Abstract
Nacobbus aberrans ranks among the “top ten” plant-parasitic nematodes of phytosanitary importance. It causes significant losses in commercial interest crops in America and is a potential risk in the European Union. The nematicidal and phytotoxic activities of seven plant extracts against N. aberrans and Solanum lycopersicum were evaluated in vitro, respectively. The chemical nature of three nematicidal extracts (EC50,48h ≤ 113 µg mL−1) was studied through NMR analysis. Plant extracts showed nematicidal activity on second-stage juveniles (J2): (≥87%) at 1000 µg mL−1 after 72 h, and their EC50 values were 71.4–468.1 and 31.5–299.8 µg mL−1 after 24 and 48 h, respectively. Extracts with the best nematicidal potential (EC50,48h < 113 µg mL−1) were those from Adenophyllum aurantium, Alloispermum integrifolium, and Tournefortia densiflora, which inhibited L. esculentum seed growth by 100% at 20 µg mL−1. Stigmasterol (1), β-sitosterol (2), and α-terthienyl (3) were identified from A. aurantium, while 1, 2, lutein (4), centaurin (5), patuletin-7-β-O-glucoside (6), pendulin (7), and penduletin (8) were identified from A. integrifolium. From T. densiflora extract, allantoin (9), 9-O-angeloyl-retronecine (10), and its N-oxide (11) were identified. The present research is the first to report the effect of T. densiflora, A. integrifolium, and A. aurantium against N. aberrans and chemically characterized nematicidal extracts that may provide alternative sources of botanical nematicides.
1. Introduction
The plant parasite nematode, Nacobbus aberrans (Thorne, 1935; Thorne & Allen, 1994), also known as false root-knot, is distributed in Argentina, Bolivia, Chile, Ecuador, Mexico, Peru, and the USA. It reduces crop yield in tomatoes (Solanum lycopersicum L.), potatoes (Solanum tuberosum), beans (Phaseolus vulgaris L.), and sugar beets (Beta vulgaris). The nematode ranks among the “top ten” plant-parasitic nematodes of phytosanitary importance [1]. It is estimated it meets the criteria to be a potential risk in the EU [2]. Management strategies of plant-parasitic nematodes are cultural practices (crop rotation), mixed-cropping, organic amendments, resistant crop cultivars, biological control [3,4,5,6], chemical nematicides, and bioactive products of plant origin. Among these strategies, natural product usage represents a vital option for controlling phytopathogenic nematodes because of their low impact on the environment and non-target organisms. In the search for botanic nematicides, some of the most recent proposals are using Stevia rebaudiana and Origanum vulgare to control Meloidogyne; in vivo experiments showed this effect [7,8]. In the case of N. aberrans, crude herbal extracts from T. lunulate [9], Cosmos sulphureus [6], Senecio salignus [6], Witheringia stramoniifolia [6], and Lantana cámara [6] showed in vitro nematicidal activity at 500 µg mL−1 (>70%) to second-stage juvenile (J2) individuals. Simultaneously, in vivo protection from infection of Lycopersicum esculentum Mill and Capsicum annumm plants occurred with extracts of Tagetes erecta [9] and Trichilia galuca [10], respectively. There are only two reports of natural compounds with toxic potential for the control of N. aberrans: capsidiol acts as a nematostatic on N. aberrans J2 (90% immobility) at 1.5 µg mL−1 after 72 h [11]; and various cadinenes affect immobility-mortality (LC50 25.4–111.4 µg mL−1) and inhibit eggs hatching (IC50 31.23–56.71 µg mL−1) [12]. Identification of substances from botanical origins capable of controlling N. aberrans lies partially on differences with other common plant-parasitic nematodes (e.g., Meloidogyne spp.), such as its endoparasitic stage (migratory and sedentary), several infective stages, and 15–30 days dehydration tolerance of J3 and J4 larvae [13]. This research identifies botanical sources of compounds with nematicidal or nematostatic characteristics by screening seven plant species for nematicidal activities against N. aberrans J2 individuals and phytotoxicity to tomato seeds. NMR analysis established the chemical nature of nematicidal extracts (EC50,48h ≤ 112.3 µg mL−1) as sterol, thiophene, flavonoid, and alkaloid-like compounds (Figure 1). Some of these compounds are known for their nematicidal or nematostatic activity against other nematode species.
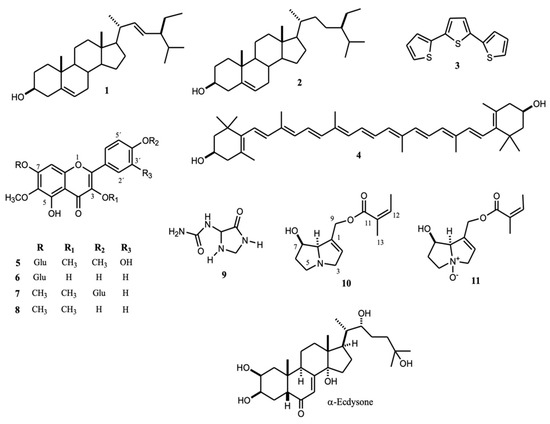
Figure 1.
Structure of compounds identified from nematicidal extracts against J2 individuals of N. aberrans (1–11) and α-ecdysone structure.
2. Results and Discussion
2.1. Nematostatic and Nematicidal Effect and EC50 of Extracts
From the nine extracts evaluated in this study (Table 1), G. mexicanum and A. integrifolium achieved the maximum paralysis effect (immobility = 85.3 ± 10% and 94.1 ± 3%, respectively) at 1000 µg mL−1 after 24 h. The remaining extracts showed their highest nematostatic effects after 36–60 h (immobility = 91.3%–100%). The immobility percentage after 72 h reached >95% for all extracts. Nematicidal (72 h *) and nematostatic effects (72 h) at 1000 µg mL−1 occurred at similar immobility percentages for A. aurantium (roots), A. integrifolium, A. subviscida, G. mexicanum, and H. terebinthinaceus, while the rest of the extracts showed their nematicidal effects at lower immobility percentages due to recovery of mobility (6.3–11%) (Table 1).

Table 1.
Effect of plant extracts at 1000 µg mL−1 on the immobility of N. aberrans J2s individual (100–150) after different exposure times.
