
Transcriptome Profiling of HCT-116 Colorectal Cancer Cells with RNA Sequencing Reveals Novel Targets for Polyphenol Nano Curcumin


Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Studies
2.1.1. Cell Culture and Reagents
2.1.2. Synthesis of Gemini-Cur nanoparticles
2.1.3. Gemini-Cur Treatments
2.1.4. RNA Extraction and Preparation
2.1.5. Library Construction and RNA Sequencing
2.2. Bioinformatics Studies
2.2.1. Quality Assessment of RNA-seq Data, Mapping and Read Annotation
2.2.2. Normalization of Read Counts, Differentially Expression Analysis (DEA) and Network Construction
2.2.3. Functional Enrichment and Gene Ontology Analysis
2.3. Exploration and Validation of CRC-Related Genes Based on Real-Time PCR and Western Blotting
3. Results
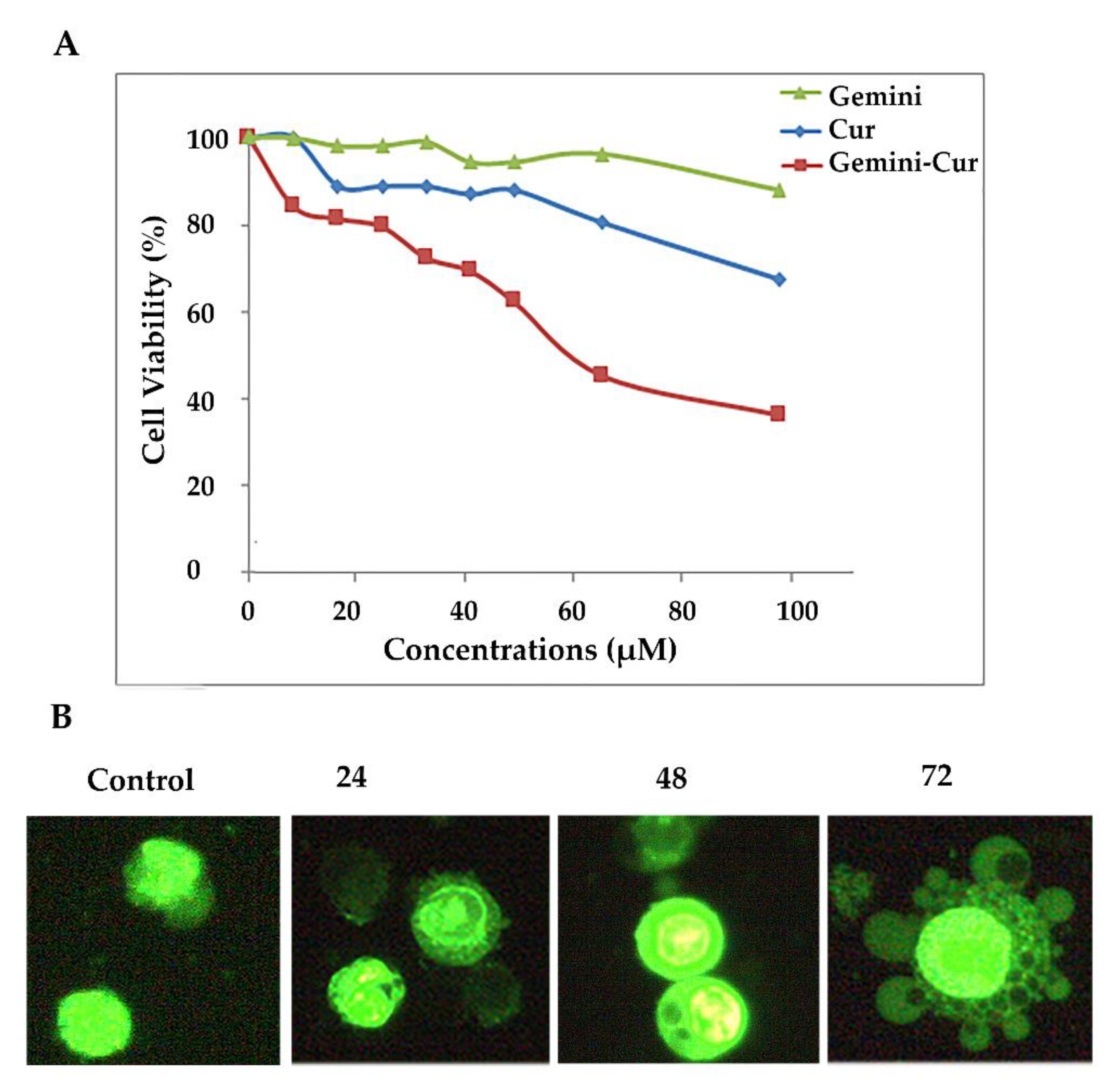
3.1. Suppressive Effect of Gemini-Cur on HCT-116 Cells
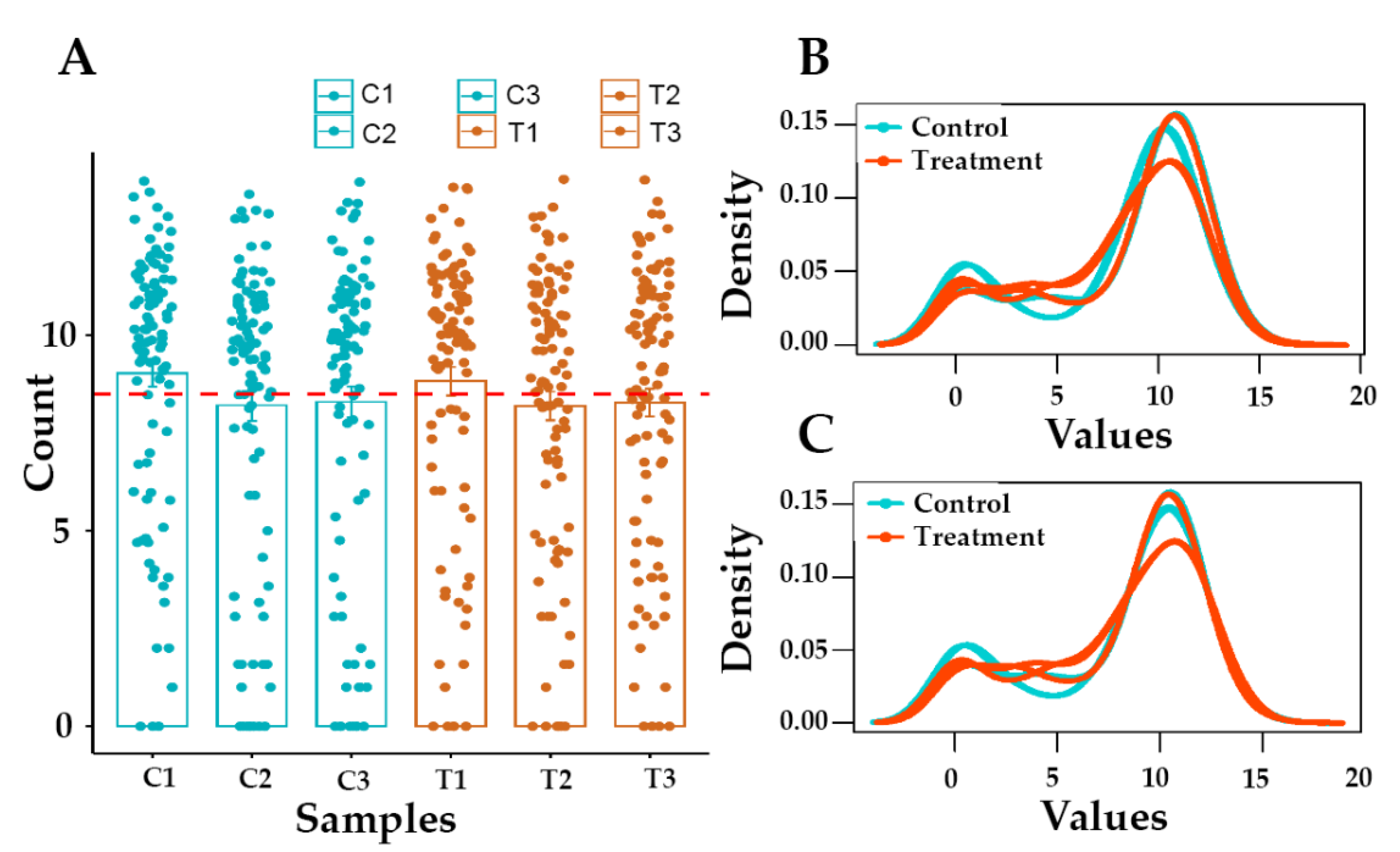
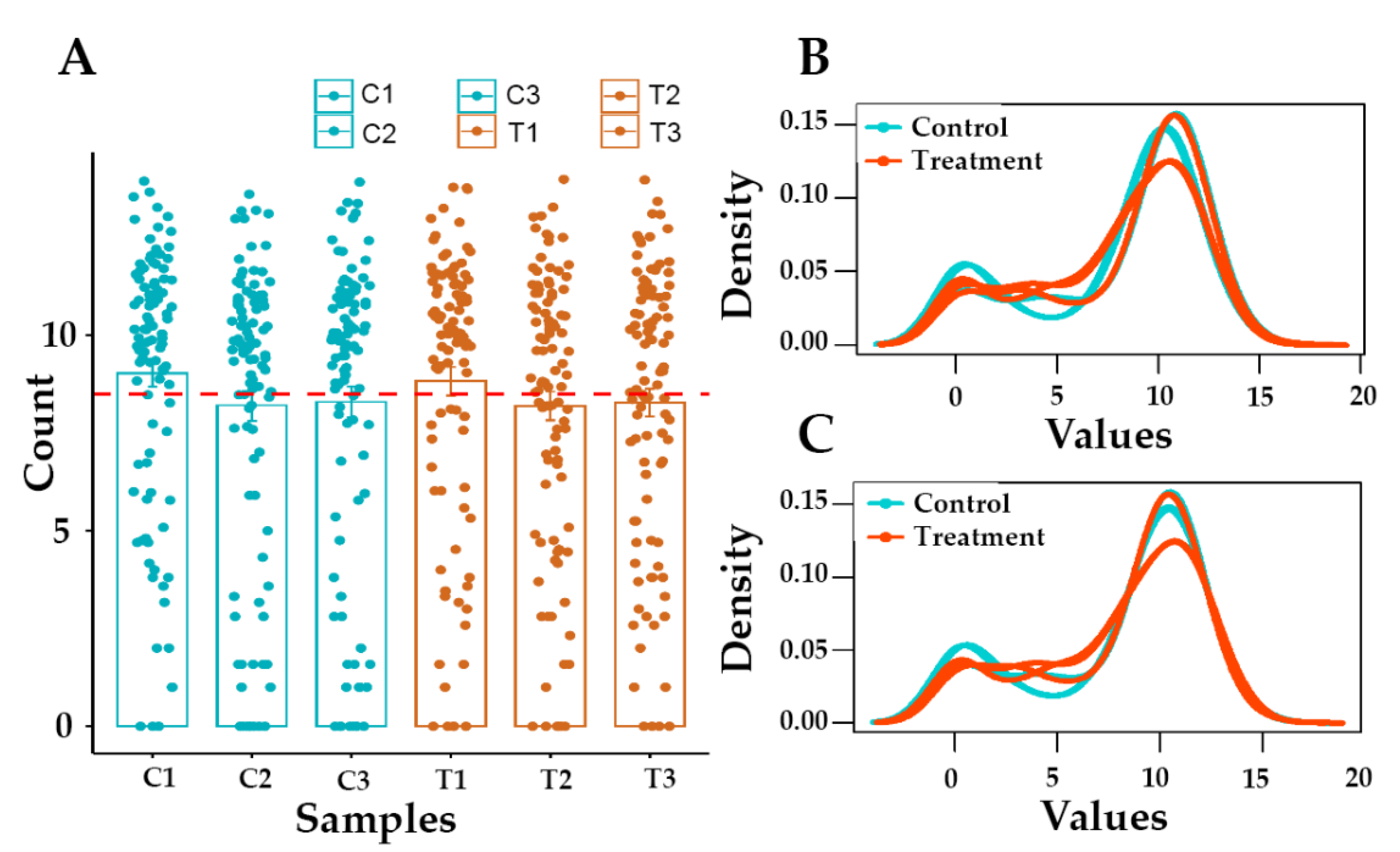
3.2. Raw Data Statistics and Quality Assessment
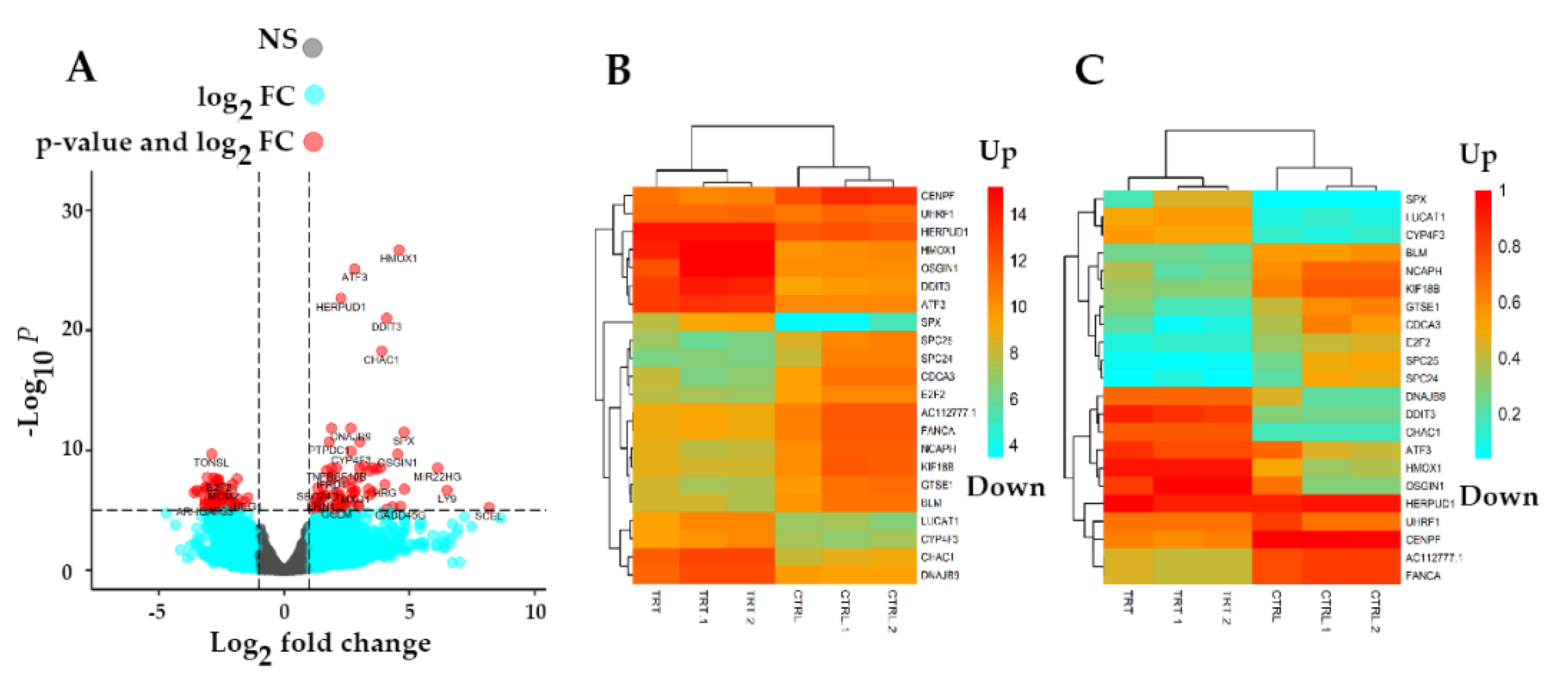
3.3. Identification of Differentially Expressed Genes (DEGs)
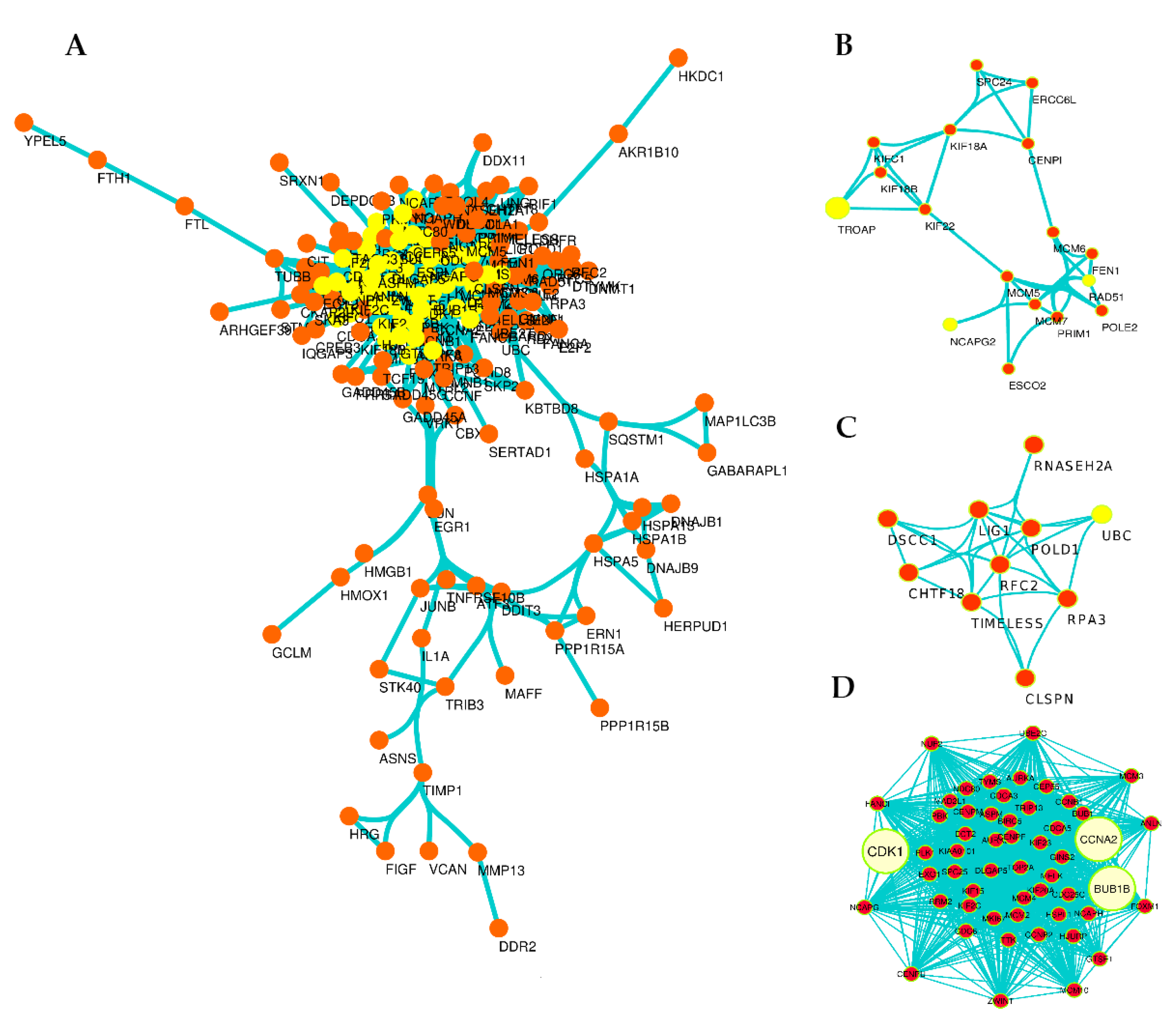
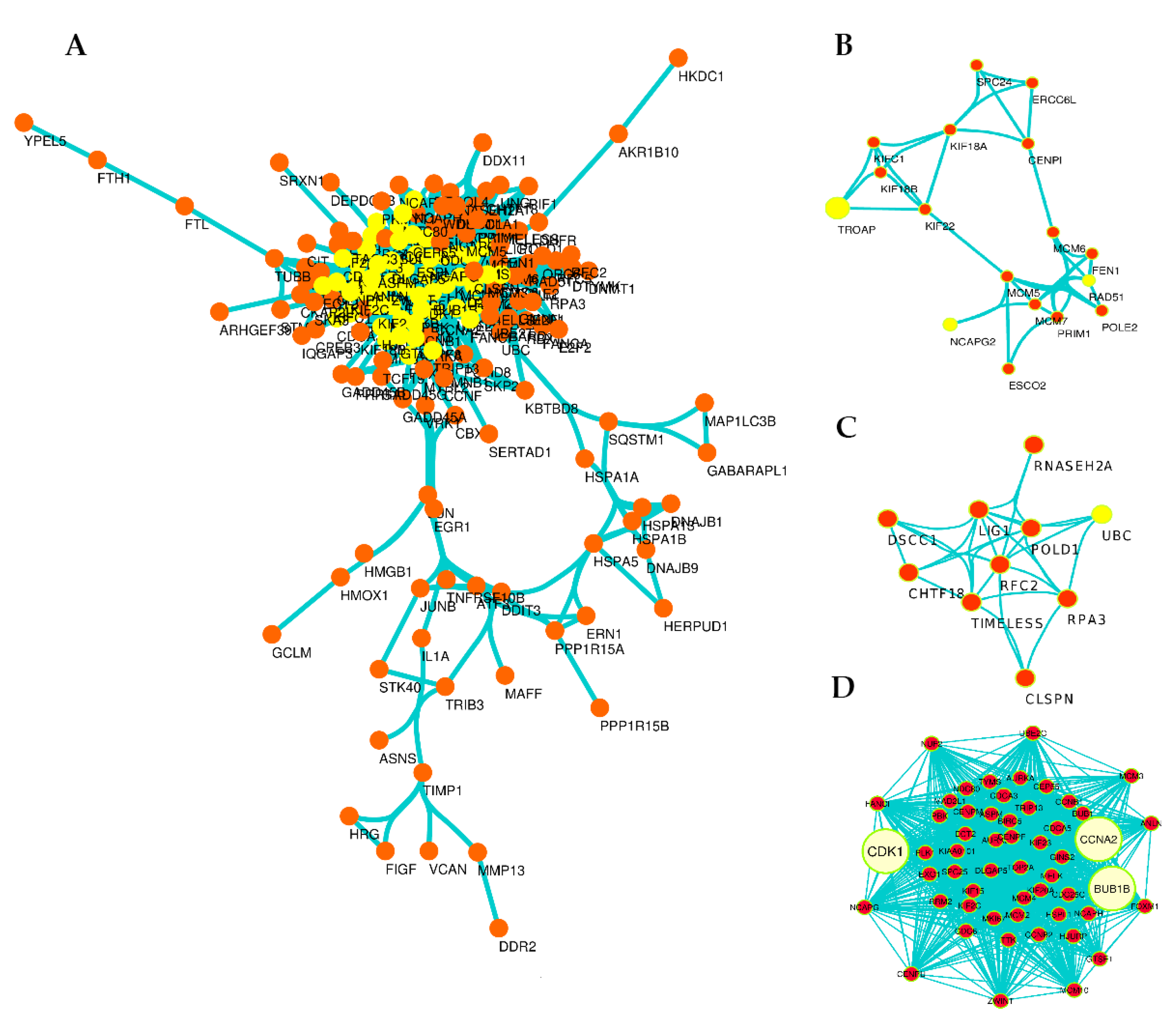
3.4. Exploration of DEGs in Protein–Protein Interaction (PPI) Network and Subnetworks (Modules)
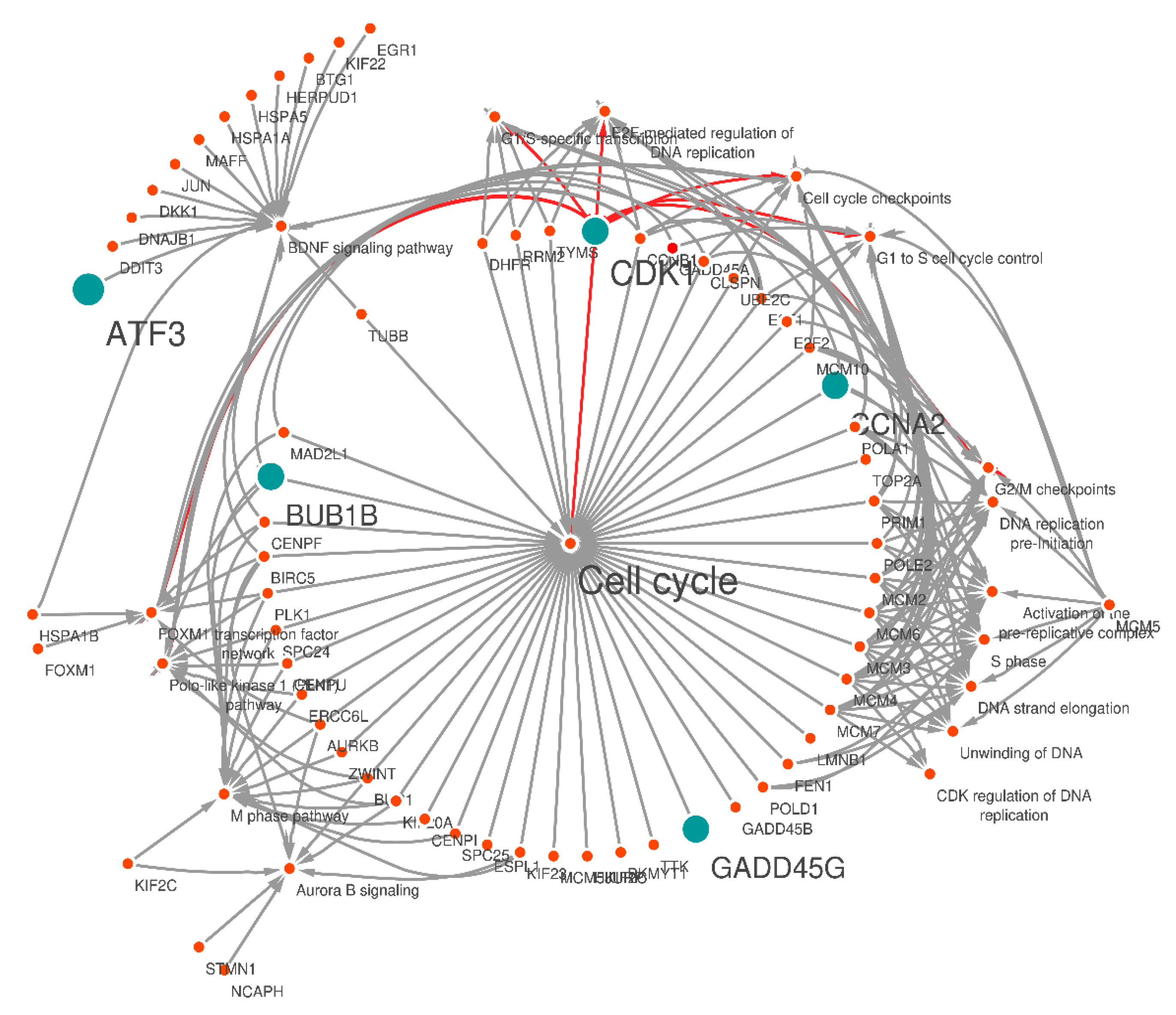
3.5. KEGG Pathway Analysis of DEGs
3.6. Functional Enrichment and Gene Ontology (GO) Analysis
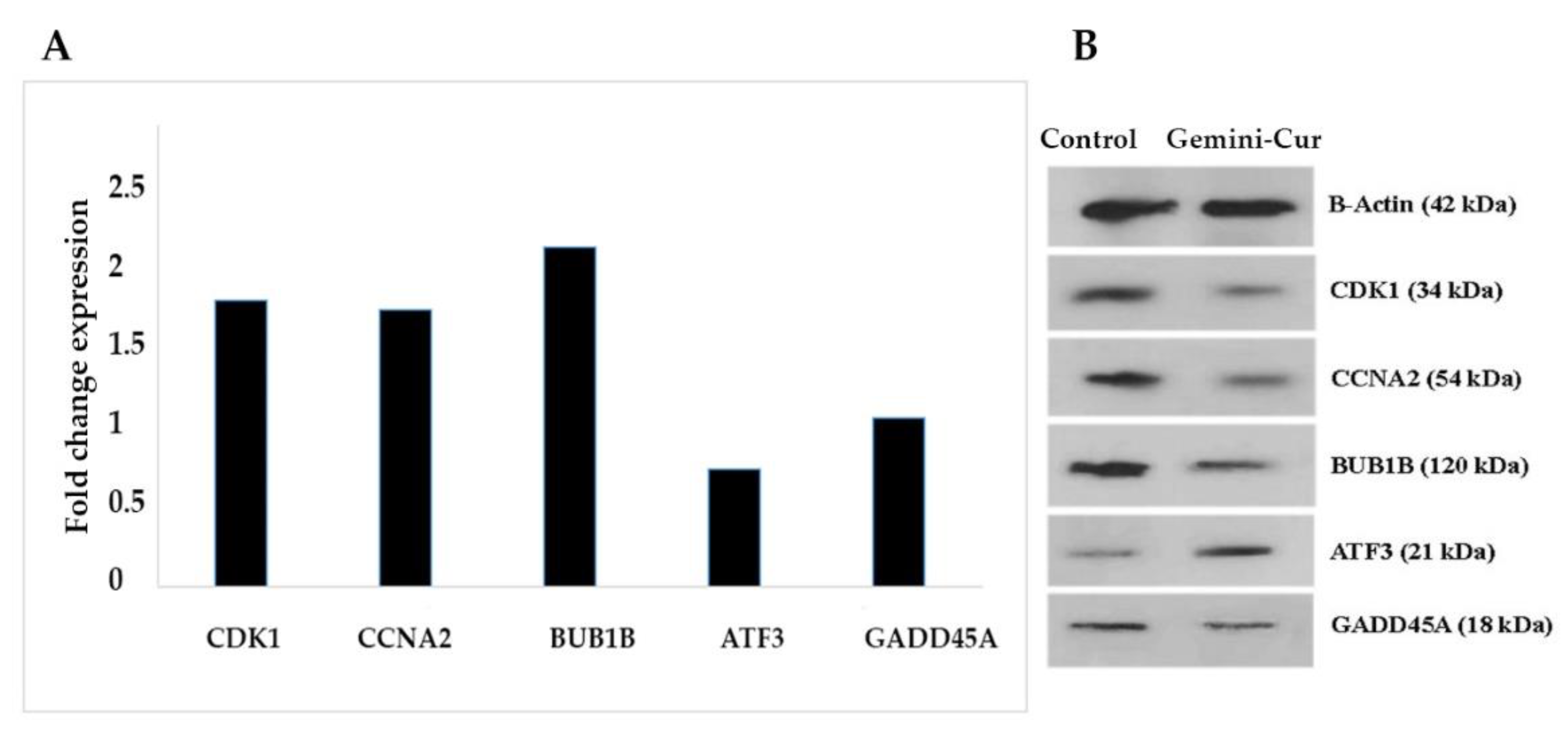
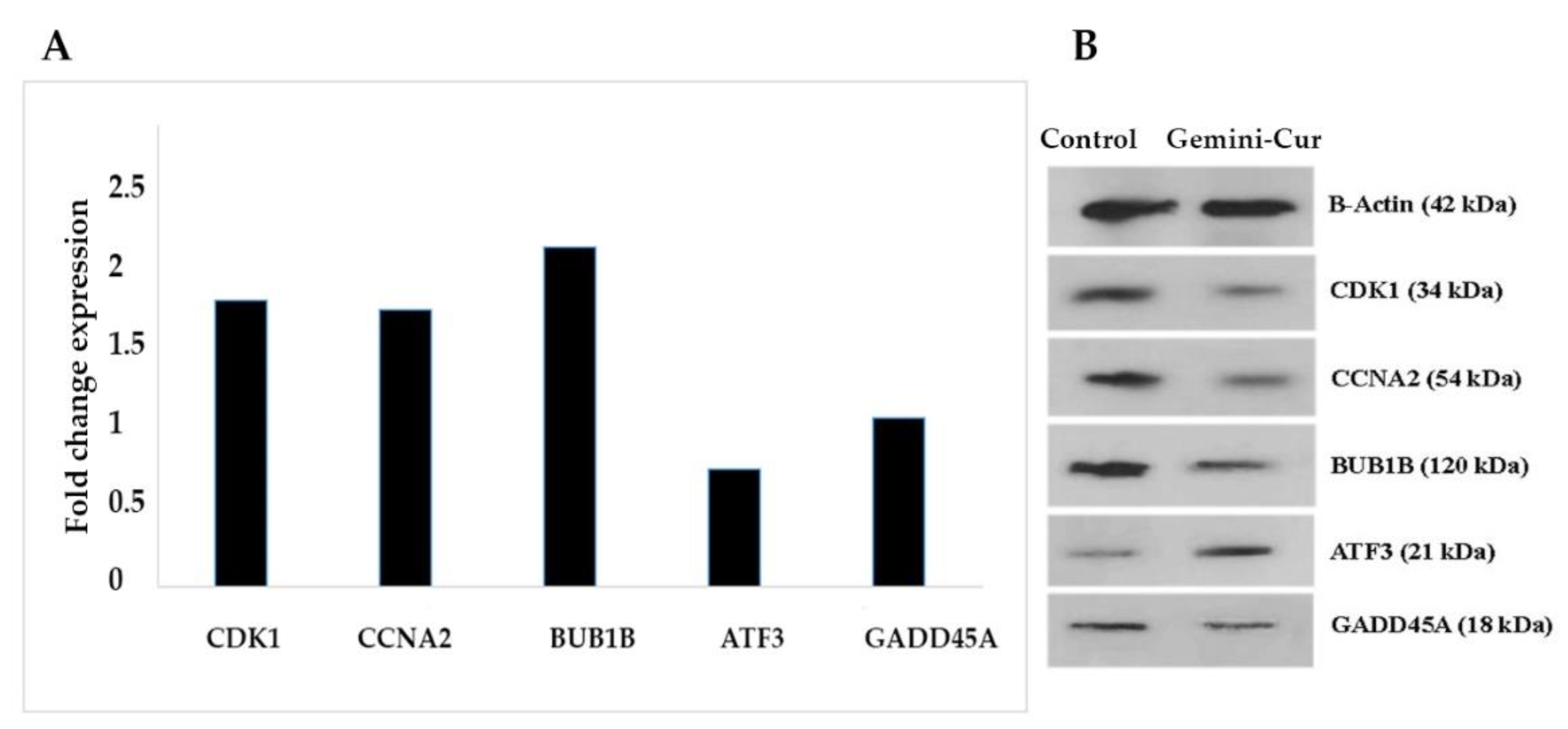
In Vitro Validation Study by Real-Time PCR and Western Blotting
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Singh, M.P.; Rai, S.; Singh, N.K.; Srivastava, S. Transcriptomic landscape of early age onset of colorectal cancer identifies novel genes and pathways in Indian CRC patients. Sci. Rep. 2021, 11, 11765. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Wu, R.; Gaspar, J.M.; Sargsyan, D.; Su, Z.