
Multiple Roles of Black Raspberry Anthocyanins Protecting against Alcoholic Liver Disease

 ,
,
Abstract
1. Introduction
2. Results and Discussion
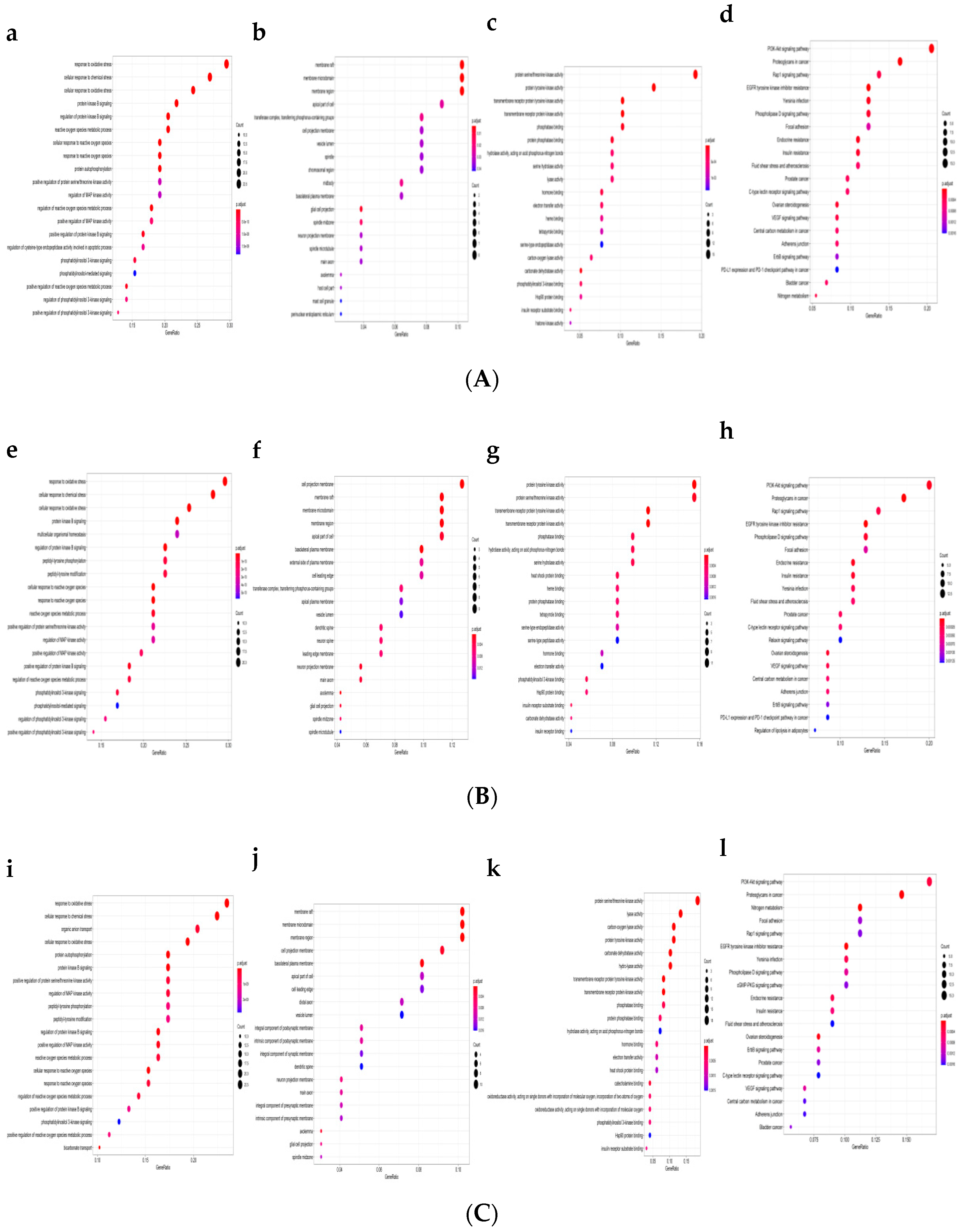
2.1. GO Functional Enrichment and Pathway Analysis
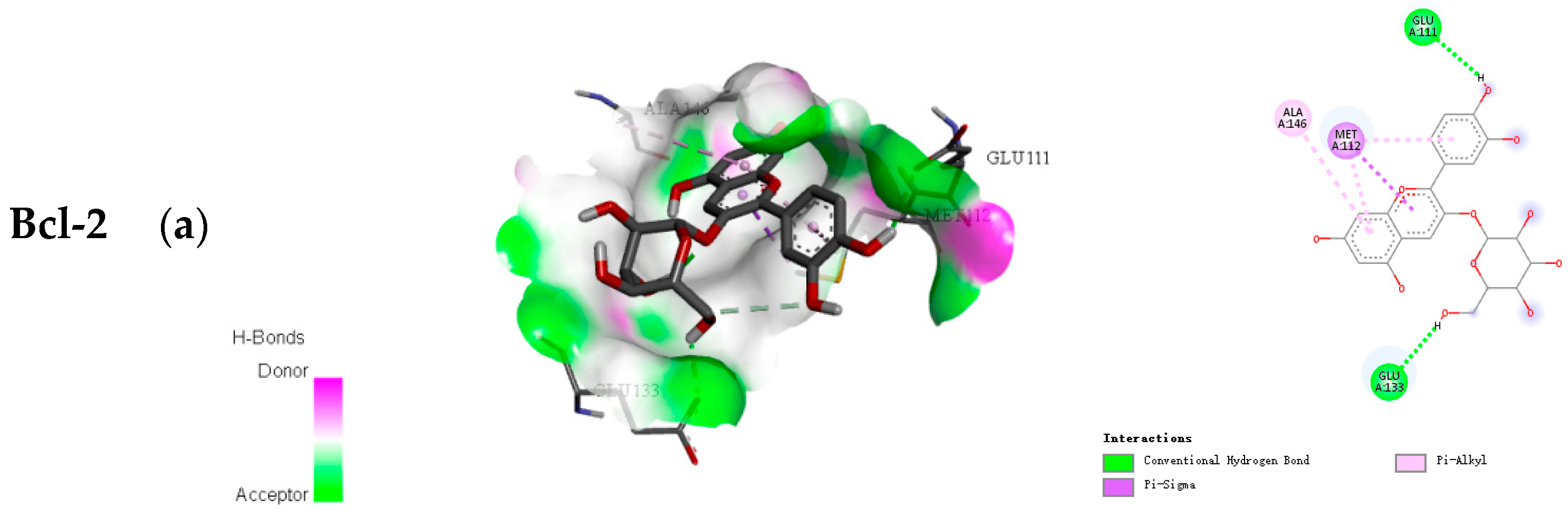
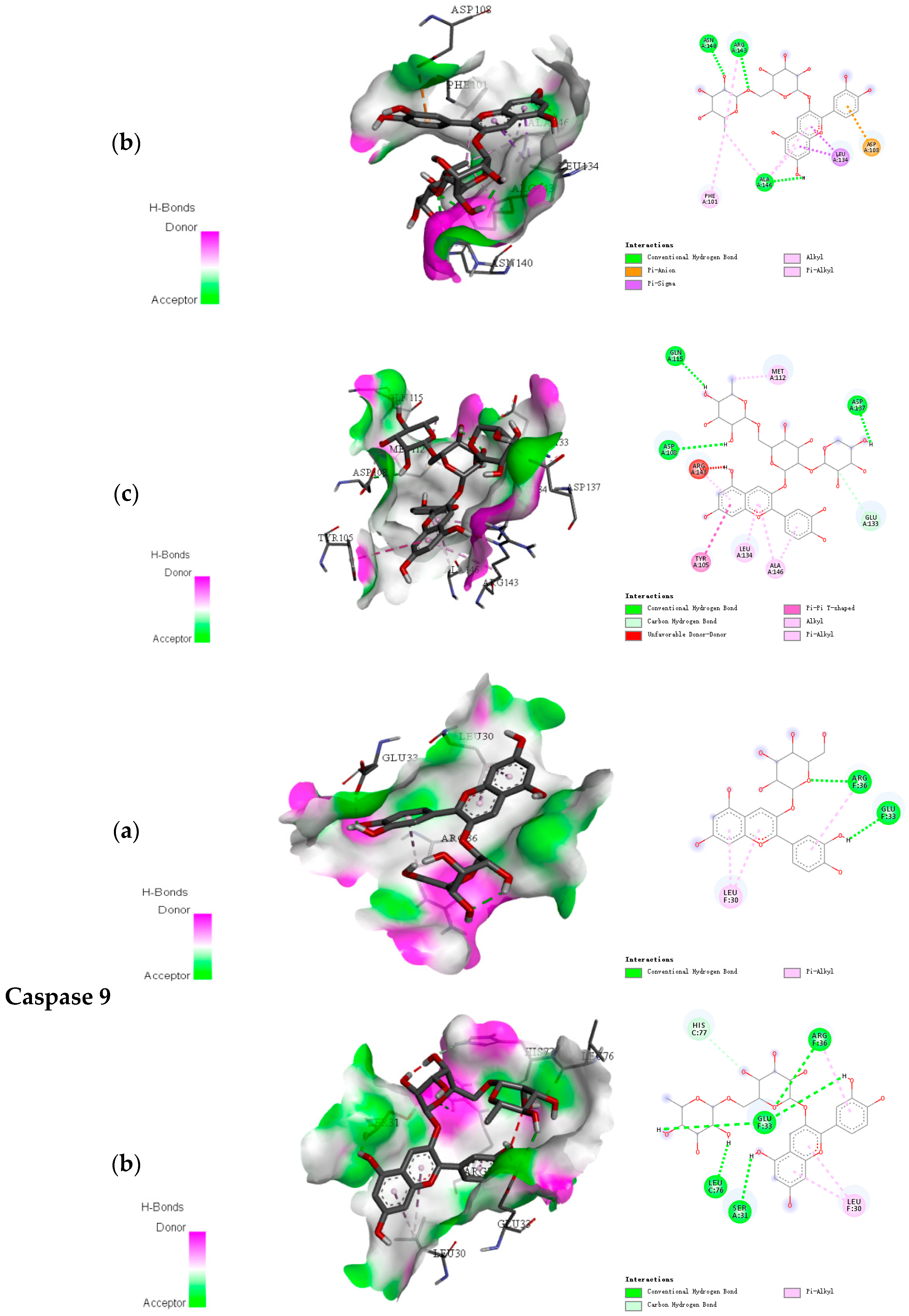
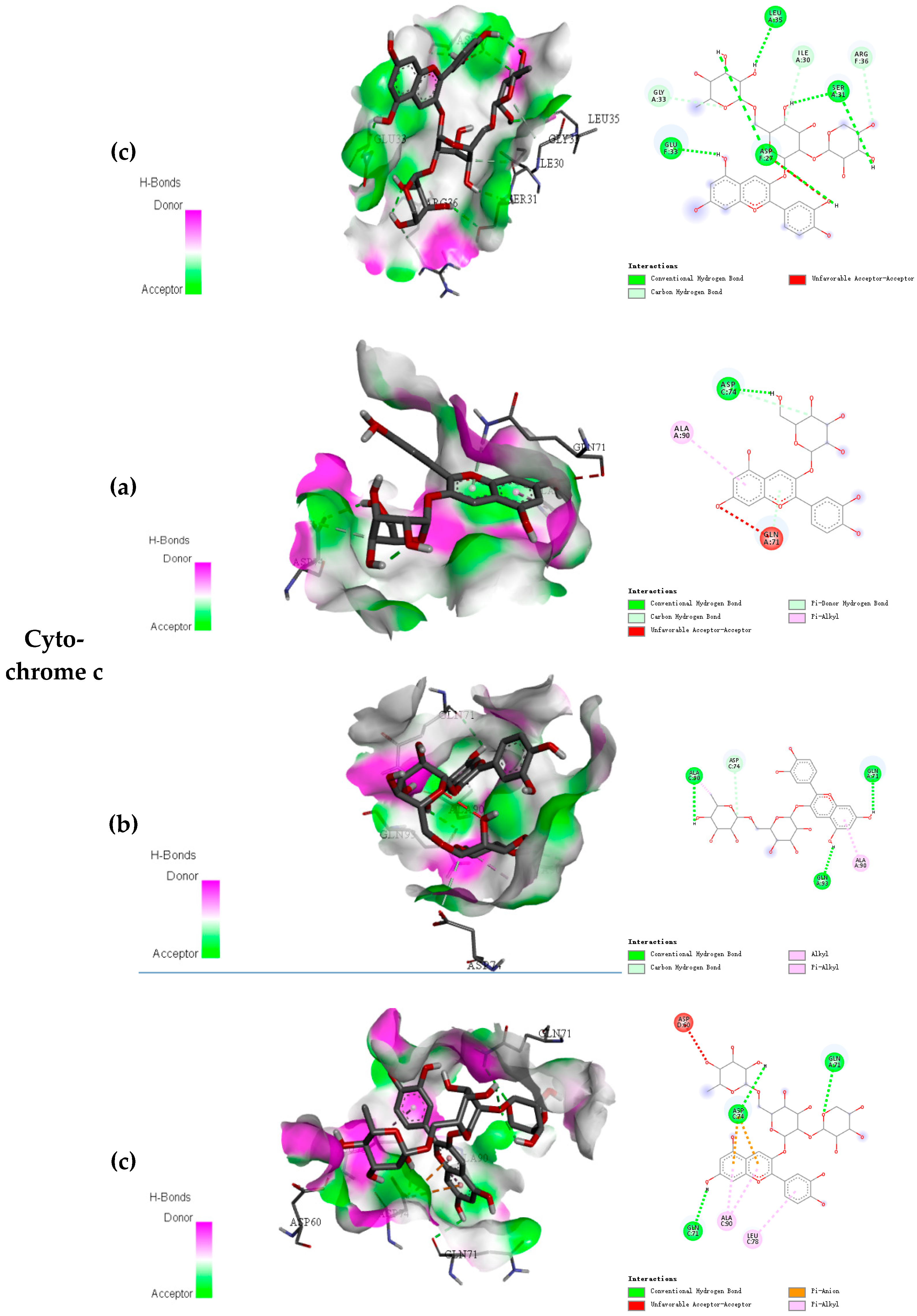
2.2. Docking Results of Anthocyanins
2.3. Antioxidant Activities
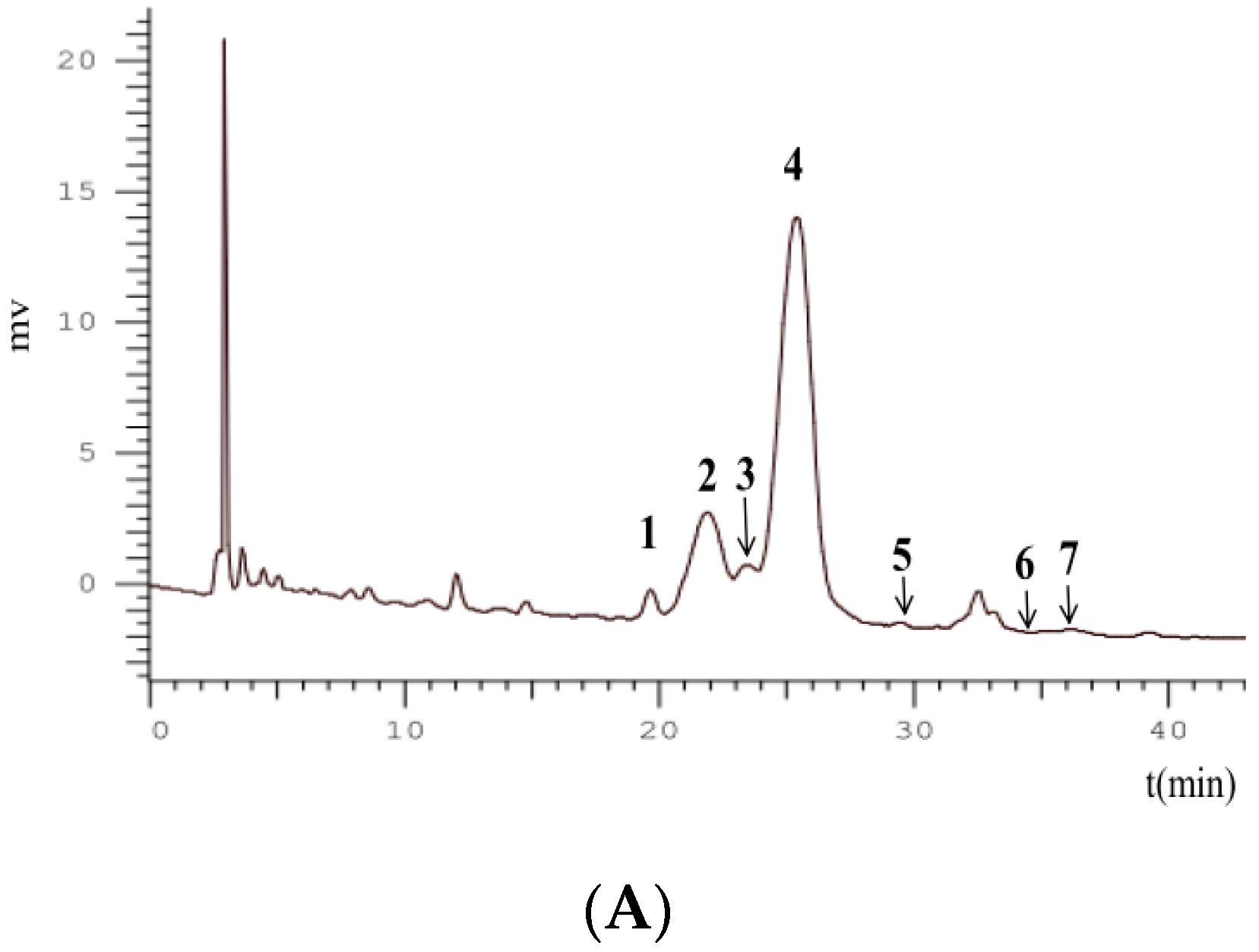
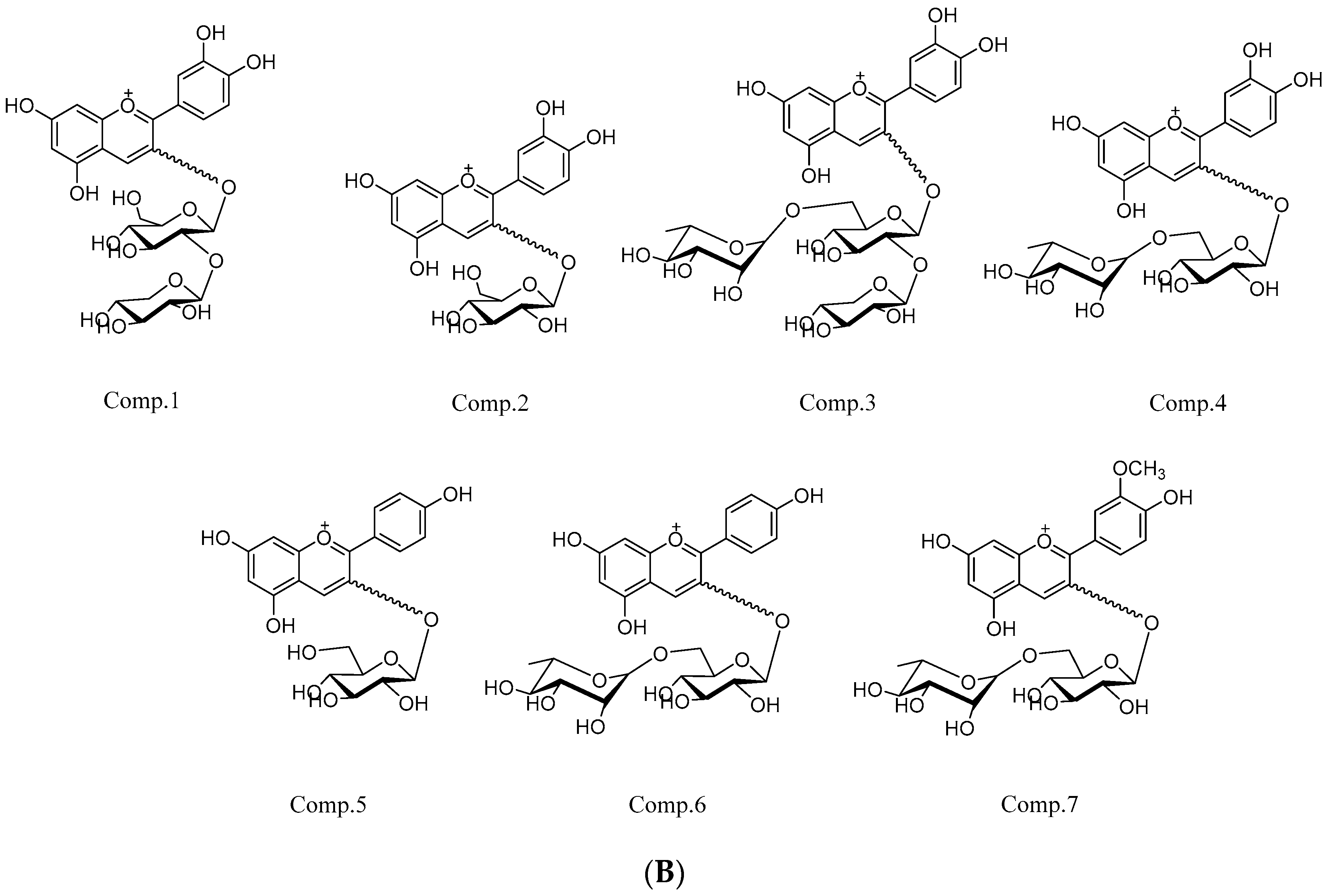
2.4. FT-ICR MS Analysis
2.5. Measurement of Biochemical Parameters
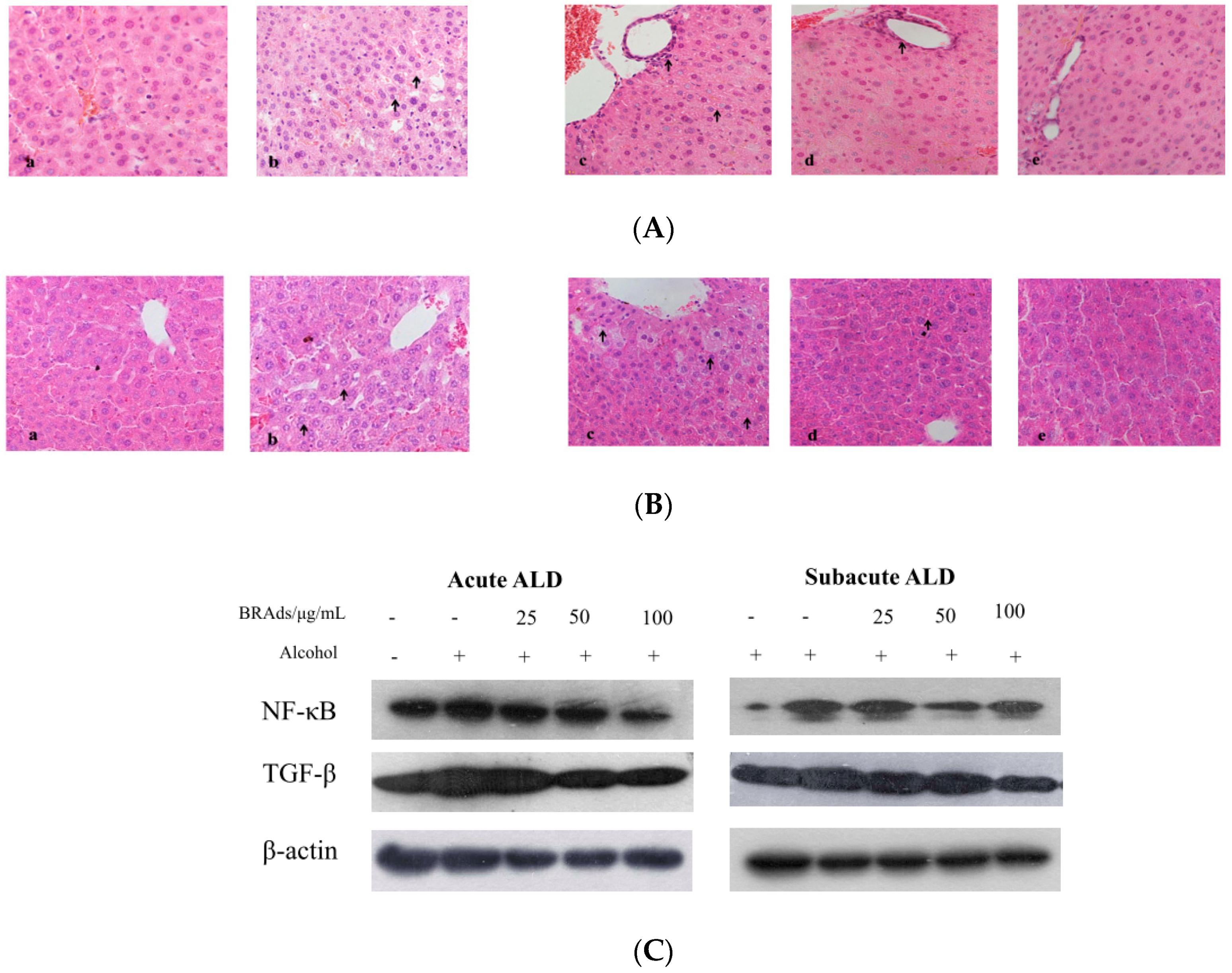
2.6. Histopathological Observation
2.7. Western Blotting Analysis
2.8. Anthocyanins in Black Raspberry-Induced Apoptosis on HepG2 Cells
2.8.1. The Antiproliferative Effect of Anthocyanins in Black Raspberry
2.8.2. Morphological Changes in HepG2 Cells
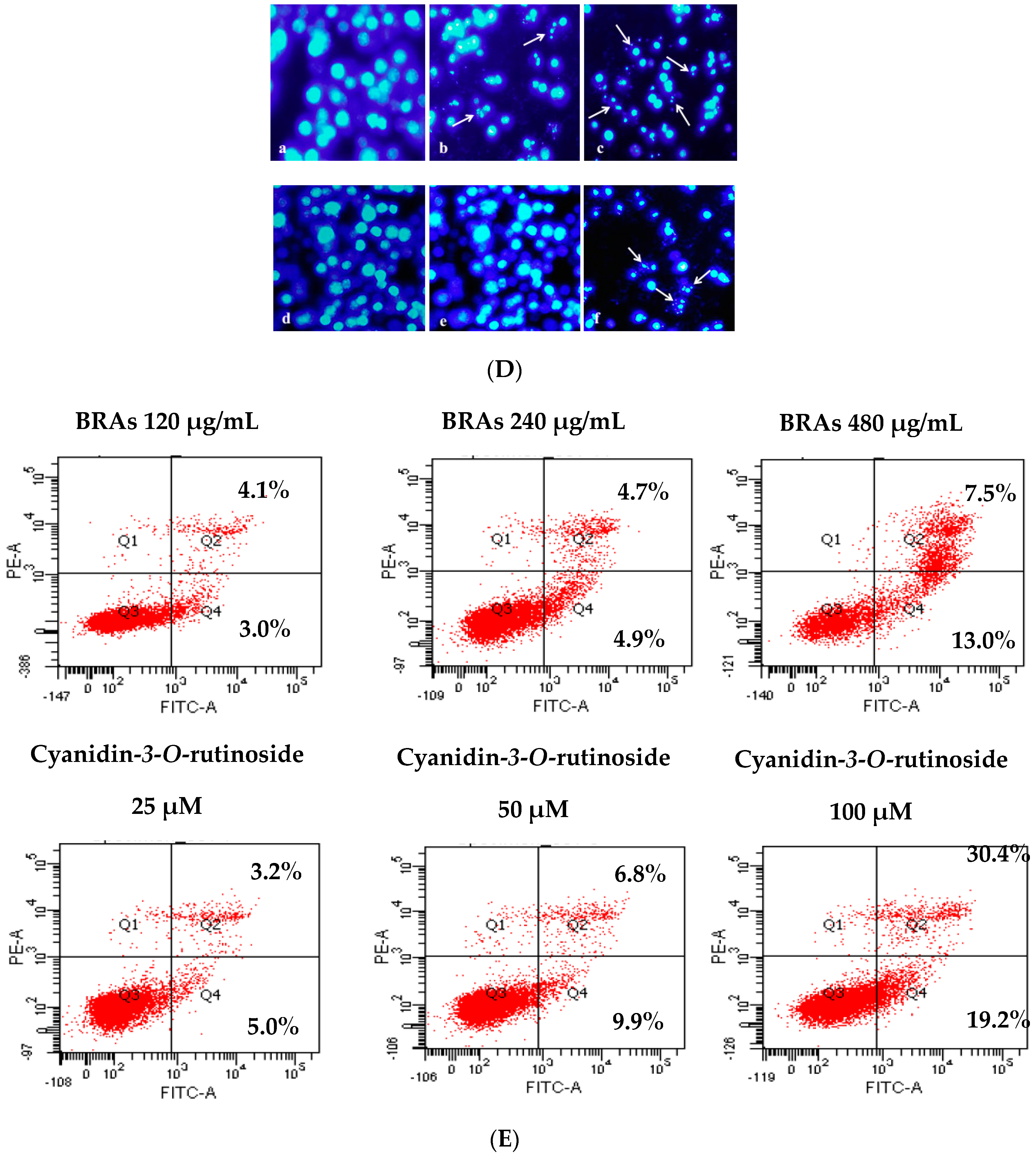
2.8.3. Anthocyanins Induce Apoptosis in HepG2 Cells
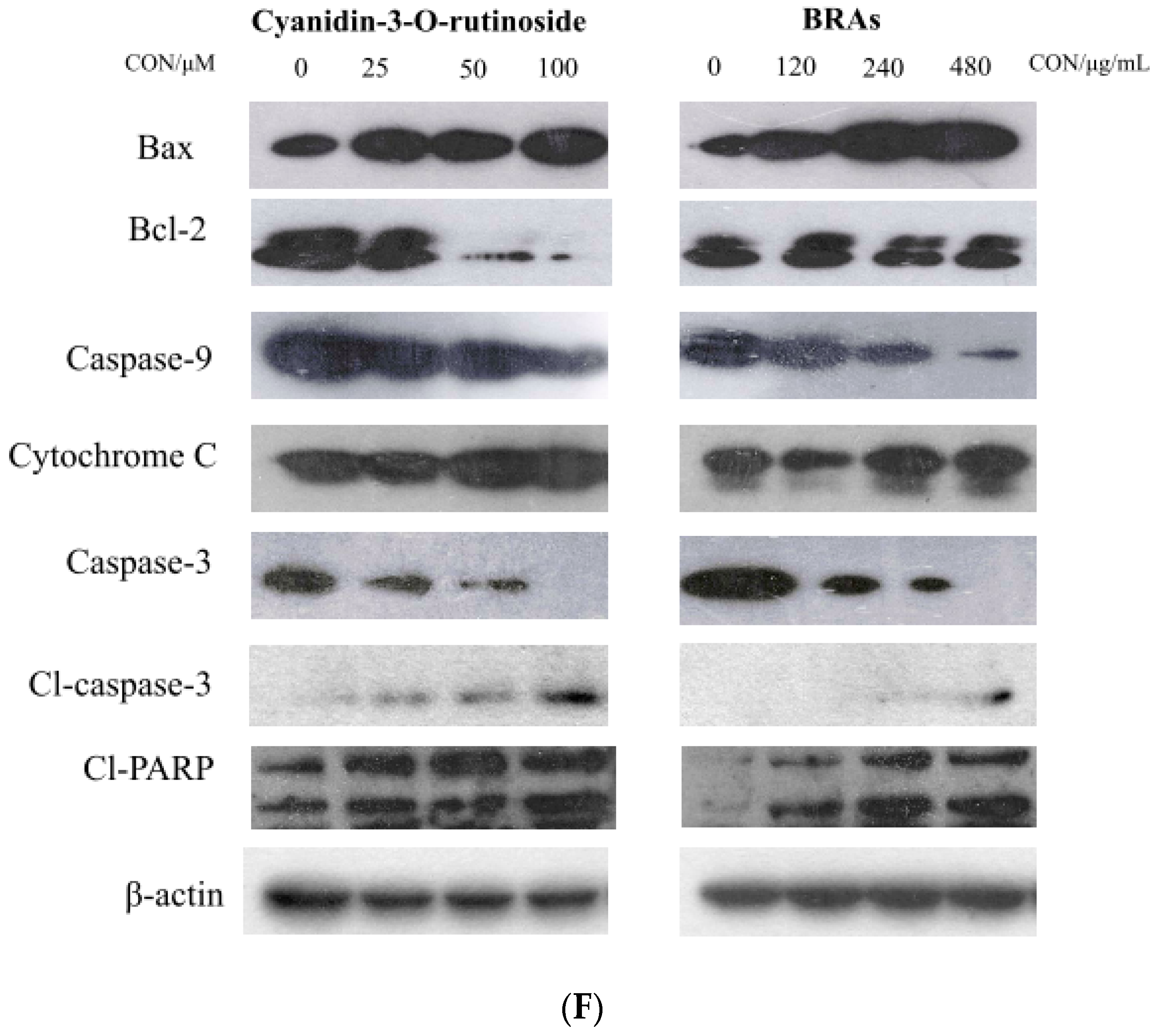
2.8.4. Effects of Anthocyanins on Expression of Cell Apoptosis-Related Proteins
3. Materials and Methods
3.1. Plant Materials and Chemicals
3.2. Gene Ontology and KEGG Analysis
3.3. In Silico Molecular Docking Study
3.4. Preparation of Black Raspberry Anthocyanins
3.5. Antioxidant Activities
3.6. HPLC-FT-ICR MS/MS Analysis
3.7. Animals and Treatment
3.7.1. Animals
3.7.2. Acute and Subacute ALD Mouse Model Design
3.7.3. Measurement of Biochemical Parameters
3.7.4. Histopathological Observation
3.7.5. Western Blot Analysis
3.8. Potential Mechanisms of BRAs against HepG2 Cells
3.8.1. Cell Viability Assay
3.8.2. Observation of Morphological Changes
3.8.3. Annexin V-FITC Apoptosis Detection by Flow Cytometry
3.8.4. Western Blot Analysis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| ABTS | 2,2′-azino-bis (3-ethyl-benzothiazoline-6-sulfonic acid) |
| ALD | Alcoholic liver disease |
| AST | Aspartate aminotransferase |
| ALT | Alanine aminotransferase |
| BRAs | Black raspberry anthocyanins |
| CHO | Serum cholesterol |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| FRAP | Ferric ion reducing antioxidant power |
| FT-ICR MS | Fourier transform-ion cyclotron resonance mass spectrometry |
| TPTZ | 2,4,6-tris(2-pyridyl)-s-triazine |
References
