
Health Potential of Clery Strawberries: Enzymatic Inhibition and Anti-Candida Activity Evaluation

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Enzyme Inhibition Activities
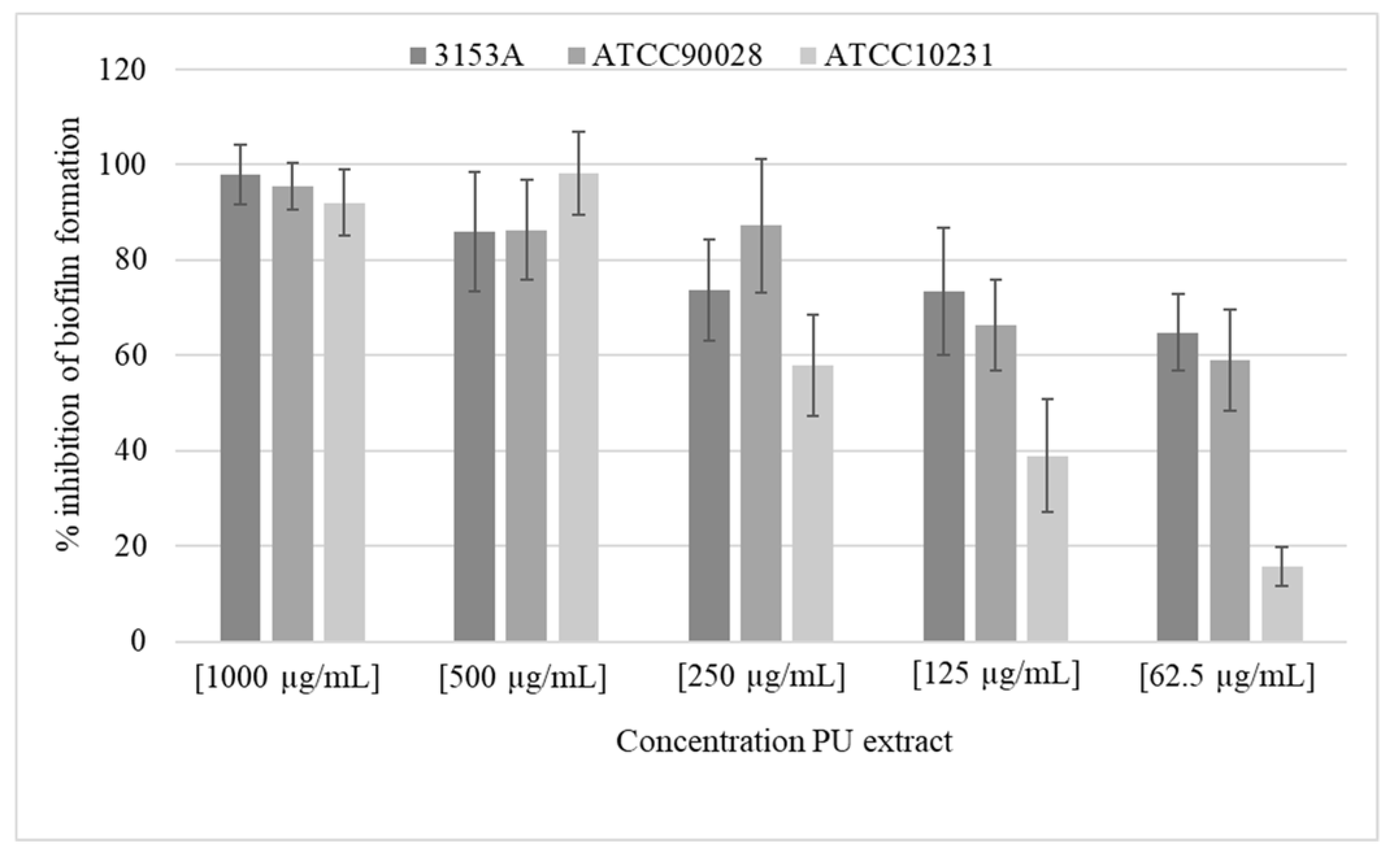
2.2. Antifungal Activity
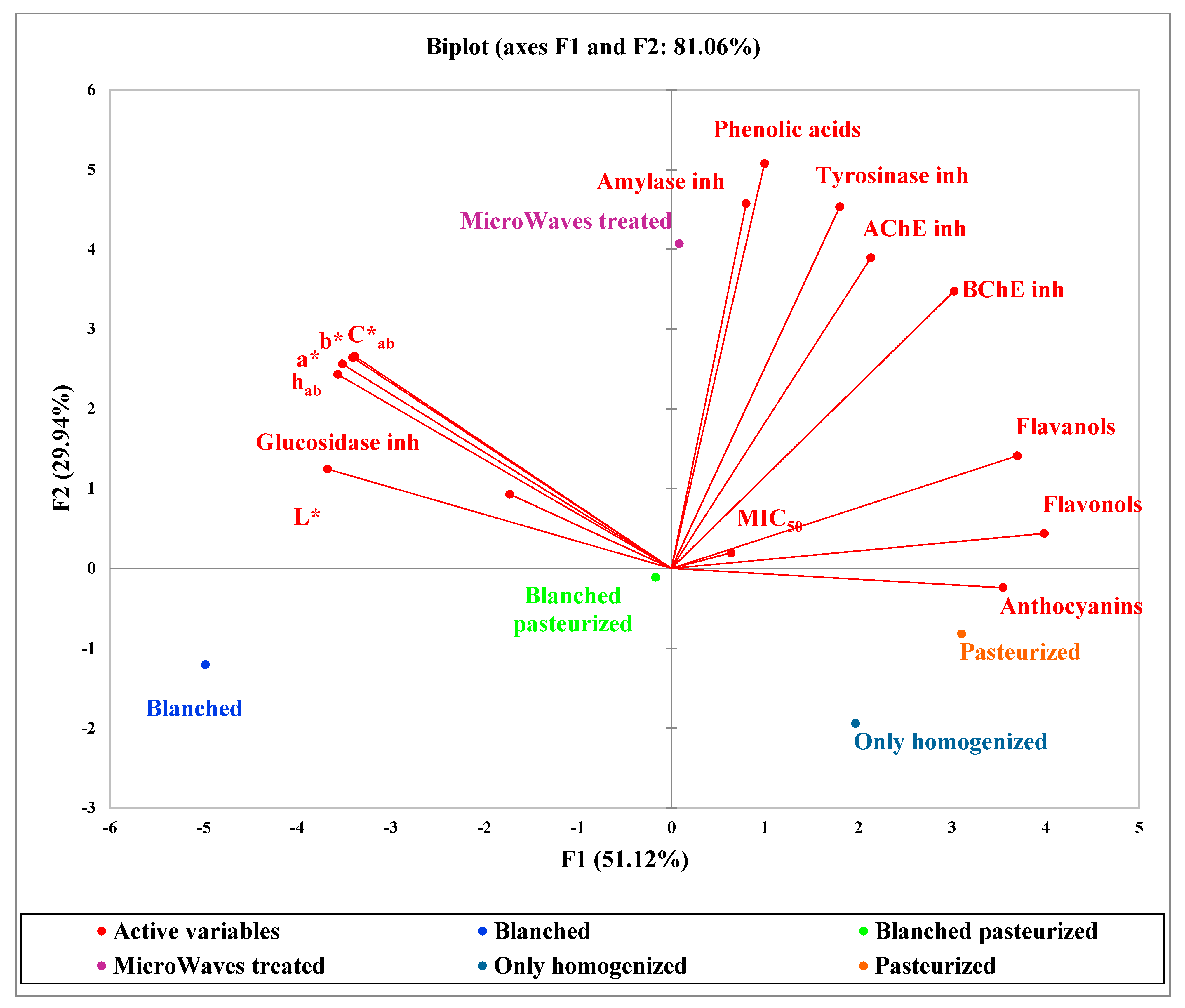
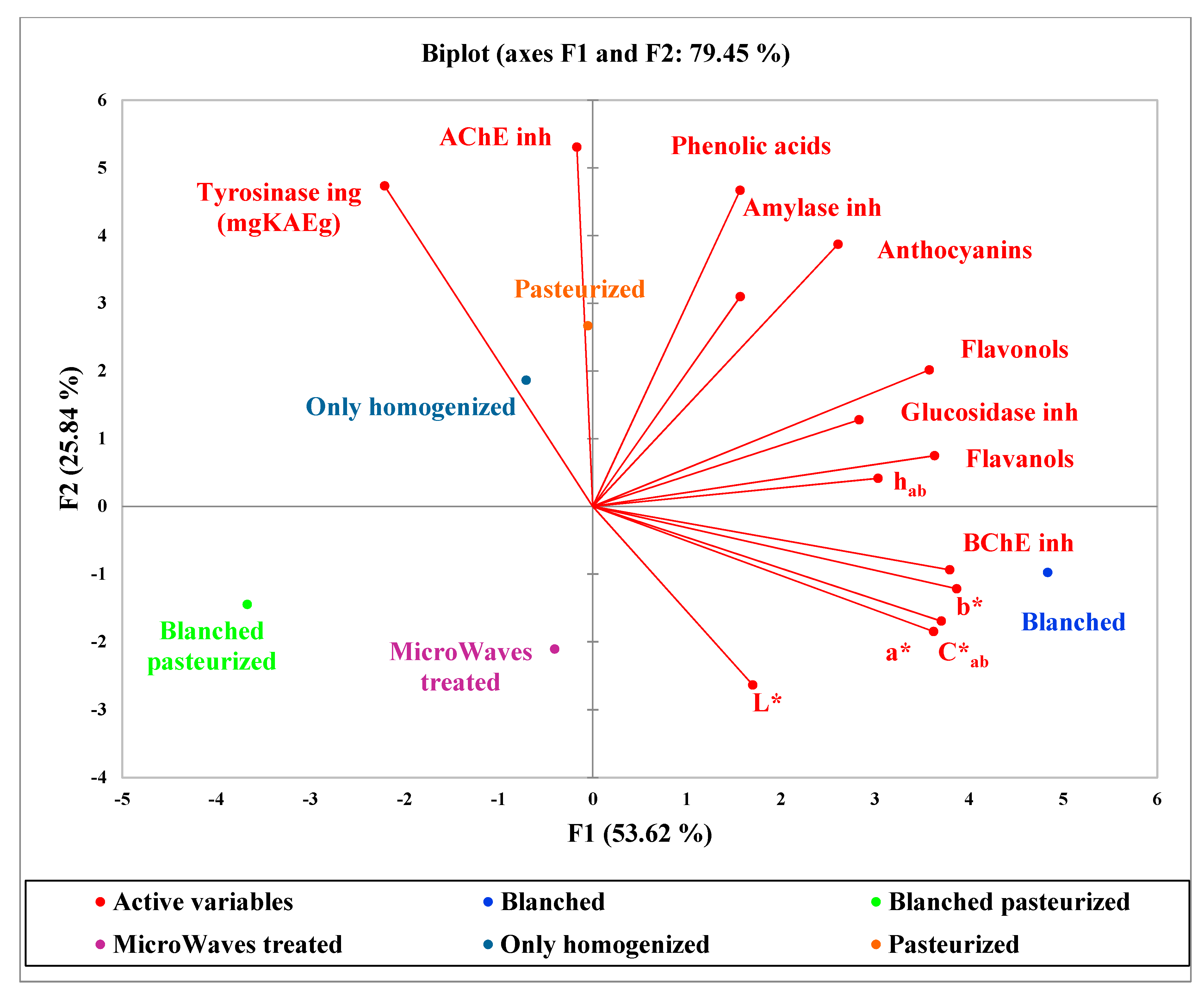
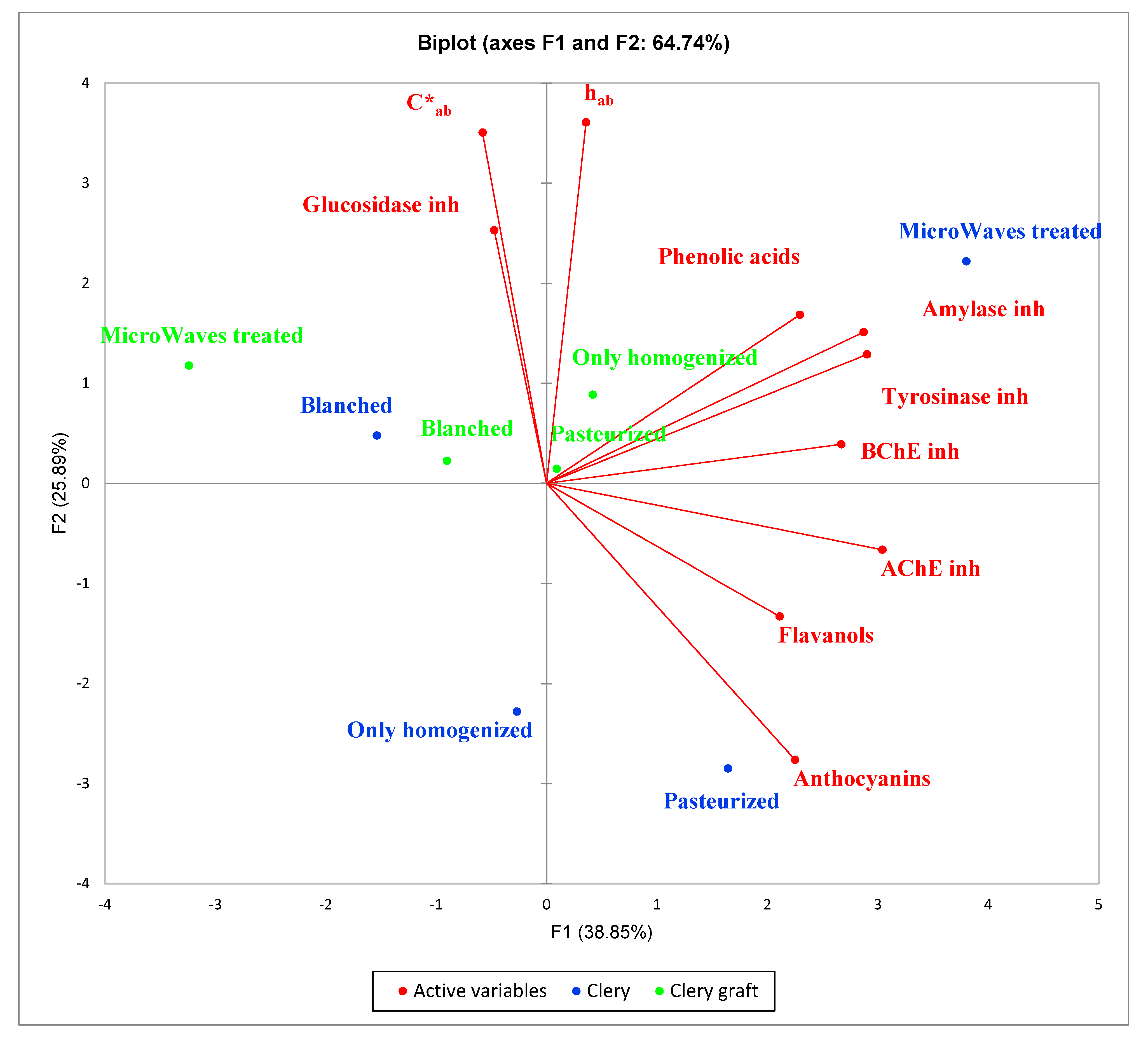
2.3. Principal Component Analysis (PCA)
3. Materials and Methods
3.1. Materials
3.2. Processing
3.3. Determination of Enzyme Inhibitory Effects
3.4. Antifungal Susceptibility Testing
3.4.1. Antifungal Susceptibility of C. albicans
3.4.2. In Vitro Activity of Compounds against C. albicans Biofilms
3.4.3. G. mellonella Survival Assay
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kovačević, D.B.; Brdar, D.; Fabečić, P.; Barba, F.J.; Lorenzo, J.M.; Putnik, P. Strategies to achieve a healthy and balanced diet: Fruits and vegetables as a natural source of bioactive compounds. In Agri-Food Industry Strategies for Healthy Diets and Sustainability; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 51–88. [Google Scholar]
- Garzoli, S.; Cairone, F.; Carradori, S.; Mocan, A.; Menghini, L.; Paolicelli, P.; Ak, G.; Zengin, G.; Cesa, S. Effects of Processing on Polyphenolic and Volatile Composition and Fruit Quality of Clery Strawberries. Antioxidants 2020, 9, 632. [Google Scholar] [CrossRef]
- Miller, K.; Feucht, W.; Schmid, M. Bioactive Compounds of Strawberry and Blueberry and Their Potential Health Effects Based on Human Intervention Studies: A Brief Overview. Nutrients 2019, 11, 1510. [Google Scholar] [CrossRef]
- Chen, T.; Shi, N.; Afzali, A. Chemopreventive Effects of Strawberry and Black Raspberry on Colorectal Cancer in Inflammatory Bowel Disease. Nutrients 2019, 11, 1261. [Google Scholar] [CrossRef]
- Koch, W. Dietary Polyphenols—Important Non-Nutrients in the Prevention of Chronic Noncommunicable Diseases. A Systematic Review. Nutrients 2019, 11, 1039. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. Gut Microbiome Dysbiosis and Immunometabolism: New Frontiers for Treatment of Metabolic Diseases. Mediat. Inflamm. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Temocico, G.; Fierascu, I.; Ortan, A.; Babeanu, N.E. Fragaria Genus: Chemical Composition and Biological Activities. Molecules 2020, 25, 498. [Google Scholar] [CrossRef]