-Y.; Zhang, C.; Gao, L.; Cheng, D.; Li, W.; Wang, C.; et al. DNA methylome and transcriptome alterations and cancer prevention by curcumin in colitis-accelerated colon cancer in mice. Carcinogenesis 2018, 39, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Passirani, C.; Vessières, A.; La Regina, G.; Link, W.; Silvestri, R. Modulating undruggable targets to overcome cancer therapy resistance. Drug Resist. Updat. 2022, 60, 100788. [Google Scholar] [CrossRef]
- Obrand, D.I.; Gordon, P.H. Incidence and patterns of recurrence following curative resection for colorectal carcinoma. Dis. Colon Rectum 1997, 40, 15–24. [Google Scholar] [CrossRef]
- André, T.; Boni, C.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Bonetti, A.; Clingan, P.; Bridgewater, J.; Rivera, F.; et al. Improved Overall Survival with Oxaliplatin, Fluorouracil, and Leucovorin as Adjuvant Treatment in Stage II or III Colon Cancer in the MOSAIC Trial. J. Clin. Oncol. 2009, 27, 3109–3116. [Google Scholar] [CrossRef] [Green Version]
- Pallag, A.; Roşca, E.; Ţiţ, D.M.; Muţiu, G.; Bungau, S.G.; Pop, O.L. Monitoring the effects of treatment in colon cancer cells using immunohistochemical and histoenzymatic techniques. Rom. J. Morphol. Embryol. 2015, 56, 1103–1109. [Google Scholar]
- Zhong, Z.; Vong, C.T.; Chen, F.; Tan, H.; Zhang, C.; Wang, N.; Cui, L.; Wang, Y.; Feng, Y. Immunomodulatory potential of natural products from herbal medicines as immune checkpoints inhibitors: Helping to fight against cancer via multiple targets. Med. Res. Rev. 2022, 42, 1246–1279. [Google Scholar] [CrossRef]
- Sivasankarapillai, V.S.; Nair, R.M.K.; Rahdar, A.; Bungau, S.; Zaha, D.C.; Aleya, L.; Tit, D.M. Overview of the anticancer activity of withaferin A, an active constituent of the Indian ginseng Withania somnifera. Environ. Sci. Pollut. Res. 2020, 27, 26025–26035. [Google Scholar] [CrossRef]
- Kumar, A.; Harsha, C.; Parama, D.; Girisa, S.; Daimary, U.D.; Mao, X.; Kunnumakkara, A.B. Current clinical developments in curcumin-based therapeutics for cancer and chronic diseases. Phytother. Res. 2021, 35, 6768–6801. [Google Scholar] [CrossRef]