Extracts at 100 µg mL−1 produced the highest immobility rates after 24 h (A. aurantium A), 36 h (A. integrifolium), 48 h (A. aurantium R), 60 h (A. cuspidata and G. mexicanum), and 72 h (A. subviscida, H. terebinthinaceus, and T. densiflora R and A) with immobility percentages ranging from 54.0 to 73.6 (Table 2). Lower concentration extracts (10 µg mL−1) produced immobility percentages <26% (Supplementary Table S1). Table 3 shows calculated EC50 values that cause 50% immobility in N. aberrans J2 individuals at 24 and 48 h. EC50 values for 24 h ranged from 53.5 to 468.1 µg mL−1 (ci, α = 0.05) and for 48 h varied from 31.5 to 299.8 µg mL−1. The most effective extracts (EC50,48h ≤ 112.3 µg mL−1) were A. aurantium R (62.3–88.3 µg mL−1), A. aurantium A (31.5–110.4 µg mL−1), A. integrifolium (47.4–107.1 µg mL−1), and T. densiflora R (59.0–112.3 µg mL−1). Chemical, non-fumigant nematicides acted as positive controls (Fluopyram and Abamectin) with EC50 values of 27.8–31.3 and 25.0–27.4 µg mL−1 at 24 and 48 h, respectively. Fluopyram is a succinate dehydrogenase inhibitor that blocks cellular respiration, while abamectin acts on glutamate-gated chloride channels, causing nematode death [14].
Table 2.
Effect of plant fractions and extracts at 100 µg mL−1 on the immobility of N. aberrans J2s individuals (100–150) after different exposure times.
Table 3.
Effective concentrations (50% immobility) at 24 and 48 h on N. aberrans.
2.2. Compounds Identified in A. aurantium and their Nematostatic Effects
Comparison of 1H and 13C NMR data allowed identification of stigmasterol (1) and β-sitosterol (2) [15,16]. Previously, we described the isolation and identification of α-terthienyl (3) and 5-(4″-hydroxy-1′-butynyl)-2–2′-bithiophene from A. aurantium roots [17]. In the present research, nematicidal extracts against N. aberrans contained 3.
Several treatments were tested at 50 and 100 µg mL−1: pure compounds 1 and 3 (Figure 1); a mixture of 1 and 2 (Mixt); α-terthienyl (aT); stigmasterol (St); and β-sitosterol (bS) (Figure 2). At 100 µg mL−1, 1, 3, and Mixt reached maximum inhibition at 72 h (93.3 ± 3%), 36 h (99.4 ± 0.6%), and 24 h (88.3 ± 8%), respectively. Commercial compounds, β-sitosterol, α-terthienyl, and stigmasterol showed lower nematostatic effects (50.6–80.7%), mainly during the first few hours of observation (12–36 h) (Figure 2). After 72 h, isolated and commercial β-stigmasterol showed 100 ± 0.0 and 94.5 ± 5.3% nematostatic effects, respectively. Treatments of isolated and commercial α-terthienyl showed the same immobility percentages (α = 0.01) after 72 h with values of 93.3 ± 3.1 and 90.6 ± 5%, respectively. Commercial α-terthienyl, stigmasterol, and β-sitosterol showed nematodes recovery values of 0.83, 0.61, and 0.61%, respectively (Table 4). Therefore, these compounds are nematostatic and nematicides at the same concentrations (α = 0.05).
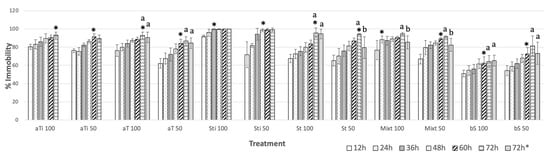
Figure 2.
Effect of compounds isolated α-terthienyl (aTi), stigmasterol (Sti), a mixture of 1 and 2 (Mixt), and commercial compounds: stigmasterol (St), α-terthienyl (aT) and β-sitosterol (bS) on the immobility of N. aberrans J2 individuals. * Maximum percentage of immobility. According to Tukey’s test, columns followed by the same letter are not significantly different (p < 0.05). Concentration 100 and 50 µg mL−1.
Table 4.
Effect of mixture stigmasterol/β-sitosterol, commercial and isolated compounds from A. aurantium roots on the immobility of N. aberrans J2 individuals after 72 h.
Thiophenes are common in the Tagetes genera [18,19]; this is a cover crop against plant-parasitic nematodes Meloidogyne and Pratilenchus (T. erecta, T. patula, T. tenuifolia, and T. minuta) [20]. Some thiophenes described as nematicide are 1-phenylhepta-1,3,5-triyne, and 5-phenyl-2-(1′-propynyl)-thiophene. These thiophenes, isolated from Coreopsis lanceolata L., effectively treated the pinewood nematode Bursaphelenchus xylophillus at 2 mM [21]. A commercial sample of α-terthienyl caused 100% mortality after 24 h at 0.125% against the infective larval stage of the cyst nematode Heterodera zeae [22]. In our experiments, α-terthienyl showed 83.2 ± 5.2% immobility after 24 h at 100 µg mL−1, or 0.01% against N. aberrans, while the commercial compound showed less effectiveness (71.74 ± 8.8%) (Figure 2). Sterols functioned on nematicidal activity as has been documented: β-sitosterol showed 60% mortality in M. incognita at 1% (after 12 h) [23] and together with stigmasterol at 5 µg mL−1 caused 74.4% and 55.3% mortality in M. incognita and Heterodera glycines, respectively [24]. In this work, isolated stigmasterol showed maximum activity after 36 h (99.4 ± 0.56%), while the commercial compound achieved it after 72 h (94.5 ± 5.3%). Synergy from impurities in the natural stigmasterol could account for the observed stronger activity. The impurities were β-sitosterol and a very similar compound but with 2 oxygen atoms, which was not identified. Similar results caused by β-sitosterol were observed with increased immobility from 68.7 ± 8.5% to 75.8 ± 12.5% when stigmasterol is present (Figure 2). A recent report about synergistic effects discusses nematicidal activity against Meloidogyne incognita of a mixture of 23a-homostigmast-5-en-3β-ol and nonacosan-10-ol. The mixture, at 50 µg mL−1, showed 93.7% mortality after 24 h, while the compounds individually, at 100 µg mL−1, exerted 50% mortality (24 h) [25,26]. The sterol kill mechanism on nematodes may disrupt steroid metabolism as stigmasterol (1) possesses a chemical similarity to α-ecdysone (Figure 1). α-ecdysone is involved in the biosynthesis and metabolism of molting and sex hormones of nematodes [27]. Such a function took part in accelerating the development of M. incognita by applying an ecdysone derivate (0.5 mM) on tomato seeds [28].
