
Direct and Indirect Antioxidant Effects of Selected Plant Phenolics in Cell-Based Assays



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
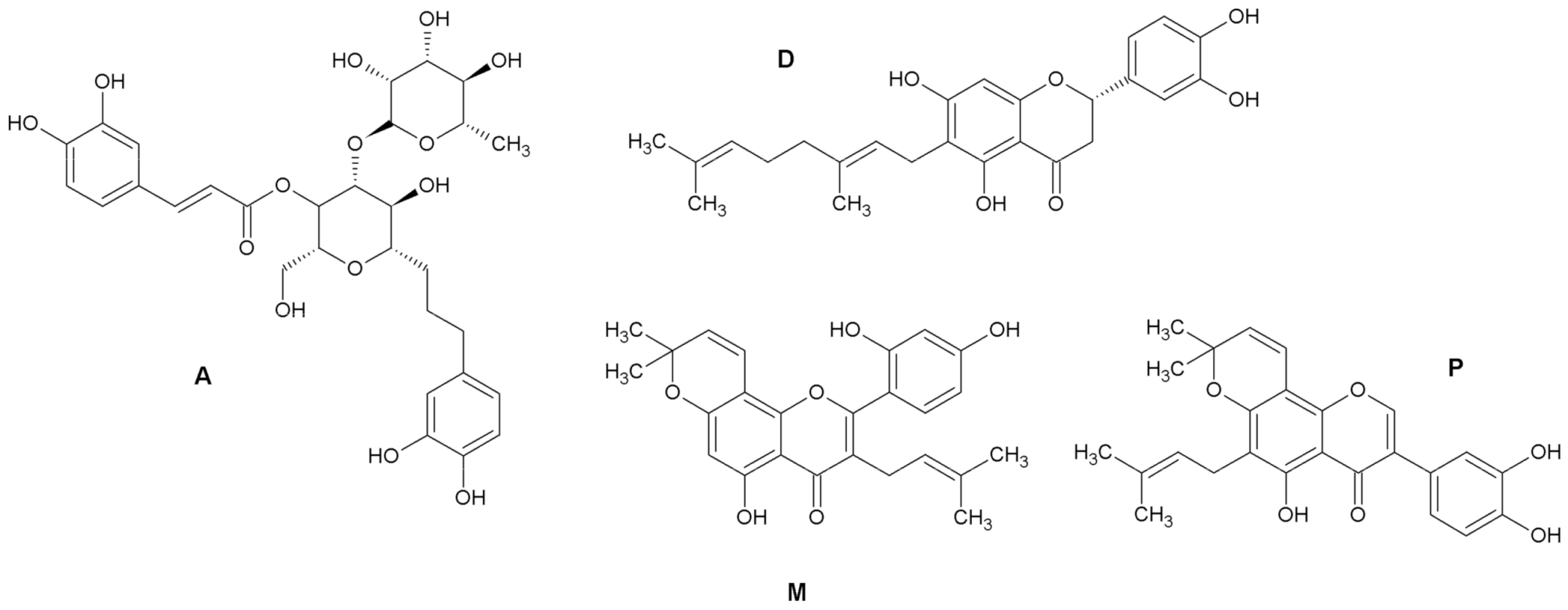
2.1. Antiproliferative Activity
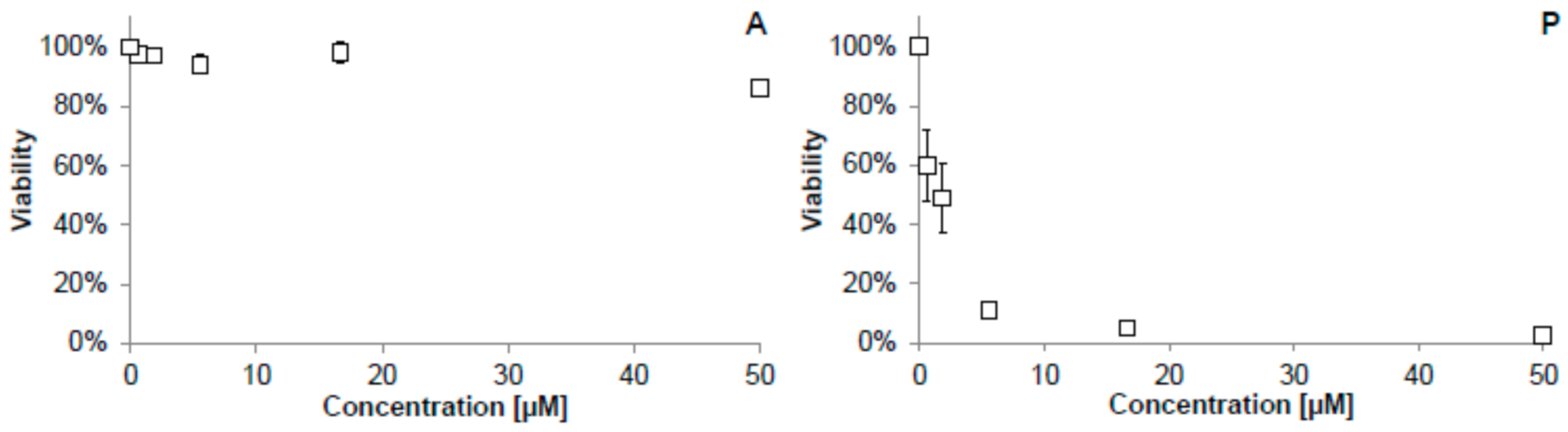
2.2. Cellular Antioxidant Activity (CAA) Assay
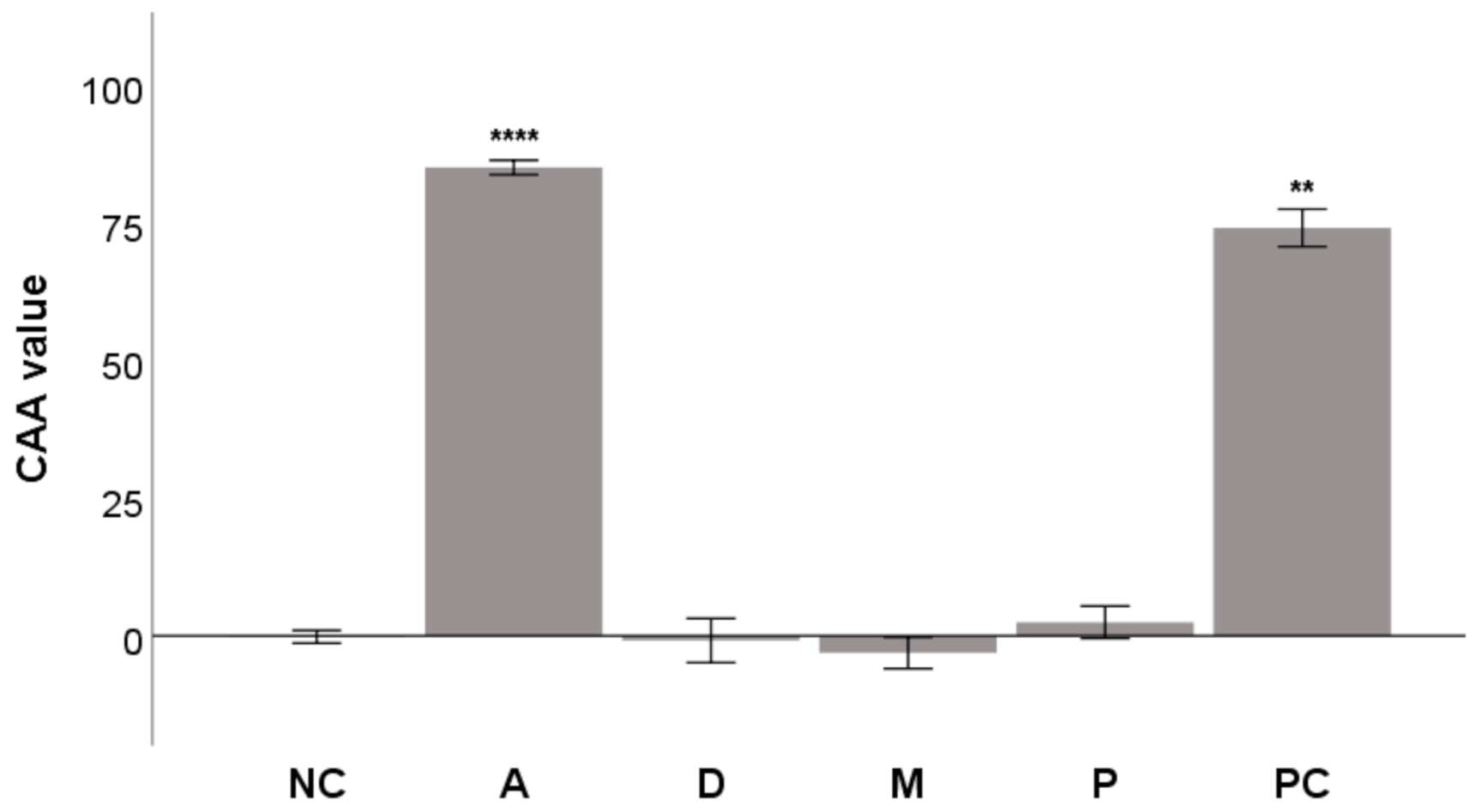
2.3. Glucose Oxidative Stress Protection (GOSP) Assay
2.4. Indirect Antioxidant Activity—Modulation of Antioxidant Enzymes
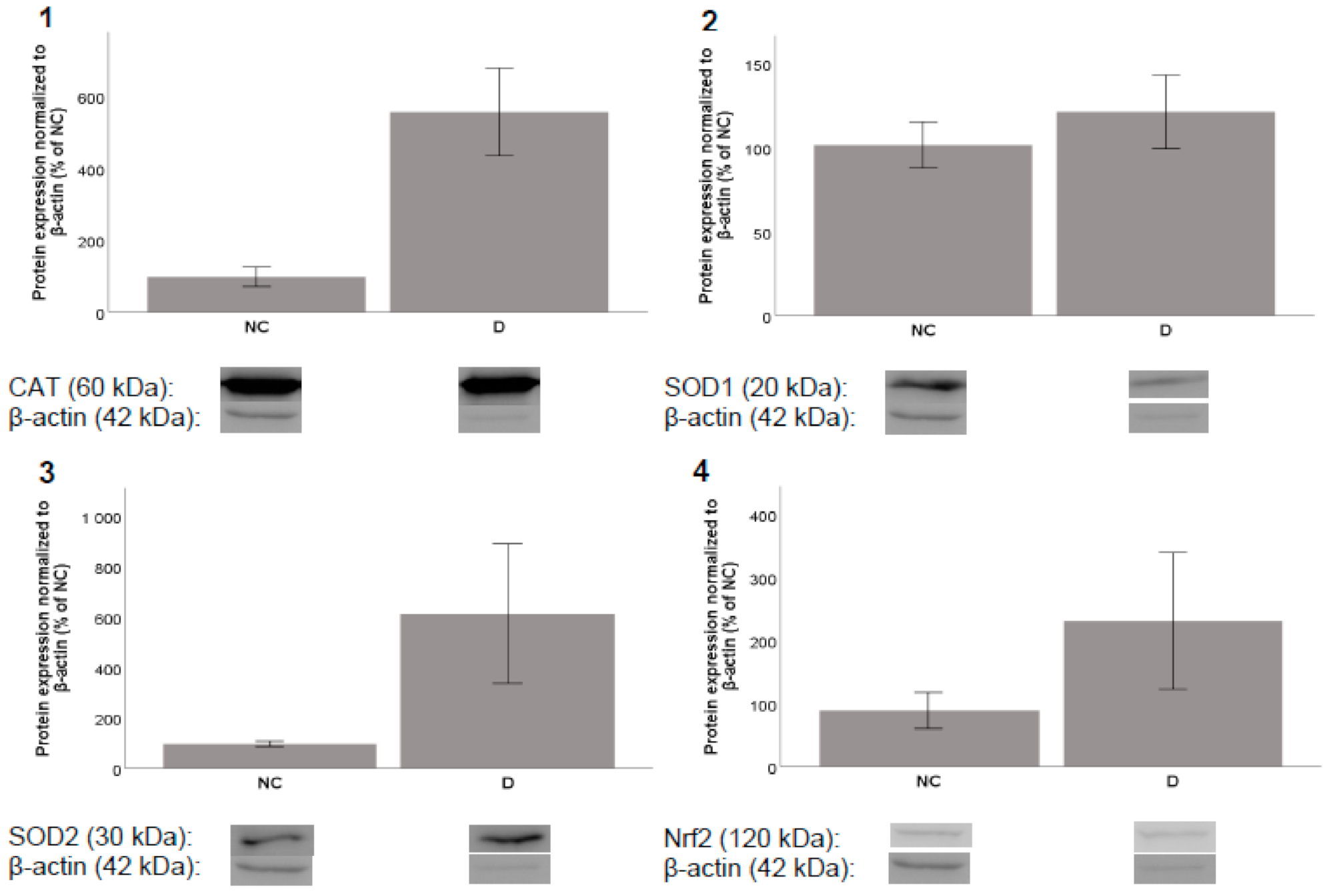
2.4.1. Protein Expression
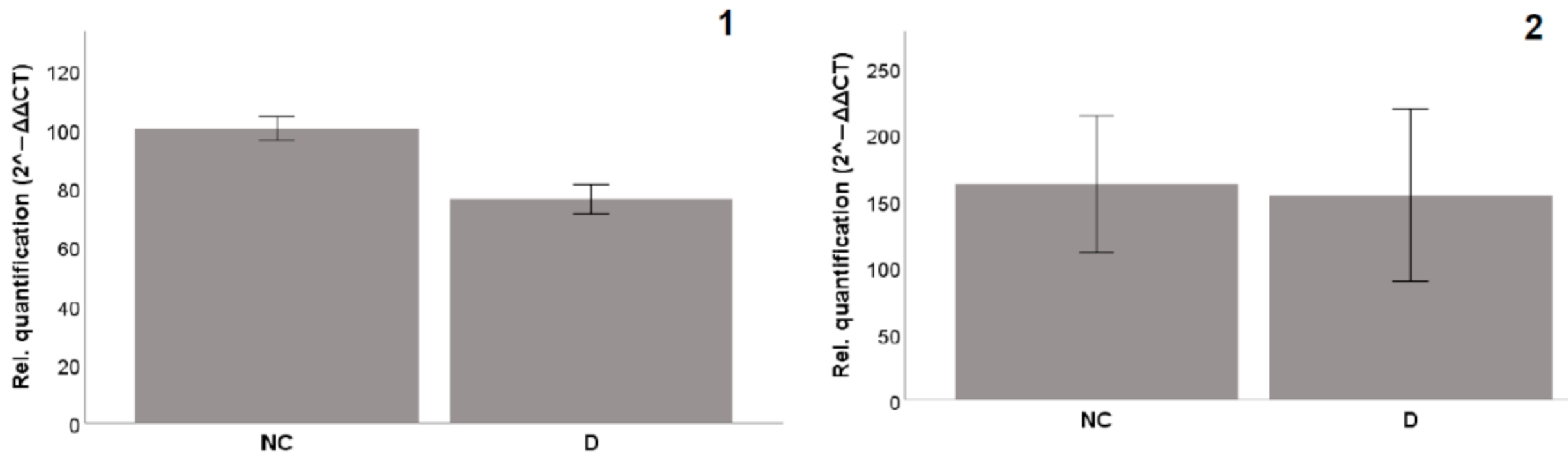
2.4.2. Gene Expression
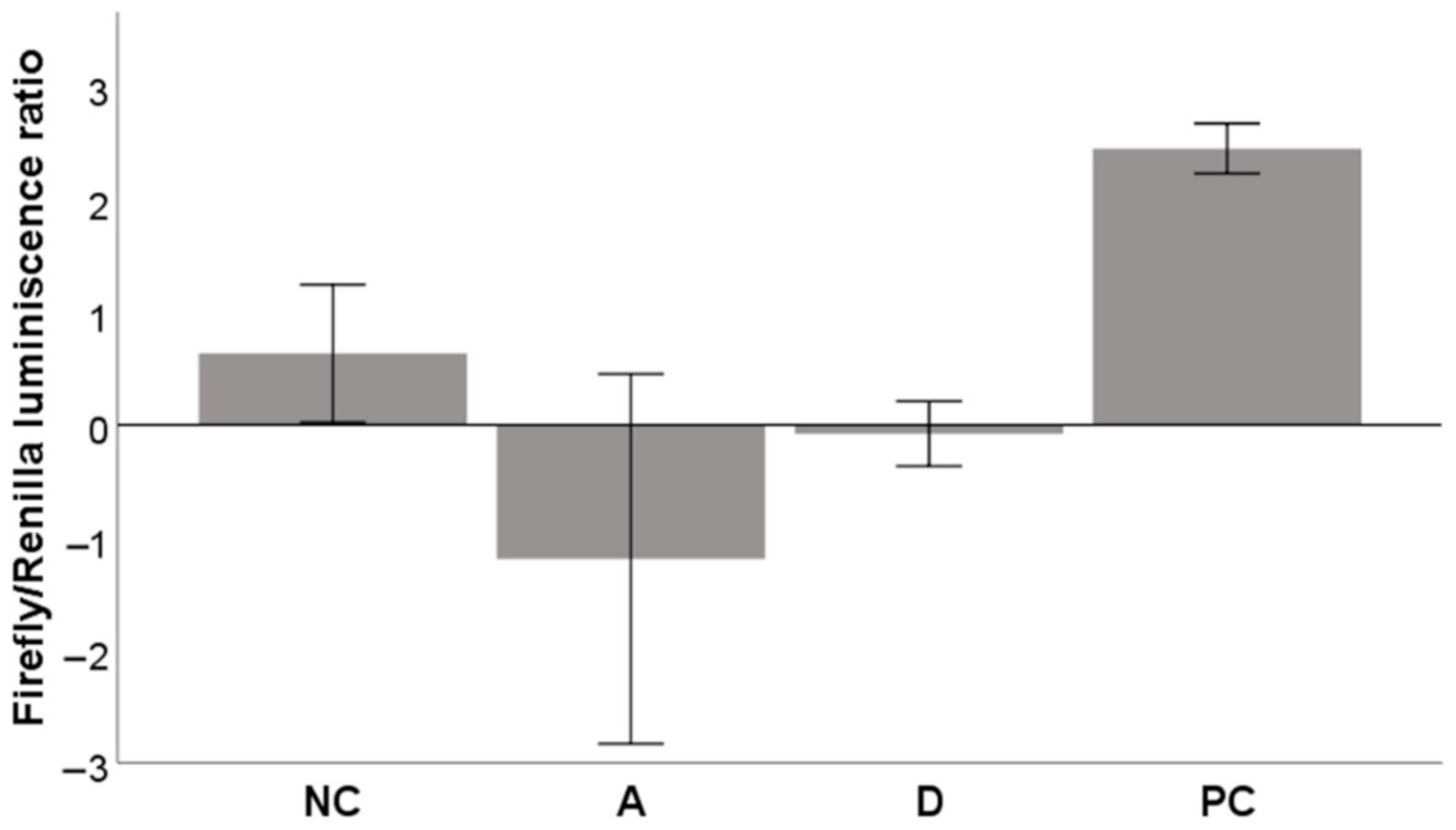
2.4.3. Activation of the Nrf2-ARE System
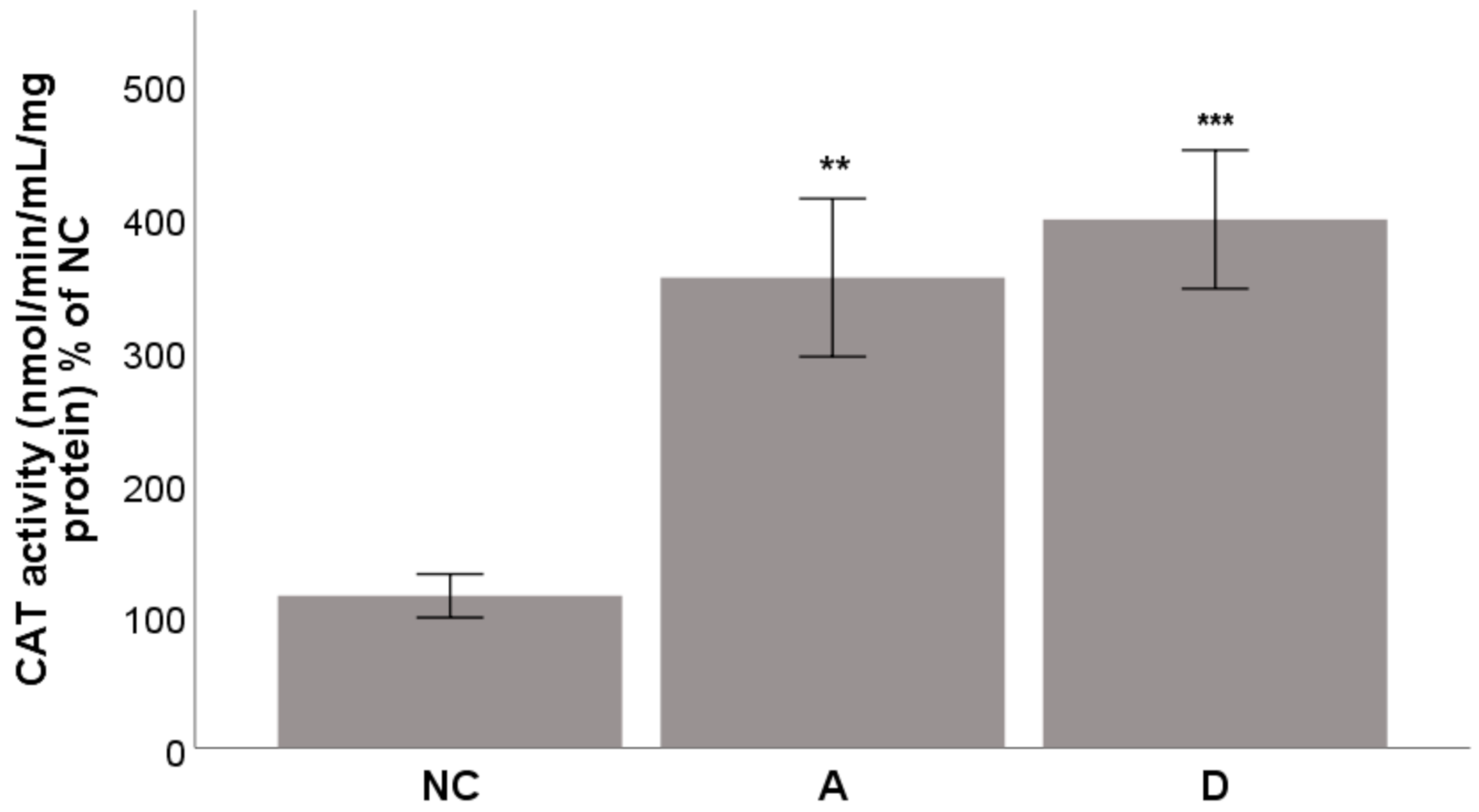
2.4.4. Activity of the Enzyme CAT
3. Materials and Methods
3.1. Test Compounds
3.2. Maintenance and Cultivation of the Cell Lines
3.3. Antiproliferative Activity
3.4. Cellular Antioxidant Activity (CAA) Assay
3.5. Glucose Oxidative Stress Protection (GOSP) Assay
3.6. Indirect Antioxidant Activity—Modulation of Antioxidant Enzymes
3.6.1. Protein Expression
3.6.2. Gene Expression
3.6.3. Activation of the Nrf2-ARE System
3.6.4. Activity of the Enzyme CAT
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Niki, E. Oxidative Stress and Antioxidants: Distress or Eustress? Arch. Biochem. Biophys. 2016, 595, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Treml, J.; Šmejkal, K. Flavonoids as Potent Scavengers of Hydroxyl Radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen Peroxide as a Central Redox Signaling Molecule in Physiological Oxidative Stress: Oxidative Eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- López-Alarcón, C.; Denicola, A. Evaluating the Antioxidant Capacity of Natural Products: A Review on Chemical and Cellular-Based Assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.-A.; Kwak, M.-K. The Nrf2 System as a Potential Target for the Development of Indirect Antioxidants. Molecules 2010, 15, 7266–7291. [Google Scholar] [CrossRef] [PubMed]