- Nocito, M.C.; De Luca, A.; Prestia, F.; Avena, P.; La Padula, D.; Zavaglia, L.; Sirianni, R.; Casaburi, I.; Puoci, F.; Chimento, A.; et al. Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability. Biomedicines 2021, 9, 1476. [Google Scholar] [CrossRef]
- D’Angelo, N.A.; Noronha, M.A.; Kurnik, I.S.; Câmara, M.C.; Vieira, J.M.; Abrunhosa, L.; Martins, J.T.; Alves, T.F.; Tundisi, L.L.; Ataide, J.A.; et al. Curcumin encapsulation in nanostructures for cancer therapy: A 10-year overview. Int. J. Pharm. 2021, 604, 120534. [Google Scholar] [CrossRef]
- Ghaderi, S.; Babaei, E.; Hussen, B.M.; Mahdavi, M.; Azeez, H.J. Gemini Curcumin Suppresses Proliferation of Ovarian Cancer OVCAR-3 Cells via Induction of Apoptosis. Anti-Cancer Agents Med. Chem. 2021, 21, 775–781. [Google Scholar] [CrossRef]
- Emami, A.; Babaei, E.; Nagishbandi, A.; Azeez, H.J.; Feizi, M.A.H.; Golizadeh, A. Cellular uptake and apoptotic properties of gemini curcumin in gastric cancer cells. Mol. Biol. Rep. 2021, 48, 7215–7222. [Google Scholar] [CrossRef]
- Karimpour, M.; Feizi, M.A.H.; Mahdavi, M.; Krammer, B.; Verwanger, T.; Najafi, F.; Babaei, E. Development of curcumin-loaded gemini surfactant nanoparticles: Synthesis, characterization and evaluation of anticancer activity against human breast cancer cell lines. Phytomedicine 2019, 57, 183–190. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Babaei, E.; Neri, F.; Feizi, M.A.H. Anti-proliferative and apoptotic effect of gemini curcumin in p53-wild type and p53-mutant colorectal cancer cell lines. Int. J. Pharm. 2021, 601, 120592. [Google Scholar] [CrossRef]
- Yu, L.-L.; Wu, J.-G.; Dai, N.; Yu, H.G.; Si, J.M. Curcumin reverses chemoresistance of human gastric cancer cells by downregulating the NF-?B transcription factor. Oncol. Rep. 2011, 26, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Inoue, M.; Uchida, Y.; Edagawa, M.; Hirata, M.; Mitamura, J.; Miyamoto, D.; Taketani, K.; Sekine, S.; Kawauchi, J.; Kitajima, S. The stress response gene ATF3 is a direct target of the Wnt/β-catenin pathway and inhibits the invasion and migration of HCT116 human colorectal cancer cells. PLoS ONE 2018, 13, e0194160. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Dong, Z.; Guo, Y.; Chen, Z.; Kuang, G.; Yang, Z. Methylation-mediated repression of GADD45A and GADD45G expression in gastric cardia adenocarcinoma. Int. J. Cancer 2013, 133, 2043–2053. [Google Scholar] [CrossRef]
- Gan, W.; Zhao, H.; Li, T.; Liu, K.; Huang, J. CDK1 interacts with iASPP to regulate colorectal cancer cell proliferation through p53 pathway. Oncotarget 2017, 8, 71618–71629. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Huang, Y.; Zhang, Y.; Zeng, X.; Yan, M.; Xia, Z.; Lai, D. Correction: DPP3/CDK1 contributes to the progression of colorectal cancer through regulating cell proliferation, cell apoptosis, and cell migration. Cell Death Dis. 2021, 12, 529, Erratum in Cell Death Dis. 2021, 12, 623. [Google Scholar] [CrossRef]
- Sung, W.-W.; Lin, Y.-M.; Wu, P.-R.; Yen, H.-H.; Lai, H.-W.; Su, T.-C.; Huang, R.-H.; Wen, C.-K.; Chen, C.-Y.; Chen, C.-J.; et al. High nuclear/cytoplasmic ratio of Cdk1 expression predicts poor prognosis in colorectal cancer patients. BMC Cancer 2014, 14, 951. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, K.; Zhang, J.; Wang, L.; Sheng, L.; Yan, L. Inhibition of CDK1 Reverses the Resistance of 5-Fu in Colorectal Cancer. Cancer Manag. Res. 2020, 12, 11271–11283. [Google Scholar] [CrossRef]
- Thoma, O.-M.; Neurath, M.F.; Waldner, M.J. Cyclin-Dependent Kinase Inhibitors and Their Therapeutic Potential in Colorectal Cancer Treatment. Front. Pharmacol. 2021, 12, 757120. [Google Scholar] [CrossRef]
- Dai, S.; Mo, Y.; Wang, Y.; Xiang, B.; Liao, Q.; Zhou, M.; Li, X.; Li, Y.; Xiong, W.; Li, G.; et al. Chronic Stress Promotes Cancer Development. Front. Oncol. 2020, 10, 1492. [Google Scholar] [CrossRef]