- Nohynek, L.J.; Alakomi, H.-L.; Kähkönen, M.P.; Heinonen, M.; Helander, I.M.; Oksman-Caldentey, K.-M.; Puupponen-Pimiä, R.H. Berry Phenolics: Antimicrobial Properties and Mechanisms of Action Against Severe Human Pathogens. Nutr. Cancer 2006, 54, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M. Target Enzyme in Alzheimer’s Disease: Acetylcholinesterase Inhibitors. Curr. Top. Med. Chem. 2019, 19, 264–275. [Google Scholar] [CrossRef]
- Okello, E.J.; Mather, J. Comparative Kinetics of Acetyl- and Butyryl-Cholinesterase Inhibition by Green Tea Catechins|Relevance to the Symptomatic Treatment of Alzheimer’s Disease. Nutrients 2020, 12, 1090. [Google Scholar] [CrossRef] [PubMed]
- Taslimi, P. Evaluation of in vitro inhibitory effects of some natural compounds on tyrosinase activity and molecular docking study: Antimelanogenesis potential. J. Biochem. Mol. Toxicol. 2020, 34, e22566. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Giménez, J.; Cuadros, T.; Bové, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepulveda, M.; Peñuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Cardullo, N.; Muccilli, V.; Pulvirenti, L.; Cornu, A.; Pouységu, L.; Deffieux, D.; Quideau, S.; Tringali, C. C-glucosidic ellagitannins and galloylated glucoses as potential functional food ingredients with anti-diabetic properties: A study of α-glucosidase and α-amylase inhibition. Food Chem. 2020, 313, 126099. [Google Scholar] [CrossRef] [PubMed]
- Kato-Schwartz, C.G.; Corrêa, R.C.G.; Lima, D.D.S.; de Sá-Nakanishi, A.B.; Gonçalves, G.D.A.; Seixas, F.A.V.; Haminiuk, C.W.; Barros, L.; Ferreira, I.C.; Bracht, A.; et al. Potential anti-diabetic properties of Merlot grape pomace extract: An in vitro, in silico and in vivo study of α-amylase and α-glucosidase inhibition. Food Res. Int. 2020, 137, 109462. [Google Scholar] [CrossRef] [PubMed]
