
Anti-Inflammatory Effects of Mitrephora sirikitiae Leaf Extract and Isolated Lignans in RAW 264.7 Cells

 ,
,
Abstract
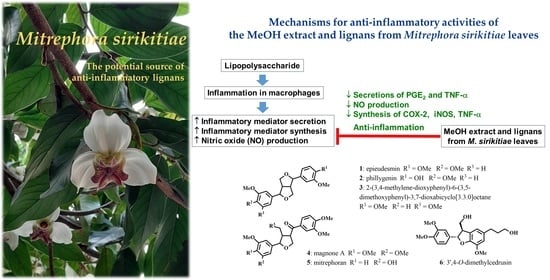
:
1. Introduction
2. Results and Discussion
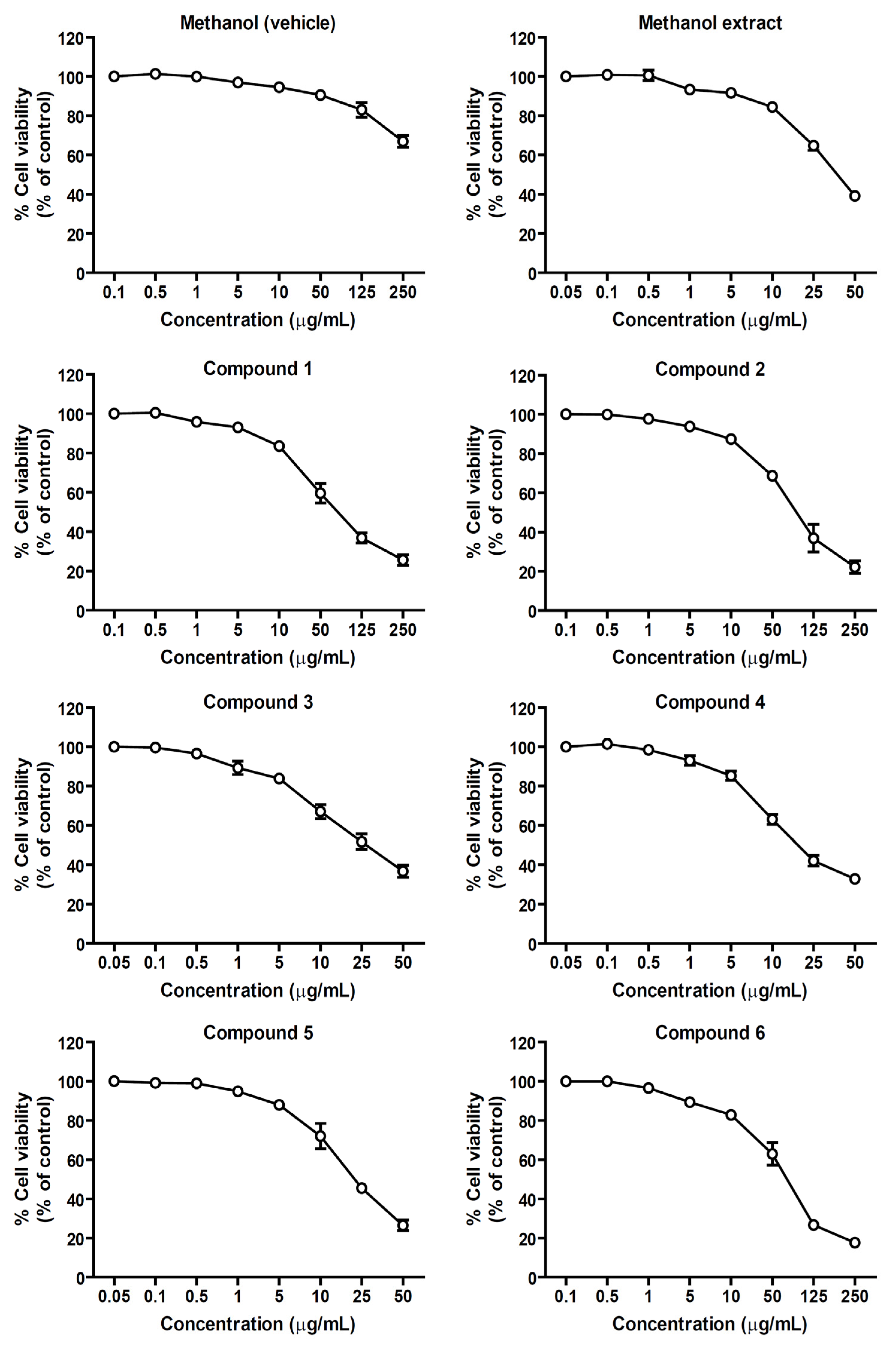
2.1. Effects of the Methanol Extract and Isolated Lignans on Cytotoxicity
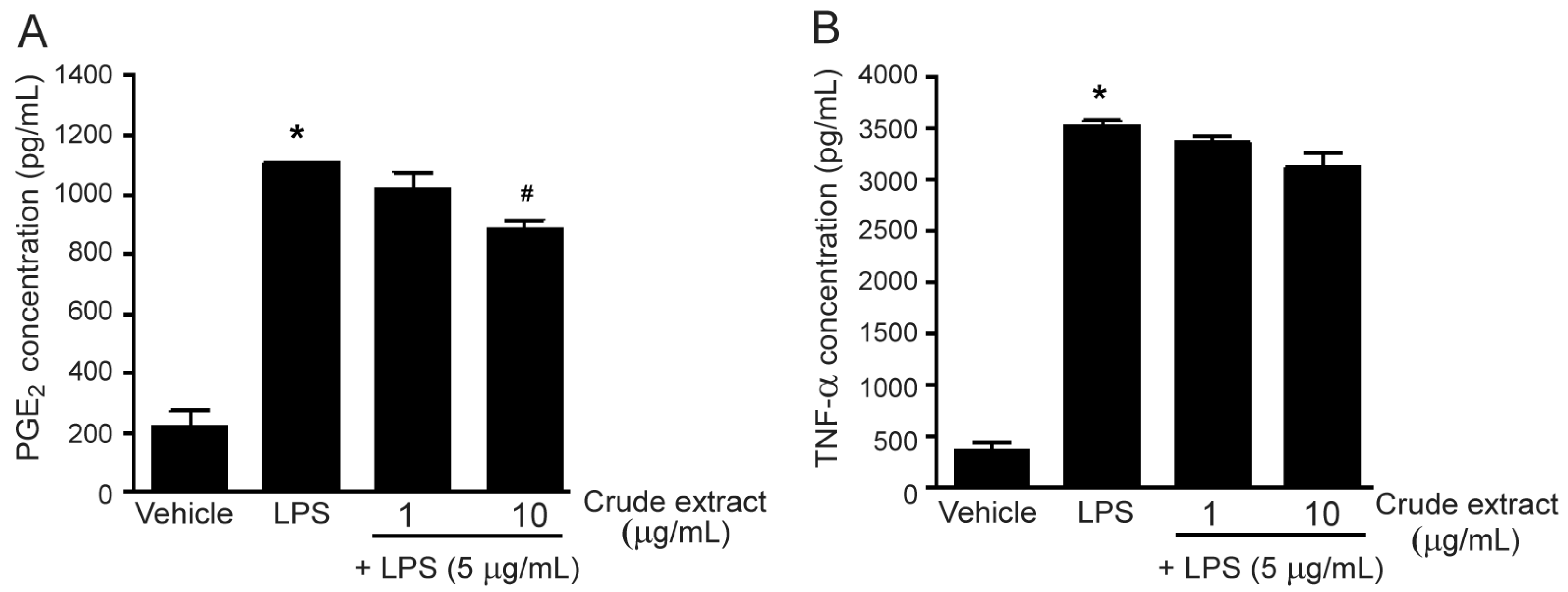
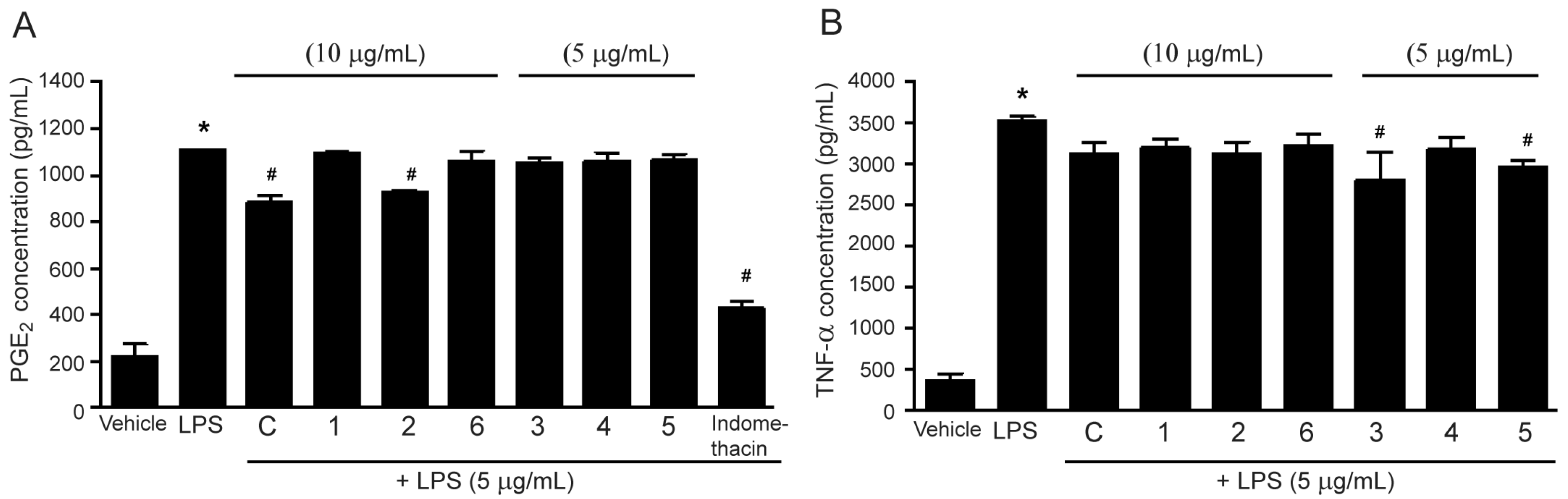
2.2. Effects of the methanol Extract and Isolated Lignans on LPS-Induced PGE2 and TNF-α Secretion
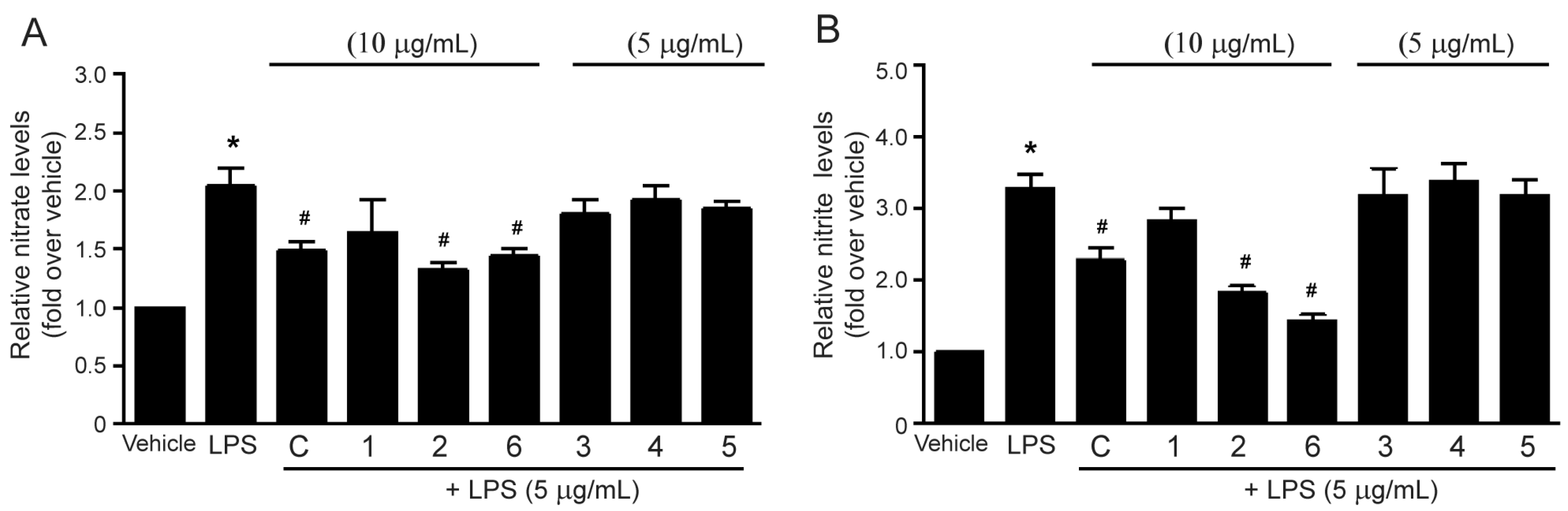
2.3. Effects of the Methanol Extract and Isolated Lignans on LPS-Induced Nitric Oxide Production
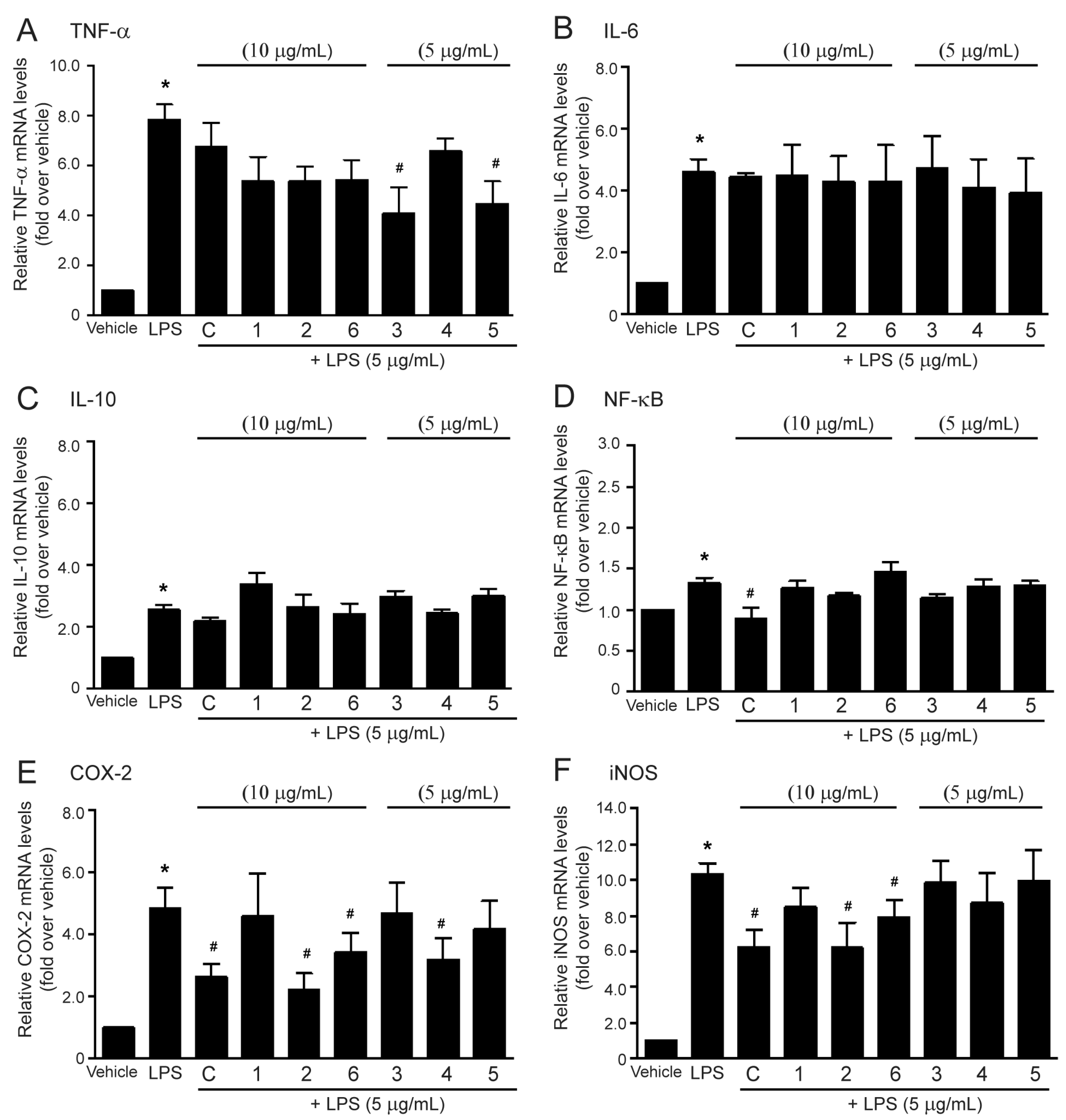
2.4. Effects of the Methanol Extract and Isolated Lignans on LPS-Induced mRNA Expressions of Inflammatory Biomarkers
3. Materials and Methods
3.1. Chemicals and Reagents
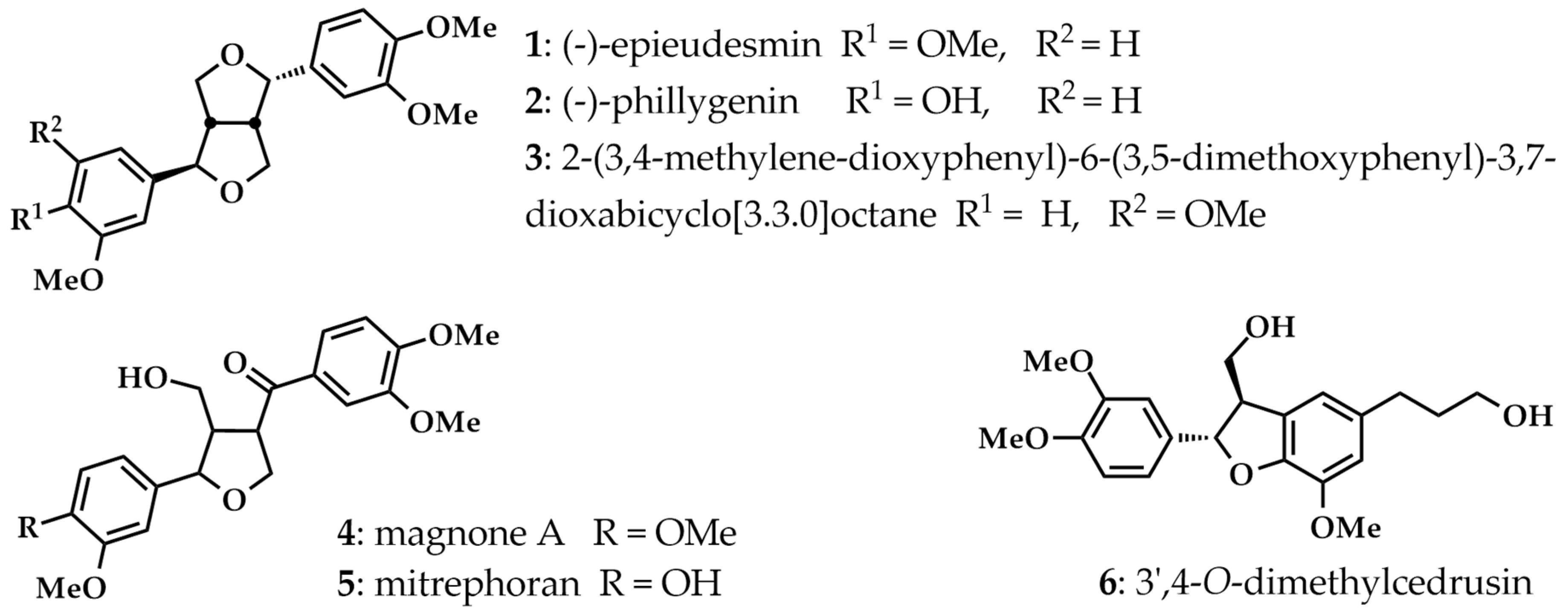
3.2. Plant Materials and Isolated Lignans
3.3. Cell Culture and Treatment
3.4. Determination of Cytotoxicity by MTT Assay
3.5. Enzyme-Linked Immunosorbent Assay (ELISA) for PGE2 and TNF-α Measurement
3.6. Measurement of Nitric Oxide Production
3.7. Measurement of mRNA Expression of Inflammatory Biomarkers
