris, A.M.; Rolfo, M.; Mangerini, R.; Arena, S.; Kirkman, H.N. Predominant role of catalase in the disposal of hydrogen peroxide within human erythrocytes. Blood 1996, 87, 1595–1599. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Ver. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2017, 54, 287–293. [Google Scholar] [CrossRef]
- Eisner, T.; Aneshansley, D.J. Spray aiming in the bombardier beetle: Photographic evidence. Proc. Natl. Acad. Sci. USA 1999, 96, 9705–9709. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Calderon, P.B. Catalase, a remarkable enzyme: Targeting the oldest antioxidant enzyme to find a new cancer treatment approach. Biol Chem. 2017, 398, 1095–1108. [Google Scholar] [CrossRef]
- Hadwan, M.H. Ensaio espectrofotométrico simples para medir a atividade da catalase em tecidos biológicos. BMC Biochem. 2018, 19, 1–7. [Google Scholar]
- Ceci, R.; Duranti, G.; Leonetti, A.; Pietropaoli, S.; Spinozzi, F.; Marcocci, L.; Amendola, R.; Cecconi, F.; Sabatini, S.; Mariottini, P.; et al. Adaptive responses of heart and skeletal muscle to spermine oxidase overexpression: Evaluation of a new transgenic mouse model. Free Radic. Biol. Med. 2017, 103, 216–225. [Google Scholar] [CrossRef]
- Nelson, S.K.; Bose, S.K.; Grunwald, G.K.; Myhill, P.; McCord, J.M. The induction of human superoxide dismutase and catalase in vivo: A fundamentally new approach to antioxidant therapy. Free Radic. Biol. Med. 2006, 40, 341–347. [Google Scholar] [CrossRef]
- Forman, H.J.; Augusto, O.; Brigelius-Flohe, R.; Dennery, P.A.; Kalyanaraman, B.; Ischiropoulos, H.; Mann, G.E.; Radi, R.; Roberts, L.J.; Vina, J.; et al. Even free radicals should follow some rules: A Guide to free radical research terminology and methodology. Free Radic. Biol. Med. 2015, 78, 233–235. [Google Scholar] [CrossRef]
- Kadiiska, M.B.; Gladen, B.C.; Baird, D.D.; Germolec, D.; Graham, L.B.; Parker, C.E.; Nyska, A.; Wachsman, J.T.; Ames, B.N.; Basu, S.; et al. Biomarkers of Oxidative Stress Study II: Are oxidation products of lipids, proteins, and DNA markers of CCl4 poisoning. Free Radic. Biol. Med. 2005, 38, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Draijer, R.; Van Dorsten, F.A.; Zebregs, Y.E.; Hollebrands, B.; Peters, S.; Duchateau, G.S.; Grün, C.H. Impact of Proteins on the Uptake, Distribution, and Excreetion of Phenolics in the Human Body. Nutrients 2016, 8, 814. [Google Scholar] [CrossRef]
- Parada, J.; Aguilera, J.M. Food microstructure affects the bioavailability of several nutrients. J. Food Sci. 2007, 72, 21–32. [Google Scholar] [CrossRef]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Stasiak, M.; Oniszczuk, A. Beneficial Effects of Phenolic Compounds on Gut Microbiota and Metabolic Syndrome. Int. J. Mol. Sci. 2021, 22, 3715. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Serrano, J.; Tabernero, M.; Arranz, S.; Díaz-Rubio, M.E.; García-Diz, L.; Goñi, I.; Saura-Calixto, F. Bioavailability of phenolic antioxidants associated with dietary fiber: Plasma antioxidant capacity after acute and long-term intake in humans. Plant Foods Hum. Nutr. 2009, 64, 102–107. [Google Scholar] [CrossRef]
- Williamson, G.; Clifford, M.N. Effect of thermal processing on free and bound phenolic compounds and antioxidant activities of hawthorn. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Costa, L.; Lean, M.; Crozier, A. Polyphenols and health: What compounds are involved. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 1–6. [Google Scholar] [CrossRef]