- Spínola, V.; Llorent-Martínez, E.J.; Castilho, P.C. Inhibition of α-amylase, α-glucosidase and pancreatic lipase by phenolic compounds of Rumex maderensis (Madeira sorrel). Influence of simulated gastrointestinal digestion on hyperglycaemia-related damage linked with aldose reductase activity and protein glycation. LWT 2020, 118, 108727. [Google Scholar] [CrossRef]
- Di Sotto, A.; Locatelli, M.; Macone, A.; Toniolo, C.; Cesa, S.; Carradori, S.; Eufemi, M.; Mazzanti, G.; Di Giacomo, S. Hypoglycemic, Antiglycation, and Cytoprotective Properties of a Phenol-Rich Extract from Waste Peel of Punica granatum L. var. Dente di Cavallo DC2. Molecules 2019, 24, 3103. [Google Scholar] [CrossRef]
- Li, Q.; Wang, C.; Tang, C.; He, Q.; Li, N.; Li, J. Dysbiosis of gut fungal microbiota is associated with mucosal inflammation in Crohn’s disease. J. Clin. Gastroenterol. 2014, 48, 513. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Y.; Shen, S.; Hou, Y.; Chen, Y.; Wang, T. The mycobiota of the human body: A spark can start a prairie fire. Gut Microbes 2020, 11, 655–679. [Google Scholar] [CrossRef]
- Pisa, D.; Alonso, R.; Juarranz, A.; Rábano, A.; Carrasco, L. Direct visualization of fungal infection in brains from patients with Alzheimer’s disease. J. Alzheimer’s Dis. 2015, 43, 613–624. [Google Scholar] [CrossRef]
- Alonso, R.; Pisa, D.; Rábano, A.; Rodal, I.; Carrasco, L. Cerebrospinal Fluid from Alzheimer’s Disease Patients Contains Fungal Proteins and DNA. J. Alzheimer’s Dis. 2015, 47, 873–876. [Google Scholar] [CrossRef]
- Pisa, D.; Alonso, R.; Rábano, A.; Rodal, I.; Carrasco, L. Different Brain Regions are Infected with Fungi in Alzheimer’s Disease. Sci. Rep. 2015, 5, 15015. [Google Scholar] [CrossRef]
- Wu, Y.; Du, S.; Johnson, J.L.; Tung, H.-Y.; Landers, C.T.; Liu, Y.; Seman, B.G.; Wheeler, R.T.; Costa-Mattioli, M.; Kheradmand, F.; et al. Microglia and amyloid precursor protein coordinate control of transient Candida cerebritis with memory deficits. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Witkowski, J.M.; Bourgade, K.; Khalil, A.; Zerif, E.; Larbi, A.; Hirokawa, K.; Pawelec, G.; Bocti, C.; Lacombe, G.; et al. Can an Infection Hypothesis Explain the Beta Amyloid Hypothesis of Alzheimer’s Disease? Front. Aging Neurosci. 