
Hypoglycemic and Antioxidant Properties of Extracts and Fractions from Polygoni Avicularis Herba

Abstract
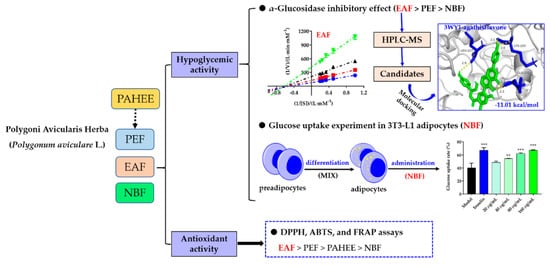
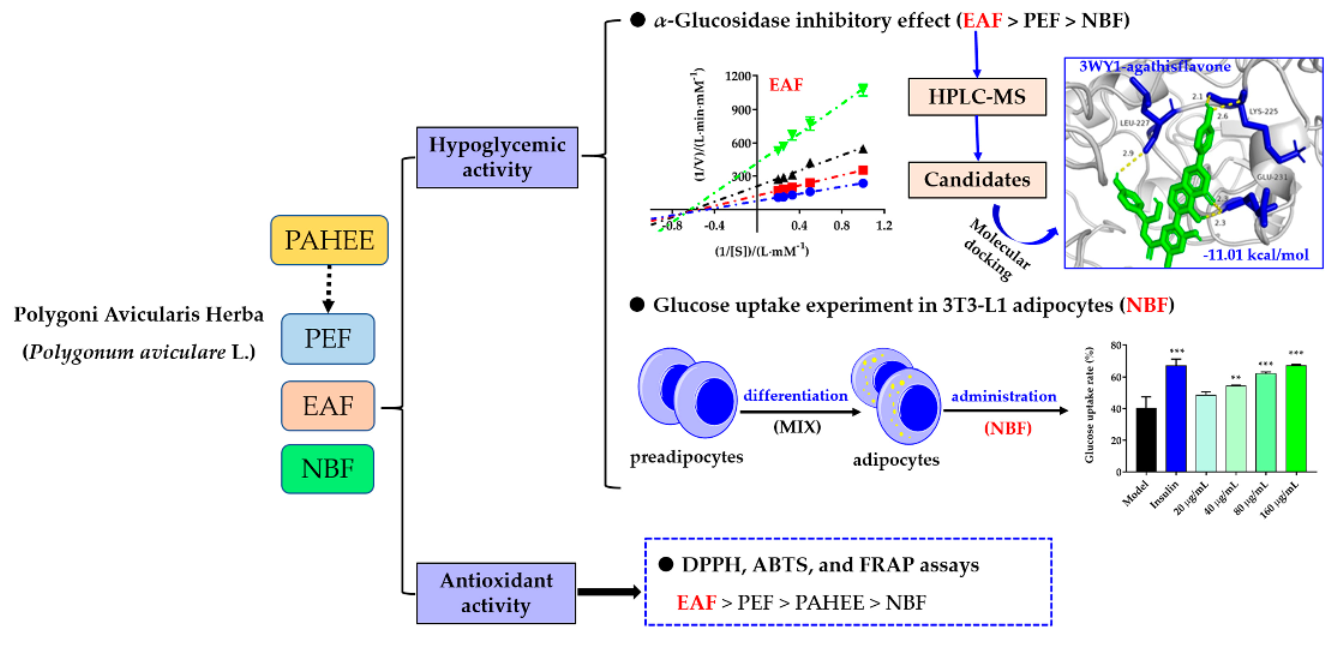
:
1. Introduction
2. Results
2.1. Total Flavonoids Content
2.2. Total Phenolics Content
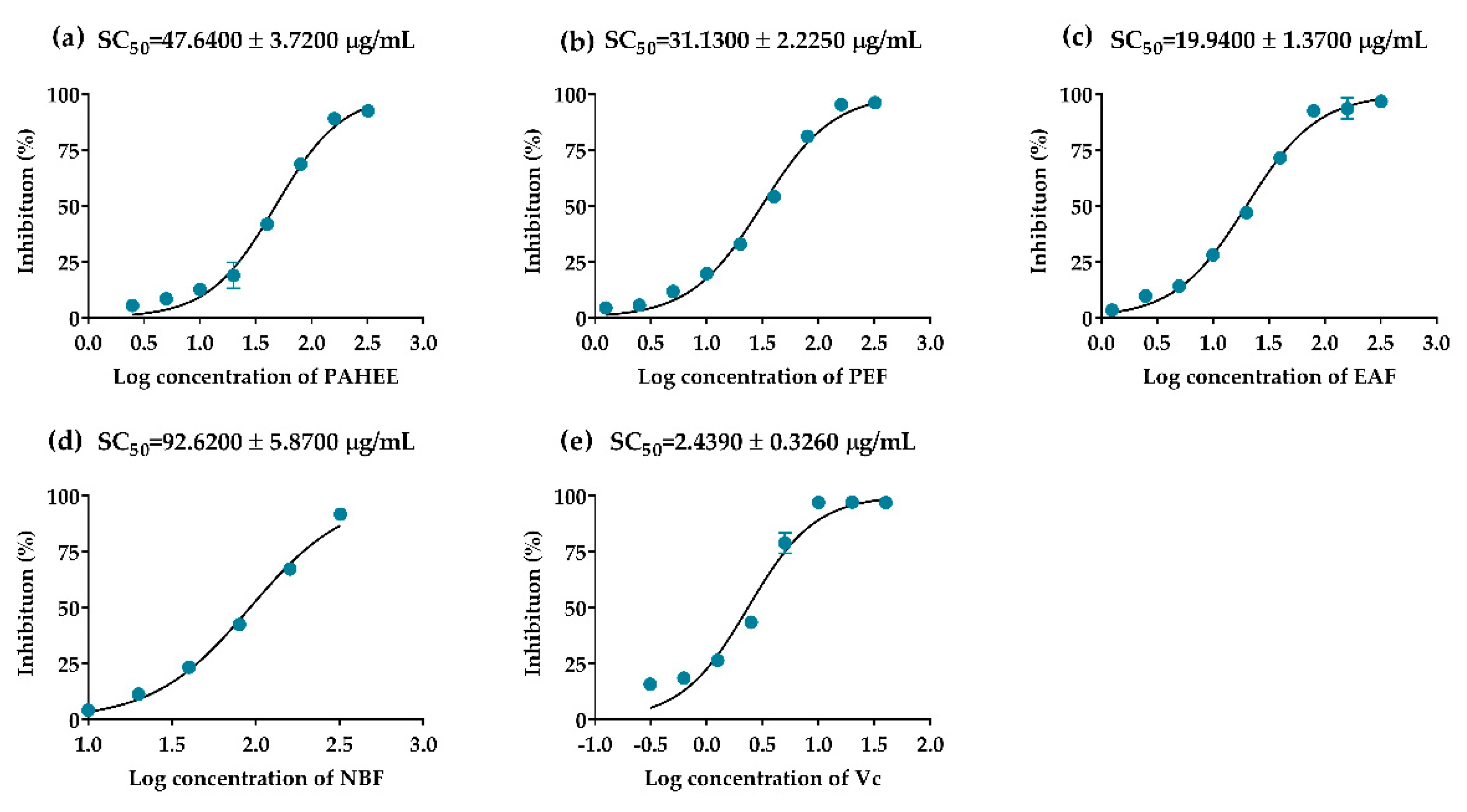
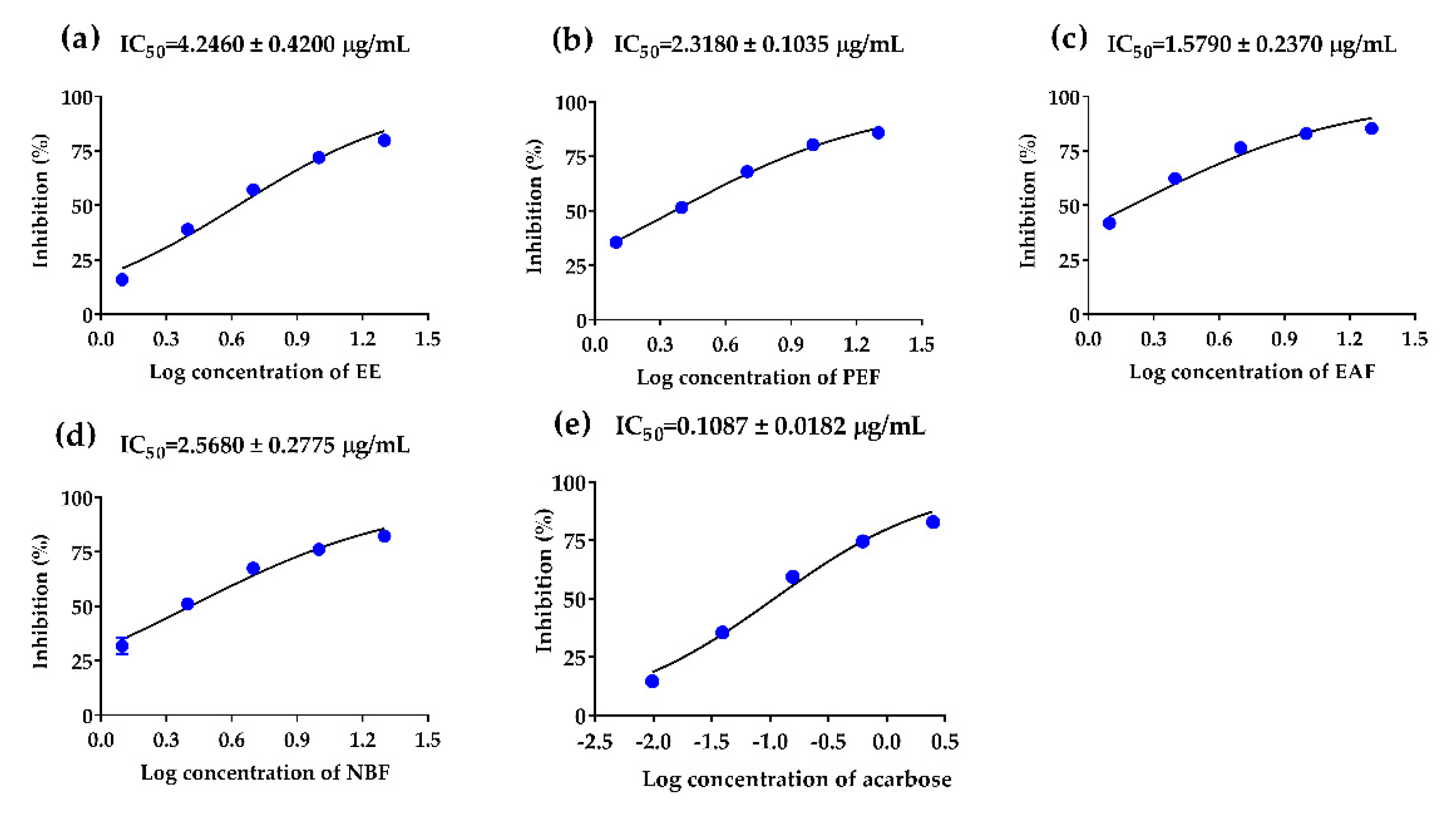
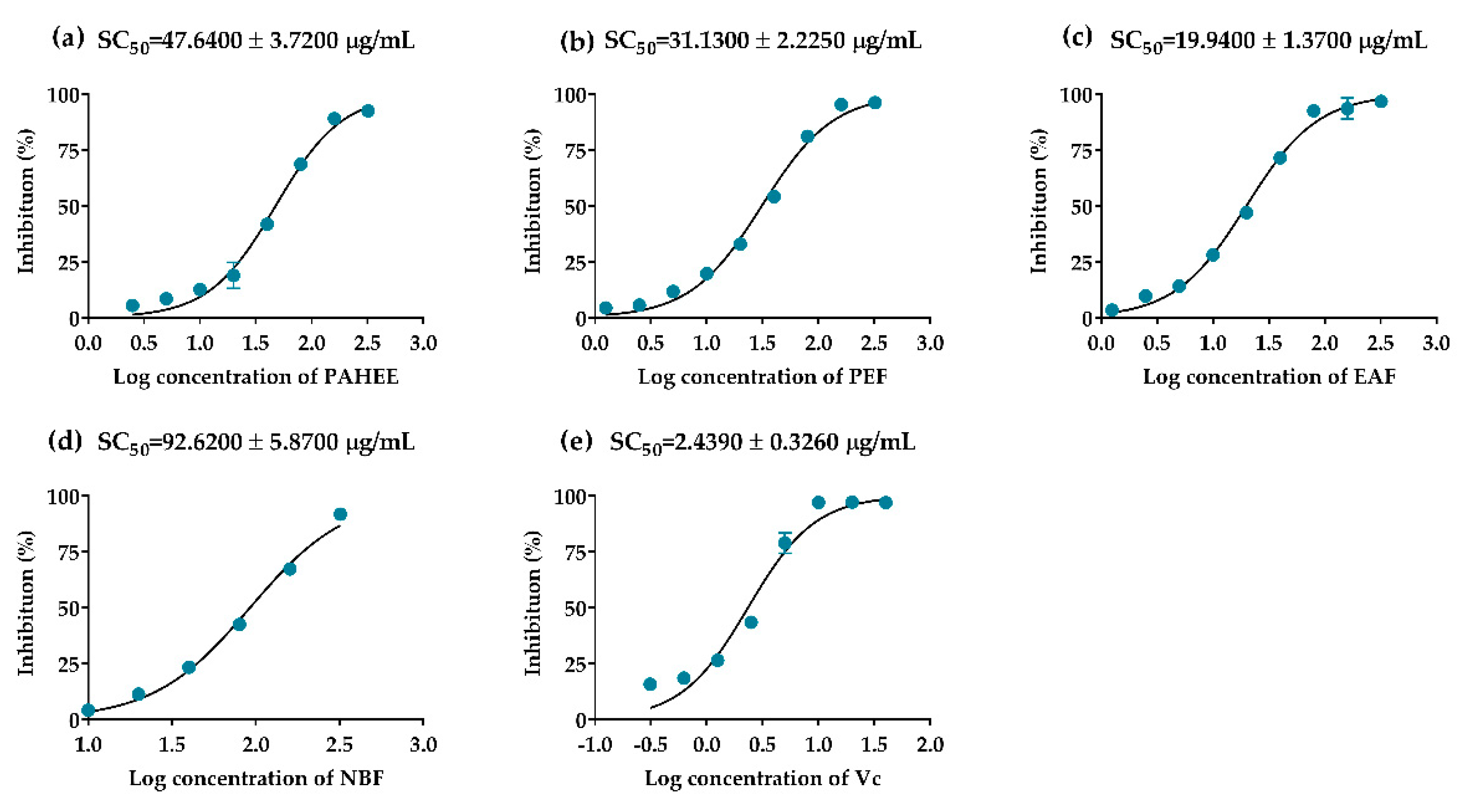
2.3. The Inhibitory Effects of PAHEE and Its Fractions on α-Glucosidase and α-Amylase
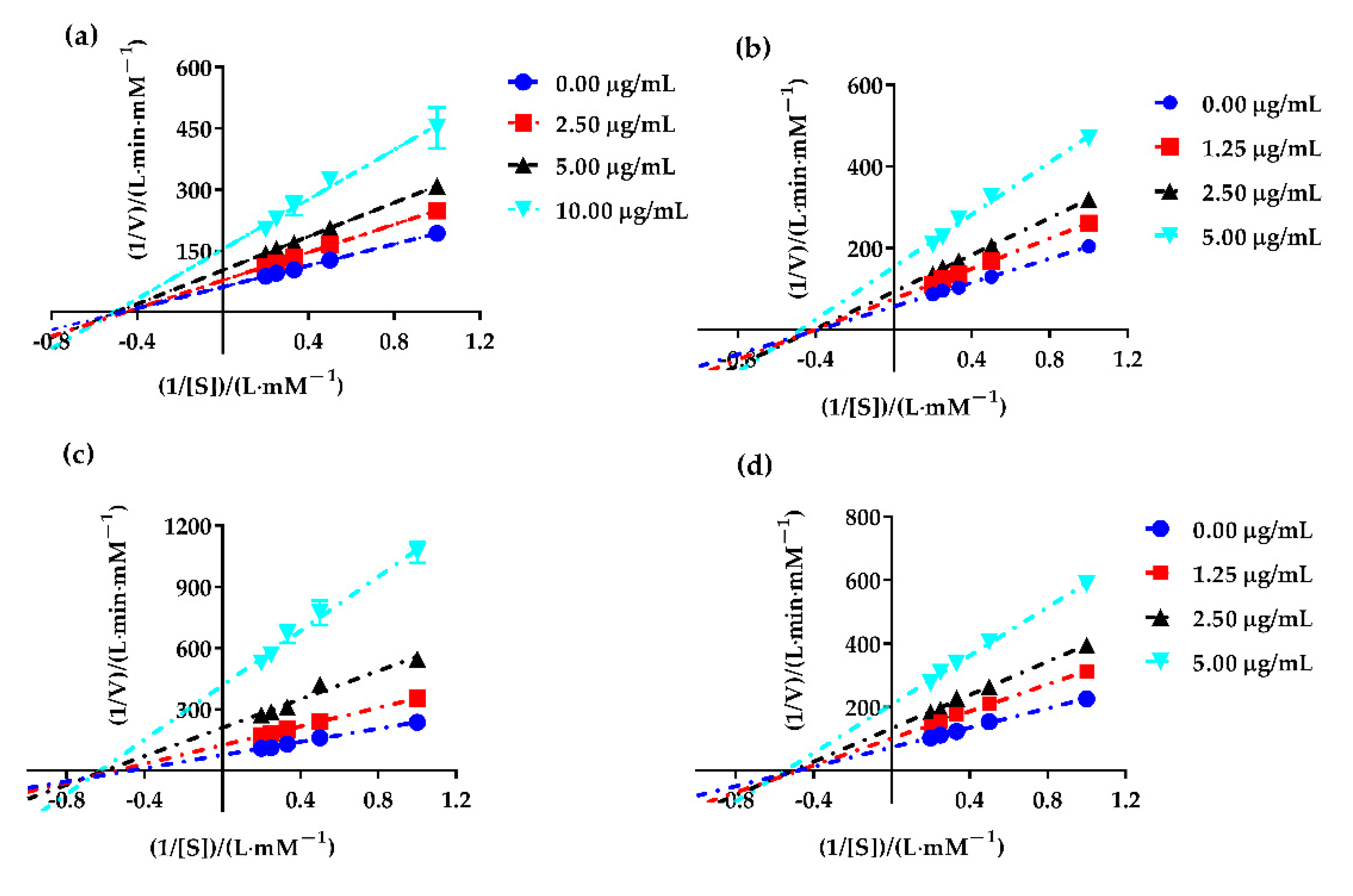
2.4. Enzyme Kinetic Equation
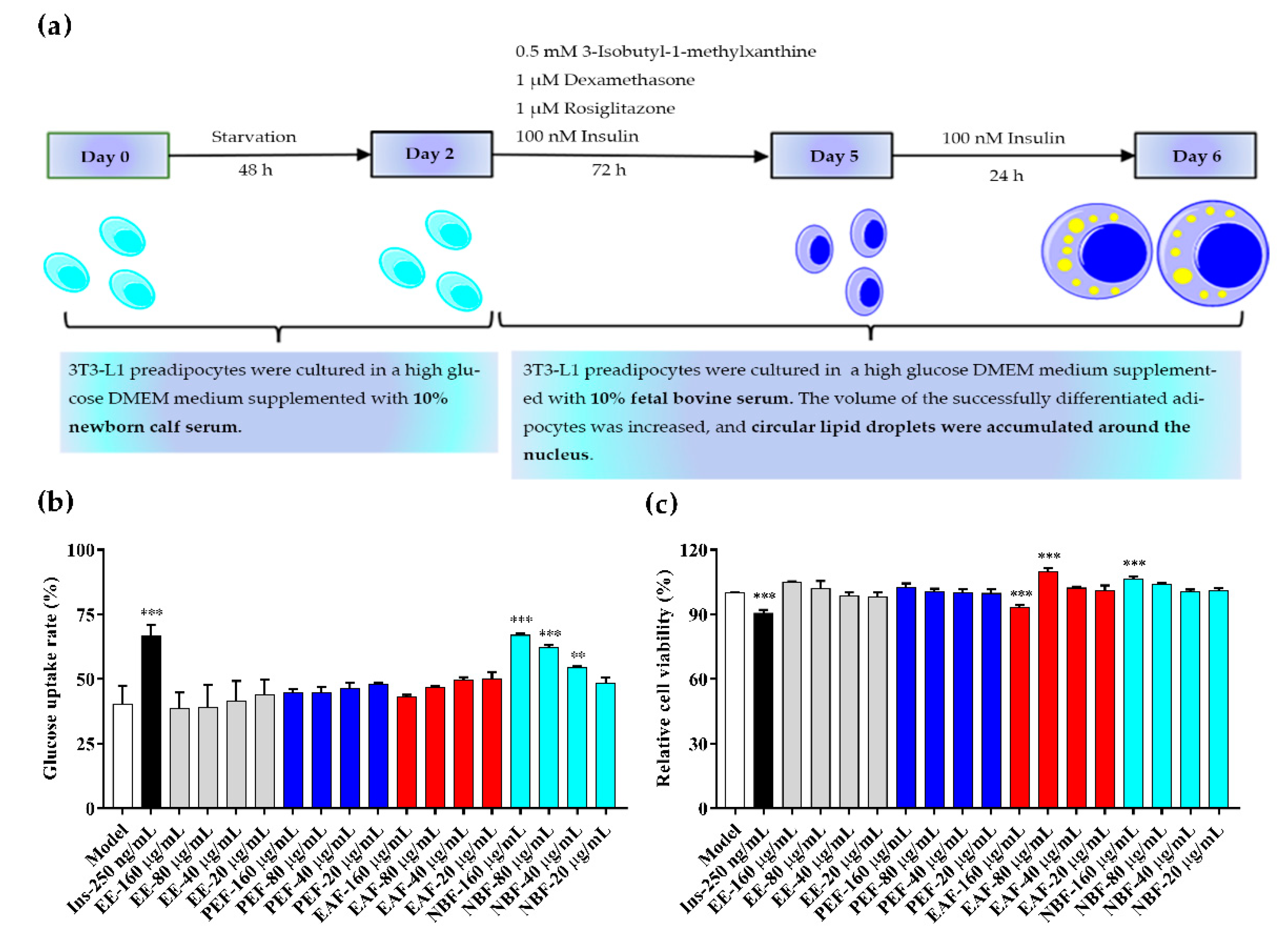
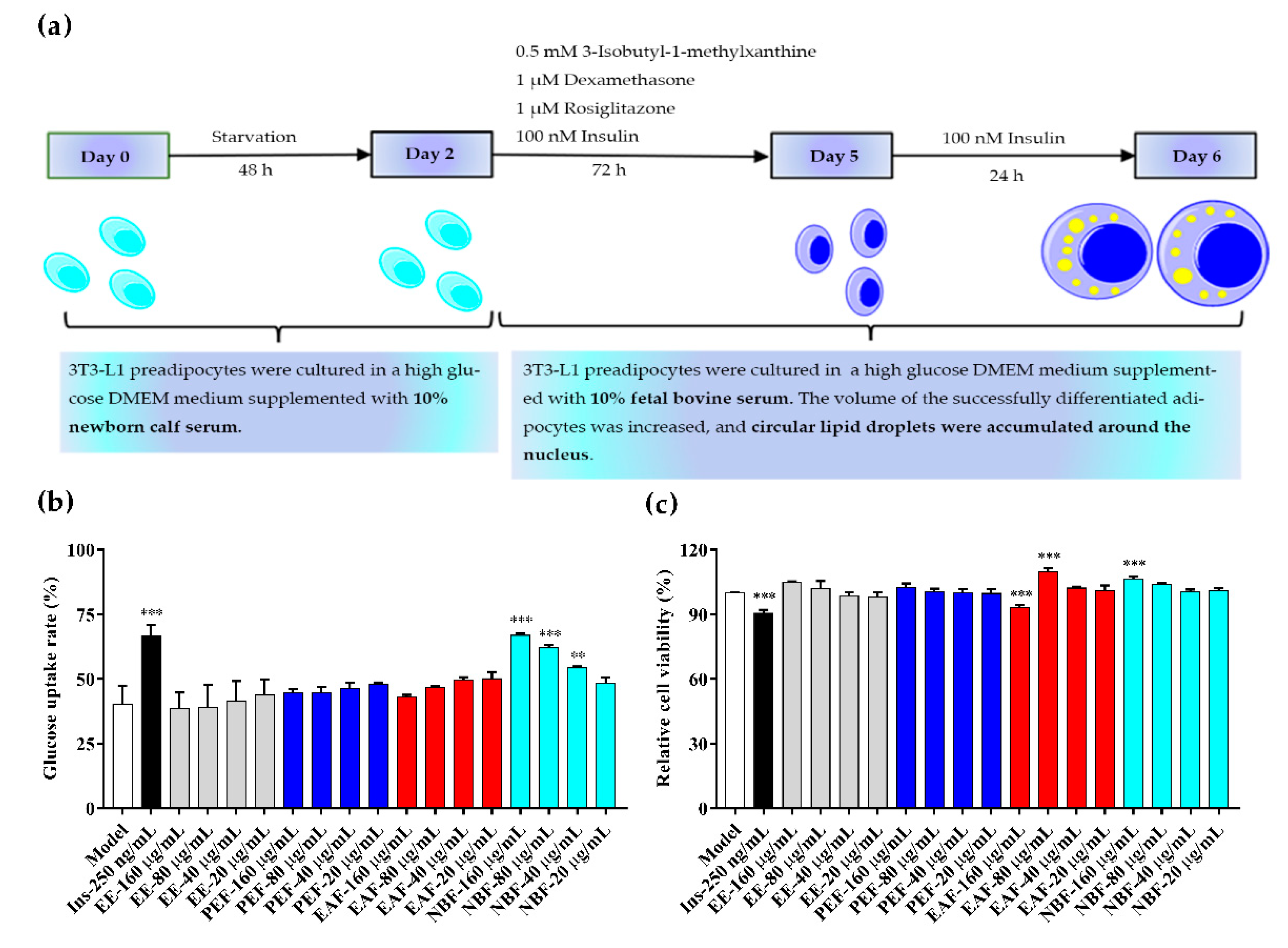
2.5. Glucose Uptake and Cell Viability Assays
2.6. HPLC-MS Analysis of EAF
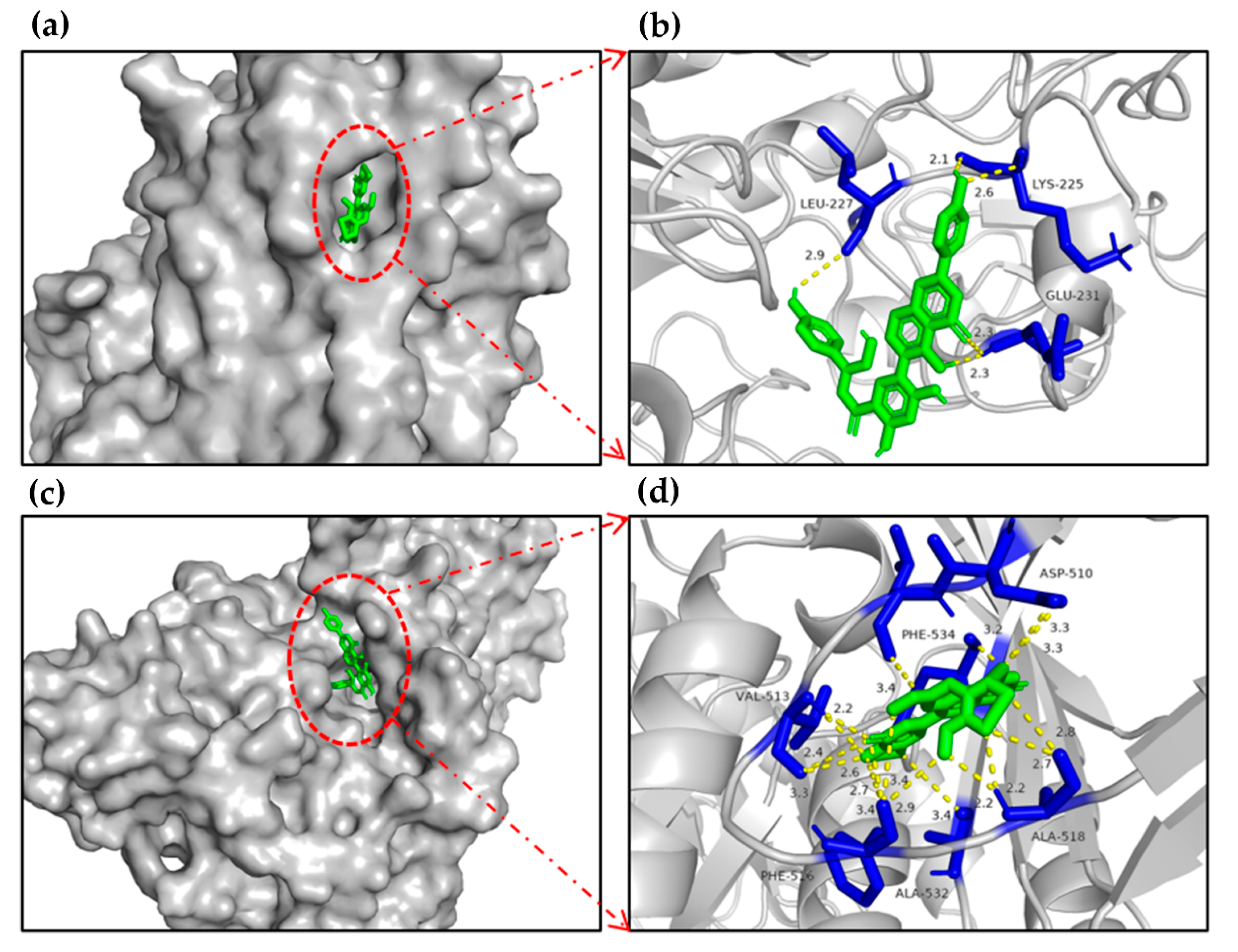
2.7. Molecular Docking of the Candidate Compounds on α-Glucosidase
2.8. DPPH-Free Radical Scavenging Assay
2.9. ABTS Radical Scavenging Assay
2.10. Ferric Reducing Antioxidation Power
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Plant Extracts
4.3. Determination of Total Flavonoids Content
4.4. Determination of Total Phenolics Content
4.5. α-Glucosidase Inhibition Experiments
4.6. Kinetic Analysis
4.7. Glucose Uptake and Cell Viability Assays
4.8. HPLC-MS Analysis of EAF
4.9. Molecular Docking of Candidate Compounds on α-Glucosidase
4.10. DPPH Free Radical Scavenging Assay
4.11. ABTS Radical Cation Scavenging Assay
4.12. Ferric Reducing Antioxidation Power
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Pharmacopoeia Commission. The Pharmacopoeia of the People’s Republic of China, 1st ed.; China Medical Science and Technology Press: Beijing, China, 2020. [Google Scholar]
- Granica, S.; Czerwińska, M.E.; Żyżyńska-Granica, B.; Kiss, A.K. Antioxidant and anti-inflammatory flavonol glucuronides from Polygonum aviculare L. Fitoterapia 2013, 91, 180–188. [Google Scholar] [CrossRef] [PubMed]
- González Begné, M.; Yslas, N.; Reyes, E.; Quiroz, V.; Santana, J.; Jimenez, G. Clinical effect of a Mexican sanguinaria extract (Polygonum aviculare L.) on gingivitis. J. Ethnopharmacol. 2001, 74, 45–51. [Google Scholar] [CrossRef]
- Salama, H.M.H.; Marraiki, N. Antimicrobial activity and phytochemical analyses of Polygonum aviculare L. (Polygonaceae), naturally growing in Egypt. Saudi. J. Biol. Sci. 2010, 17, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.Y. Antioxidant activity of extract from Polygonum aviculare L. Biol. Res. 2006, 39, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.S.; Kim, S.Y.; Kong, B.J.; Kim, K.J.; Noh, G.Y.; Im, N.R.; Lim, J.W.; Ha, J.H.; Kim, J.; Park, S.N. Cell penetrating peptide conjugated liposomes as transdermal delivery system of Polygonum aviculare L. extract. Int. J. Pharm. 2015, 483, 26–37. [Google Scholar] [CrossRef]