- Kawai, Y. Understanding metabolic conversions and molecular actions of flavonoids in vivo: Toward new strategies for effective utilization of natural polyphenols in human health. J. Med. Investig. 2018, 65, 162–165. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, 00370. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; González-Aguilar, G.A. The Role of Dietary Fiber in the Bioaccessibility and Bioavailability of Fruit and Vegetable Antioxidants. J. Food Sci. 2011, 76, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Lafay, S.; Gil-Izquierdo, A. Bioavailability of phenolic acids. Phytochem. Rev. 2008, 7, 301. [Google Scholar] [CrossRef]
- Lafay, S.; Gil-Izquierdo, A.; Manach, C.; Morand, C.; Besson, C.; Scalbert, A. Chlorogenic acid is absorbed in its intact form in the stomach of rats. J. Nutr. 2006, 136, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, L.; Martins, C.A.; Sampaio, G.R.; Monteiro, M.P.; César, L.M.; Mioto, B.M.; Mori, C.S.; Mendes, T.M.N.; Ribeiro, M.L.; Arçari, D.P.; et al. Bioavailability of catechins from guaraná (Paullinia cupana) and its effect on antioxidant enzymes and other oxidative stress markers in healthy human subjects. Food Funct. 2016, 7, 2970–2978. [Google Scholar] [CrossRef]
- Yao, H.T.; Hsu, Y.R.; Lii, C.K.; Lin, A.H.; Chang, K.H.; Yang, H.T. Effect of commercially available green and black tea beverages on drug-metabolizing enzymes and oxidative stress in Wistar rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 70, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Lara-Guzmán, O.J.; Álvarez-Quintero, R.; Osorio, E.; Naranjo-Cano, M.; Muñoz-Durango, K. CGMS method to quantify bioavailable phenolic compounds and antioxidant capacity determination of plasma after acute coffee consumption in human volunteers. Food Res. Int. Ott. Ont. 2016, 89, 219–226. [Google Scholar] [CrossRef]
- Yeh, H.; Chuang, C.; Chen, H.; Wan, C.; Chen, T.; Lin, L. Bioactive components analysis of two various gingers (Zingiber officinale Roscoe) and antioxidant effect of ginger extracts. LWT Food Sci. Technol. 2014, 55, 329–334. [Google Scholar] [CrossRef]
- Sauberlich, H. Bioavailability of vitamins. Prog. Food Nutr. Sci. 1985, 9, 1–33. [Google Scholar]
- Naidu, K.A. Vitamin C in human health and disease is still a mystery—An overview. Nutr. J. 2003, 2, 1–7. [Google Scholar] [CrossRef]
- Borel, P.; Preveraud, D.; Desmarchelier, C. Bioavailability of vitamin E in humans: An update. Nutr. Rev. 2013, 71, 319–331. [Google Scholar] [CrossRef]
- Iwata, N.; Okazaki, M.; Xuan, M.; Kamiuchi, S.; Matsuzaki, H.; Hibino, Y. Orally Administrated Ascorbic Acid Suppresses Neuronal Damage and Modifies Expression of SVCT2 and GLUT1 in the Brain of Diabetic Rats with Cerebral Ischemia-Reperfusion. Nutrients 2014, 6, 1554–1577. [Google Scholar] [CrossRef]
- Bayrak, B.B.; Çatal, T.; Öztay, F.; Yanardağ, R.; Bolkent, Ş. Efficacy of antioxidant vitamins (vitamin C, vitamin E, beta-carotene) and selenium supplement on D-galactosamine-induced lung injury. IUFS J. Biol. 2016, 75, 11–18. [Google Scholar]
- Catal, T.; Tunali, S.; Bolkent, S.; Yanardag, R. An Antioxidant Combination Improves Histopathological Alterations and Biochemical Parameters in D-Galactosamine-Induced Hepatotoxicity in Rats. Eur. J. Biol. 2017, 76, 14–19. [Google Scholar] [CrossRef]