2.3. Compounds Identified in A. integrifolium
Structural identification of 1, 2, 4–8 was made by comparing their 1H and 13C NMR data with those described [15,16,29,30,31,32,33,34]. Identification of 5–8 required HSQC and HMBC experiments to confirm the structures and assign 1H and 13C signals (Supplementary Table S2).
Previous studies of A. integrifolium include the presence of an acetylene compound from stems [35]. Additionally, methanol extracts from stems showed inhibition of a topoisomerase enzyme (JN394, −81.19 ± 2.12% and JN362a, 126.06 ± 12.02%), and no significant antioxidant (AE = 4.18%) nor antimicrobial activities (MIC ≥ 6250 µg mL−1) [36]. In this research, the identification of quercetagetin derivatives 5–8 in A. integrifolium matches the genus’s phytochemistry (also named Calea genus) [37]. Previous studies have shown the nematicidal potential of flavonoids. For example, rutin exhibited the same mortality percentage as the positive control (carbofuran): 100% mortality at a 0.5% concentration (24 h) against the cyst nematode Heterodera zeae. Moreover, quercetin showed 50% mortality, and patuletin 7-β-O-glycoside showed no significant activity (20%, 24 h, light) [22]. However, this compound exerts moderate activity against Meloidogyne incognita J2 individuals (LC50,48h = 0.506%), while kaempferol, isorhamnetin, rutin, myricetin, and fisetin, among others, exhibited an LC50,48h value similar to carbofuran (LC50,48h = 0.0506%) [38]. Nematicidal activity of flavonoids may be due to acetylcholinesterase inhibition (AChE) because nematodes possess some neurotransmitters common in mammals like acetylcholine, serotonin, or glutamate [39]. Flavonoids as quercetin, genistein, and luteolin 7-O-glycoside inhibited AChE by 76.2, 65.7, and 54.9%, respectively [40], while a methoxylated quercetagetin showed a minor effect (inhibition 23.73 ± 1.94%) [41]. However, a glycosylated derivative, quercetagetin-7-O-(6-O-caffeoyl-β-D-glucopyranoside, possessed significant activity (IC50 12.54 ± 0.50 µg mL−1) against AChE isolated from Caenorhabditis elegans and Spodoptera litura. The IC50 value found came close to the value for chloropyrifos (2.32 ± 0.06 µg mL−1) [42]. We hypothesize flavonoid (5–8) isolated from A. integrifolium exerted their effects on the cholinergic nervous system of N. aberrans J2 individuals, while sterols (triterpenes) (1, 2) acted on the hormonal system. Therefore, the nematostatic (EC50,48h = 47.4–107.1 µg mL−1) and nematicidal (at 1000 µg mL−1, 99.4 ± 1%) effects observed could be the synergetic interaction between sterols and flavonoids. Synergic effects have been described on triterpenes (as saponins) and polyphenols combinations, which improved nematicidal activity in vivo against various nematode species (Meloidogyne, Xiphinema, Tylenchorhynchus, Criconemoides, and Pratylenchus) [43].
2.4. Compounds Identified in T. densiflora
Studies of the chemical composition of extracts from the Tournefortia genera revealed phenolic compounds [44] and alkaloids [45]. We identified allantoin (9) and pyrrolizidine alkaloids 10 and 11 (PAs), initially identified in the methanolic extract from its NMR data. The 13C NMR spectrum showed signals at δ 96.70, 96.28, and 96.21 (Figure 3) possibly related to C-8 of N-oxide PAs [46,47] and signals at δ 76.74 and 76.66 probably due to C-8 of PAs [48]. The detection of PAs in T. densiflora roots required alkaloid extraction and chromatographic separation to identify 10 and 11. 1H and 13C NMR data comparison with similar data from retronecine and PAs N-oxides [48,49,50,51], as well as HMQC and HMBC experiments, identified 9-O-angeloyl-retronecine N-oxide (11). The DEPTQ spectrum revealed the following functional groups: two CH3, four CH2, four CH, and three quaternary carbon atoms. Two double bonds were observed from signals at δ 132.9 (C), 121.4 (CH), 127.6 (C), and 139.0 (CH), while an angeloyl ester moiety was deduced from the 13C signal at δ 167.1 and the 1H NMR signal at δ 6.16 as qq (J = 1.6, 8.4Hz, H-12), which showed HMBC correlation with signals at δ 20.1 (C-12) and 16.0 (C-13). The signal at δ 95.5, caused by CH (C-8) next to quaternary nitrogen (N-oxide), and the signals at δ 34.8 (C-6), 68.5 (C-5), 77.8 (C-3), 60.9 (C-9), and 69.8 (C-7) were consistent with a necine base (as N-oxide) carrying a double-bond between C1 (132.9) and C2 (121.4). HMBC correlations support this proposal, and the presumed position of the angeloyl group at C-9, as the correlation of H-9 (δ 4.78 and 4.72) with the ester carbon C-10 (δ 167.1) showed (Table 5). Compound 10 displayed a 13C NMR spectrum very similar to 11. The main differences lay in the chemical shifts of C-3, C-5, and C-8 (atoms near the quaternary nitrogen), which appear in 10 at 60.4, 53.4, and 77.8 ppm, respectively. In 11, these signals shifted to higher frequencies at δ 77.8 (C-3), 68.5 (C-5), and 95.5 (C-8) ppm (Table 5).
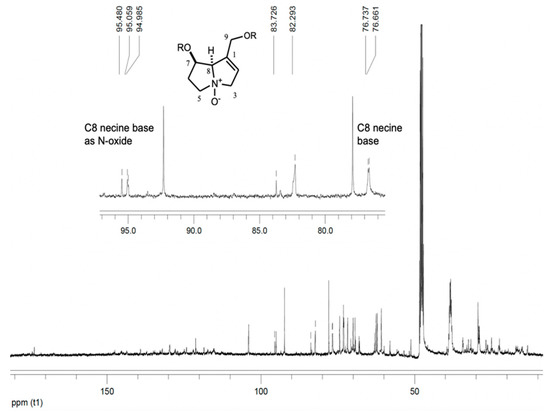
Figure 3.
13C NMR spectrum of methanol extract of T. densiflora roots. 100 MHz, CD3OD.