- Sun, H.; Mu, T.; Liu, X.; Zhang, M.; Chen, J. Purple Sweet Potato (Ipomoea batatas L.) Anthocyanins: Preventive Effect on Acute and Subacute Alcoholic Liver Damage and Dealcoholic Effect. J. Agric. Food Chem. 2014, 62, 2364–2373. [Google Scholar] [CrossRef]
- Rehm, J.; Mathers, C.; Popova, S.; Thavorncharoensap, M.; Teerawattananon, Y.; Patra, J. Global burden of disease and injury and economic cost attributable to alcohol use and alcohol-use disorders. Lancet 2009, 373, 2223–2233. [Google Scholar] [CrossRef]
- Goossens, N.; Hoshida, Y. Is hepatocellular cancer the same disease in alcoholic and non-alcoholic fatty liver diseases? Gastroenterology 2016, 150, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Status Report on Alcohol and Health 2018; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Bertola, A.; Mathews, S.; Ki, S.H.; Wang, H.; Gao, B. Mouse model of chronic and binge ethanol feeding (the NIAAA model). Nat. Protoc. 2013, 8, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Gowd, V.; Jia, Z.; Chen, W. Anthocyanins as promising molecules and dietary bioactive components against diabetes—A review of recent advances. Trends Food Sci. Tech. 2017, 68, 1–13. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, D.; Bao, T.; Xie, J.; Chen, W. A simple and rapid method for the preparation of pure delphinidin-3-O-sambubioside from Roselle and its antioxidant and hypoglycemic activity. J. Funct. Foods 2017, 39, 9–17. [Google Scholar] [CrossRef]
- Su, H.; Xie, L.; Xu, Y.; Ke, H.; Bao, T.; Li, Y.; Chen, W. Pelargonidin-3-O-glucoside Derived from Wild Raspberry Exerts Antihyperglycemic Effect by Inducing Autophagy and Modulating Gut Microbiota. J. Agric. Food Chem. 2020, 68, 13025–13037. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Qin, P.; Ren, G. Effect of Anthocyanin-Rich Extract from Black Rice (Oryza sativa L. Japonica) on Chronically Alcohol-Induced Liver Damage in Rats. J. Agric. Food Chem. 2010, 58, 3191–3196. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, P.; Zhu, Y.; Liu, X.; Hu, X.; Chen, F. Blueberry anthocyanins extract inhibits acrylamide-induced diverse toxicity in mice by preventing oxidative stress and cytochrome P450 2E1 activation. J. Funct. Foods 2015, 14, 95–101. [Google Scholar] [CrossRef]