- Donhowe, E.G.; Kong, F. Beta-carotene: Digestion, microencapsulation, and in vitro bioavailability. Food Bioprocess Technol. 2014, 7, 338–354. [Google Scholar] [CrossRef]
- Desmarchelier, C.; Borel, P. Overview of carotenoid bioavailability determinants: From dietary factors to host genetic variations. Trends Food Sci. Technol. 2017, 69, 270–280. [Google Scholar] [CrossRef]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Rühl, R.; Keijer, J.; Borel, P. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef]
- Ho, N.; Inbaraj, B.S.; Chen, B.H. Utilization of microemulsions from Rhinacanthus nasutus (L.) Kurz to improve carotenoid bioavailability. Sci. Rep. 2016, 6, 25426. [Google Scholar] [CrossRef]
- Aschoff, J.K.; Rolke, C.L.; Breusing, N.; Bosy-Westphal, A.; Högel, J.; Carle, R.; Schweiggert, R.M. Bioavailability of β-cryptoxanthin is greater from pasteurized orange juice than from fresh oranges–a randomized cross-over study. Mol. Nutr. Food Res. 2015, 59, 1896–1904. [Google Scholar] [CrossRef]
- Van het Hof, K.H.; West, C.E.; Weststrate, J.A.; Hautvast, J.A.J. Dietary Factors That Affect the Bioavailability of Carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar] [CrossRef]
- Gupta, R.; Kopec, R.E.; Schwartz, S.J.; Balasubramaniam, V. Combined pressure–temperature effects on carotenoid retention and bioaccessibility in tomato juice. J. Agric. Food Chem. 2011, 59, 7808–7817. [Google Scholar] [CrossRef]
- Buniowska, M.; Carbonell-Capella, J.M.; Frigola, A.; Esteve, M.J. Bioaccessibility of bioactive compounds after non-thermal processing of an exotic fruit juice blend sweetened with Stevia rebaudiana. Food Chem. 2017, 221, 1834–1842. [Google Scholar] [CrossRef]
- Ranga Rao, A.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. In vivo bioavailability and antioxidant activity of carotenoids from microalgal biomass—A repeated dose study. Food Res. Int. 2013, 54, 711–717. [Google Scholar] [CrossRef]
- Ravi, H.; Baskaran, V. Chitosan-glycolipid nanocarriers improve the bioavailability of fucoxanthin via up-regulation of PPARγ and SRB1 and antioxidant activity in rat model. J. Funct. Foods 2017, 28, 215–226. [Google Scholar] [CrossRef]
- Campos, K.K.D.; Araújo, G.R.; Martins, T.L.; Bandeira, A.C.B.; Costa, G.P.; Talvani, A.; Garcia, C.C.M.; Oliveira, L.A.M.; Costa, D.C.; Bezerra, F.S. The antioxidant and anti-inflammatory properties of lycopene in mice lungs exposed to cigarette smoke. J. Nutr. Biochem. 2017, 48, 9–20. [Google Scholar] [CrossRef]
- Gökmen, V.; Serpen, A.; Fogliano, V. Direct measurement of the total antioxidant capacity of foods: The ‘QUENCHER’ approach. Trends Food Sci. Technol. 2009, 20, 278–288. [Google Scholar] [CrossRef]
- Del Pino-García, R.; García-Lomillo, J.; Rivero-Pérez, M.D.; González-SanJosé, M.L.; Muñiz, P. Adaptation and Validation of QUick, Easy, New, CHEap, and Reproducible (QUENCHER) Antioxidant Capacity Assays in Model Products Obtained from Residual Wine Pomace. J. Agric. Food Chem. 2015, 63, 6922–6931. [Google Scholar] [CrossRef]
- Kitrytė, V.; Šaduikis, A.; Venskutonis, P.R. Assessment of antioxidant capacity of brewer’s spent grain and its supercritical carbon dioxide extract as sources of valuable dietary ingredients. J. Food Eng. 2015, 167, 18–24. [Google Scholar] [CrossRef]