Table 5.
1H (400 MHz) and 13C NMR (100 MHz) data for 9 (MeOD), 10 and 11 (DMSO-d6).
Previous research demonstrated toxicity to Saccharomyces cerevisae from methanolic extracts from roots (70 mgmL−1) [52] and mycelial growth inhibition of Alternaria alternata (69.07 ± 2.0%) and Fusarium solani (52.42 ± 2.0%) [46]. Pyrrolizidine alkaloids (PAs) play a defensive role in the plant as antifeedants against herbivores [53]. Research shows PAs and PAs N-oxides are nematicides against Meloidogyne incognita, Heterodera schachtii, Pratylenchus penetrans, Plasmarhabditis hermaphrodita, and Rhabditis sp. [54]. Additionally, allantoin (9) possesses nematicidal activity (51.3% mortality) in Meloidogyne incognita and Heterodera glycines at 5 µg mL−1 [24]. These toxic activities and common occurrence of PAs in the Boraginaceae family [45] confirmed that PAs and allantoin are responsible for the toxic effects of the T. densiflora root extract observed on N. aberrans J2 individuals. PAs toxicity relies on their oxidation to pyrrolizinium ions by the cytochrome-P450 enzyme in the nematode. These ions are highly reactive as electrophiles and react with DNA, proteins, and other important macromolecules [55].
2.5. Phytotoxicity Test
Most nematicidal extracts against N. aberrans extracts showed 100% inhibition (−100%) of L. esculentum radicle growth at 20 µg mL−1, except extracts from H. terenbinthinaceus (−37%) (Figure 4). T. densiflora R extracts were the most phytotoxic with 40 and 38% inhibition at 0.02 µg mL−1 (Figure 4). Potentially, these extracts could act as soil disinfection agents. Also, A. cuspidata, A. subviscida, and T. densiflora A extracts showed hormetic effects: a 20 µg mL−1 solution inhibited radicle growth inhibition while a 0.02 µg mL−1 solution promoted it.
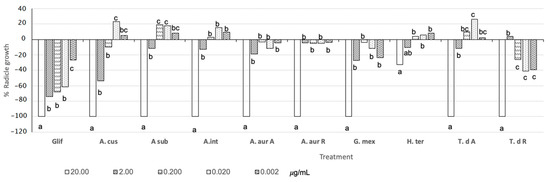
Figure 4.
Phytotoxic activity on L. esculentum of plant extracts.
Nematostatic and nematicidal effects observed by treatments with EC50,48h < 113 µg mL−1 relate to secondary metabolites like sterols, flavonoids, thiophenes, or alkaloids (PAs and allantoin), possibly biosynthesized in plants as a stress response. In general, sterols are involved in plants’ growth and fertility as hormonal precursors and cell membranes’ functional components. Nematodes also need sterols for their survival, but they cannot biosynthesize them de novo, so the nematodes readily absorb sterols. For example, Meloidogyne arenaria, M. incognita, and Pratilenchus agilis incorporate and transform sterols into necessary derivatives in their growth and reproduction [56]. Thus, nematodes should elicit a biological response to some sterols. Also, plant–nematode interactions require flavonoids and might be required for nematode reproduction. However, some flavonoids with specific structural arrangements have shown toxic effects on specific targets such as enzymes. Finally, thiophenes could inhibit enzymes like superoxide dismutase [57] and damage DNA [58]. The transformation of secondary metabolites to more toxic compounds also happened with PAs, as mentioned before.
3. Materials and Methods
3.1. General Experimental Procedures
NMR measurements were carried out on Bruker ASCENDTM 400 (400 MHz proton frequency) spectrometer (Bruker, Germany) at 298 K using 5 mm probes at 22 °C from CD3OD or DMSOd6 solutions. Chemical shifts (δ = ppm) were referenced to 2.50 (1H) and 39.43 (13C) ppm (DMSOd6) or to 3.30 (1H) and 36.067 (13C) ppm (CD3OD). Coupling constants are given in Hz. Signals are described as s (singlet), d (double), t (triple), and q (quartet).
3.2. Chemicals
All reagents and solvents (ACS grade), LiChroprep RP-18, and SiO2 supports for column and plate chromatography were obtained from Merck (MA, USA). Amberlite XAD16, α-terthienyl, β-sitosterol, stigmasterol, deuterated solvents, and dimethyl sulfoxide (DMSO-Hybri-Max) were obtained from Sigma Chemical (St. Louis, MO, USA).
3.3. Plant Species
The plant species were collected in Oaxaca, Mexico (See Table 6), and voucher specimens were deposited in the Herbarium of Forest Sciences, Universidad Autonoma de Chapingo, Texcoco (Estado de México, México). The scientific name, collection site, voucher number, plant part used, and extraction solvent are listed in Table 6.
Table 6.
Plants used in experiments.
3.4. Preparation of Extracts
Extracts were prepared according to procedures previously described [52]. All extracts were kept at 4 °C and protected from light and moisture until further use.
3.5. Isolation of 1–3 from A. aurantium Extract
The methanol extract (0.582 g) from aerial parts of A. aurantium was subjected to column chromatography (CC) using n-hexane-ethyl acetate mixtures. The fractions eluted with an 8:2 mixture, were re-chromatographed on CC with n-hexane-ethyl acetate (95:5) to yield 63.2 mg (10.8%), 30 mg (5.15%), and 3.02 mg (0.52%) of stigmasterol (1) and a mixture of stigmasterol (1)/β-sitosterol (2), and α-terthienyl (3), respectively (Figure 1). The purity of stigmasterol and α-terthienyl was approximately 95 and 98%, respectively. Purity was approximate since 1H NMR spectra by comparison of integration areas of 1 and 3 with those corresponding to impurities. The methanol extract from roots (7.215 g) was dissolved in acetone, and the solution yielded a solid residue (0.347 g), which was subjected to CC using n-hexane-ethyl acetate mixtures to obtain 40 mg (0.55%) and 23.7 mg (0.32%) of 1 and 3 respectively.
