
Chemical-Genetic Interactions of Bacopa monnieri Constituents in Cells Deficient for the DNA Repair Endonuclease RAD1 Appear Linked to Vacuolar Disruption


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Mutation Frequency But Not Chromosome Instability Is Enhanced Following Exposure to B. monnieri Extracts
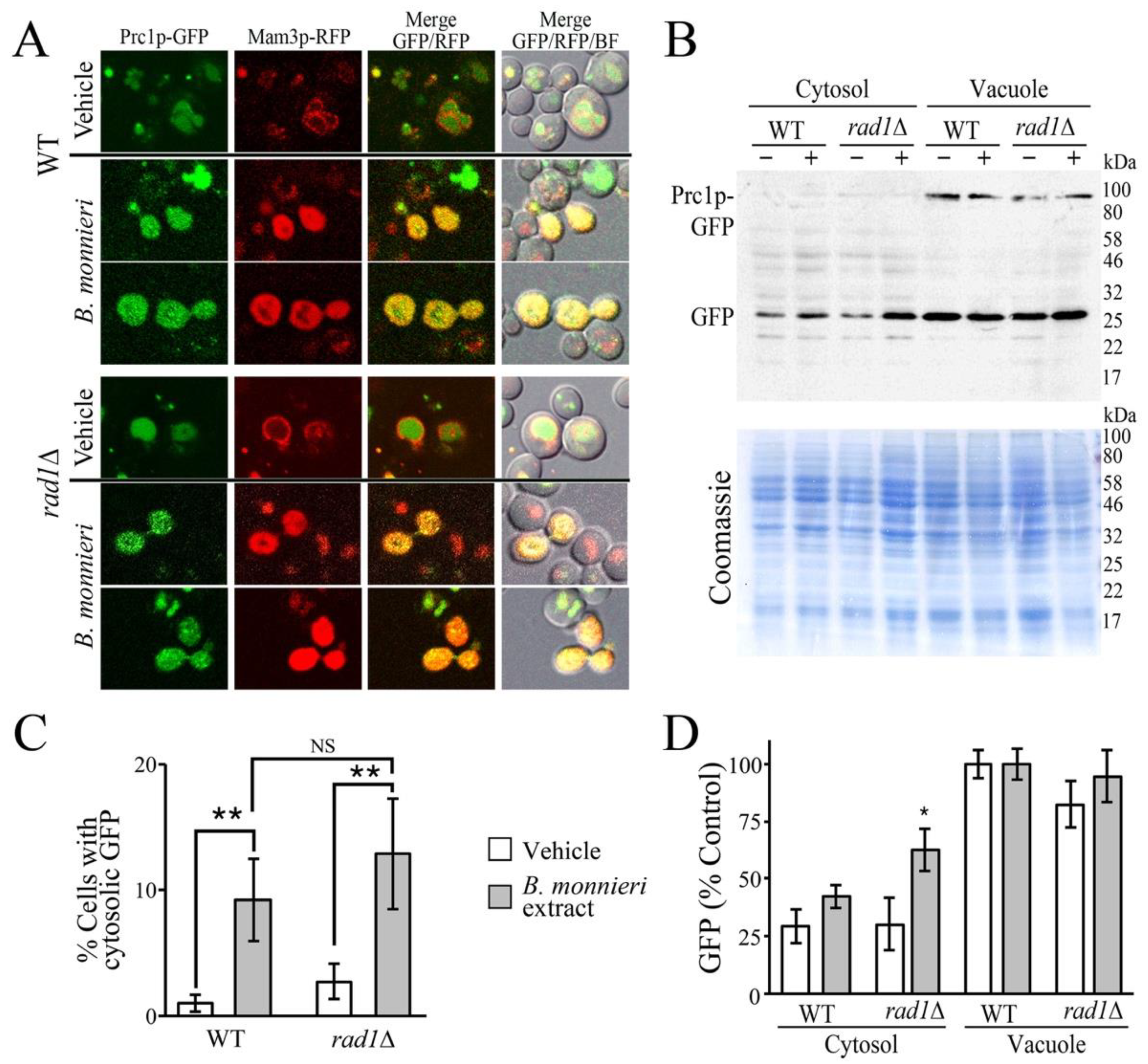
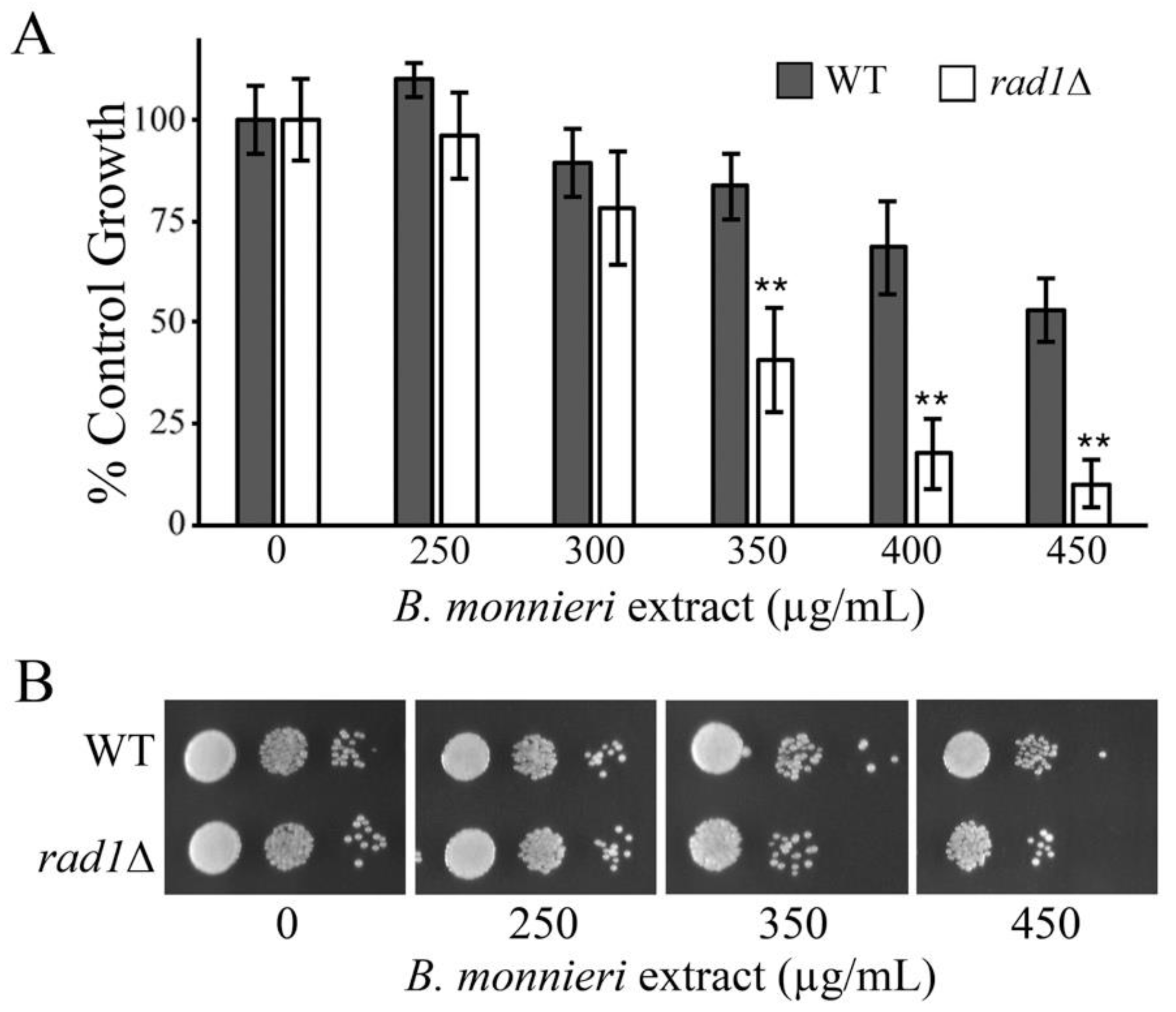
2.2. B. monnieri Extracts Promote Vacuole Damage
2.3. Vacuole Permeabilization with Chloroquine Does Not Mimic Effects of B. monnieri Extracts
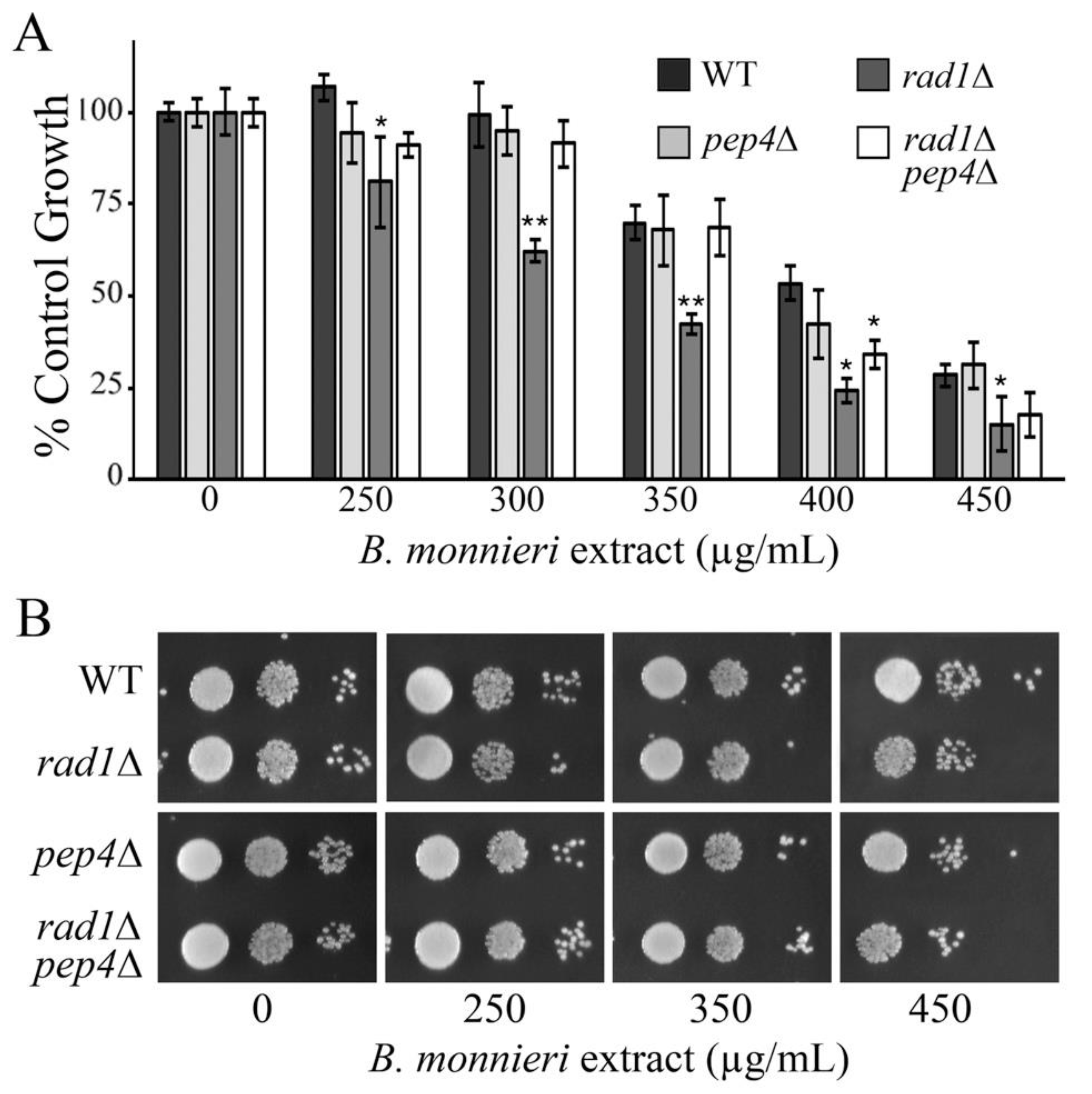