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Tripathi, P.; Kashyap, L.; Singh, V. The role of nitric oxide in infammatory reactions. FEMS Immunol. Med. Microbiol. 2007, 51, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in inflammatory disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S.; et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol. Med. Rep. 2018, 17, 5484–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.Y.; Wen, M.H. Lipopolysaccharide-mediated reactive oxygen species and signal transduction in the regulation of interleukin-1 gene expression. J. Biol. Chem. 2002, 277, 22131–22139. [Google Scholar] [CrossRef] [Green Version]
- Libby, P. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr. Rev. 2007, 65, 140–146. [Google Scholar] [CrossRef]
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [Green Version]
- Weerasooriya, A.D.; Chalermglin, P.; Saunders, R.M.K. Mitrephora sirikitiae (Annonaceae): A remarkable new species endemic to northern Thailand. Nord. J. Bot. 2004, 24, 201–206. [Google Scholar] [CrossRef]
- Anantachoke, N.; Lovacharaporn, D.; Reutrakul, V.; Michel, S.; Gaslonde, T.; Piyachaturawat, P.; Suksen, K.; Prabpai, S.; Nuntasaen, N. Cytotoxic compounds from the leaves and stems of the endemic Thai plant Mitrephora sirikitiae. Pharm. Biol. 2020, 58, 490–497. [Google Scholar] [CrossRef]