3.6. Identification of Compounds from A. integrifolium
Methanol extract of A. integrifolium (23.1 g) was partitioned with ethyl acetate (3 times) to obtain 10.1 g of ethyl acetate soluble fraction (ESF) and 13 g of methanol soluble fraction (MSF). ESF was subjected to column chromatography (SiO2) and eluted with mixtures of AcOEt: n-hexanes to obtain a mixture of chlorophylls “a” and “b” (80 mg), and a dark solid (155.8 mg). The solid was re-chromatographed (SiO2) with the same eluents to obtain lutein (4, 9.3 mg) (Figure 1) and a mixture (27.1 mg) of stigmasterol (1) and β-sitosterol (2). MFS (13 g) was solubilized in water and supported on a column of Amberlite XAD16; after two washes with water, the compounds retained were eluted with methanol to obtain a residue (1.5 g) free from simple carbohydrates. The residue (1.0 g) was eluted in a chromatography column (C18) using methanol:water mixtures as eluent. Chromatographic separation yielded 72 mg of four quercetagetin derivatives in binaries mixtures; its approximate composition was calculated by integrating 1H NMR areas of their characteristic signals. These compounds were identified as centaurin (5, 28.1 mg), patuletin-7-β-O-glucoside (6, 1.7 mg), pendulin (7, 6.4 mg), and penduletin (8, 1.0 mg) (Figure 1) from the analysis of their NMR data (Supplementary Table S2).
3.7. Identification of Compounds from T. densiflora Roots
The methanol extract (1.14 g), previously defatted with n-hexane and AcOEt, was subjected to C18 column chromatography and eluted with water. The eluent was identified by its NMR data [59] as allantoin (9, 33 mg, Figure 1, Table 5). PAs extraction required 8 g of the methanolic extract to be stirred with 1 M HCl (44 mL, 20 min); the mixture was filtered, the filtrate adjusted to pH 10 (KOH 1 M), and extracted with ethyl acetate. Organic fraction (180 mg) was subjected to chromatographic column (CC-SiO2) and eluted with CHCl3: methanol mixtures to obtain 9-O-angeloyl-retronecine (10, 11.5 mg, approx. 80% purity) and their N-oxide (11, 12.1 mg, approx. 90% purity).
3.8. Screening of Nematicidal and Nematostatic Activities
3.8.1. Nematodes
Mature egg masses of N. aberrans were extracted from infected roots of tomato plants (Lycopersicum esculentum Mill., 1768 or Solanum lycopersicum), propagated at Colegio de Postgraduados, Montecillo, Texcoco, Mexico. Egg masses were gently washed with water to remove adhered soil and a NaOCl 0.53% solution until the gelatinous matrix dissolved. Then they were washed with distilled water on a mesh sieve (#400) and incubated in distilled water at 25 °C for 5 days. Emerging J2 individuals were used in all experiments.
3.8.2. Assay
Test solutions were prepared in DMSO with 0.5% Tween 20 at 10, 100, 1000 µg mL−1 for extracts, while concentrations at 100 µg mL−1 were used for compounds and fractions. Fluopyram 50% (Verango, Bayer) and abamectin 5.41% (Oregon 60C-FMC) at 5, 10, 15, 25, 30 and 50 µg mL−1 (dissolved in distilled water) were tested as positive control. Treatments (5 µL) and between 100 and 150 J2 individuals in 95 µL of water were added to 96-well plates (Falcon, USA) and incubated at 25 °C. DMSO with 0.5% Tween 20 (5 µL) in 95 µL of water was used as blank. Previously, non-effect on J2 mobility was shown at 24, 36, 48, 60, and 72 h with the solvents used (Supplementary Table S3). Percentages of J2 immobility were recorded after 12, 24, 36, 48, 60, and 72 h by counting mobile and immobile J2 individuals under a stereomicroscope at 240X. A nematode was considered immobile if the nematode failed to respond to stimulation with a needle. After that, the J2 individuals at 1000 µg mL−1 (extracts) and 100 µg mL−1 (commercial stigmasterol, α-terthienyl, and β-sitosterol) were washed on a 400-mesh filter with distilled water to remove the excess test substance (extracts and commercial compounds). The treatments were replaced with distilled water to allow a possible recovery of the J2 individuals after 24 h. If they remained immobile, they were assumed to be dead, and the effect was considered nematicide. If any J2 individual regained mobility, the effect was considered nematostatic (paralysis). All treatments (extracts, isolated and commercial compounds) and control were replicated five times, and the experiments were performed two times. The immobility percentage was calculated using the equation: i = 100 × (1 − nt/nc); where i = immobility percentage, nt = active J2 in the treatment, and nc = active J2 in the blank [60].
3.9. Phytotoxicity Assay
Experiments were conducted with L. esculentum F1 seeds var. Sheva according to the methodology described [61]. Prior to evaluation, all extracts were dissolved in a 0.5% DMSO/H2O solution at 20, 2.0, 0.2, 0.02, and 0.002 µg mL−1 concentrations to obtain solids-free solutions. Commercial herbicide (Glyphosate) was used as a positive control at the same concentrations, while 0.5% DMSO/H2O was used as blank (100% growth).
3.10. Statistical Analysis
All experimental data were subjected to an analysis of variance (ANOVA) using Statistica Pro (Stat Soft, Japan). Treatment means were tested with Tukey’s HSD multiple comparison test at 0.05% or 0.01% probability levels.
4. Conclusions
To our knowledge, our results show for the first time the nematicidal activity against N. aberrans from T. densiflora, A. integrifolium, and A. aurantium extracts. In this research, we identified several compounds present in the nematicidal extracts against J2 individuals of N. aberrans containing: (a) flavonoids (A. integrifolium); (b) triterpene-type compounds (A. aurantium, A. integrifolium), (c) thiophene-type compounds (A. aurantium) and (d) alkaloids (T. densiflora). We identify 5–8 and 9–10 from A. integrifolium and T. densiflora, respectively. Moreover, we described the phytotoxic effect of all extracts on tomato radicle growth. Further research of these plant extracts will allow us to identify more compounds responsible for the nematicidal activity and provide alternative nontoxic crop protection chemicals.
Supplementary Materials
The following are available online. Table S1: Effect of plant extracts at 10 µg mL−1 on the immobility of N. aberrans J2s individuals after different exposure times; Table S2: 1H and 13C data for compounds 5–8. 400 MHz, 100 MHz CD3OD; Table S3: Effect of DMSO with 0.5% Tween on immobility of N. aberrans J2s after different exposure times.