- Reisman, S.A.; Yeager, R.L.; Yamamoto, M.; Klaassen, C.D. Increased Nrf2 Activation in Livers from Keap1-Knockdown Mice Increases Expression of Cytoprotective Genes That Detoxify Electrophiles More than Those That Detoxify Reactive Oxygen Species. Toxicol. Sci. 2009, 108, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Chen, C.; Yu, R.; Owuor, E.D.; Kong, A.N. Activation of Antioxidant-Response Element (ARE), Mitogen-Activated Protein Kinases (MAPKs) and Caspases by Major Green Tea Polyphenol Components during Cell Survival and Death. Arch. Pharm. Res. 2000, 23, 605–612. [Google Scholar] [CrossRef]
- Kweon, M.-H.; In Park, Y.; Sung, H.-C.; Mukhtar, H. The Novel Antioxidant 3-O-Caffeoyl-1-Methylquinic Acid Induces Nrf2-Dependent Phase II Detoxifying Genes and Alters Intracellular Glutathione Redox. Free Radic. Biol. Med. 2006, 40, 1349–1361. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Kundu, J.K.; Na, H.-K. Nrf2 as a Master Redox Switch in Turning on the Cellular Signaling Involved in the Induction of Cytoprotective Genes by Some Chemopreventive Phytochemicals. Planta Med. 2008, 74, 1526–1539. [Google Scholar] [CrossRef]
- Lunder, M.; Roškar, I.; Hošek, J.; Štrukelj, B. Silver Fir (Abies Alba) Extracts Inhibit Enzymes Involved in Blood Glucose Management and Protect against Oxidative Stress in High Glucose Environment. Plant Foods Hum. Nutr. 2019, 74, 47–53. [Google Scholar] [CrossRef]
- Chandrasekaran, K.; Swaminathan, K.; Chatterjee, S.; Dey, A. Apoptosis in HepG2 Cells Exposed to High Glucose. Toxicol. In Vitro 2010, 24, 387–396. [Google Scholar] [CrossRef]
- Xu, D.-P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.-J.; Li, H.-B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Koo, K.A.; Kim, S.H.; Oh, T.H.; Kim, Y.C. Acteoside and Its Aglycones Protect Primary Cultures of Rat Cortical Cells from Glutamate-Induced Excitotoxicity. Life Sci. 2006, 79, 709–716. [Google Scholar] [CrossRef]