- Laus, M.N.; Di Benedetto, N.A.; Caporizzi, R.; Tozzi, D.; Soccio, M.; Giuzio, L.; De Vita, P.; Flagella, Z.; Pastore, D. Evaluation of Phenolic Antioxidant Capacity in Grains of Modern and Old Durum Wheat Genotypes by the Novel QUENCHERABTS Approach. Plant Foods Hum. Nutr. Dordr. Neth. 2015, 70, 207–214. [Google Scholar] [CrossRef]
- Shahbaz, H.M.; Park, E.J.; Kim, G.R.; Akram, K.; Kwon, J.H. Assessment of antioxidant potential of pomegranate fruit by-products via a direct approach using a simple quencher method. J. AOAC Int. 2016, 99, 599–603. [Google Scholar] [CrossRef]
- Keller, J.N.; Kindy, M.S.; Holtsberg, F.W.; Clair, D.K.S.; Yen, H.C.; Germeyer, A.; Steiner, S.M.; Bruce-Keller, A.J.; Hutchins, J.B.; Mattson, M.P. Mitochondrial manganese superoxide dismutase prevents neural apoptosis and reduces ischemic brain injury: Suppression of peroxynitrite production, lipid peroxidation, and mitochondrial dysfunction. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 687–697. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Qin, H.; Sankarapandi, S.; Becher, M.W.; Wong, P.C.; Zweier, J.L. Overexpression of human copper, zinc-superoxide dismutase (SOD1) prevents postischemic injury. Proc. Natl. Acad. Sci. USA 1998, 95, 4556–4560. [Google Scholar] [CrossRef]
- Fujita, H.; Fujishima, H.; Chida, S.; Takahashi, K.; Qi, Z.; Kanetsuna, Y.; Breyer, M.D.; Harris, R.C.; Yamada, Y.; Takahashi, T. Reduction of renal superoxide dismutase in progressive diabetic nephropathy. J. Am. Soc. Nephrol. JASN 2009, 20, 1303–1313. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Sah, S.K.; Zouboulis, C.C.; Kim, T.Y. Inhibitory effects of superoxide dismutase 3 on Propionibacterium acnes-induced skin inflammation. Sci Rep. 2018, 8, 4024. [Google Scholar] [CrossRef]
- Agrahari, G.; Sah, S.K.; Nguyen, C.T.; Choi, S.S.; Kim, H.Y.; Kim, T.Y. Superoxide Dismutase 3 Inhibits LL-37/KLK-5-Mediated Skin Inflammation through Modulation of EGFR and Associated Inflammatory Cascades. J. Investig. Dermatol. 2020, 140, 656–665. [Google Scholar] [CrossRef]
- Groner, Y.; Elroy-Stein, O.; Bernstein, Y.; Dafni, N.; Levanon, D.; Danciger, E.; Neer, A. Molecular genetics of Down’s syndrome: Overexpression of transfected human Cu/Zn-superoxide dismutase gene and the consequent physiological changes. Cold Spring Harb. Symp. Quant. Biol. 1986, 51, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Amstad, P.; Peskin, A.; Shah, G.; Mirault, M.E.; Moret, R.; Zbinden, I.; Cerutti, P. The balance between copper-zinc superoxide dismutase and catalase affects the sensitivity of mouse epidermal cells to oxidative stress. Biochemistry 1991, 30, 9305–9313. [Google Scholar] [CrossRef] [PubMed]
- Amstad, P.; Moret, R.; Cerutti, P. Glutathione peroxidase compensates for the hypersensitivity of Cu, Zn-superoxide dismutase overproducers to oxidant stress. J. Biol. Chem. 1994, 269, 1606–1609. [Google Scholar] [CrossRef]
- Kowald, A.; Lehrach, H.; Klipp, E. Alternative pathways as mechanism for the negative effects associated with overexpression of superoxide dismutase. J. Theor. Biol. 2006, 238, 828–840. [Google Scholar] [CrossRef]
- Sinet, P.M. Metabolism of oxygen derivatives in Down’s syndrome. Ann. N. Y. Acad. Sci. 1982, 396, 83–94. [Google Scholar] [CrossRef]