- Morrison, M.-C.; Liang, W.; Mulder, P.; Verschuren, L.; Toet, E.-K.; Heeringa, P.; Wieling, P.-Y.; Kooistra, T.; Kleemann, R. Mirtoselect, an anthocyanin-rich bilberry extract, attenuates non-alcoholic steatohepatitis and associated fifibrosis in ApoE* 3Leiden mice. J. Hepatol. 2015, 62, 1180–1186. [Google Scholar] [CrossRef]
- Shi, L.; Liu, Y.; Tan, D.; Yan, T.; Song, D.; Hou, M.; Meng, X. Blueberry anthocyanins ameliorate cyclophosphamide-induced liver damage in rats by reducing inflflammation and apoptosis. J. Funct. Foods 2014, 11, 71–81. [Google Scholar] [CrossRef]
- Ke, H.; Bao, T.; Chen, W. Polysaccharide from Rubus chingii Hu affords protection against palmitic acid-induced lipotoxicity in human hepatocytes. Int. J. Biol. Macromol. 2019, 133, 1063–1071. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Xie, J.; Chen, W. Malvidin-3-O-arabinoside ameliorates ethyl carbamate-induced oxidative damage by stimulating AMPK-mediated autophagy. Food Funct. 2020, 11, 10317–10328. [Google Scholar] [CrossRef]
- Xiao, T.; Guo, Z.; Bi, X.; Zhao, Y. Polyphenolic profile as well as anti-oxidant and anti-diabetes effects of extracts from freeze-dried black raspberries. J. Funct. Foods 2017, 31, 179–187. [Google Scholar] [CrossRef]
- Medda, R.; Lyros, O.; Schmidt, J.L.; Jovanovic, N.; Nie, L.; Link, B.J.; Otterson, M.F.; Stoner, G.D.; Shaker, R.; Rafiee, P. Anti inflammatory and anti angiogenic effect of black raspberry extract on human esophageal and intestinal microvascular endothelial cells. Microvasc. Res. 2015, 97, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Knobloch, T.J.; Seamon, L.G.; Stoner, G.D.; Cohn, D.E.; Paskett, E.D.; Fowler, J.M.; Weghorst, C.M. A black raspberry extract inhibits proliferation and regulates apoptosis in cervical cancer cells. Gynecol. Oncol. 2011, 123, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; States, V.A.; Li, S.; Zhang, X.; Martin, R.C.G. The Effect of Black Raspberry Extracts on MnSOD Activity in Protection Against Concanavalin a Induced Liver Injury. Nutr. Cancer 2014, 66, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fan, T.-P.; Jia, W.; Lu, A.; Zhang, W. Network Pharmacology in Traditional Chinese Medicine. Evidence-Based Complement. Altern. Med. 2014, 2014, 138460. [Google Scholar] [CrossRef]
