
In Vitro and In Vivo Protective Effects of Lentil (Lens culinaris) Extract against Oxidative Stress-Induced Hepatotoxicity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
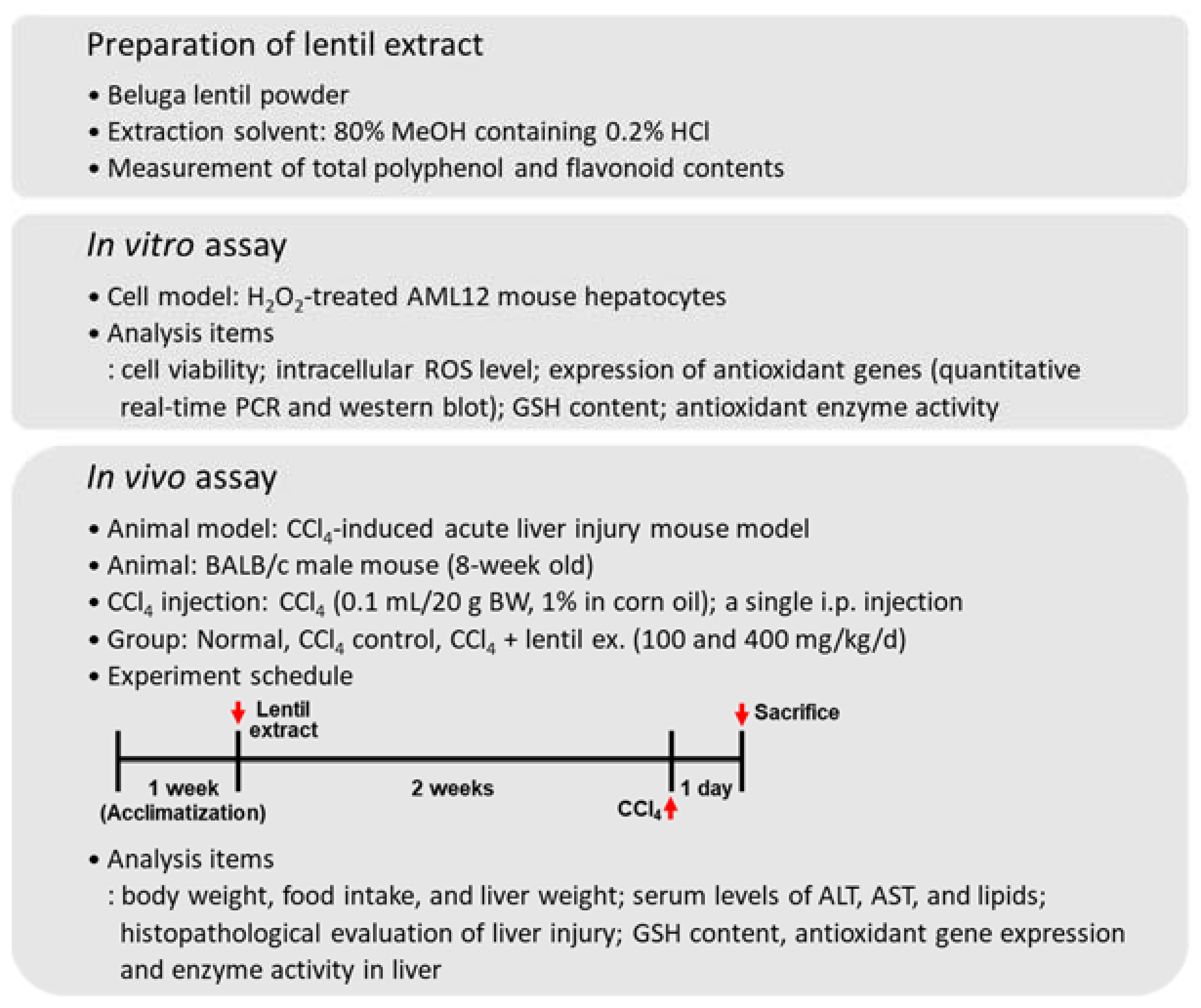
2.1. Materials and Preparation of Lentil Extract
2.2. Cell Cultures
2.3. Animals and Experimental Design
2.4. Statistical Analysis
3. Results and Discussion
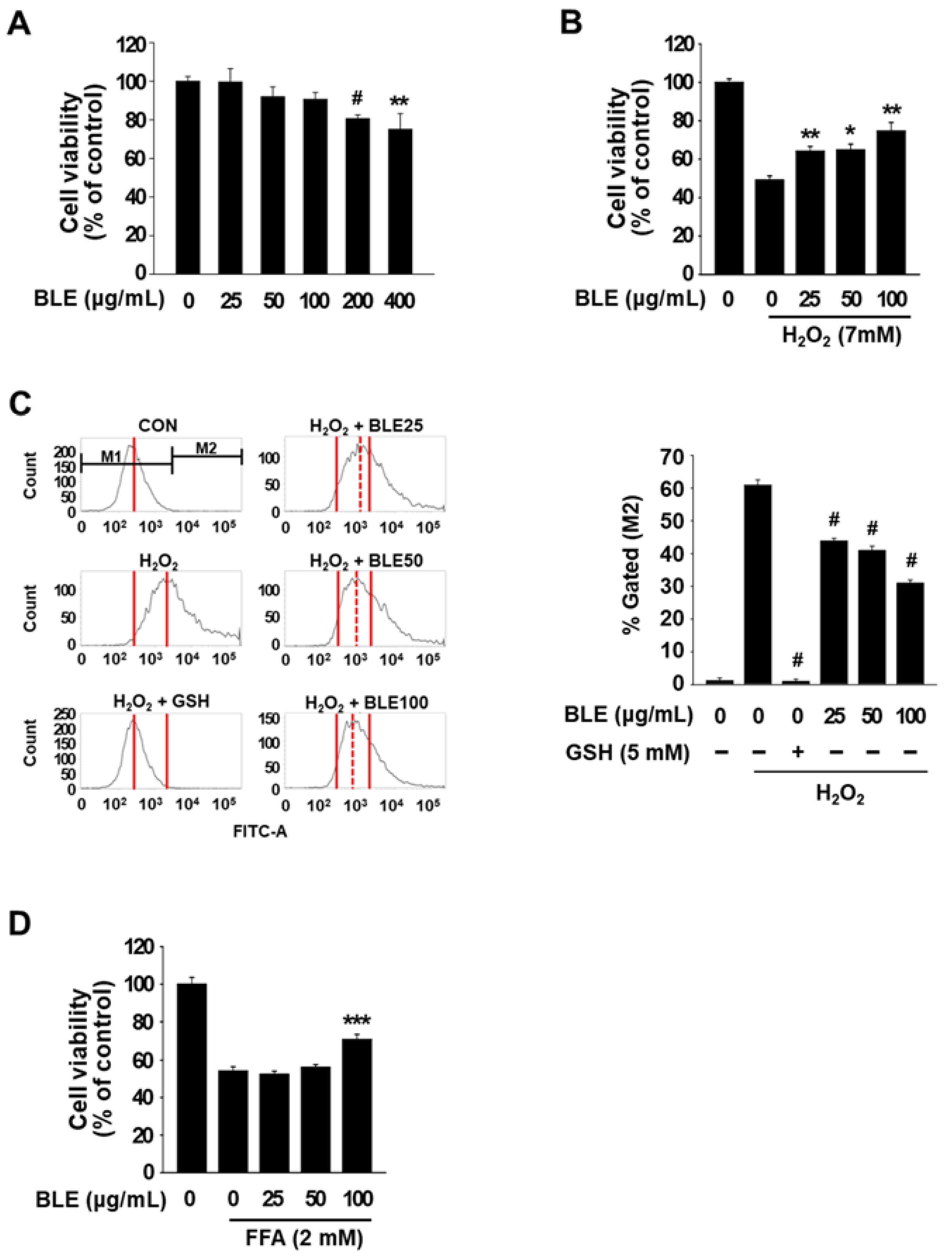
3.1. BLE Protects AML12 Cells against Oxidative Stress-Induced Cytotoxicity
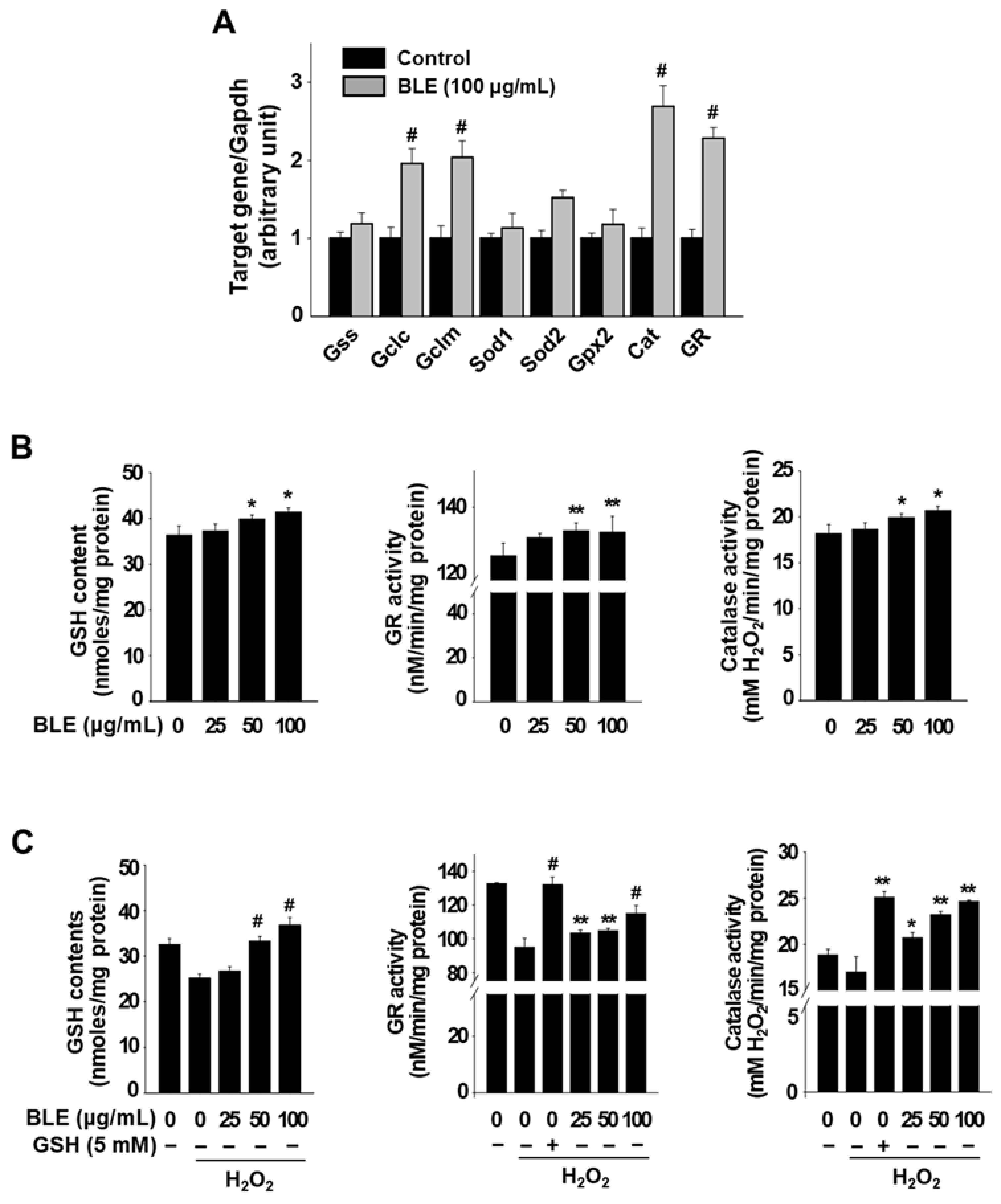
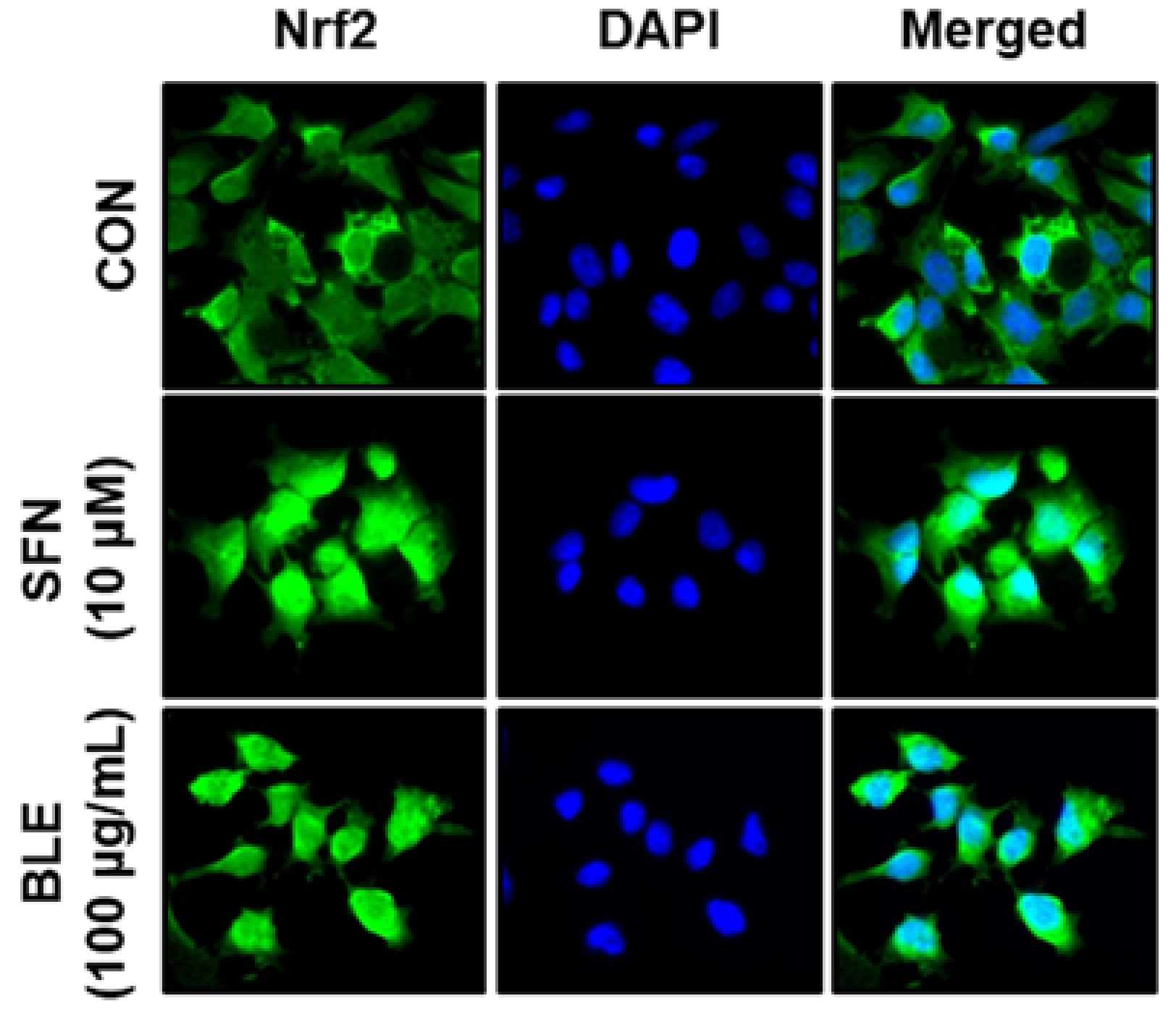
3.2. BLE Upregulates the Expression of Antioxidant Genes, in Part, via the Activation of Nrf2 in AML12 Cells
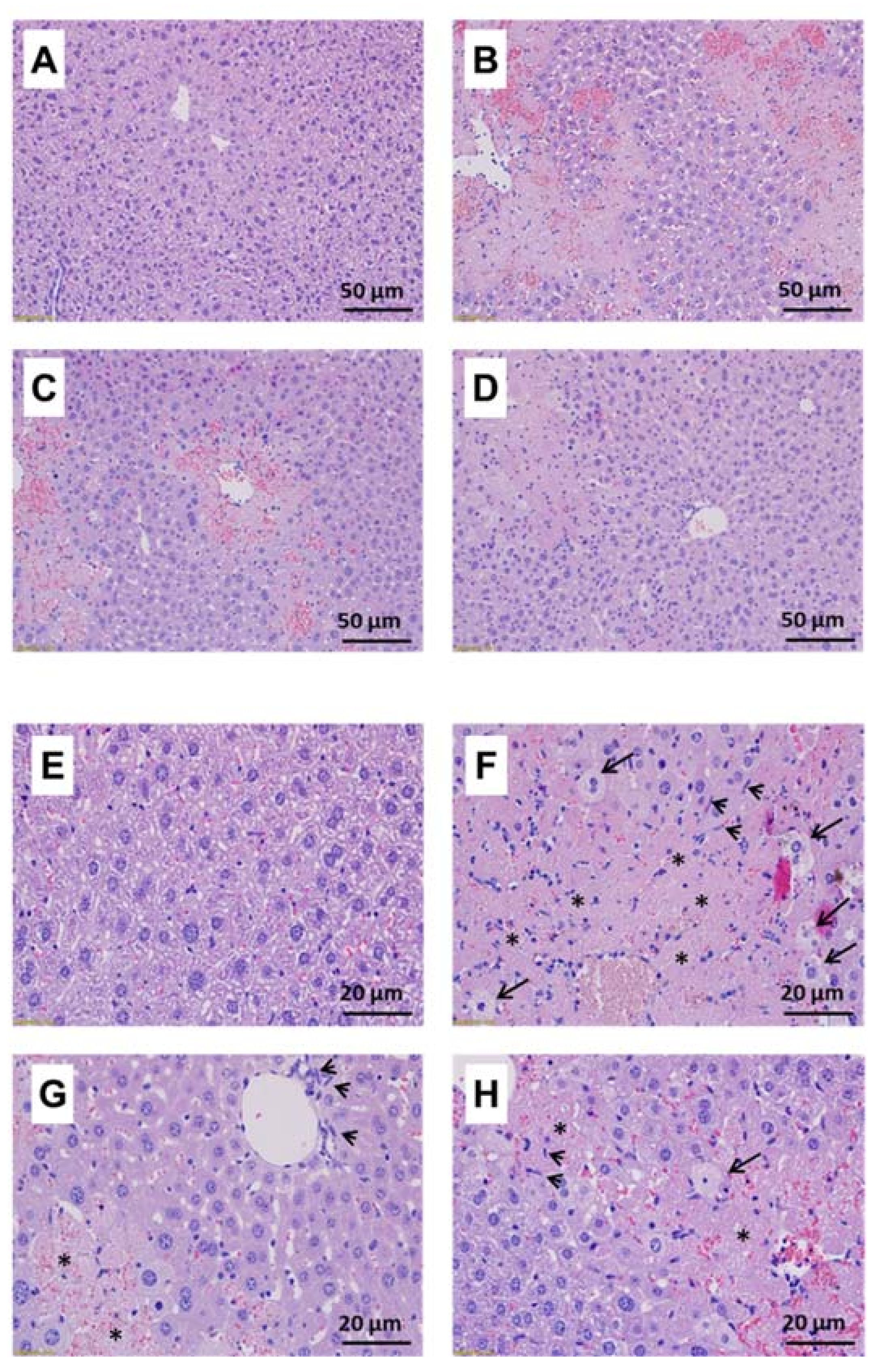
3.3. Protective Effects of BLE against CCl4-Induced Acute Hepatotoxicity in BALB/c Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Sample Availability
References
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [Green Version]
- Adewusi, E.A.; Afolayan, A. A review of natural products with hepatoprotective activity. J. Med. Plants Res. 2010, 4, 1318–1334. [Google Scholar]