- Abaspour, A.R.; Taghikhani, M.; Parizade, M.R.; Moohebati, M.; Ghafoori, F.; Mehramiz, M.; Tayefi, M.; Avan, A.; Ghalandari, M.; Ferns, G.A.A.; et al. Association between Serum Glutathione Peroxidases and Superoxide Dismutases mRNA Level with Coronary Artery Disease. Health 2017, 9, 252–260. [Google Scholar] [CrossRef]
- Salem, K.; McCormick, M.L.; Wendlandt, E.; Zhan, F.; Goel, A. Copper–zinc superoxide dismutase-mediated redox regulation of bortezomib resistance in multiple myeloma. Redox Biol. 2014, 4, 23–33. [Google Scholar] [CrossRef]
- Glorieux, C.; Zamocky, M.; Sandoval, J.M.; Verrax, J.; Calderon, P.B. Regulation of catalase expression in healthy and cancerous cells. Free Radic. Biol. Med. 2015, 87, 84–97. [Google Scholar] [CrossRef]
- Röhrdanz, E.; Schmuck, G.; Ohler, S.; Kahl, R. The influence of oxidative stress on catalase and MnSOD gene transcription in astrocytes. Brain Res. 2001, 900, 28–136. [Google Scholar] [CrossRef]
- Röhrdanz, E.; Kahl, R. Alterations of antioxidant enzyme expression in response to hydrogen peroxide. Free Radic. Biol. Med. 1998, 24, 27–38. [Google Scholar] [CrossRef]
- Sen, P.; Mukherjee, S.; Bhaumik, G.; Das, P.; Ganguly, S.; Choudhury, N.; Raha, S. Enhancement of catalase activity by repetitive low-grade H2O2 exposures protects fibroblasts from subsequent stress-induced apoptosis. Mutat. Res. 2003, 529, 87–94. [Google Scholar] [CrossRef]
- Giustarini, D.; Tsikas, D.; Colombo, G.; Milzani, A.; Dalle-Donne, I.; Fanti, P.; Rossi, R. Pitfalls in the analysis of the physiological antioxidant glutathione (GSH) and its disulfide (GSSG) in biological samples: An elephant in the room. Anal. Tools Protoc. Oxidative Stress 2016, 1019, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Hedrén, E.; Diaz, V.; Svanberg, U. Estimation of carotenoid accessibility from carrots determined by an in vitro digestion method. Eur. J. Clin. Nutr. 2002, 56, 425. [Google Scholar] [CrossRef] [PubMed]
- Mason, S.A.; Della Gatta, P.A.; Snow, R.J.; Russell, A.P.; Wadley, G.D. Ascorbic acid supplementation improves skeletal muscle oxidative stress and insulin sensitivity in people with type 2 diabetes: Findings of a randomized controlled study. Free Radic. Biol. Med. 2016, 93, 227–238. [Google Scholar] [CrossRef]
- Kagan, V.E.; Tyurina, Y.Y. Recycling and Redox Cycling of Phenolic Antioxidants. Ann. N. Y. Acad. Sci. 2006, 854, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Bayir, H.; Kagan, V.E.; Tyurina, Y.Y.; Tyurin, V.; Ruppel, R.A.; Adelson, P.D.; Graham, S.H.; Janesko, K.; Clark, R.S.B.; Kochanek, P.M. Assessment of Antioxidant Reserves and Oxidative Stress in Cerebrospinal Fluid after Severe Traumatic Brain Injury in Infants and Children. Pediatr. Res. 2002, 51, 571. [Google Scholar] [CrossRef]
- Bast, A.; Haenen, G.R.M. Ten misconceptions about antioxidants. Trends Pharmacol. Sci. 2013, 34, 430–436. [Google Scholar] [CrossRef]
- Heaney, R.P. Nutrients, endpoints, and the problem of proof. J. Nutr. 2008, 138, 1591–1595. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Method | Advantages | Disadvantages |
|---|---|---|
| DPPH | Easy procedure Low cost Fast reaction | EC50 concept could be hard to interpret No standardization among time to assess or endpoint reaction |
| ABTS | Low cost Fast reaction Can be used to assess pH effect on activity, since is stable pH | Now standardized, which difficult comparison among results It is necessary an extra step to generate free radical from ABTS salt The generated free radical is not stable for long periods of time |