Author Contributions
Conceptualization, R.V.-A., I.C.d.P.-V., and B.H.-C.; Formal analysis, B.H.-C.; Funding acquisition, B.H.-C.; Investigation, R.V.-A., H.C.-S., M.d.R.G.-O., R.S.-C., C.V.-C., and K.I.L.-d.L.; Methodology, R.V.-A., H.C.-S., M.d.R.G.-O., N.F.S.-S., and R.S.-C.; Project administration, B.H.-C.; Resources, I.C.d.P.-V., N.F.S.-S., and R.S.-C.; Supervision, M.V.R.-M.; Validation, M.V.R.-M.; Visualization, I.C.d.P.-V.; Writing–original draft, R.V.-A., H.C.-S., and B.H.-C.; Writing–review & editing, M.V.R.-M., I.C.d.P.-V., and B.H.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CONACyT grant number 225188, SAGARPA-CONACyT grant number 2016-1-277609, and SEP-PROMEP/103.5/12/6525.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Raul Velasco Azorsa thanks CONACyT for Masters in Science Fellowship (No. 318148). C.V.-C. thanks Prodep-SEP for postdoctoral Fellowship. We thank M.Sc. Ernestina Cedillo-Portugal from Universidad Autónoma Chapingo, Gerardo A. Salazar-Chávez, and Jaime Jiménez-Ramirez from the Universidad Nacional Autónoma de México for the identification of the plant species studied.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are available from the corresponding author.
References
- Singh, S.K.; Hodda, M.; Ash, G.J. Plant-parasitic nematodes of potential phytosanitary importance, their main hosts and reported yield losses. EPPO Bull. 2013, 43, 334–374. [Google Scholar] [CrossRef]
- Flores-Camacho, R.; Manzanilla-López, R.H.; Cid del Prado-Vera, I.; Martínez-Garza, A. Control de Nacobbus aberrans (Thorne) Thorne y Allen con Pochonia chlamydosporia (Goddard) Gams y Zare. Rev. Mex. Fitopatol. 2007, 25, 26–34. [Google Scholar]
- Pérez-Rodríguez, I.; Franco-Navarro, F.; Cid del Prado-Vera, I.; Zavaleta-Mejía, E. Control de Nacobbus aberrans en chile ancho (Capsicum annuum L.) mediante el uso combinado de enmiendas orgánicas, hongos nematófagos y nematicidas. Nematropica 2011, 41, 122–129. [Google Scholar]
- Lax, P.; Marro, N.; Agaras, B.; Valverde, C.; Doucet, M.E.; Becerra, A. Biological control of the false root-knot nematode Nacobbus aberrans by Pseudomonas protegens under controlled conditions. Crop Prot. 2013, 52, 97–102. [Google Scholar] [CrossRef]
- Caccia, M.; Lax, P.; Doucet, M.E. Effect of entomopathogenic nematodes on the plants-parasitic nematode Nacobbus aberrans. Biol. Fertil. Soils 2013, 49, 105–109. [Google Scholar] [CrossRef]
- Vázquez-Sánchez, M.; Medina-Medrano, J.R.; Cortez-Madrigal, H.; Angoa-Pérez, M.V.; Muñoz-Ruíz, C.V.; Villar-Luna, E. Nematicidal activity of wild plant extracts against second-stage juveniles of Nacobbus aberrans. Nematropica 2018, 48, 136–144. [Google Scholar]
- Ntalli, N.; Kasiotis, K.M.; Baira, E.; Stamatis, C.L.; Machera, K. Nematicidal activity of Stevia rebaudiana (Bertoni) assisted by phytochemical analysis. Toxins 2020, 12, 319. [Google Scholar] [CrossRef] [PubMed]
- Ntalli, N.G.; Ozalexandridou, E.X.; Kasiotis, K.M.; Samara, M.; Golfinopoulos, S.K. Nematicidal activity and phytochemistry of Greek Lamiaceae species. Agronomy 2020, 10, 1119. [Google Scholar] [CrossRef]
- Zavaleta-Mejía, E.; Gómez, R.O. Effect of Tagetes erecta L.-Tomato (Lycopersicon esculentum Mill.) intercropping on some tomato pests. Fitopatología 1995, 30, 33–45. [Google Scholar]
- Mareggiani, G.; Zamuner, N.; Michetti, M.; Franzetti, D.; Collavino, M. Impact of natural extracts on target and non-target soil organisms. Bol. San. Veg. Plagas 2005, 31, 443–448. [Google Scholar]
- Godínez-Vidal, D.; Soto-Hernández, M.; Rocha-Sosa, M.; Lozoya-Gloria, E.; Rojas-Martínez, R.I.; Guevara-Olvera, L.; Zavaleta-Mejía, E. Contenido de capsidiol en raíces de chile CM-334 infectadas por Nacobbus aberrans y su efecto en juveniles del segundo estadio. Nematropica 2010, 40, 227–237. [Google Scholar]
- Rodríguez-Chávez, J.L.; Franco-Navarro, F.; Delgado, G. In vitro nematicidal activity of natural and semisynthetic cadinenes from Heterotheca inuloides against the plant-parasitic nematode Nacobbus aberrans (Tylenchida: Pratylenchidae). Pest Manag. Sci. 2019, 75, 1734–1742. [Google Scholar] [CrossRef]
- Cristóbal-Alejo, J.; Mora-Aguilera, G.; Manzanilla-Lopez, R.H.; Marbán-Méndoza, N.; Sánchez-Garcia, P.; Del Prado-Vera, I.C.; Evans, K. Epidemiology and integrated control of Nacobbus aberrans on tomato in Mexico. Nematology 2006, 8, 727–737. [Google Scholar] [CrossRef]
- Feist, E.; Kearn, J.; Gaihre, Y.; O’Connor, V.; Holden-Dye, L. The distinct profiles of the inhibitory effects of fluensulfone, abamectin, aldicarb and fluopyram on Globodera pallida hatching. Pestic. Biochem. Physiol. 2020, 165, 104541. [Google Scholar] [CrossRef]