- Šmejkal, K.; Grycová, L.; Marek, R.; Lemière, F.; Jankovská, D.; Forejtníková, H.; Vančo, J.; Suchý, V. C-Geranyl Compounds from Paulownia Tomentosa Fruits. J. Nat. Prod. 2007, 70, 1244–1248. [Google Scholar] [CrossRef]
- Veselá, D.; Kubınová, R.; Muselık, J.; Žemlička, M.; Suchý, V. Antioxidative and EROD Activities of Osajin and Pomiferin. Fitoterapia 2004, 75, 209–211. [Google Scholar] [CrossRef]
- Cheng, P.-S.; Hu, C.-C.; Wang, C.-J.; Lee, Y.-J.; Chung, W.-C.; Tseng, T.-H. Involvement of the Antioxidative Property of Morusin in Blocking Phorbol Ester–Induced Malignant Transformation of JB6 P+ Mouse Epidermal Cells. Chem.-Biol. Interact. 2017, 264, 34–42. [Google Scholar] [CrossRef]
- Šmejkal, K.; Babula, P.; Šlapetová, T.; Brognara, E.; Dall’Acqua, S.; Žemlička, M.; Innocenti, G.; Cvačka, J. Cytotoxic Activity of C-Geranyl Compounds from Paulownia Tomentosa Fruits. Planta Med. 2008, 74, 1488–1491. [Google Scholar] [CrossRef]
- Hošek, J.; Bartos, M.; Chudík, S.; Dall’Acqua, S.; Innocenti, G.; Kartal, M.; Kokoška, L.; Kollár, P.; Kutil, Z.; Landa, P.; et al. Natural Compound Cudraflavone B Shows Promising Anti-Inflammatory Properties In Vitro. J. Nat. Prod. 2011, 74, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Hošek, J.; Závalová, V.; Šmejkal, K.; Bartoš, M. Effect of Diplacone on LPS-Induced Inflammatory Gene Expression in Macrophages. Folia Biol. (Praha) 2010, 56, 124–130. [Google Scholar] [PubMed]
- Zelová, H.; Hanáková, Z.; Čermáková, Z.; Šmejkal, K.; Dalĺ Acqua, S.; Babula, P.; Cvačka, J.; Hošek, J. Evaluation of Anti-Inflammatory Activity of Prenylated Substances Isolated from Morus Alba and Morus Nigra. J. Nat. Prod. 2014, 77, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Kollar, P.; Bárta, T.; Hošek, J.; Souček, K.; Závalová, V.M.; Artinian, S.; Talhouk, R.; Šmejkal, K.; Suchý, P.; Hampl, A. Prenylated Flavonoids from Morus Alba L. Cause Inhibition of G1/S Transition in THP-1 Human Leukemia Cells and Prevent the Lipopolysaccharide-Induced Inflammatory Response. Evid.-Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Lee, K.-W.; Kim, H.J.; Lee, Y.S.; Park, H.-J.; Choi, J.-W.; Ha, J.; Lee, K.-T. Acteoside Inhibits Human Promyelocytic HL-60 Leukemia Cell Proliferation via Inducing Cell Cycle Arrest at G0/G1 Phase and Differentiation into Monocyte. Carcinogenesis 2007, 28, 1928–1936. [Google Scholar] [CrossRef]
- Sgarbossa, A.; Dal Bosco, M.; Pressi, G.; Cuzzocrea, S.; Dal Toso, R.; Menegazzi, M. Phenylpropanoid Glycosides from Plant Cell Cultures Induce Heme Oxygenase 1 Gene Expression in a Human Keratinocyte Cell Line by Affecting the Balance of NRF2 and BACH1 Transcription Factors. Chem.-Biol. Interact. 2012, 199, 87–95. [Google Scholar] [CrossRef]
- Nam, S.-Y.; Kim, H.-M.; Jeong, H.-J. Attenuation of IL-32-Induced Caspase-1 and Nuclear Factor-ΚB Activations by Acteoside. Int. Immunopharmacol. 2015, 29, 574–582. [Google Scholar] [CrossRef]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Reale, M.; Menghini, L.; Vinciguerra, I.; Lutiis, M.A.D.; Felaco, M.; Grilli, A. Antiinflammatory Effects in THP-1 Cells Treated with Verbascoside. Phytother. Res. 2010, 24, 1398–1404. [Google Scholar] [CrossRef]
- Son, I.H.; Chung, I.-M.; Lee, S.I.; Yang, H.D.; Moon, H.-I. Pomiferin, Histone Deacetylase Inhibitor Isolated from the Fruits of Maclura Pomifera. Bioorg. Med. Chem. Lett. 2007, 17, 4753–4755. [Google Scholar] [CrossRef]
- Yang, R.; Hanwell, H.; Zhang, J.; Tsao, R.; Meckling, K.A. Antiproliferative Activity of Pomiferin in Normal (MCF-10A) and Transformed (MCF-7) Breast Epithelial Cells. J. Agric. Food Chem. 2011, 59, 13328–13336. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: Oxford, UK, 2007; ISBN 978-0-19-856869-8. [Google Scholar]