- Li, Y.; Hu, D.; Qi, J.; Cui, S.; Chen, W. Lysosomal reacidification ameliorates vinyl carbamate-induced toxicity and disruption on lyso-somal pH. J. Agric. Food Chem. 2020, 68, 8951–8961. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-Y.; Yun, D.-H.; Lee, J.-H.; Kwon, Y.-B.; Lee, Y.-M.; Lee, D.-H.; Kim, D.-K. Extract of Triticum aestivum Sprouts Suppresses Acetaminophen-Induced Hepatotoxicity in Mice by Inhibiting Oxidative Stress. Molecules 2021, 26, 6336. [Google Scholar] [CrossRef]
- Liu, J.; Li, D.; Zhang, T.; Tong, Q.; Ye, R.D.; Lin, L. SIRT3 protects hepatocytes from oxidative injury by enhancing ROS scavenging and mito-chondrial integrity. Cell Death Dis. 2017, 8, e3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H. Polyphenol Contents and Antioxidant Activities of Lentil Extracts from Different Cultivars. J. Korean Soc. Food Sci. Nutr. 2016, 45, 973–979. [Google Scholar] [CrossRef]
- Yang, L.; Wei, J.; Sheng, F.; Li, P. Attenuation of Palmitic Acid–Induced Lipotoxicity by Chlorogenic Acid through Activation of SIRT1 in Hepatocytes. Mol. Nutr. Food Res. 2019, 63, e1801432. [Google Scholar] [CrossRef] [PubMed]