- Forgo, P.; Kövér, K.E. Gradient enhanced selective experiments in the 1H NMR chemical shift assignment of the skeleton and side-chain resonances of stigmasterol, a phytosterol derivative. Steroids 2004, 69, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ododo, M.M.; Choudhury, M.K.; Dekebo, A.H. Structure elucidation of β-sitosterol with antibacterial activity from the root bark of Malva parviflora. SpringerPlus 2016, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Martínez, M.; Hernández-Ramírez, V.I.; Hernández-Carlos, B.; Chávez-Munguía, B.; Calderón-Oropeza, M.A.; Talamás-Rohana, P. Antiamoebic activity of Adenophyllum aurantium (L.) Strother and its effect on the actin cytoskeleton of Entamoeba histolytica. Front. Pharmacol. 2016, 7, 169. [Google Scholar] [CrossRef] [PubMed]
- Downum, K.R.; Keil, D.J.; Rodríguez, E. Distribution of acetylenic thiophenes in the pectidinae. Biochem. Syst. Ecol. 1985, 13, 109–113. [Google Scholar] [CrossRef]
- Marotti, I.; Marotti, M.; Piccaglia, R.; Nastri, A.; Grandi, S.; Dinelli, G. Thiophene occurrence in different Tagetes species: Agricultural biomasses as sources of biocidal substances. J. Sci. Food Agric. 2010, 90, 1210–1217. [Google Scholar] [CrossRef]
- Hooks, C.R.R.; Wang, K.H.; Ploeg, A.; McSorley, R. Using marigold (Tagetes spp.) as a cover crop to protect crops from plant-parasitic nematodes. Appl. Soil Ecol. 2010, 46, 307–320. [Google Scholar] [CrossRef]
- Kimura, Y.; Hiraoka, K.; Kawano, T.; Fujioka, S.; Shimada, A. Nematicidal activities of acetylene compounds from Coriopsis lanceolata L. Zeitschrift für Naturforschung C 2008, 63, 843–847. [Google Scholar] [CrossRef]
- Faizi, S.; Fayyaz, S.; Bano, S.; Yawar Iqbal, E.; Lubna, L.; Siddiqi, H.; Naz, A. Isolation of nematicidal compounds from Tagetes patula L. yellow flowers: Structure-activity relationship studies against cyst nematode Heterodera zeae infective stage larvae. J. Agric. Food Chem. 2011, 59, 9080–9093. [Google Scholar] [CrossRef] [PubMed]
- Ferheen, S.; Akhtar, M.; Ahmed, A.G.; Anwar, M.A.; Kalhoro, M.A.; Afza, N.; Malik, A. Nematicidal potential of the Galinsoga parviflora. Pak. J. Sci. Ind. Res. Ser. B Biol. Sci. 2011, 54, 83–87. [Google Scholar]
- Barbosa, L.C.A.; Barcelos, F.F.; Demuner, A.J.; Santos, M.A. Chemical constituents from Mucuma aterrima with activity against Meloidogyne incognita and Heterodera glycines. Nematropica 1999, 29, 81–88. [Google Scholar]
- Naz, I.; Khan, M.R. Nematicidal activity of nonacosane-10-ol and 23a-homostigmast-5-en-3β-ol isolated from the roots of Fumaria parviflora (Fumariaceae). J. Agric. Food Chem. 2013, 61, 5689–5695. [Google Scholar] [CrossRef]
- Naz, I.; Saifullah, S.; Palomares-Rius, J.E.; Ahmad, M.; Ali, A.; Rashid, M.U.; Bibi, F. Combined nematocidal effect of nonacosan-10-ol and 23a-homostigmast-5-en-3β-ol on Meloidogyne incognita (kofoid and white) chitwood. J. Anim. Plant Sci. 2016, 26, 1633–1640. [Google Scholar]
- Chitwood, D.J.; McClure, M.A.; Feldlaufer, M.F.; Lusby, W.R.; Oliver, T.E. Sterol composition and ecdysteroid content of eggs of the root-knot nematodes Meloidogyne incognita and M. arenaria. J. Nematol. 1987, 19, 352. [Google Scholar]
- Udalova, Z.V.; Zinov’eva, S.V.; Valis’eva, I.S.; Paseshnichenko, V.A. Correlation between the structure of plant steroids and their effects on phytoparasitic nematodes. Appl. Biochem. Microbiol. 2004, 40, 93–97. [Google Scholar] [CrossRef]
- Moss, G.P. Carbon-13 NMR spectra of carotenoids. Pure Appl. Chem. 1976, 47, 97–102. [Google Scholar] [CrossRef][Green Version]
- El-Raey, M.A.; Ibrahim, G.E.; Eldahshan, O.A. Lycopene and lutein; A review for their chemistry and medicinal uses. J. Pharmacogn. Phytochem. 2013, 2, 245–254. [Google Scholar]
- Chiang, Y.M.; Chuang, D.Y.; Wang, S.Y.; Kuo, Y.H.; Tsai, P.W.; Shyur, L.F. Metabolite profiling and chemopreventive bioactivity of plant extracts from Bidens pilosa. J. Ethnopharmacol. 2004, 95, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Tapia, A.; Theoduloz, C.; Rodríguez, J.; López, S.; Feresin, G.E. Free radical scavengers and antioxidants from Tagetes mendocina. Zeitschrift für Naturforschung C 2004, 59, 345–353. [Google Scholar] [CrossRef]
- Arisawa, M.; Hatashita, T.; Numata, Y.; Tanaka, M.; Sasaki, T. Cytotoxic principles from Chrysosplenium flagelliferum. Int. J. Pharmacogn. 1997, 35, 141–143. [Google Scholar] [CrossRef]
- Bai, N.; He, K.; Zhou, Z.; Lai, C.S.; Zhang, L.; Quan, Z.; Shao, X.; Pan, M.-H.; Ho, C.T. Flavonoids from Rabdosia rubescens exert anti-inflammatory and growth inhibitory effect against human leukemia HL-60 cells. Food Chem. 2010, 122, 831–835. [Google Scholar] [CrossRef]
- Bohlmann, F.; Zdero, C. C17-Acetylenverbindungen aus Calea integrifolia. Phytochemistry 1976, 15, 1177. [Google Scholar] [CrossRef]
- Lira-De León, K.I.; Herrera-Martínez, M.; Ramírez-Mares, M.V.; Hernández-Carlos, B. Evaluation of anticancer potential of eight vegetal species from the state of Oaxaca. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 61–73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lima, T.C.; de Jesus Souza, R.; da Silva, F.A.; Biavatti, M.W. The genus Calea L.: A review on traditional uses, phytochemistry and biology activities. Phytother. Res. 2018, 32, 769–795. [Google Scholar] [CrossRef]