- Zheng, Z.S.; Yang, A.M.; Zhang, F.L.; Shang, H.L.; Li, Q. Study on the antioxidant and antibacterial activities of the extracts of Polygonum aviculare L. Acta Chin. Med. Pharmacol. 2019, 47, 55–58. [Google Scholar]
- Sung, Y.Y.; Yoon, T.; Yang, W.K.; Kim, S.J.; Kim, D.S.; Kim, H.K. The antiobesity effect of Polygonum aviculare L. ethanolic extract in high-fat diet-induced obese mice. Evid. Based Complement. Altern. Med. 2013, 2013, 626397. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Kim, C.S.; Park, K.M.; Chang, P.S. Inhibitory characteristics of flavonol-3-O-glycosides from Polygonum aviculare L. (common knotgrass) against porcine pancreatic lipase. Sci. Rep. 2019, 9, 18080. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.F. Clinical observation on treatment of 25 cases of diabetes with Polygonum aviculare L. J. Nantong Univ. 1995, 15, 274–275. [Google Scholar]
- Chen, Y.; Lan, Y. Polygonum aviculare L. for diabetes. Hubei J. Tradit. Chin. Med. 2000, 22, 32. [Google Scholar]
- Yin, M.H.; Kang, D.G.; Choi, D.H.; Kwon, T.O.; Lee, H.S. Screening of vasorelaxant activity of some medicinal plants used in Oriental medicines. J. Ethnopharmacol. 2005, 99, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Liu, L.; Zhu, D.D. Textual research on Chinese herbal medicine and modern pharmacology of Polygonum aviculare L. J. Liaoning Univ. Tradit. Chin. Med. 2020, 22, 4–7. [Google Scholar]
- Yang, J.L.; Huang, L.D.; Zhang, Y.Z.; Wu, J. Research advances of Polygonum aviculare L. Anhui Med. Pharm. J. 2016, 20, 1025–1029. [Google Scholar]
- Das, R.R.; Rahman, M.A.; Al-Araby, S.Q.; Islam, M.S.; Rashid, M.M.; Babteen, N.A.; Alnajeebi, A.M.; Alharbi, H.; Jeandet, P.; Rafi, M.; et al. The antioxidative role of natural compounds from a green coconut mesocarp undeniably contributes to control diabetic complications as evidenced by the associated genes and biochemical indexes. Oxid. Med. Cell. Longev. 2021, 2021, 9711176. [Google Scholar] [CrossRef]