- Siciliano, T.; Bader, A.; Vassallo, A.; Braca, A.; Morelli, I.; Pizza, C.; De, T. Secondary Metabolites from Ballota Undulata (Lamiaceae). Biochem. Syst. Ecol. 2005, 33, 341–351. [Google Scholar] [CrossRef]
- Li, X.; Xie, Y.; Li, K.; Wu, A.; Xie, H.; Guo, Q.; Xue, P.; Maleshibek, Y.; Zhao, W.; Guo, J.; et al. Antioxidation and Cytoprotection of Acteoside and Its Derivatives: Comparison and Mechanistic Chemistry. Molecules 2018, 23, 498. [Google Scholar] [CrossRef]
- Lee, H.-D.; Kim, J.H.; Pang, Q.Q.; Jung, P.-M.; Cho, E.J.; Lee, S. Antioxidant Activity and Acteoside Analysis of Abeliophyllum Distichum. Antioxidants 2020, 9, 1148. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.A.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J.; Ischiropoulos, H. Measuring Reactive Oxygen and Nitrogen Species with Fluorescent Probes: Challenges and Limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef]
- Hošek, J.; Toniolo, A.; Neuwirth, O.; Bolego, C. Prenylated and Geranylated Flavonoids Increase Production of Reactive Oxygen Species in Mouse Macrophages but Inhibit the Inflammatory Response. J. Nat. Prod. 2013, 76, 1586–1591. [Google Scholar] [CrossRef]
- Orhan, I.E.; Senol, F.S.; Demirci, B.; Dvorska, M.; Smejkal, K.; Zemlicka, M. Antioxidant Potential of Some Natural and Semi-Synthetic Flavonoid Derivatives and the Extracts from Maclura Pomifera (Rafin.) Schneider (Osage Orange) and Its Essential Oil Composition. Turk. J. Chem. 2016, 41, 403–411. [Google Scholar] [CrossRef]
- Bozkurt, İ.; Dilek, E.; Erol, H.S.; Çakir, A.; Hamzaoğlu, E.; Koç, M.; Keleş, O.N.; Halici, M.B. Investigation on the Effects of Pomiferin from Maclura Pomifera on Indomethacin-Induced Gastric Ulcer: An Experimental Study in Rats. Med. Chem. Res. 2017, 26, 2048–2056. [Google Scholar] [CrossRef]
- Hwang, H.-S.; Winkler-Moser, J.K.; Tisserat, B.; Harry-O’kuru, R.E.; Berhow, M.A.; Liu, S.X. Antioxidant Activity of Osage Orange Extract in Soybean Oil and Fish Oil during Storage. J. Am. Oil Chem. Soc. 2021, 98, 73–87. [Google Scholar] [CrossRef]