2018, 10, 224. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; Da Pieve, S.; Butler, F. Impact of high pressure processing on total antioxidant activity, phenolic, ascorbic acid, anthocyanin content and colour of strawberry and blackberry purées. Innov. Food Sci. Emerg. Technol. 2009, 10, 308–313. [Google Scholar] [CrossRef]
- Karacam, C.H.; Sahin, S.; Oztop, M.H. Effect of high pressure homogenization (microfluidization) on the quality of Ottoman Strawberry (F. Ananassa) juice. LWT 2015, 64, 932–937. [Google Scholar] [CrossRef]
- Cesa, S.; Carradori, S.; Bellagamba, G.; Locatelli, M.; Casadei, M.A.; Masci, A.; Paolicelli, P. Evaluation of processing effects on anthocyanin content and colour modifications of blueberry (Vaccinium spp.) extracts: Comparison between HPLC-DAD and CIELAB analyses. Food Chem. 2017, 232, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Patsilinakos, A.; Ragno, R.; Carradori, S.; Petralito, S.; Cesa, S. Carotenoid content of Goji berries: CIELAB, HPLC-DAD analyses and quantitative correlation. Food Chem. 2018, 268, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, I.D. Galleria mellonellaas a model host to study virulence of Candida. Virulence 2014, 5, 237–239. [Google Scholar] [CrossRef]
- Arvanitis, M.; Glavis-Bloom, J.; Mylonakis, E. Invertebrate models of fungal infection. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 1378–1383. [Google Scholar] [CrossRef]
- Al Aboody, M.S.; Mickymaray, S. Anti-Fungal Efficacy and Mechanisms of Flavonoids. Antibiotics 2020, 9, 45. [Google Scholar] [CrossRef]
- Grussu, D.; Stewart, D.; McDougall, G.J. Berry Polyphenols Inhibit α-Amylase in Vitro: Identifying Active Components in Rowanberry and Raspberry. J. Agric. Food Chem. 2011, 59, 2324–2331. [Google Scholar] [CrossRef]
- Xiao, J.; Ni, X.; Kai, G.; Chen, X. A Review on Structure–Activity Relationship of Dietary Polyphenols Inhibiting α-Amylase. Crit. Rev. Food Sci. Nutr. 2013, 53, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and Enzyme Inhibitory Potential of Two Potentilla species (P. speciosa L. and P. reptans Willd.) and Their Chemical Composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Taff, H.T.; Nett, J.E.; Andes, D.R. Comparative analysis of Candidabiofilm quantitation assays. Sabouraudia 2012, 50, 214–218. [Google Scholar] [CrossRef]