- Tan, Y.; Chang, S.; Zhang, Y. Comparison of α-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.H.; Yu, J.S.; Lee, H.S.; Choi, C.I.; Kim, K.H. Antidiabetic flavonoids from fruits of Morus alba promoting insulin-stimulated glucose uptake via Akt and AMP-activated protein kinase activation in 3T3-L1 adipocytes. Pharmaceutics 2021, 13, 526. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Vasconcelos, A.; Wu, D.T.; Li, H.B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.Y. Citrus flavonoids as promising phytochemicals targeting diabetes and related complications: A systematic review of in vitro and in vivo studies. Nutrients 2020, 12, 2907. [Google Scholar] [CrossRef]
- Leite, D.; Camilo, C.J.; Nonato, C.; Carvalho, N.; Salazar, G.; de Morais, S.M.; Costa, J. Chemical profile and evaluation of the antioxidant and anti-acetylcholinesterase activities of Annona squamosa L. (Annonaceae) extracts. Foods 2021, 10, 2343. [Google Scholar] [CrossRef]
- Azemi, A.K.; Mokhtar, S.S.; Sharif, S.; Rasool, A. Clinacanthus nutans attenuates atherosclerosis progression in rats with type 2 diabetes by reducing vascular oxidative stress and inflammation. Pharm. Biol. 2021, 59, 1432–1440. [Google Scholar] [CrossRef]
- Leong, X.F. Lipid oxidation products on inflammation-mediated hypertension and atherosclerosis: A mini review. Front. Nutr. 2021, 8, 717740. [Google Scholar] [CrossRef]
- Piera-Velazquez, S.; Jimenez, S.A. Oxidative stress induced by reactive oxygen species (ROS) and NADPH oxidase 4 (NOX4) in the pathogenesis of the fibrotic process in systemic sclerosis: A promising therapeutic target. J. Clin. Med. 2021, 10, 4791. [Google Scholar] [CrossRef] [PubMed]
- Rosini, E.; Pollegioni, L. Reactive oxygen species as a double-edged sword: The role of oxidative enzymes in antitumor therapy. Biofactors, 2021; in press. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.T.; Zhou, H.; Liu, Y.M.; Lin, A.H. Inhibitory effect of phellodendrine on α-glucosidase in vitro. J. Nanjing. Univ. Tradit. Chin. Med. 2020, 36, 853–858. [Google Scholar]
- Lim, B.O. Effects of wogonin, wogonoside, and 3,5,7,2’,6’-pentahydroxyflavone on chemical mediator production in peritoneal exduate cells and immunoglobulin E of rat mesenteric lymph node lymphocytes. J. Ethnopharmacol. 2003, 84, 23–29. [Google Scholar] [CrossRef]
- Gök, H.N.; Orhan, N.; Özüpek, B.; Pekacar, S.; Selvi, Ş.N.; Orhan, D.D. Standardization of Juniperus macrocarpa Sibt. & Sm. and Juniperus excelsa M. Bieb. Extracts with Carbohydrate Digestive Enzyme Inhibitory and Antioxidant Activities. Iran. J. Pharm. Res. 2021, 20, 441–455. [Google Scholar]