2.4. Deletion of PEP4 Limits Toxicity of B. monnieri Extracts in rad1∆ Yeast
2.5. Mitochondrial Structure Is Not Disrupted by Exposure to B. monnieri Extracts
2.6. Stabilization of Plasma Membrane Does Not Prevent Toxicity from B. monnieri Extracts
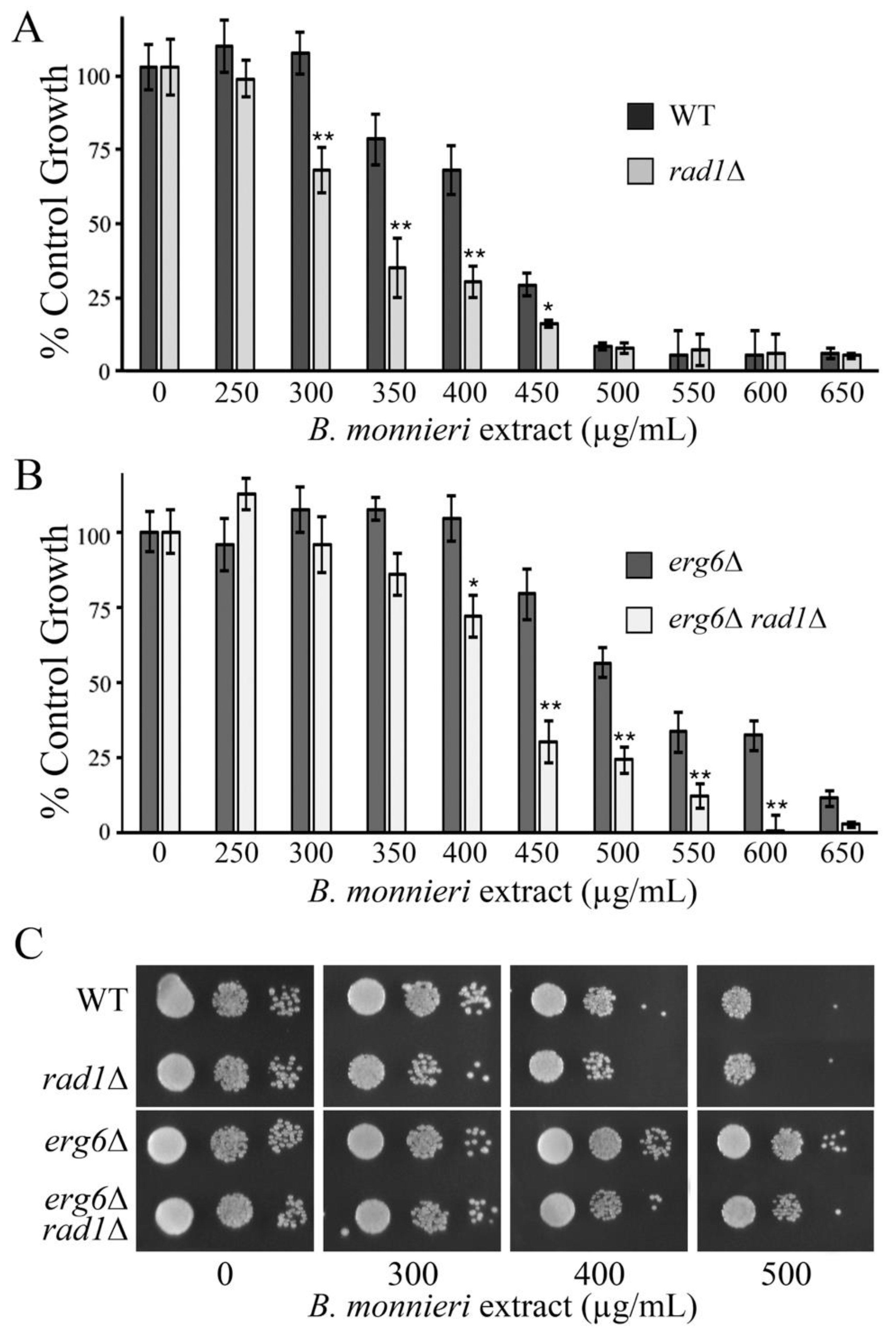
2.7. Toxicity of B. monnieri Extracts Is Enhanced in Yeast Competent for Sterol Production
2.8. The Yeast Meta-Caspase Yca1p Is Required for Toxicity of B. monnieri Extracts in rad1∆ Cells
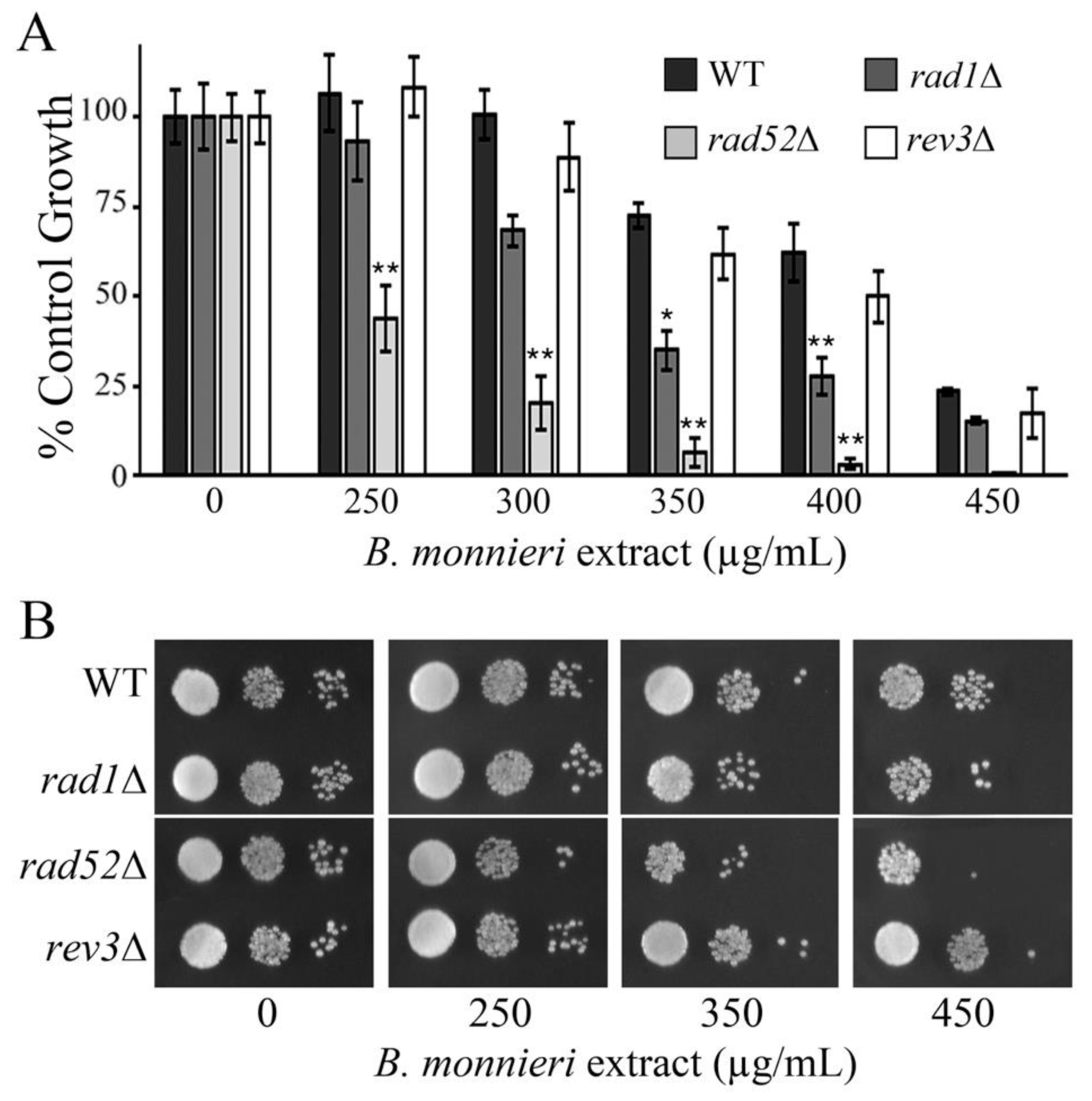
2.9. Yeast Deleted for RAD52 Exhibit Synthetic Lethal Interactions with B. monnieri Extracts