- Barker, D. Lignans. Molecules 2019, 24, 1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Toledo, E.; Delgado-Rodríguez, M.; Gaforio, J.J. Naturally lignan-rich foods: A dietary tool for health promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.H.; Zhao, P.; Wang, M.; Liang, Q. Naturally occurring furofuran lignans: Structural diversity and biological activities. Nat. Prod. Res. 2019, 33, 1357–1373. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Dziurka, M.; Warzecha, A.; Kubica, P.; Klimek-Szczykutowicz, M.; Ekiert, H. Targeted lignan profiling and anti-inflammatory properties of Schisandra rubriflora and Schisandra chinensis extracts. Molecules 2018, 23, 3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.; Lee, J.G.; Lee, S.H.; Kim, Y.S.; Kim, H.P. Anti-inflammatory activity of phylligenin, a lignan from the fruits of Forsythia koreana, and its cellular mechanism of action. J. Ethnopharmacol. 2008, 118, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Shin, J.S.; Chung, K.S.; Lee, Y.G.; Baek, N.I.; Lee, K.T. Anti-inflammatory mechanisms of koreanaside A, a lignan isolated from the flower of Forsythia koreana, against LPS-induced macrophage activation and DSS-induced colitis mice: The crucial role of AP-1, NF-κB, and JAK/STAT signaling. Cells 2019, 8, 1163. [Google Scholar] [CrossRef] [Green Version]