- Zhang, Z.; Lv, G.; Du, L. Avicularin Reduces the Expression of Mediators of Inflammation and Oxidative Stress in Bradykinin-Treated MG-63 Human Osteoblastic Osteosarcoma Cells. Med. Sci. Monit. 2020, 26, e921957. [Google Scholar] [CrossRef]
- Zhu, X.; Qiu, Z.; Ouyang, W.; Miao, J.; Xiong, P.; Mao, D.; Feng, K.; Li, M.; Luo, M.; Xiao, H.; et al. Hepatic transcriptome and proteome analyses provide new insights into the regulator mechanism of dietary avicularin in diabetic mice. Food. Res. Int. 2019, 125, 108570. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, R.; Shi, W.; Li, L.; Liu, H.; Liu, Z.; Wu, L. The Multifunctional Benefits of Naturally Occurring Delphinidin and Its Glycosides. J. Agric. Food Chem. 2019, 67, 11288–11306. [Google Scholar] [CrossRef]
- Promyos, N.; Temviriyanukul, P.; Suttisansanee, U. Investigation of Anthocyanidins and Anthocyanins for Targeting α-Glucosidase in Diabetes Mellitus. Prev. Nutr. Food. Sci. 2020, 25, 263–271. [Google Scholar] [CrossRef]
- Xiao, Z.; Yang, R.; Wang, H.; Cui, X.; Zhang, Y.; Yuan, Y.; Yue, T.; Li, P. Inhibitory properties of polyphenols in Malus "Winter Red" crabapple fruit on α-glucosidase and α-amylase using improved methods. J. Food. Biochem. 2021, 45, e13942. [Google Scholar] [CrossRef]
- Egbuna, C.; Awuchi, C.G.; Kushwaha, G.; Rudrapal, M.; Patrick-Iwuanyanwu, K.C.; Singh, O.; Odoh, U.E.; Khan, J.; Jeevanandam, J.; Kumarasamy, S.; et al. Bioactive Compounds Effective Against Type 2 Diabetes Mellitus: A Systematic Review. Curr. Top. Med. Chem. 2021, 21, 1067–1095. [Google Scholar] [CrossRef] [PubMed]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H.; de Mejia, E.G. Phenolic Compounds from Fermented Berry Beverages Modulated Gene and Protein Expression to Increase Insulin Secretion from Pancreatic β-Cells in Vitro. J. Agric. Food Chem. 2016, 64, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasam, B.; Vareed, S.K.; Olson, L.K.; Nair, M.G. Insulin secretion by bioactive anthocyanins and anthocyanidins present in fruits. J. Agric. Food Chem. 2005, 53, 28–31. [Google Scholar] [CrossRef]
- Huang, H.; Xin, H.; Liu, X.; Xu, Y.; Wen, D.; Zhang, Y.; Zhu, Y.Z. Novel anti-diabetic effect of SCM-198 via inhibiting the hepatic NF-κB pathway in db/db mice. Biosci. Rep. 2012, 32, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Ding, Z.; Zhang, K.; Duan, W.; Qin, Y.; Su, Z.; Bi, Y. Optimization of extraction flavonoids from Exocarpium Citri Grandis and evaluation its hypoglycemic and hypolipidemic activities. J. Ethnopharmacol. 2020, 262, 113178. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhao, J.G.; Zhang, Y.Q. The flavonoid-rich ethanolic extract from the green cocoon shell of silkworm has excellent antioxidation, glucosidase inhibition, and cell protective effects in vitro. Food. Nutr. Res. 2020, 64, 29219. [Google Scholar] [CrossRef]