- Simona, O.; Letitia, O. Anthocyanins, from biosynthesis in plant to human health. Acta Univ. Cinbinesis 2011, 15, 1–3. [Google Scholar]
- Nordmann, R.; Ribière, C.; Rouach, H. Implication of free radical mechanisms in ethanol-induced cellular injury. Free Radic. Biol. Med. 1992, 12, 219–240. [Google Scholar] [CrossRef]
- Wang, W.; Li, J.; Wang, Z.; Gao, H.; Su, L.; Xie, J.; Chen, X.; Liang, H.; Wang, C.; Han, Y. Oral Hepatoprotective Ability Evaluation of Purple Sweet Potato Anthocyanins on Acute and Chronic Chemical Liver Injuries. Cell Biophys. 2014, 69, 539–548. [Google Scholar] [CrossRef]
- Xiao, T.; Guo, Z.; Sun, B.; Zhao, Y. Identification of Anthocyanins from Four Kinds of Berries and Their Inhibition Activity to α-Glycosidase and Protein Tyrosine Phosphatase 1B by HPLC–FT-ICR MS/MS. J. Agric. Food Chem. 2017, 65, 6211–6221. [Google Scholar] [CrossRef]
- Pingitore, F.; Tang, Y.; Kruppa, A.G.H.; Keasling, J.D. Analysis of Amino Acid Isotopomers Using FT-ICR MS. Anal. Chem. 2007, 79, 2483–2490. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Ren, C.; Mallery, S.R.; Ugalde, C.M.; Pei, P.; Saradhi, U.V.; Stoner, G.D.; Chan, K.K.; Liu, Z. A rapid and sensitive LC–MS/MS method for quantification of four anthocyanins and its application in a clinical pharmacology study of a bioadhesive black raspberry gel. J. Chromatogr. B 2009, 877, 4027–4034. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xin, X.; Zhang, H.; Yuan, Q. Phytochemical properties and antioxidant capacities of commercial raspberry varieties. J. Funct. Foods 2013, 5, 508–515. [Google Scholar] [CrossRef]
- Wu, X.; Pittman, I.H.E.; Prior, R.L. Fate of Anthocyanins and Antioxidant Capacity in Contents of the Gastrointestinal Tract of Weanling Pigs Following Black Raspberry Consumption. J. Agric. Food Chem. 2006, 54, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Zhu, W.; Li, Z.; Ling, S. Effect of juice and fermented vinegar from Hovenia dulcis peduncles on chronically alcohol-induced liver damage in mice. Food Funct. 2012, 3, 628. [Google Scholar] [CrossRef]
- Wondergem, R.; Davis, J. Ethanol Increases Hepatocyte Water Volume. Alcohol. Clin. Exp. Res. 1994, 18, 1230–1236. [Google Scholar] [CrossRef]
- Cai, Z.; Song, L.; Qian, B.; Xu, W.; Ren, J.; Jing, P.; Oey, I. Understanding the effect of anthocyanins extracted from purple sweet potatoes on alcohol-induced liver injury in mice. Food Chem. 2018, 245, 463–470. [Google Scholar] [CrossRef]