- Treml, J.; Šmejkal, K.; Hošek, J.; Žemlička, M. Determination of Antioxidant Activity Using Oxidative Damage to Plasmid DNA—Pursuit of Solvent Optimization. Chem. Pap. 2013, 67, 484–489. [Google Scholar] [CrossRef]
- Zima, A.; Hošek, J.; Treml, J.; Muselík, J.; Suchý, P.; Pražanová, G.; Lopes, A.; Žemlička, M. Antiradical and Cytoprotective Activities of Several C-Geranyl-Substituted Flavanones from Paulownia Tomentosa Fruit. Molecules 2010, 15, 6035–6049. [Google Scholar] [CrossRef]
- Moon, H.-I.; Jeong, M.H.; Jo, W.S. Protective Activity of C-Geranylflavonoid Analogs from Paulownia Tomentosa against DNA Damage in 137Cs Irradiated AHH-1Cells. Nat. Prod. Comm. 2014, 9. [Google Scholar] [CrossRef]
- Malaník, M.; Treml, J.; Leláková, V.; Nykodýmová, D.; Oravec, M.; Marek, J.; Šmejkal, K. Anti-Inflammatory and Antioxidant Properties of Chemical Constituents of Broussonetia Papyrifera. Bioorg. Chem. 2020, 104, 104298. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lyu, D.H.; Koo, U.; Nam, K.-W.; Hong, S.S.; Kim, K.O.; Kim, K.H.; Lee, D.; Mar, W. Protection of Prenylated Flavonoids from Mori Cortex Radicis (Moraceae) against Nitric Oxide-Induced Cell Death in Neuroblastoma SH-SY5Y Cells. Arch. Pharm. Res. 2012, 35, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-G.; Matsuzaki, K.; Takamatsu, S.; Kitanaka, S. Inhibitory Effects of Constituents from Morus Alba Var. Multicaulis on Differentiation of 3T3-L1 Cells and Nitric Oxide Production in RAW264.7 Cells. Molecules 2011, 16, 6010–6022. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.-H.; Wang, J.-J.; Lin, H.-C.; Wang, J.-P.; Lin, C.-N. Chemistry and Biological Activities of Constituents from Morus Australis. Biochim. Biophys. Acta Gen. Subj. 1999, 1428, 293–299. [Google Scholar] [CrossRef]