- Gan, Y.; Li, Y.; Li, T.; Shu, G.; Yin, G. CCNA2 acts as a novel biomarker in regulating the growth and apoptosis of colorectal cancer. Cancer Manag. Res. 2018, 10, 5113–5124. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, L.; Liu, Y.; Yang, L.; Jiang, D.; Li, K.; Xie, S.; Wang, X.; Wang, S. Comprehensive Analysis of Cyclin Family Gene Expression in Colon Cancer. Front. Oncol. 2021, 11, 674394. [Google Scholar] [CrossRef]
- Ding, X.; Duan, H.; Luo, H. Identification of Core Gene Expression Signature and Key Pathways in Colorectal Cancer. Front. Genet. 2020, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- Vairapandi, M.; Balliet, A.G.; Hoffman, B.; Liebermann, D.A. GADD45b and GADD45g are cdc2/cyclinB1 kinase inhibitors with a role in S and G2/M cell cycle checkpoints induced by genotoxic stress. J. Cell. Physiol. 2002, 192, 327–338. [Google Scholar] [CrossRef]
- Taylor, W.R.; Stark, G.R. Regulation of the G2/M transition by p53. Oncogene 2001, 20, 1803–1815. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′→3′) | PCR Product |
|---|---|---|
| CDK1 | F: 5-AGCCGGGATCTACCATACC-3 R: 5-CATGGCTACCACTTGACCTG-3 | 126 |
| CCNA2 | F: 5-GGACAAAGCTGGCCTGAATC-3 R: 5-CTGTTGTGCATGCTGTGGTG-3 | 116 |
| BUB1B | F: 5-CAATTCCAAGCTCGAGTGTC-3 R: 5-GATGATTGGAGCTCTTGCTG-3 | 146 |
| GADD45G | F: 5-GTCAGCCAAAGTCTTGAACG-3 R: 5-GCACTATGTCGATGTCGTTC-3 | 145 |
| ATF3 | F: 5-CAGCACCTTGCCCCAAAATC -3 R: 5-TGGATGGCAAACCTCAGCTC-3 | 171 |
| β-actin | F: 5-CAGCACCTTGCCCCAAAATC -3 R: 5-TGGATGGCAAACCTCAGCTC-3 | 184 |
| Gene Symbol | STATUS | Base Mean | log2 Fold Change | lfcSE | Stat | p Value | padj |
|---|---|---|---|---|---|---|---|
| HMOX1 | Up | 14082.63 | 4.581563 | 0.393634 | 11.63913 | 2.61 × 10−31 | 9.29 × 10−27 |
| DDIT3 | Up | 7029.257 | 4.083832 | 0.394679 | 10.34723 | 4.31 × 10−25 | 7.68 × 10−21 |
| ATF3 | Up | 5160.583 | 2.807268 | 0.27917 | 10.05577 | 8.66 × 10−24 | 1.01 × 10−19 |
| CHAC1 | Up | 2994.215 | 3.896157 | 0.400672 | 9.724061 | 2.38 × 10−22 | 2.12 × 10−18 |
| HERPUD1 | Up | 12540.28 | 2.267609 | 0.254122 | 8.92332 | 4.53 × 10−19 | 3.23 × 10−19 |