- Köroğlu, E.; Canbakan, B.; Atay, K.; Hatemi, I.; Tuncer, M.; Dobrucalı, A.; Sonsuz, A.; Gültepe, I.; Şentürk, H. Role of oxidative stress in the pathogenesis of non-alcoholic fatty liver dis-ease: Implications for prevention and therapy. Antioxidants 2021, 10, 174. [Google Scholar]
- Ganesan, K.; Xu, B. Polyphenol-Rich Lentils and Their Health Promoting Effects. Int. J. Mol. Sci. 2017, 18, 2390. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Wang, Y.; Zhao, E.; Wu, K.; Li, W.; Shi, L.; Wang, D.; Xie, G.; Yin, Y.; Deng, M.; et al. Maresin 1, a Proresolving Lipid Mediator, Mitigates Carbon Tetrachloride-Induced Liver Injury in Mice. Oxid. Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Yan, C.; Sun, W.; Wang, X.; Long, J.; Liu, X.; Feng, Z.; Liu, J. Punicalagin attenuates palmitate-induced lipotoxicity in HepG2 cells by activating the Keap1-Nrf2 antioxidant defense system. Mol. Nutr. Food Res. 2016, 60, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- López, A.; El-Naggar, T.; Dueñas, M.; Ortega, T.; Estrella, I.; Hernández, T.; Carretero, M.E. Influence of processing in the phenolic composition and health-promoting properties of lintils (Lens culinaris L.). J. Food Process. Preserv. 2017, 41, e13113. [Google Scholar] [CrossRef]