- Li, D.-D.; Deng, L.; Hu, G.-H.; Zhao, L.-X.; Hu, D.-D.; Jiang, Y.-Y.; Wang, Y. Using Galleria mellonella–Candida albicans Infection Model to Evaluate Antifungal Agents. Biol. Pharm. Bull. 2013, 36, 1482–1487. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Treatment | Sample | Treatment |
|---|---|---|---|
| M | Homogenized with mixer | U | Homogenized with Ultraturrax® |
| M def | Defrosted and homogenized with mixer | U def | Defrosted and homogenized with Ultraturrax® |
| MP | Homogenized with mixer and pasteurized | UP | Homogenized with Ultraturrax® and pasteurized |
| BM | Blanched and homogenized with mixer | BU | Blanched and homogenized with Ultraturrax® |
| BMP | Blanched, homogenized with mixer and pasteurized | BUP | Blanched, homogenized with Ultraturrax® and pasteurized |
| WM | Treated with microwaves and homogenized with mixer | WU | Treated with microwaves and homogenized with Ultraturrax® |
| PM | Pasteurized and homogenized with mixer | PU | Pasteurized and homogenized with Ultraturrax® |
| PM def | Defrosted, pasteurized and homogenized with mixer | PU def | Defrosted, pasteurized and homogenized with Ultraturrax® |
| PM imm. | Immature fruits pasteurized and homogenized with mixer | PU imm. | Immature fruits pasteurized and homogenized with Ultraturrax® |
| M graft | Graft Clery/F. vesca homogenized with mixer | U graft | Graft Clery/F. vesca homogenized with Ultraturrax® |
| M def graft | Graft Clery/F. vesca defrosted and homogenized with mixer | U def graft | Graft Clery/F. vesca defrosted and homogenized with Ultraturrax® |
| MP graft | Graft Clery/F. vesca homogenized with mixer and pasteurized | UP graft | Graft Clery/F. vesca homogenized with Ultraturrax® and pasteurized |
| BM graft | Graft Clery/F. vesca defrosted and homogenized with mixer | BU graft | Graft Clery/F. vesca defrosted and homogenized with Ultraturrax® |