- Dong, D.D.; Li, H.; Jiang, K.; Qu, S.J.; Tang, W.; Tan, C.H.; Li, Y.M. Diverse lignans with anti-inflammatory activity from Urceola rosea. Fitoterapia 2019, 134, 96–100. [Google Scholar] [CrossRef]
- Lee, D.Y.; Seo, K.H.; Jeong, R.H.; Lee, S.M.; Kim, G.S.; Noh, H.J.; Kim, S.Y.; Kim, G.W.; Kim, J.Y.; Baek, N.I. Anti-inflammatory lignans from the fruits of Acanthopanax sessiliflorus. Molecules 2013, 18, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.J.; Chen, X.L.; Li, X.S.; Su, J.; He, J.B.; Wang, Y.H.; Li, Y.; Cheng, Y.X. Two dimeric lignans with an unusual α,β-unsaturated ketone motif from Zanthoxylum podocarpum and their inhibitory effects on nitric oxide production. Bioorg. Med. Chem. Lett. 2011, 21, 373–376. [Google Scholar] [CrossRef]
- Rhee, S.H. Lipopolysaccharide: Basic biochemistry, intracellular signaling, and physiological impacts in the gut. Intest. Res. 2014, 12, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Jang, D.; Murrell, G. Nitric oxide in arthritis. Free Radic. Biol Med. 1998, 24, 1511–1519. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar] [PubMed]