| ORAC | Once the method is standardized allows comparison Takes in account both time and antioxidant reaction Use biologically relevant antioxidants | Expensive pH-sensitive Quantification of results takes long time Results can vary across different equipment |
| FRAP | Used in samples of plant origin. | Uses high volume of reagent. |
| TRAP | It is sensitive to all antioxidants present in the sample. | It is a complex and time-consuming technique that requires a high degree of knowledge and experience. |
| Compound | Study Design | Bioavailability | Assays | Results | Related Antioxidant Activity | Reference |
|---|---|---|---|---|---|---|
| Guaraná powdered seed (Paullinia cupana) | Humans (healthy overweight adults) n = 12 | Detected 1 h after intake and remained after plasma clearance (plasma HPLC) | ORAC | ↑ORAC | Guaraná is a rich source of bioavailable catechins and contributes to reducing the oxidative stress parameters of clinically health overweight individuals by direct antioxidant action and up-regulation of antioxidant enzymes. | Yonekura et al. (2016) [178] |
| Single dose (3 g/90 mg catechins and 60 mg epicatechins equivalent) daily for 15 days | SOD/CAT/GPx activities | ↑CAT/GPx ↔SOD | ||||
| Blood samples: overnight-fasting and 1 h after intake | Ex vivo LDL oxidation/H2O2 induced DNA damage in lymphocytes (Comet Assay) | ↓LDL oxidation (only in the first day of study) ↓DNA damage (only after 1 h intake) | ||||
| Green (GT) and Black Tea (BT) (commercially acquired) | Male Wistar rats (n = 18) | Not evaluated | Drug metabolizing enzymes activity (hepatic and pulmonary) | ↑P450 (CYP) 1A1 (hep) ↑UDP-glucoronosyltransferase (hep/pulm) ↑CYP 1A2 (hep) ↓CYP 2C (hep) ↓CYP 2E1 (hep) ↓CYP 3A (hep) | Feeding both tea drinks to rats modulated drug metabolizing enzymes at a transcriptional level and reduced oxidative stress in the liver and lungs, but green tea was more effective in reducing oxidative stress. Their possible interactions with drugs or toxic compounds should be taken into account. | Yao et al. (2014) [179] |
| Ad libitum with food and as water replacement for 5 weeks | GSH, GSSG and GSH/GSSG ratio | ↓[GSH] (hep/pulm) ↓Lipid peroxide (pulm/GT) | ||||
| Blood and tissue (liver and lungs) samples after being euthanized | GPx and GSR TBARS and ROS | |||||
| DNA-binding activity of nuclear factors | ||||||
| Coffee (caffeic and ferulic acids) | Humans (n = 20) | Detected 1 h after intake (plasma HPLC) | ORAC | ↑ORAC | Lara-Guzmán et al. (2016) [180] | |
| FRAP/TRAP | ↑FRAP/↔TRAP | The experiments on plasma with caffeic and ferulic acids showed a significant increase in the antioxidant activity as well as delay of LDL oxidation. | ||||
| After acute consumption (1 h/400 mL) | Ex vivo LDL oxidation | ↓LDL oxidation | ||||
| Vitamin C supplementation | Male Sprague Dawley rats (n = 6–7 per group) | Detected in plasma and cerebral tissue (spectrophotometry) | Histopathology | Diabetic state: ↑infarct volume and edema Vit. C: ↓damage | Daily intake of ascorbic acid attenuates the exacerbation of cerebral ischemic injury in a diabetic state, which may be attributed to anti-apoptotic and anti-inflammatory effects via the improvement of augmented oxidative stress in the brain. Ascorbic acid supplementation may protect endothelial function against the exacerbated ischemic oxidative injury and improve its transport through SVCT2 in the cortex. | Iwata et al. (2014) [184] |