- Fan, S.; Yang, G.; Zhang, J.; Li, J.; Bai, B. Optimization of ultrasound-assisted extraction using response surface methodology for simultaneous quantitation of six flavonoids in flos sophorae immaturus and antioxidant activity. Molecules 2020, 25, 1767. [Google Scholar] [CrossRef] [Green Version]
- Masood, S.; Rehman, A.U.; Ihsan, M.A.; Shahzad, K.; Sabir, M.; Alam, S.; Ahmed, W.; Shah, Z.H.; Alghabari, F.; Mehmood, A.; et al. Antioxidant potential and α-glucosidase inhibitory activity of onion (Allium cepa L.) peel and bulb extracts. Braz. J. Biol. 2021, 83, 00264. [Google Scholar] [CrossRef]
- Cheng, W.; Liu, L.; Zhu, D.D. Research on Chinese medicine Polygonum aviculare L. materia medica and modern pharmacology. J. Liaoning Univ. Tradit. Chin. Med. 2020, 22. [Google Scholar] [CrossRef]
- Mu, R.F.; Niu, Y.F.; Wang, Q.; Zhou, H.M.; Hu, J.; Qin, W.Y.; Xiong, W.Y. Eriocalyxin B inhibits adipogenesis in 3T3-L1 adipocytes by cell cycle arrest. Nat. Prod. Bioprospect. 2020, 10, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Huang, W.X.; Lü, H.M.; Qin, J.; Ke, B.; Shen, W.Z. Identification of Cald1 as a novel regulator of Linggui Zhugan decoction for improving insulin resistance in vivo and in vitro. J. Tradit. Chin. Med. 2021, 41, 706–716. [Google Scholar] [PubMed]
- Choi, E.; Baek, S.; Baek, K.; Kim, H.K. Psidium guajava L. leaf extract inhibits adipocyte differentiation and improves insulin sensitivity in 3T3-L1 cells. Nutr. Res. Pract. 2021, 15, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Hai, P.; Su, Y.L. Measuring of the total flavonoid content and anti-oxidation in polygonum aviculare and its processing products. Chin. J. Ethnomed. Ethnopharm. 2008, 17, 3–6. [Google Scholar]
- Shahwan, M.; Alhumaydhi, F.; Ashraf, G.M.; Hasan, P.; Shamsi, A. Role of polyphenols in combating Type 2 Diabetes and insulin resistance. Int. J. Biol. Macromol. 2022, 206, 567–579. [Google Scholar] [CrossRef]
- Wang, L.K.; Chen, H.G.; Huang, Z.L.; Huang, J.J.; Xia, Z.Y.; Zhao, S.Q.; Huang, H.R.; Zheng, J.X. Antioxidant activities and contents of total flavonoids and polyphenols from different parts of Millettia speciose Champ. Chin. Arch. Tradit. Chin. Med. 2022, 40, 139–142. [Google Scholar]