- Baldi, E.; Burra, P.; Plebani, M.; Salvagnini, M. Serum malondialdehyde and mitochondrial aspartate aminotransferase activity as markers of chronic alcohol intake and alcoholic liver disease. Ital. J. Gastroenterol. 1993, 25, 429–432. [Google Scholar]
- Diosse, L.; Levy, D.; Cupples, L.-A.; Ellison, R.-C. Total serum bilirubin and risk of cardiovascular disease in the Franningham off spring study. Am. J. Cardiol. 2001, 87, 1196–1200. [Google Scholar]
- Sozio, M.; Crabb, D.W. Alcohol and lipid metabolism. Am. J. Physiol. Metab. 2008, 295, E10–E16. [Google Scholar] [CrossRef]
- Yu, W.; Zeng, Y.; Zheng, W.; Chen, Y. The investigation on protective effects of grape seed proanthocyanidins on alcoholic liver injury in rats based on Nrf2/OH-1/NF-κB pathway. J. Emerg. Tradit. Chin. Med. 2019, 28, 1021–1023. [Google Scholar]
- Lingappan, K. NF-κB in oxidative stress. Curr. Opin. Toxicol. 2017, 7, 81–86. [Google Scholar] [CrossRef]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef]
- Ni, M.-M.; Xu, T.; Wang, Y.-R.; He, Y.-H.; Zhou, Q.; Huang, C.; Meng, X.-M.; Li, J. Inhibition of IRF3 expression reduces TGF-β1-induced proliferation of hepatic stellate cells. J. Physiol. Biochem. 2015, 72, 9–23. [Google Scholar] [CrossRef]
- Török, N.J. Recent advances in the pathogenesis and diagnosis of liver fibrosis. J. Gastroenterol. 2008, 43, 315–321. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, X.; Ding, J. Characteristics of liver cancer stem cells and clinical correlations. Cancer Lett. 2016, 379, 230–238. [Google Scholar] [CrossRef] [PubMed]
- De Wang, X.; Sun, Y.Y.; Zhao, C.; Qu, F.Z.; Zhao, Y.Q. 12-Chloracetyl-PPD, a novel dammarane derivative, shows anti-cancer activity via delay the progression of cell cycle G2/M phase and reactive oxygen species-mediate cell apoptosis. Eur. J. Pharmacol. 2017, 798, 49–56. [Google Scholar] [CrossRef]
- Larsen, B.-D.; Sørensen, C.-S. The caspase-activated DNase: Apoptosis and beyond. FEBS J. 2017, 284, 1160–1170. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. Bcl-2-regulated apoptosis: Mechanism and therapeutic potential. Curr. Opin. Immunol. 2007, 19, 488–496. [Google Scholar] [CrossRef]