| Streptozotocin-induced type 1 diabetes | IHC (SVCT2, GLUT-1, cleaved caspase-3, TNF-α, IL-1β | ↓cleaved caspase-3 ↓inflammatory cytokines (TNF-α and IL-1β) | ||||
| 100 mg/kg of ascorbic acid (gavage) for 2 weeks before cerebral ischemia-reperfusion protocol | PCR (SVCT2 and GLUT1) | ↑SVCT2 (neurons and endothelial cells) ↑GLUT1 (endothelial cells) | ||||
| Cerebral ischemia-reperfusion (infarct induction) | Superoxide production | ↓superoxide radical | ||||
| Multivitamin supplementation (vit. C + vit. E + sodium selenite + β-carotene) | Female Sprague Dawley rats (n = 40) | Not evaluated | Histopathology | ↓honeycomb-like injury ↓edema ↔inflammatory infiltrate | The antioxidant combination protected lung tissue against damage by enhancing biochemical parameters and pulmonary edema, while no significant effect on protection of pulmonary inflammation was observed. The antioxidant vitamin supplementation with selenium can be used in the prevention of acute lung injury. | Bayrak et al. (2016) [185] |
| Vitamin C (100 mg/kg/day), vitamin E (100 mg/kg/day), sodium selenite (0.2 mg/kg/day and β-carotene (15 mg/kg/day) via gavage for 3 days before the injury protocol | ||||||
| D-galactosamine-induced (DGaIN) acute lung injury | GSH/GPx/PON TF/LDH/CAT/SOD/MPO/XO / | ↑GSH/GPx/PON ↓TF/LDH/CAT/SOD/MPO/XO/Na+/K+ ATPase | ||||
| LPO (MDA) | ↓LPO | |||||
| Multivitamin supplementation (vit. C + vit. E + sodium selenite + β-carotene) | Female Sprague Dawley rats (n = 40) | Not evaluated | Histopathology | ↓edema ↓necrosis ↓inflammatory infiltrate | The combination of antioxidants suppressed histopathological changes in the liver and biochemical parameters in D-GaIN-induced hepatotoxicity rats. The antioxidant vitamin supplementation with selenium can be used in the prevention of acute hepatotoxicity. | Catal et al. (2017) [186] |
| Vitamin C (100 mg/kg/day), vitamin E (100 mg/kg/day), sodium selenite (0.2 mg/kg/day and β-carotene (15 mg/kg/day) via gavage for 3 days before the injury protocol | Blood GSH/CAT Blood AST/ALT/ALP/GGT/LDH Blood sialic and uric acid Hepatic CAT/SOD/GSH/GPx Hepatic GST | ↑blood GSH and CAT ↓blood AST/ALT/ALP/GGT/LDH ↓blood sialic and uric acid ↑hepatic CAT/SOD/GSH/GPx ↔hepatic GST | ||||
| D-galactosamine-induced (DGaIN) acute liver injury | LPO (MDA) | ↓hepatic LPO | ||||
| Histopathology | ||||||
| Carotenoids derived from microalgal biomass (Spirulina platensis, Haematococcus pluvialis and Botryococcus braunii) | Male Wistar rats (n = 25) | Detected in plasma after 2 h, in the liver after 4 h and in the eyes after 6 h (HPLC/LC-MS) Carotenoid’s accumulation: liver > eyes > plasmaAstaxanthin from H. pluvialis had better bioavailability and accumulation than other groups. | Plasma and hepatic SOD, CAT, peroxidase and lipid peroxidation (TBARS) | ↑plasma and hepatic SOD, CAT and peroxidase ↓Lipid peroxidation | Rao et al. (2013) [194] | |
| Sigle dose of microalgal biomass (200 µM equivalent of β-carotene, astaxanthin and lutein) via gavage for 15 days | These results indicate that the astaxanthin from H. pluvialis has better bioavailability and better antioxidant properties compared to other carotenoids. Microalgae biomass is capable of preventing lipid peroxidation through scavenging free radicals and hydroxyl radicals in living cells and also restoring antioxidant enzyme activity. | |||||