- Pradhan, B.; Patra, S.; Behera, C.; Nayak, R.; Jit, B.P.; Ragusa, A.; Jena, M. Preliminary investigation of the antioxidant, anti-diabetic, and anti-Inflammatory activity of enteromorpha intestinalis extracts. Molecules 2021, 26, 1171. [Google Scholar] [CrossRef]
- Banothu, V.; Neelagiri, C.; Adepally, U.; Lingam, J.; Bommareddy, K. Phytochemical screening and evaluation of in vitro antioxidant and antimicrobial activities of the indigenous medicinal plant Albizia odoratissima. Pharm. Biol. 2017, 55, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- Aloo, S.O.; Ofosu, F.K.; Oh, D.H. Effect of germination on alfalfa and buckwheat: Phytochemical profiling by UHPLC-ESI-QTOF-MS/MS, bioactive compounds, and in vitro studies of their diabetes and obesity-related functions. Antioxidants 2021, 10, 1613. [Google Scholar] [CrossRef]
- Mechchate, H.; Es-Safi, I.; Conte, R.; Hano, C.; Amaghnouje, A.; Jawhari, F.Z.; Radouane, N.; Bencheikh, N.; Grafov, A.; Bousta, D. In vivo and in vitro antidiabetic and anti-inflammatory properties of Flax (Linum usitatissimum L.) seed polyphenols. Nutrients 2021, 13, 2759. [Google Scholar] [CrossRef]
- Zhao, X.; Tao, J.; Zhang, T.; Jiang, S.; Wei, W.; Han, H.; Shao, Y.; Zhou, G.; Yue, H. Resveratroloside alleviates postprandial hyperglycemia in diabetic mice by competitively inhibiting α-glucosidase. J. Agric. Food Chem. 2019, 67, 2886–2893. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Addepalli, R.; Netzel, M.E.; Tinggi, U.; Fletcher, M.T.; Sultanbawa, Y.; Osborne, S.A. Antioxidant-rich extracts of terminalia ferdinandiana interfere with estimation of cell viability. Antioxidants 2019, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, A.; Singh, B.; Arora, R.; Arora, S. In vitro evaluation of the α-glucosidase inhibitory potential of methanolic extracts of traditionally used antidiabetic plants. BMC Complement. Altern. Med. 2019, 19, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Saburi, W.; Gai, Z.; Kato, K.; Ojima-Kato, T.; Yu, J.; Komoda, K.; Kido, Y.; Matsui, H.; Mori, H.; et al. Structural analysis of the α-glucosidase HaG provides new insights into substrate specificity and catalytic mechanism. Acta. Crystallogr. D Biol. Crystallogr. 2015, 71, 1382–1391. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and AutoDockTools4: Automated docking with selective receptor flexiblity. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsamo, D.; Tamokou, J.D.; Kengne, I.C.; Ngnokam, C.; Djamalladine, M.D.; Voutquenne-Nazabadioko, L.; Ngnokam, D. Antimicrobial and antioxidant secondary metabolites from Trifolium baccarinii Chiov. (fabaceae) and their mechanisms of antibacterial action. Biomed. Res. Int. 2021, 2021, 3099428. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | TFC (mg/g) | TPC (mg/g) |
|---|---|---|
| PAHEE | 68.7 ± 0.3 C | 43.2 ± 0.6 C |
| PEF | 88.5 ± 4.4 B | 62.2 ± 1.1 B |
| EAF | 159.7 ± 2.5 A | 107.6 ± 2.0 A |
| NBF | 34.7 ± 2.0 D | 29.7 ± 0.4 D |
| Category | Compound Name | Formula | Mass (g/mol) | Reference Mass (g/mol) | RT (min) | Area (μV·s) | Score (%) | Binding Energy (kcal/mol) | References |
|---|---|---|---|---|---|---|---|---|---|
| Flavonoids | 3,5,7,2′,6′-Pentahydroxyflavone | C15H10O7 | 302.0 | 302.0 | 22.7 | 100671889 | 99.4 | −9.55 | [25] a |
| 5,7,2′,3′-Tetrahydroxyflavone | C15H10O6 | 286.0 | 286.1 | 27.0 | 65396294 | 96.3 | −7.15 | * | |
| 3′-Methoxydaidzein | C16H12O5 | 284.1 | 284.1 | 48.2 | 43163766 | 97.0 | −8.80 | * | |
| Agathisflavone | C30H18O10 | 538.1 | 538.1 | 42.0 | 39711957 | 99.5 | −11.35 | [26] a,b | |
| 6-Hydroxykaempferol-7-O-glucoside | C21H20O12 | 464.1 | 464.1 | 17.9 | 67100153 | 98.9 | −8.08 | * | |
| Avicularin | C20H18O11 | 434.1 | 434.1 | 22.7 | 57090941 | 99.0 | −10.02 | [27] a; [28] b | |
| Desmanthin 2 | C28H24O16 | 616.1 | 616.1 | 16.5 | 29675775 | 98.1 | −8.45 | * | |
| 3,5,7,3′,4′ -Pentahydroxyflavone-3-L-rhamnoside | C21H20O11 | 448.1 | 448.1 | 24.1 | 26798354 | 96.8 | −10.20 | * | |
| 3,7,4′,5-Tetrahydroxyflavone-3-L-rhamnoside | C21H20O10 | 432.1 | 432.1 | 28.5 | 15851984 | 98.8 | −8.16 | * | |
| Melicitrin | C20H18O12 | 450.1 | 450.1 | 17.6 | 11126608 | 98.9 | −6.79 | * | |
| 2′′-O-Galloylisoorientin | C28H24O15 | 600.1 | 600.1 | 30.5 | 10835996 | 97.8 | −9.78 | * | |
| Delphinidin | C15H11O7 | 303.1 | 303.1 | 28.7 | 47200508 | 99.5 | −7.50 | [29] a,b; [30] b | |
| Cyanidin | C15H11O6 | 287.1 | 287.1 | 34.4 | 32042323 | 99.2 | −8.03 | [31] b; [32] b | |
| Pelargonidin | C15H11O5 | 271.1 | 271.1 | 35.5 | 27253529 | 97.6 | −8.43 | [33] b | |
| Peonidin | C16H13O6 | 301.1 | 301.1 | 36.7 | 14049096 | 99.3 | −6.65 | * | |
| Delphinidin-3-O-β-D-galactoside | C21H21O12 | 465.1 | 465.1 | 17.9 | 85247661 | 98.9 | −11.58 | * | |
| Delphinidin-3-arabinoside | C20H19O11 | 435.1 | 435.1 | 22.7 | 70570638 | 99.0 | −8.52 | [34] b | |