- Yao, F.-R.; Sun, C.; Chang, S.K.C. Morton Lentil Extract Attenuated Angiotensin II-Induced Cardiomyocyte Hypertrophy via Inhibition of Intracellular Reactive Oxygen Species Levels in Vitro. J. Agric. Food Chem. 2010, 58, 10382–10388. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K.C. Phenolic Substance Characterization and Chemical and Cell-Based Antioxidant Activities of 11 Lentils Grown in the Northern United States. J. Agric. Food Chem. 2010, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W. Antioxidant activity of ethanol and water extracts from lentil (Lens culinaris). J. Food Nutr. Res. 2015, 3, 667–669. [Google Scholar]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1–Nrf2 system. Free. Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Scholten, D.; Trebicka, J.; Liedtke, C.; Weiskirchen, R. The carbon tetrachloride model in mice. Lab. Anim. 2015, 49, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Apte, U. Liver Regeneration: Basic Mechanisms, Relevant Models and Clinical Applications, 1st ed.; Academic Press: San Diego, CA, USA, 2015; Chapter 1. [Google Scholar]
- Kalantari, H.; Houshmand, G.; Hasanvand, A.; Kalantar, M.; Goudarzi, M.; Haghighian, H.K. Ameliorative Effects of Red Lentil Extract on Sodium Arsenite-induced Oxidative Stress in Rats. Jundishapur J. Nat. Pharm. Prod. 2017, 12 (Supp. 3), e64309. [Google Scholar] [CrossRef]
- Abdel-Salam, N.A.; Ghazy, N.M.; Sallam, S.M.; Radwan, M.; Wanas, A.; ElSohly, M.A.; El-Demellawy, M.A.; Abdel-Rahman, N.M.; Piacente, S.; Shenouda, M.L. Flavonoids of Alcea rosea L. and their immune stimulant, antioxidant and cytotoxic activities on hepatocellular carcinoma HepG-2 cell line. Nat. Prod. Res. 2017, 32, 702–706. [Google Scholar] [CrossRef]
- Binu, P.; Gifty, K.; Vineetha, R.C.; Abhilash, S.; Arathi, P.; Nair, R.H. Eugenol, a plant-derived phenolic nutraceutical, protects thiol (SH) group in myo-cardium from ROS-mediated oxidation under chemotherapeutic stress induced by arsenic trioxide—A in vivo model study. Drug Chem. Toxicol. 2018, 26, 1–6. [Google Scholar]
- Shahwar, D.; Bhat, T.M.; Ansari, M.Y.K.; Chaudhary, S.; Aslam, R. Retracted Article: Health functional compounds of lentil (Lens culinaris Medik): A review. Int. J. Food Prop. 2017, 