| WM graft | Graft Clery/F. vesca treated with microwaves and homogenized with mixer | WU graft | Graft Clery/F. vesca treated with microwaves and homogenized with Ultraturrax® |
| PM graft | Graft Clery/F. vesca pasteurized and homogenized with mixer | PU graft | Graft Clery/F. vesca pasteurized and homogenized with Ultraturrax® |
| BMP graft | Graft Clery/F. vesca defrosted and homogenized with mixer | BUP graft | Graft Clery/F. vesca defrosted and homogenized with Ultraturrax® |
| Sample | AChE Inhibition | BChE Inhibition | Tyrosinase Inhibition | α-Amylase Inhibition | α-Glucosidase Inhibition |
|---|---|---|---|---|---|
| M | 1.16 ± 0.01 jkl | 2.40 ± 0.33 ab | 1.92 ± 0.01 abcdefg | 12.84 ± 4.19 defg | 1.00 ± 0.01 d |
| U | 1.15 ± 0.01 kl | 2.23 ± 0.48 ab | 1.94 ± 0.01 abcde | 16.87 ± 2.28 abcde | 1.00 ± 0.01 d |
| M def | 1.40 ± 0.01 efg | 2.17 ± 0.27 ab | 1.79 ± 0.01 hijk | 17.02 ± 0.31 abcd | 1.00 ± 0.01 d |
| U def | 1.17 ± 0.01 jk | 2.17 ± 0.27 ab | 2.01 ± 0.02 a | 21.18 ± 0.13 a | 1.00 ± 0.01 d |
| MP | 1.23 ± 0.02 ijk | 2.37 ± 0.26 ab | 1.81 ± 0.05 ghijk | 10.65 ± 2.28 fg | ni |
| UP | 1.19 ± 0.01 jk | 2.27 ± 0.36 ab | 1.87 ± 0.04 cdefghi | 14.10 ± 2.66 cdef | 1.00 ± 0.01 d |
| BM | 1.25 ± 0.03 hij | 2.967 ± 0.43 a | 1.95 ± 0.02 abcd | 20.12 ± 0.17 ab | 1.00 ± 0.01 d |
| BU | 1.51 ± 0.05 d | 2.18 ± 0.07 ab | 1.97 ± 0.02 abc | 19.82 ± 0.81 ab | 1.01 ± 0.01 d |
| BMP | 1.21 ± 0.01 ijk | 2.13 ± 0.35 ab | 1.94 ± 0.09 abcde | 12.57 ± 0.92 defg | 1.00 ± 0.01 d |
| BUP | 1.25 ± 0.0 hij | 2.40 ± 0.38 ab | 1.83 ± 0.02 efghij | 15.54 ± 1.63 bcde | ni |
| WM | 1.07 ± 0.01 lm | 2.25 ± 0.45 ab | 1.92 ± 0.05 abcdefg | 12.10 ± 2.61 efg | 1.00 ± 0.01 d |
| WU | 1.03 ± 0.01 m | 1.99 ± 0.06 ab | 1.60 ± 0.02 l | 3.76 ± 0.05 h | 1.01 ± 0.01 d |
| PM | 1.16 ± 0.02 jkl | 2.06 ± 0.30 ab | 1.87 ± 0.02 cdefghi | 8.83 ± 2.09 g | ni |
| PU | 1.34 ± 0.02 gh | 2.07 ± 0.27 ab | 1.73 ± 0.02 jk | 14.25 ± 0.39 cdef | 1.00 ± 0.01 d |
| M graft | 1.14 ± 0.01 kl | 2.35 ± 0.20 ab | 1.77 ± 0.0 1ijk | 16.82 ± 0.56 abcde | 1.00 ± 0.01 d |
| U graft | 1.46 ± 0.01 def | 2.19 ± 0.32 ab | 1.86 ± 0.02 cdefghi | 18.32 ± 1.03 abc | 1.01 ± 0.01 d |