| SPX | Up | 260.3101 | 4.781735 | 0.578882 | 8.260295 | 1.45 × 10−16 | 8.64 × 10−13 |
| DNAJB9 | Up | 2736.219 | 2.665123 | 0.342986 | 7.770348 | 7.83 × 10−15 | 3.99 × 10−11 |
| OSGIN1 | Up | 13215.95 | 4.534403 | 0.586017 | 7.737659 | 1.01 × 10−14 | 4.51 × 10−11 |
| LUCAT1 | Up | 608.1075 | 3.02517 | 0.396356 | 7.632456 | 2.30 × 10−14 | 9.13 × 10−11 |
| CYP4F3 | Up | 547.2113 | 2.671816 | 0.369558 | 7.22977 | 4.84 × 10−13 | 1.57 × 10−09 |
| TONSL | Down | 2059.716 | −2.89597 | 0.395525 | −7.32183 | 2.45 × 10−13 | 8.72 × 10−10 |
| MCM4 | Down | 5519.294 | −2.8152 | 0.421581 | −6.67772 | 2.43 × 10−11 | 4.5 5× 10−08 |
| AC112777.1 | Down | 221.6533 | −3.06376 | 0.459517 | −6.66735 | 2.60 × 10−11 | 4.64 × 10−08 |
| FANCA | Down | 1932.18 | −2.67477 | 0.406894 | −6.57362 | 4.91 × 10−11 | 7.47 × 10−08 |
| E2F2 | Down | 629.9052 | −2.62378 | 0.402383 | −6.52061 | 7.00 × 10−11 | 9.25 × 10−08 |
| BLM | Down | 995.8815 | −2.69881 | 0.413601 | −6.52516 | 6.79 × 10−11 | 9.25 × 10−08 |
| NCAPH | Down | 1459.514 | −3.16292 | 0.490122 | −6.45332 | 1.09 × 10−10 | 1.32 × 10−07 |
| KIF18B | Down | 1583.582 | −3.07757 | 0.477409 | −6.44639 | 1.15 × 10−10 | 1.32 × 10−07 |
| UHRF1 | Down | 2198.428 | −2.78382 | 0.432416 | −6.43782 | 1.21 × 10−10 | 1.32 × 10−07 |
| CDCA3 | Down | 933.7762 | −3.48616 | 0.542823 | −6.42228 | 1.34 × 10−10 | 1.41 × 10−07 |
| Gene Symbol | Expression Based on RNA-seq | Degree (K) | Betweenness (BC) | Closeness Centrality (CC) | MCODE: Score | No; Cluster |
|---|---|---|---|---|---|---|
| CDK1 | Down | 106 | 0.94541487 | 0.885964912 | 34.77520814 | 1 |
| CCNA2 | Down | 96 | 0.299119893 | 0.789115646 | 34.77520814 | 1 |
| BUB1B | Down | 90 | 0.296979671 | 0.724550898 | 34.77520814 | 1 |
| GADD45G | Up | 5 | 0 | 0 | 5 | 5 |
| ATF3 | Up | 6 | 0 | 0.411764706 | 2.7 | 8 |
| Pathway | Gene Symbol | Padj |
|---|---|---|
| Cell cycle | TOP2A; ERCC6L; MCM7; PRIM1; HJURP; BUB1B; MCM10; TTK; PKMYT1; TYMS; AURKB; LMNB1; CCNB1; POLD1; E2F1; E2F2; CLSPN; BUB1; CENPURRM2; GADD45B; GADD45A; UBE2C; TUBB; PLK1; KIF23; ZWINT; GADD45G; DHFR, CCNA2; POLA1; CENPF; ESPL1; CENPI; POLE2; CDK1; MCM3; MCM4; BIRC5; MCM5; KIF2C; KIF20A; MCM6; SPC24; MCM2; SPC25; MAD2L1 | 1.82 × 10−34 |