- Gu, X.; Yao, Y.; Cheng, R.; Zhang, Y.; Dai, Z.; Wan, G.; Yang, Z.; Cai, W.; Gao, G.; Yang, X. Plasminogen K5 activates mitochondrial apoptosis pathway in endothelial cells by regulating Bak and Bcl-xL subcellular distribution. Apoptosis 2011, 16, 846–855. [Google Scholar] [CrossRef]
- Mazumder, S.; Plesca, A.; Almasan, A. Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis. Methods Mol. Biol. 2008, 414, 13–21. [Google Scholar] [CrossRef]
- Gaude, P.; Dessimoz, C. Gene Ontology: Pitfalls, Biases, and Remedies. Methods Mol. Biol. 2017, 1446, 189–205. [Google Scholar]
- Zhang, G.; Wang, H.; Zhu, K.; Yang, Y.; Li, J.; Jiang, H.; Liu, Z. Investigation of candidate molecular biomarkers for expression profile analysis of the Gene expression omnibus (GEO) in acute lymphocytic leukemia (ALL). Biomed. Pharmacother. 2019, 120, 109530. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Ma, X.; Zhao, X.; Zhang, S. Baicalein induces apoptosis and autophagy of breast cancer cells via inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des. Dev. Ther. 2018, 12, 3961–3972. [Google Scholar] [CrossRef]
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.; Mills, G.B. Exploiting the PI3K/AKT Pathway for Cancer Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar] [CrossRef]
- Feng, L.-X.; Sun, P.; Mi, T.; Liu, M.; Liu, W.; Yao, S.; Cao, Y.-M.; Yu, X.-L.; Wu, W.-Y.; Jiang, B.-H.; et al. Agglutinin isolated from Arisema heterophyllum Blume induces apoptosis and autophagy in A549 cells through inhibiting PI3K/Akt pathway and inducing ER stress. Chin. J. Nat. Med. 2016, 14, 856–864. [Google Scholar] [CrossRef]
- Yuriev, E.; Ramsland, P.A. Latest developments in molecular docking: 2010–2011 in review. J. Mol. Recognit. 2013, 26, 215–239. [Google Scholar] [CrossRef]
- De Sá, M.; Justino, V.; Spranger, M.I.; Zhao, Y.Q.; Han, L.; Sun, B.S. Extraction Yields and Anti-oxidant Activity of Proanthocyanidins from Different Parts of Grape Pomace: Effect of Mechanical Treatments. Phytochem. Anal. 2013, 25, 134–140. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Sun, B.; Neves, A.C.; Fernandes, T.A.; Fernandes, A.L.; Mateus, N.; De Freitas, V.; Leandro, C.; Spranger, M.I. Evolution of Phenolic Composition of Red Wine during Vinification and Storage and Its Contribution to Wine Sensory Properties and Antioxidant Activity. J. Agric. Food Chem. 2011, 59, 6550–6557. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targets | Compound | Binding Free Energy (kcal/mol) | Numbers of Bonds |