- Bano, S.; Iqbal, E.Y.; Lubna; Zil-ur-Rehman, S.; Fayyaz, S.; Faizi, S. Nematicidal activity of flavonoids with structure activity relationship (SAR) studies against root knot nematode Meloidogyne incognita. Eur. J. Plant Pathol. 2020, 157, 299–309. [Google Scholar] [CrossRef]
- Isaac, R.E.; MacGregor, D.; Coates, D. Metabolism and inactivation of neurotransmitters in nematodes. Parasitology 1996, 113, S157–S173. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.; Kartal, M.; Tosun, F.; Şener, B. Screening of various phenolic acids and flavonoid derivatives for their anticholinesterase potential. Zeitschrift für Naturforschung C 2007, 62, 829–832. [Google Scholar] [CrossRef]
- Promchai, T.; Saesong, T.; Ingkaninan, K.; Laphookhieo, S.; Pyne, S.G.; Limtharakul, T. Acetylcholinesterase inhibitory activity of chemical constituents isolated from Miliusa thorelii. Phytochem. Lett. 2018, 23, 33–37. [Google Scholar] [CrossRef]
- Li, M.; Gao, X.; Lan, M.; Liao, X.; Su, F.; Fan, L.; Zhao, Y.; Hao, X.; Wu, G.; Ding, X. Inhibitory activities of flavonoids from Eupatorium adenophorum against acetylcholinesterase. Pestic. Biochem. Physiol. 2020, 170, 104701. [Google Scholar] [CrossRef] [PubMed]
- San Martín, R.; Magunacelaya, J.C. Control of plant-parasitic nematodes with extracts of Quillaja saponaria. Nematology 2005, 7, 577–585. [Google Scholar]
- Correia Da Silva, T.B.; Souza, V.K.T.; Da Silva, A.P.F.; Lyra-Lemos, R.P.; Conserva, L.M. Determination of the phenolic content and antioxidant potential of crude extracts and isolated compounds from leaves of Cordia multispicata and Tournefortia bicolor. Pharm. Biol. 2010, 48, 63–69. [Google Scholar] [CrossRef]
- El-Shazly, A.; Wink, M. Diversity of pyrrolizidine alkaloids in the Boraginaceae structures, distribution, and biological properties. Diversity 2014, 6, 188–282. [Google Scholar] [CrossRef]
- Ogihara, K.; Miyagi, Y.; Higa, M.; Yogi, S. Pyrrolizidine alkaloids from Messerschmidia argentea. Phytochemistry 1997, 44, 545–547. [Google Scholar] [CrossRef]
- Constantinidis, T.; Harvala, C.; Skaltsounis, A.L. Pyrrolizidine N-oxide alkaloids of Heliotropium hirsutissimum. Phytochemistry 1993, 32, 1335–1337. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Roitman, J.N.; Benson, M.; Lundin, R.E. 13C NMR spectroscopy of pyrrolizidine alkaloids. Phytochemistry 1982, 21, 439–443. [Google Scholar] [CrossRef]
- Hammouda, F.M.; Ismail, S.I.; Hassan, N.M.; Tawfiq, W.A.; Kamel, A. Pyrrolizidine alkaloids from Alkanna orientalis (L.) Boiss. Qatar Univ. Sci. J. 1992, 12, 80–82. [Google Scholar]
- Logie, C.G.; Grue, M.R.; Liddell, J.R. Proton NMR spectroscopy of pyrrolizidine alkaloids. Phytochemistry 1994, 37, 43–109. [Google Scholar] [CrossRef]
- Roeder, E. Carbon-13 NMR spectroscopy of pyrrolizidine alkaloids. Phytochemistry 1990, 29, 11–29. [Google Scholar] [CrossRef]
- Lira-De León, K.I.; Ramírez-Mares, M.V.; Sánchez-López, V.; Ramírez Lepe, M.; Salas-Coronado, R.; Santos-Sánchez, N.F.; Valadez-Blanco, R.; Hernández-Carlos, B. Effect of crude plant extracts from some Oaxacan flora on two deleterious fungal phytopathogens and extracts compatibility with a biofertilizer strain. Front. Microbiol. 2014, 5, 383. [Google Scholar] [CrossRef]
- van Dam, N.M.; Vuister, L.W.; Bergshoeff, C.; de Vos, H.; Van der Meijden, E.D. The “Raison D’être” of pyrrolizidine alkaloids in Cynoglossum officinale: Deterrent effects against generalist herbivores. J. Chem. Ecol. 1995, 21, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Thoden, T.C.; Boppré, M.; Hallmann, J. Effects of pyrrolizidine alkaloids on the performance of plant-parasitic and free-living nematodes. Pest Manag. Sci. 2009, 65, 823–830. [Google Scholar] [CrossRef]
- Schramm, S.; Köhler, N.; Rozhon, W. Pyrrolizidine alkaloids: Biosynthesis, biological activities and occurrence in crop plants. Molecules 2019, 24, 498. [Google Scholar] [CrossRef]
- Chitwood, D.J.; Lusby, W.R. Metabolism of plants sterol by Nematodes. Lipids 1991, 26, 619–626. [Google Scholar] [CrossRef]
- Nivsarkar, M.; Kumar, G.P.; Laloraya, M.; Laloraya, M.M. Superoxide dismutase in the anal gills of the mosquito larvae of Aedes aegypti: Its inhibition by α-terthienyl. Arch. Insect Biochem. Physiol. 1991, 16, 249–255. [Google Scholar] [CrossRef]
- MacRae, W.D.; Chan, G.F.Q.; Wat, C.K.; Towers, G.H.N.; Lam, J. Examination of naturally occurring polyacetylenes and α-terthienyl for their ability to induce cytogenetic damage. Experientia 1980, 36, 1096–1097. [Google Scholar] [CrossRef]
- Sripathi, S.K.; Gopal, P.; Lalitha, P. Allantoin from the leaves of Pisonia grandis R. Br. Int. J. Pharm. Life Sci. 2011, 2, 815–817. [Google Scholar]
- Argentieri, M.P.; D’Addabbo, T.; Tava, A.; Agostinelli, A.; Jurzysta, M.; Avato, P. Evaluation of nematicidal properties of saponins from Medicago spp. Eur. J. Plant Pathol. 2008, 120, 189–197. [Google Scholar] [CrossRef]
- Hernández-Carlos, B.; González-Coloma, A.; Orozco-Valencia, A.U.; Ramírez-Mares, M.V.; Andrés-Yeves, M.F.; Joseph-Nathan, P. Bioactive saponins from Microsechium helleri and Sycios bulbosus. Phytochemistry 2011, 72, 743–751. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).