- Kawahito, S.; Kitahata, H.; Oshita, S. Problems Associated with Glucose Toxicity: Role of Hyperglycemia-Induced Oxidative Stress. World J. Gastroenterol. 2009, 15, 4137–4142. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Morón, E.; Abad-Jiménez, Z.; Martínez de Marañón, A.; Iannantuoni, F.; Escribano-López, I.; López-Domènech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J. Clin. Med. 2019, 8, 1385. [Google Scholar] [CrossRef]
- El-Marasy, S.A.; El-Shenawy, S.M.; Moharram, F.A.; El-Sherbeeny, N.A. Antidiabetic and Antioxidant Effects of Acteoside from Jacaranda Mimosifolia Family Biognoniaceae in Streptozotocin–Nicotinamide Induced Diabetes in Rats. Open Access Maced. J. Med. Sci. 2020, 8, 125–133. [Google Scholar] [CrossRef]
- Yu, S.Y.; Lee, I.-S.; Jung, S.-H.; Lee, Y.M.; Lee, Y.-R.; Kim, J.-H.; Sun, H.; Kim, J.S. Caffeoylated Phenylpropanoid Glycosides from Brandisia Hancei Inhibit Advanced Glycation End Product Formation and Aldose Reductase In Vitro and Vessel Dilation in Larval Zebrafish In Vivo. Planta Med. 2013, 79, 1705–1709. [Google Scholar] [CrossRef]
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.-A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef]
- Huan, S.K.-H.; Wang, K.-T.; Lee, C.-J.; Sung, C.-H.; Chien, T.-Y.; Wang, C.-C. Wu-Chia-Pi Solution Attenuates Carbon Tetrachloride-Induced Hepatic Injury through the Antioxidative Abilities of Its Components Acteoside and Quercetin. Molecules 2012, 17, 14673–14684. [Google Scholar] [CrossRef]
- Carocho, M.; Ferreira, I.C.F.R. A Review on Antioxidants, Prooxidants and Related Controversy: Natural and Synthetic Compounds, Screening and Analysis Methodologies and Future Perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Galati, G.; O’Brien, P.J. Potential Toxicity of Flavonoids and Other Dietary Phenolics: Significance for Their Chemopreventive and Anticancer Properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and Prooxidant Properties of Flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Kanner, J. Polyphenols by Generating H2O2, Affect Cell Redox Signaling, Inhibit PTPs and Activate Nrf2 Axis for Adaptation and Cell Surviving: In Vitro, In Vivo and Human Health. Antioxidants 2020, 9, 797. [Google Scholar] [CrossRef]