|---|---|---|---|
| Bcl-2 | Cyanidin-3-O-glucoside | −7.3 | 6 |
| Cyanidin-3-O-rutinoside | −8.2 | 11 | |
| Cyanidin-3-O-xylosylrutinoside | −8.3 | 11 | |
| Caspase 9 | Cyanidin-3-O-glucoside | −7.6 | 4 |
| Cyanidin-3-O-rutinoside | −8.0 | 6 | |
| Cyanidin-3-O-xylosylrutinoside | −8.3 | 9 | |
| Cytochrome c | Cyanidin-3-O-glucoside | −7.2 | 5 |
| Cyanidin-3-O-rutinoside | −7.7 | 9 | |
| Cyanidin-3-O-xylosylrutinoside | −7.7 | 10 |
| No. | RT (min) | Observed Mass (Da) | Calculated Mass (Da) | Error (ppm) | MS/MS | Formula | Identification | References |
|---|---|---|---|---|---|---|---|---|
| 1 | 19.85 | 581.15071 | 581.15010 | −1.03 | 287.05478 | C26H29O15 | Cyanidin-3-O-sambubioside | [25] |
| 2 | 21.99 | 449.10530 | 449.10784 | −0.71 | 287.05440 | C21H21O11 | Cyanidin-3-O-glucoside | [26] |
| 3 | 23.51 | 727.20315 | 727.20811 | −1.04 | 287.05455 | C32H39O19 | Cyanidin-3-O-xylosyl-rutinoside | [26] |
| 4 | 25.78 | 595.16217 | 595.16575 | −0.23 | 287.05520 | C27H31O15 | Cyanidin-3-O-rutinoside | [26] |
| 5 | 29.50 | 433.11031 | 433.11292 | −0.16 | 271.05945 | C21H21O10 | Pelargonidin-3-O-glucoside | [26] |
| 6 | 34.50 | 579.16687 | 579.17083 | −1.46 | 271.05954 | C27H31O14 | Pelargonidin-3-O-rutinoside | [26] |
| 7 | 36.10 | 609.18196 | 609.18140 | −0.93 | 301.07027 | C28H33O15 | Peonidin-3-O-rutinoside | [27] |
| Treatment Group | Final Weight (g) | Liver Index (%) | AST (U/L) | ALT (U/L) | CHOL (U/L) | TBIL (U/L) | LDL (U/L) |
|---|---|---|---|---|---|---|---|
| (a) Subacute ALD mice | |||||||
| Control | 35.11 ± 0.91 | 4.10 ± 0.21 | 81.40 ± 1.45 | 28.60 ± 1.56 | 1.92 ± 0.78 | 1.23 ± 0.52 | 0.28 ± 0.18 |
| Model | 34.67 ± 0.62 * | 5.07 ± 0.47 *** | 114.14 ± 2.17 *** | 57.71 ± 1.14 *** | 2.59 ± 1.01 *** | 1.73 ± 1.08 *** | 0.48 ± 0.15 *** |
| Low-dose BRAs | 35.18 ± 0.72 | 4.86 ± 0.22 ### | 107.13 ± 2.02 # | 41.50 ± 1.11 ## | 2.44 ± 1.13 # | 1.45 ± 1.02 ### | 0.42 ± 0.13 |
| Middle-dose BRAs | 35.24 ± 0.64 | 4.54 ± 0.38 ### | 101.67 ± 1.67## | 37.33 ± 1.03 ### | 2.39 ± 0.76 ### | 1.31 ± 0.94 ### | 0.40 ± 0.12 |
| High-dose BRAs | 35.12 ± 0.56 | 4.33 ± 0.32 ### | 87.13 ± 1.32 ### | 31.38 ± 0.97 ### | 2.14 ± 0.81 ### | 1.26 ± 0.75 ### | 0.38 ± 0.09 # |