20, S1–S15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antiox-idant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Takeoka, G.R.; Dao, L.T.; Tamura, H.; Harden, L.A. Delphinidin 3-O-(2-O-β-d-Glucopyranosyl-α-l-arabinopyranoside): A Novel Anthocyanin Identified in Beluga Black Lentils. J. Agric. Food Chem. 2005, 53, 4932–4937. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Polyphenols (mg GAE (1)/g Extract) | Total Flavonoids (mg QE (2)/g Extract) | |
|---|---|---|
| BLE | 32.09 ± 0.37 | 19.32 ± 0.08 |
| Normal | CCl4 | BLE100 | BLE400 | |
|---|---|---|---|---|
| ALT (U/L) | 73.15 ± 30.80 c | 9472.38 ± 2755.53 a | 10,420.50 ± 5724.78 a | 4860.90 ± 2041.11 b |
| AST (U/L) | 493.63 ± 334.75 b | 5158.45 ± 2333.14 a | 6829.04 ± 2995.22 a | 6010.76 ± 1140.44 a |
| T-Chol (mg/dL) | 141.50 ± 19.07 ns | 130.50 ± 20.13 | 132.43 ± 42.26 | 116.43 ± 20.28 |
| TG (mg/dL) | 169.50 ± 51.99 ab | 245.63 ± 81.19 a | 145.29 ± 81.40 b | 148.43 ± 46.97 b |
| HDL-C (mg/dL) | 105.73 ± 14.15 ns | 89.73 ± 12.15 | 99.43 ± 26.66 | 76.39 ± 30.46 |