| E2F-mediated regulation of DNA replication | DHFR; POLA1; CCNB1; RRM2; PRIM1; E2F1; CDK1; TYMS | 1.67 × 10−08 |
| G1/S-specific transcription | DHFR; POLA1; RRM2; CDK1; E2F1; TYMS | 1.79 × 10−07 |
| Cell cycle checkpoints | MCM7; UBE2C; BUB1B; MCM10; CCNB1; CDK1; MCM3; MCM4; MCM5; CLSPN; MCM6; MCM2; MAD2L1 | 2.84 × 10−09 |
| G2/M checkpoints | CCNB1; MCM7; CDK1; MCM3; MCM4; MCM10; MCM5; CLSPN; MCM6; MCM2 | 2.89 × 10−10 |
| FOXM1 transcription factor network | CCNA2; CCNB1; CENPF; PLK1; CDK1; BIRC5; FOXM1; AURKB; HSPA1B | 4.54 × 10−09 |
| G1 to S cell cycle control | DHFR; POLA1; RRM2; CDK1; E2F1; TYMS | 3.83 × 10−12 |
| GO Term | Source | Padj | Gene Symbol |
|---|---|---|---|
| microtubule cytoskeleton organization involved in mitosis | BP | 3.94 × 10−23 | ERCC6L; BUB1B; CDCA8; TTK; CENPA; TACC3; BIRC5; CENM; KIF2C; SPC24; MAD2L1; |
| mitotic spindle organization | BP | 1.75 × 10−22 | ERCC6L; BUB1B; CDCA8; TTK; CENPA; KIF2C; SPC24; MAD2L1; SPC25 |
| mitotic sister chromatid segregation | BP | 1.37 × 10−21 | SPAG5; CDCA5; P LK1; NCAPG2; CDCA8; NCAPG; PSRC1; ESPL1; KIFC1; PRC1; CDK1; KIF2C |
| DNA metabolic process | BP | 8.96 × 10−20 | TOP2A; BLM; FEN1; RNASEH2A; MCM7; UHRF1; HMGB2; MCM10; TYMS; CDK1; MCM4; MCM5; MCM6; MCM2 |
| positive regulation of cell cycle process | BP | 9.66 × 10−10 | UBE2C; TUBB; CCNF; PLK1; CDC7; CDC25C; PKMYT1; FOXM1; AURKA; CCNA2; CCNB2; CCNB1; CDK1; E2F1; TACC3; NEK2; CDKN3 |
| spindle | CC | 2.01 × 10−18 | SPAG5; CKAP2L; PLK1; BUB1B; CDC7; KIF23; TTK; KIF22; SKA3; AURKB; AURKA; CDC20; CENPF; CDK1; TACC3; BIRC5; KIF2C; KIF20A |
| nucleus | CC | 2.20 × 10−13 | TOP2A; ARHGAP11A; FEN1; MCM7; DSCC1; CCNF; NCAPG2; HMGB2; MCM10; CCNA2; ASPM; MCM5; KIF20A; MCM6; PRR11; MCM2; BLM; RAD51; PRC1; UBE2T; CDK1; TRIP13; MAD2L1 |