| M def graft | 1.66 ± 0.03 c | 2.62 ± 0.43 ab | 2.03 ± 0.05 a | 17.20 ± 0.57 abcd | 1.24 ± 0.08 b |
| U def graft | 1.37 ± 0.03 fg | 2.25 ± 0.05 ab | 2.00 ± 0.05 ab | 16.69 ± 2.79 abcde | 1.06 ± 0.01 d |
| MP graft | 1.16 ± 0.01 jkl | 2.97 ± 0.18 a | 1.71 ± 0.02 kl | 14.05 ± 1.46 cdef | 1.00 ± 0.01 d |
| UP graft | 1.25 ± 0.04 hij | 2.50 ± 0.33 ab | 1.81 ± 0.02 ghijk | 15.48 ± 0.57 bcde | 1.00 ± 0.01 d |
| BM graft | 1.66 ± 0.03 c | 2.15 ± 0.25 ab | 1.97 ± 0.05 abc | 16.13 ± 0.07 bcde | 1.00 ± 0.01 d |
| BU graft | 1.40 ± 0.05 efg | 1.99 ± 0.26 ab | 1.94 ± 0.03 abcdef | 16.30 ± 0.39 bcde | 1.00 ± 0.01 d |
| BMP graft | 1.31 ± 0.01 ghi | 2.61 ± 0.52 ab | 1.84 ± 0.03 defghij | 18.33 ± 0.33 abc | 1.00 ± 0.01 d |
| BUP graft | 1.79 ± 0.05 b | 2.22 ± 0.15 ab | 1.86 ± 0.02 cdefghi | 17.86 ± 1.48 abc | 1.44 ± 0.05 a |
| WM graft | 1.88 ± 0.02 ab | 2.60 ± 0.53 ab | 1.94 ± 0.09 abcde | 17.94 ± 1.54 abc | 1.01 ± 0.01 d |
| WU graft | 1.95 ± 0.05 a | 2.13 ± 0.53 ab | 1.89 ± 0.03 bcdefgh | 15.66 ± 0.23 bcde | 1.00 ± 0.01 d |
| PM graft | 1.21 ± 0.02 ijk | 1.81 ± 0.13 b | 1.82 ± 0.03 fghij | 15.81 ± 1.12 bcde | 1.02 ± 0.01 d |
| PU graft | 1.48 ± 0.04 de | 2.29 ± 0.10 ab | 1.81 ± 0.01 ghijk | 12.73 ± 0.34 defg | 1.14 ± 0.03 c |
| Galantamine | 0.003 ± 0.0001 n | 0.007 ± 0.0001 c | nt | nt | nt |
| Kojic acid | nt | nt | 0.08 ± 0.01 m | nt | nt |
| Acarbose | nt | nt | nt | 0.86 ± 0.01 h | 1.28 ± 0.04 b |
| Sample | Candida albicans | |||
|---|---|---|---|---|
| ATCC 90028 | ATCC10231 | ATCC24433 | 3153A | |
| Median MIC50 (μg/mL) | ||||
| M | 312.5 | 7.8 | 11.7 | 156.25 |
| U | 93.75 | 23.42 | 5.85 | 62.5 |
| MP | 257.8 | 7.8 | 3.9 | 62.5 |
| UP | 93.75 | 7.8 | 3.9 | 62.5 |
| BM | 156.25 | 7.8 | 5.85 | 62.5 |
| BU | 187.5 | 11.7 | 3.9 | 93.75 |
| BMP | 93.75 | 11.7 | 7.8 | 62.5 |
| BUP | 46.87 | 3.9 | 3.9 | 62.5 |
| WM | 312.5 | 7.8 | 5.85 | 62.5 |
| WU | 281.25 | 7.8 | 7.8 | 93.75 |
| PM | 312.5 | 11.7 | 11.7 | 156.25 |
| PU | 281.25 | 5.85 | 2.92 | 62.5 |
| PM def. | 375 | 46.87 | 5.85 | 31.25 |
| PU def. | 187.5 | 46.87 | 9.75 | 62.5 |