- Ben-Baruch, A. Inflammation-associated immune suppression in cancer: The roles played by cytokines, chemokines and additional mediators. Semin. Cancer Biol. 2006, 16, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Clària, J.; Romano, M. Pharmacological intervention of cyclooxygenase-2 and 5-lipoxygenase pathways. Impact on inflammation and cancer. Curr. Pharm. Des. 2005, 11, 3431–3447. [Google Scholar] [CrossRef]
- Teponno, R.B.; Kusari, S.; Spiteller, M. Recent advances in research on lignans and neolignans. Nat. Prod. Rep. 2016, 33, 1044–1092. [Google Scholar] [CrossRef] [Green Version]
- During, A.; Debouche, C.; Raas, T.; Larondelle, Y. Among plant lignans, pinoresinol has the strongest antiinflammatory properties in human intestinal Caco-2 cells. J. Nutr. 2012, 142, 1798–1805. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.S.; Hira, K.; Ahil, S.B.; Kulkarni, O.P.; Araya, H.; Fujimoto, Y. New synthetic coumarinolignans as attenuators of pro-inflammatory cytokines in LPS-induced sepsis and carrageenan-induced paw oedema models. Inflammopharmacology 2020, 28, 1365–1373. [Google Scholar] [CrossRef]
- Sato, V.H.; Sungthong, B.; Nuamnaichati, N.; Rinthong, P.; Mangmool, S.; Sato, H. In vivo and in vitro evidence for the antihyperuricemic, anti-inflammatory and antioxidant effects of a Traditional Ayurvedic Medicine, Triphala. Nat. Prod. Commun. 