| intracellular non-membrane-bounded organelle | CC | 4.72 × 10−10 | TOP2A; FEN1; MCM7; CDCA5; HJURP; HMGB2; BUB1B; CDCA8; MCM10; TTK; MKI67; PKMYT1; AURKB; PLK1; VRK1; KIF23; ESCO2; PIMREG; CIT; CENPF; PSRC1; PRC1; UBE2T; KIF2C; KIF20A; TRIP13; SPC24; MCM2 |
| cyclin-dependent protein kinase holoenzyme complex | CC | 4.89 × 10−05 | CCNA2; CCNB2; CCNB1; CCNF; CDK1 |
| serine/threonine protein kinase complex | CC | 1.32 × 10−04 | CCNA2; CCNB2; CCNB1; CCNF; CDK1 |
| histone kinase activity | MF | 7.17 × 10−04 | CDK1; AURKB; AURKA |
| protein serine/threonine kinase activity | MF | 0.001470884 | PLK1; CDK1; PBK; NEK2; VRK1; CDC7; TTK; PASK; PKMYT1; AURKB; CIT; AURKA |
| kinase binding | MF | 0.004957845 | CAV1; PLK1; VRK1; CDC25C; FOXM1; AURKB; CIT; AURKA; ARHGAP33; CCNA2 |
| cyclin-dependent protein serine/threonine kinase regulator activity | MF | 0.00820989 | CCNA2; CCNB2; CCNB1; CCNF |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azeez, H.J.; Neri, F.; Hosseinpour Feizi, M.A.; Babaei, E. Transcriptome Profiling of HCT-116 Colorectal Cancer Cells with RNA Sequencing Reveals Novel Targets for Polyphenol Nano Curcumin. Molecules 2022, 27, 3470. https://doi.org/10.3390/molecules27113470
Azeez HJ, Neri F, Hosseinpour Feizi MA, Babaei E. Transcriptome Profiling of HCT-116 Colorectal Cancer Cells with RNA Sequencing Reveals Novel Targets for Polyphenol Nano Curcumin. Molecules. 2022; 27(11):3470. https://doi.org/10.3390/molecules27113470
Chicago/Turabian StyleAzeez, Hewa Jalal, Francesco Neri, Mohammad Ali Hosseinpour Feizi, and Esmaeil Babaei. 2022. "Transcriptome Profiling of HCT-116 Colorectal Cancer Cells with RNA Sequencing Reveals Novel Targets for Polyphenol Nano Curcumin" Molecules 27, no. 11: 3470. https://doi.org/10.3390/molecules27113470
APA StyleAzeez, H. J., Neri, F., Hosseinpour Feizi, M. A., & Babaei, E. (2022). Transcriptome Profiling of HCT-116 Colorectal Cancer Cells with RNA Sequencing Reveals Novel Targets for Polyphenol Nano Curcumin. Molecules, 27(11), 3470. https://doi.org/10.3390/molecules27113470