| Pelargonidin-3-galactoside | C21H21O10 | 433.1 | 433.1 | 28.5 | 19201678 | 99.2 | / | [35] b | |
| Delphinidin-3′-O-(2″-O-galloyl-β-galactoside) | C28H25O16 | 617.1 | 617.1 | 17.3 | 13797523 | 98.2 | −6.29 | -- | |
| Phenolics | 2-Methoxy-4-(3-methoxy-1-propenyl)-phenol | C11H14O3 | 194.1 | 194.1 | 11.73 | 16988535 | 97.9 | −5.14 | * |
| Alkaloids | Leonurine | C14H21N3O5 | 311.2 | 311.2 | 26.7 | 27644760 | 98.6 | / | [36] b |
| Gnoscopine | C24H27NO6 | 425.2 | 425.2 | 46.5 | 24023457 | 97.1 | / | -- | |
| 8-Acetyldolaconine | C26H39NO6 | 461.3 | 461.3 | 31.8 | 18923730 | 98.6 | / | -- | |
| Quinones | 1,8-Dihydroxy-4-hydroxymethyl anthraquinone | C15H10O5 | 270.1 | 270.1 | 35.5 | 23901981 | 97.6 | / | -- |
| 1,6-Dihydroxy-2,4-dimethoxyanthraquinone V | C16H12O6 | 300.1 | 300.1 | 36.7 | 12128297 | 99.3 | / | -- | |
| Abieta-8,12-dien-11,14-dione | C20H28O2 | 300.2 | 300.2 | 37.8 | 11252041 | 98.6 | / | -- | |
| Terpenoids | (12R)-12-Hydroxy cascarill one | C20H30O3 | 318.2 | 318.2 | 43.2 | 12287901 | 96.6 | / | -- |
| Samples | DPPH (SC50/μg/mL) | ABTS (mmol Trolox/L) | FRAP (mmol Trolox/L) |
|---|---|---|---|
| PAHEE | 47.64 ± 3.72 B | 0.83 ± 0.03 C | 0.35 ± 0.00 C |
| PEF | 31.13 ± 2.23 C | 1.12 ± 0.01 B | 0.54 ± 0.00 B |
| EAF | 19.94 ± 1.37 D | 1.30 ± 0.00 A | 0.76 ± 0.04 A |
| NBF | 96.62 ± 5.87 A | 0.42 ± 0.12 D | 0.14 ± 0.00 D |
| Ascorbic acid | 2.44 ± 0.33 E | -- | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Han, M.; Zhao, X.; Chen, X.; Wang, H.; Ni, J.; Zhang, Y. Hypoglycemic and Antioxidant Properties of Extracts and Fractions from Polygoni Avicularis Herba. Molecules 2022, 27, 3381. https://doi.org/10.3390/molecules27113381
Zhang K, Han M, Zhao X, Chen X, Wang H, Ni J, Zhang Y. Hypoglycemic and Antioxidant Properties of Extracts and Fractions from Polygoni Avicularis Herba. Molecules. 2022; 27(11):3381. https://doi.org/10.3390/molecules27113381
Chicago/Turabian StyleZhang, Kun, Mei Han, Xia Zhao, Xuelin Chen, Hanlei Wang, Jiyan Ni, and Yumei Zhang. 2022. "Hypoglycemic and Antioxidant Properties of Extracts and Fractions from Polygoni Avicularis Herba" Molecules 27, no. 11: 3381. https://doi.org/10.3390/molecules27113381
APA StyleZhang, K., Han, M., Zhao, X., Chen, X., Wang, H., Ni, J., & Zhang, Y. (2022). Hypoglycemic and Antioxidant Properties of Extracts and Fractions from Polygoni Avicularis Herba. Molecules, 27(11), 3381. https://doi.org/10.3390/molecules27113381