2017, 12, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Wongthai, N.; Tanticharakunsiri, W.; Mangmool, S.; Ochaikul, D. Characteristics and antioxidant activity of royal lotus pollen, butterfly pea flower, and oolong tea kombucha beverages. Asia Pac. J. Sci. Technol. 2021, 26, APST-26-04-17. [Google Scholar] [CrossRef]
- Nuamnaichati, N.; Parichatikanond, W.; Mangmool, S. Cardioprotective effects of glucagon-like peptide-1 (9-36) against oxidative injury in H9c2 cardiomyoblasts: Potential role of the PI3K/Akt/NOS pathway. J. Cardiovasc. Pharmacol. 2022, 79, e50–e63. [Google Scholar] [CrossRef] [PubMed]
- Mangmool, S.; Kunpukpong, I.; Kitphati, W.; Anantachoke, N. Antioxidant and anticholinesterase activities of extracts and phytochemicals of Syzygium antisepticum leaves. Molecules 2021, 26, 3295. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene-Specific Primers | Sequences | |
|---|---|---|
| COX-2 | Sense | 5′-tgcatgtggctgtggatgtcatcaa-3′ |
| Antisense | 5′-cactaagacagacccgtc atctcca-3′ | |
| IL-6 | Sense | 5′-gacaaagccagagtccttcagagag-3′ |
| Antisense | 5′-ctaggtttgccgagtagatctc-3′ | |
| IL-10 | Sense | 5′-gctggacaacatactgctaacc-3′ |
| Antisense | 5′-atttccgataaggcttggcaa-3′ | |
| iNOS | Sense | 5′-gtgttccaccaggagatgttg-3′ |
| Antisense | 5′-ctcctgcccactgagttcgtc-3′ | |
| NF-κB | Sense | 5′-gaaattcctgatccagacaaaaac-3′ |
| Antisense | 5′-atcacttcaatggcctctgtgtag-3′ | |