| (b) Acute ALD mice | |||||||
| Control | 35.14 ± 0.87 | 4.19 ± 0.22 | 81.43 ± 1.47 | 28.58 ± 1.26 | 1.94 ± 0.88 | 1.25 ± 0.49 | 0.29 ± 0.19 |
| Model | 34.78 ± 0.57 * | 5.26 ± 0.41 *** | 133.27 ± 2.74 *** | 62.56 ± 1.45 *** | 3.31 ± 1.45 *** | 1.94 ± 0.98 *** | 0.80 ± 0.54 *** |
| Low-dose BRAs | 35.13 ± 0.63 | 4.95 ± 0.34 ### | 122.16 ± 2.14 ## | 52.64 ± 1.32 ## | 3.06 ± 1.23 ## | 1.85 ± 0.79 | 0.73 ± 0.46 |
| Middle-dose BRAs | 35.11 ± 0.45 | 4.73 ± 0.31 ### | 113.77 ± 1.78 ### | 44.43 ± 1.13 ### | 3.01 ± 1.17 ### | 1.61 ± 0.54 # | 0.65 ± 0.43 |
| High-dose BRAs | 35.19 ± 0.55 | 4.29 ± 0.28 ### | 92.75 ± 1.23 ###,* | 32.80 ± 1.07 ### | 2.28 ± 1.02 ### | 1.33 ± 0.51 ### | 0.48 ± 0.32 # |
| Cell Line | IC50 (n = 3) | |||
|---|---|---|---|---|
| BRAs | Cyanidin-3-O-Rutinoside | Silymarin | Mitomycin C | |
| t-HSC/Cl-6 | 202.91 ± 10.17 a | 17.87 ± 2.43 a 30.04 ± 3.54 b | 192.19 ± 14.22 a | - |
| HepG2 | 198.63 ± 9.68 a | 13.46 ± 1.78 a 22.62 ± 2.89 b | - | 8.06 ± 1.12 a 24.10 ± 2.37 b |
| Hep3B | 181.00 ± 12.34 a | 9.95 ± 1.81 a 16.73 ± 2.37 b | - | 8.01 ± 1.15 a 23.96 ± 2.14 b |
| GSE-1 | 335.10 ± 18.65 a | 37.14 ± 4.07 a 62.42 ± 9.68 b | - | 9.11 ± 1.26 a 27.26 ± 2.72 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, T.; Luo, Z.; Guo, Z.; Wang, X.; Ding, M.; Wang, W.; Shen, X.; Zhao, Y. Multiple Roles of Black Raspberry Anthocyanins Protecting against Alcoholic Liver Disease. Molecules 2021, 26, 2313. https://doi.org/10.3390/molecules26082313
Xiao T, Luo Z, Guo Z, Wang X, Ding M, Wang W, Shen X, Zhao Y. Multiple Roles of Black Raspberry Anthocyanins Protecting against Alcoholic Liver Disease. Molecules. 2021; 26(8):2313. https://doi.org/10.3390/molecules26082313
Chicago/Turabian StyleXiao, Ting, Zhonghua Luo, Zhenghong Guo, Xude Wang, Meng Ding, Wei Wang, Xiangchun Shen, and Yuqing Zhao. 2021. "Multiple Roles of Black Raspberry Anthocyanins Protecting against Alcoholic Liver Disease" Molecules 26, no. 8: 2313. https://doi.org/10.3390/molecules26082313
APA StyleXiao, T., Luo, Z., Guo, Z., Wang, X., Ding, M., Wang, W., Shen, X., & Zhao, Y. (2021). Multiple Roles of Black Raspberry Anthocyanins Protecting against Alcoholic Liver Disease. Molecules, 26(8), 2313. https://doi.org/10.3390/molecules26082313