- Treml, J.; Leláková, V.; Šmejkal, K.; Paulíčková, T.; Labuda, Š.; Granica, S.; Havlík, J.; Jankovská, D.; Padrtová, T.; Hošek, J. Antioxidant Activity of Selected Stilbenoid Derivatives in a Cellular Model System. Biomolecules 2019, 9, 468. [Google Scholar] [CrossRef]
- Hanáková, Z.; Hošek, J.; Babula, P.; Dall’Acqua, S.; Václavík, J.; Šmejkal, K. C-Geranylated Flavanones from Paulownia Tomentosa Fruits as Potential Anti-Inflammatory Compounds Acting via Inhibition of TNF-α Production. J. Nat. Prod. 2015, 78, 850–863. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Treml, J.; Šalamúnová, P.; Hanuš, J.; Hošek, J. The Effect of Curcumin Encapsulation into Yeast Glucan Particles on Antioxidant Enzyme Expression In Vitro. Food Funct. 2021. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Treml, J.; Večeřová, P.; Herczogová, P.; Šmejkal, K. Direct and Indirect Antioxidant Effects of Selected Plant Phenolics in Cell-Based Assays. Molecules 2021, 26, 2534. https://doi.org/10.3390/molecules26092534
Treml J, Večeřová P, Herczogová P, Šmejkal K. Direct and Indirect Antioxidant Effects of Selected Plant Phenolics in Cell-Based Assays. Molecules. 2021; 26(9):2534. https://doi.org/10.3390/molecules26092534
Chicago/Turabian StyleTreml, Jakub, Petra Večeřová, Petra Herczogová, and Karel Šmejkal. 2021. "Direct and Indirect Antioxidant Effects of Selected Plant Phenolics in Cell-Based Assays" Molecules 26, no. 9: 2534. https://doi.org/10.3390/molecules26092534
APA StyleTreml, J., Večeřová, P., Herczogová, P., & Šmejkal, K. (2021). Direct and Indirect Antioxidant Effects of Selected Plant Phenolics in Cell-Based Assays. Molecules, 26(9), 2534. https://doi.org/10.3390/molecules26092534