| TNF-α | Sense | 5′-atgagcacagaaagcatgatc-3′ |
| Antisense | 5′-tacaggcttgtcactcgaatt-3′ | |
| GAPDH | Sense | 5′-gcctgcttcaccaccttc-3′ |
| Antisense | 5′-ggctctccagaacatcatcc-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangmool, S.; Limpichai, C.; Han, K.K.; Reutrakul, V.; Anantachoke, N. Anti-Inflammatory Effects of Mitrephora sirikitiae Leaf Extract and Isolated Lignans in RAW 264.7 Cells. Molecules 2022, 27, 3313. https://doi.org/10.3390/molecules27103313
Mangmool S, Limpichai C, Han KK, Reutrakul V, Anantachoke N. Anti-Inflammatory Effects of Mitrephora sirikitiae Leaf Extract and Isolated Lignans in RAW 264.7 Cells. Molecules. 2022; 27(10):3313. https://doi.org/10.3390/molecules27103313
Chicago/Turabian StyleMangmool, Supachoke, Chayaporn Limpichai, Khine Kyi Han, Vichai Reutrakul, and Natthinee Anantachoke. 2022. "Anti-Inflammatory Effects of Mitrephora sirikitiae Leaf Extract and Isolated Lignans in RAW 264.7 Cells" Molecules 27, no. 10: 3313. https://doi.org/10.3390/molecules27103313
APA StyleMangmool, S., Limpichai, C., Han, K. K., Reutrakul, V., & Anantachoke, N. (2022). Anti-Inflammatory Effects of Mitrephora sirikitiae Leaf Extract and Isolated Lignans in RAW 264.7 Cells. Molecules, 27(10), 3313. https://doi.org/10.3390/molecules27103313