| LDL-C (mg/dL) | 8.85 ± 1.78 ns | 11.05 ± 4.04 | 10.36 ± 3.52 | 17.93 ± 15.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, Y.-S.; Lee, S.-H.; Chun, S.Y.; Kim, D.H.; Jang, B.I.; Han, M.-H.; Lee, S.-O. In Vitro and In Vivo Protective Effects of Lentil (Lens culinaris) Extract against Oxidative Stress-Induced Hepatotoxicity. Molecules 2022, 27, 59. https://doi.org/10.3390/molecules27010059
Jung Y-S, Lee S-H, Chun SY, Kim DH, Jang BI, Han M-H, Lee S-O. In Vitro and In Vivo Protective Effects of Lentil (Lens culinaris) Extract against Oxidative Stress-Induced Hepatotoxicity. Molecules. 2022; 27(1):59. https://doi.org/10.3390/molecules27010059
Chicago/Turabian StyleJung, Yeon-Seop, So-Hee Lee, So Young Chun, Dae Hwan Kim, Byung Ik Jang, Man-Hoon Han, and Syng-Ook Lee. 2022. "In Vitro and In Vivo Protective Effects of Lentil (Lens culinaris) Extract against Oxidative Stress-Induced Hepatotoxicity" Molecules 27, no. 1: 59. https://doi.org/10.3390/molecules27010059
APA StyleJung, Y.-S., Lee, S.-H., Chun, S. Y., Kim, D. H., Jang, B. I., Han, M.-H., & Lee, S.-O. (2022). In Vitro and In Vivo Protective Effects of Lentil (Lens culinaris) Extract against Oxidative Stress-Induced Hepatotoxicity. Molecules, 27(1), 59. https://doi.org/10.3390/molecules27010059