| Fucoxanthin (FUCO) solubilized in glycolipid (GL) and absorbed from chitosan nanogels (NG) | Rat model Groups: control, FUCO, FUCO+NG-GL, FUCO+NG+GL | Plasma FUCO (HPLC). After single dose: ↑ 227.5% After repeated doses: ↑ 292.4% (compared to control and -GL groups). | CAT/GST/SODLipid peroxidation (TBARS) | ↑CAT/GST/SOD (+GL > ☐GL > controls) ↓Lipid peroxidation (controls > ☐GL > +GL) | The advantage of CS-NGs + GL for improved FUCO bioavailability via passive and active transport through PPARc mediated SRB1 activation was demonstrated. Elevated plasma and tissue levels of FUCO in these groups could be the reason for a higher activity of antioxidant enzymes and lower lipid peroxides. | Ravi and Baskaram (2017) [195] |
| Single dose study: 48 h Repeated dose study: 14 days Dietary feeding study: 1 month | Plasma FUCO (HPLC) after dietary feeding: ↑ 57.5% (compared to ☐GL) ↑ 400% (compared to seaweed) ↑ 287% (compared to control) | PPARγ/SRB1 | ↑PPARγ/SRB1(FUCO and GL had an agonist action) | |||
| Lycopene | Murine emphysema model (n = 40) | Not evaluated | Histopathology | Inhibited emphysema-like features when treated with LY | Lycopene acts as an antioxidant and anti-inflammatory through the neutralization of reactive species production in vitro and in vivo, the restoration of the GSH/GSSG ratio, decreasing oxidative damage, decreasing pro-inflammatory cytokines through decreased cell influx and the direct suppression of cytokine production. | Campos et al. (2017) [196] |
| Cigarette smoke exposure 3 times a day for 60 days | SOD/CAT/GPxGSH/GSSG ratio | ↔SOD/↓CAT/↑GPx ↑GSH/GSSG ratio | ||||
| Treatment with lycopene (LY) diluted in sunflower oil (25 and 50 mg/kg/day) | TBARS | ↓TBARS | ||||
| IFN-γ/TNF-α/IL-10 (bronchoalveolar fluid) | ↓ IFN-γ/TNF-α/IL-10 | |||||
| Leukocyte influx (bronchoalveolar fluid) | ↓ Leukocyte influx |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendonça, J.d.S.; Guimarães, R.d.C.A.; Zorgetto-Pinheiro, V.A.; Fernandes, C.D.P.; Marcelino, G.; Bogo, D.; Freitas, K.d.C.; Hiane, P.A.; de Pádua Melo, E.S.; Vilela, M.L.B.; et al. Natural Antioxidant Evaluation: A Review of Detection Methods. Molecules 2022, 27, 3563. https://doi.org/10.3390/molecules27113563
Mendonça JdS, Guimarães RdCA, Zorgetto-Pinheiro VA, Fernandes CDP, Marcelino G, Bogo D, Freitas KdC, Hiane PA, de Pádua Melo ES, Vilela MLB, et al. Natural Antioxidant Evaluation: A Review of Detection Methods. Molecules. 2022; 27(11):3563. https://doi.org/10.3390/molecules27113563
Chicago/Turabian StyleMendonça, Jenifer da Silva, Rita de Cássia Avellaneda Guimarães, Verônica Assalin Zorgetto-Pinheiro, Carolina Di Pietro Fernandes, Gabriela Marcelino, Danielle Bogo, Karine de Cássia Freitas, Priscila Aiko Hiane, Elaine Silva de Pádua Melo, Marcelo Luiz Brandão Vilela, and et al. 2022. "Natural Antioxidant Evaluation: A Review of Detection Methods" Molecules 27, no. 11: 3563. https://doi.org/10.3390/molecules27113563
APA StyleMendonça, J. d. S., Guimarães, R. d. C. A., Zorgetto-Pinheiro, V. A., Fernandes, C. D. P., Marcelino, G., Bogo, D., Freitas, K. d. C., Hiane, P. A., de Pádua Melo, E. S., Vilela, M. L. B., & Nascimento, V. A. d. (2022). Natural Antioxidant Evaluation: A Review of Detection Methods. Molecules, 27(11), 3563. https://doi.org/10.3390/molecules27113563