| PM imm. | 375 | 23.42 | 4.85 | 31.25 |
| PU imm. | 187.5 | 62.5 | 15.6 | 93.75 |
| Median MIC90 (μg/mL) | ||||
| M | >1000 | 62.5 | 23.42 | 156.25 |
| U | 1000 | 78.12 | 15.6 | 62.5 |
| MP | >1000 | 62.5 | 7.8 | 62.5 |
| UP | 750 | 62.5 | 7.8 | 62.5 |
| BM | 1000 | 187.5 | 7.8 | 62.5 |
| BU | 1000 | 93.75 | 23.42 | 93.75 |
| BMP | 1000 | 62.5 | 11.7 | 62.5 |
| BUP | 1000 | 93.75 | 7.8 | 62.5 |
| WM | >1000 | 187.5 | 15.6 | 62.5 |
| WU | 1000 | 62.5 | 19.52 | 93.75 |
| PM | >1000 | 62.5 | 23.42 | 281.25 |
| PU | >1000 | 250 | 7.8 | 62.5 |
| PM def. | >1000 | 93.75 | 7.8 | 31.25 |
| PU def. | 1000 | 187.5 | 15.6 | 93.75 |
| PM imm. | >1000 | 62.5 | 7.8 | 31.25 |
| PU imm. | 1000 | 187.5 | 31.25 | 125 |
| % of Survival Rate | |||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| Control larvae | 100% | 100% | 100% |
| Pierced larvae | 100% | 100% | 100% |
| PBS | 100% | 60% | 60% |
| PU 1000 µg/mL | 100% | 100% | 100% |
| C. albicans ATCC 10231 | 40% | 0 | 0 |
| C. albicans ATCC 10231 + PU 1000 µg/mL | 100% | 60% | 40% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cairone, F.; Simonetti, G.; Orekhova, A.; Casadei, M.A.; Zengin, G.; Cesa, S. Health Potential of Clery Strawberries: Enzymatic Inhibition and Anti-Candida Activity Evaluation. Molecules 2021, 26, 1731. https://doi.org/10.3390/molecules26061731
Cairone F, Simonetti G, Orekhova A, Casadei MA, Zengin G, Cesa S. Health Potential of Clery Strawberries: Enzymatic Inhibition and Anti-Candida Activity Evaluation. Molecules. 2021; 26(6):1731. https://doi.org/10.3390/molecules26061731
Chicago/Turabian StyleCairone, Francesco, Giovanna Simonetti, Anastasia Orekhova, Maria Antonietta Casadei, Gokhan Zengin, and Stefania Cesa. 2021. "Health Potential of Clery Strawberries: Enzymatic Inhibition and Anti-Candida Activity Evaluation" Molecules 26, no. 6: 1731. https://doi.org/10.3390/molecules26061731
APA StyleCairone, F., Simonetti, G., Orekhova, A., Casadei, M. A., Zengin, G., & Cesa, S. (2021). Health Potential of Clery Strawberries: Enzymatic Inhibition and Anti-Candida Activity Evaluation. Molecules, 26(6), 1731. https://doi.org/10.3390/molecules26061731