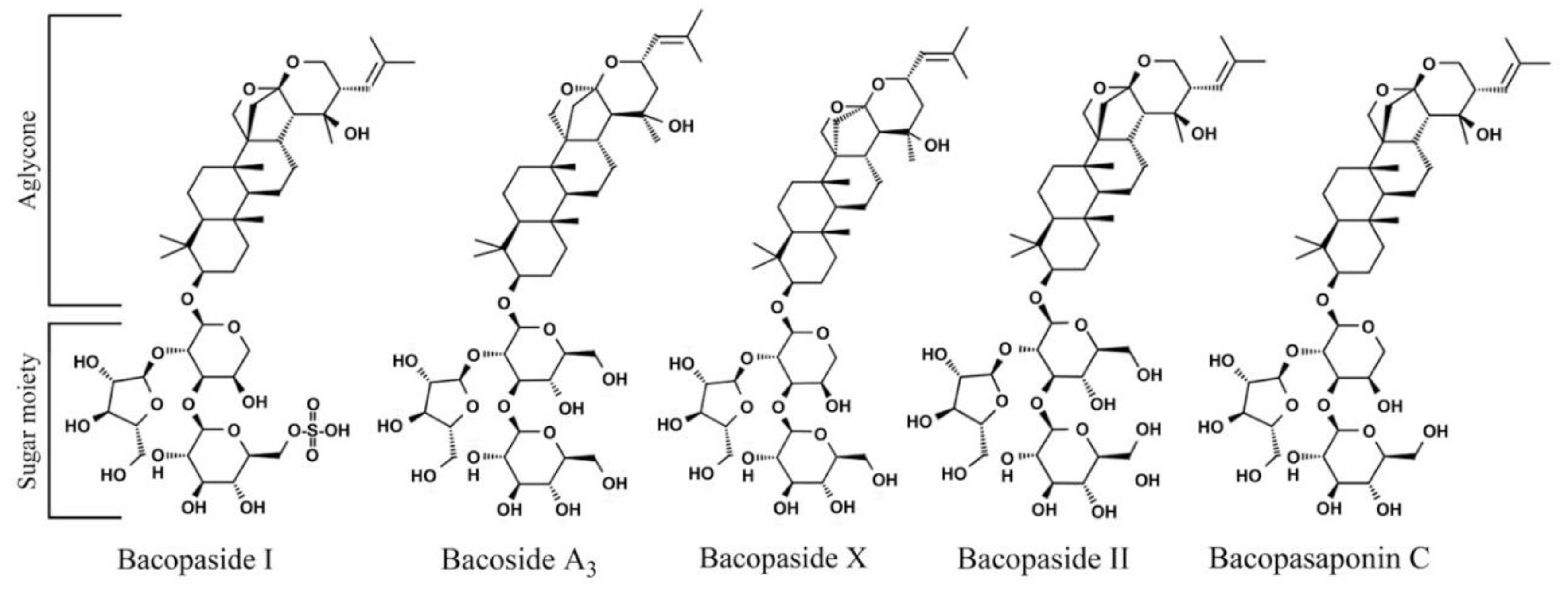
2.10. Bacopasaponin C Preferentially Limits the Growth of RAD1 Deleted Yeast
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Plasmids
4.2. Growth Tests
4.3. Analysis of Mutation Rates and Chromosome Instability
4.4. Fluorescence Imaging
4.5. Cellular Fractionation
4.6. B. monnieri Extracts and Pure Compounds
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Watson, A.J.; Collins, P.D. Colon cancer: A civilization disorder. Dig. Dis. 2011, 29, 222–228. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer Incidence and Mortality Worldwide; International Agency for Research on Cancer: Lyon, France, 2013. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef]
- Watanabe, T.; Itabashi, M.; Shimada, Y.; Tanaka, S.; Ito, Y.; Ajioka, Y.; Hamaguchi, T.; Hyodo, I.; Igarashi, M.; Ishida, H.; et al. Japanese Society for Cancer of the Colon and Rectum (JSCCR) guidelines 2010 for the treatment of colorectal cancer. Int. J. Clin. Oncol. 2012, 17, 1–29. [Google Scholar] [CrossRef] [PubMed]
- de Gramont, A.; Schmoll, H.J.; Cervantes, A.; Tournigand, C. The evolving role of oxaliplatin in the management of colorectal cancer. Colorectal Dis. 2003, 5, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Ling, W.; Fan, J.; Ma, Y.; Ma, Y.; Wang, H. Capecitabine-based chemotherapy for metastatic colorectal cancer. J. Cancer Res. Clin. Oncol. 2011, 137, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef]
- Raymond, E.; Faivre, S.; Woynarowski, J.M.; Chaney, S.G. Oxaliplatin: Mechanism of action and antineoplastic activity. Semin. Oncol. 1998, 25, 4–12. [Google Scholar]
- Teixeira, L.; Barry, S.; Debourdeau, P.; Cohen, A.; Tournigand, C. Cardiotoxicity of 5-fluorouracil. Bull. Cancer 2004, 91, 154–158. [Google Scholar]
- Ng, M.; Cunningham, D.; Norman, A.R. The frequency and pattern of cardiotoxicity observed with capecitabine used in conjunction with oxaliplatin in patients treated for advanced colorectal cancer (CRC). Eur. J. Cancer 2005, 41, 1542–1546. [Google Scholar] [CrossRef]
- Han, R.; Yang, Y.M.; Dietrich, J.; Luebke, A.; Mayer-Proschel, M.; Noble, M. Systemic 5-fluorouracil treatment causes a syndrome of delayed myelin destruction in the central nervous system. J. Biol. 2008, 7, 12. [Google Scholar] [CrossRef]
- McQuade, R.M.; Bornstein, J.C.; Nurgali, K. Anti-Colorectal Cancer Chemotherapy-Induced Diarrhoea: Current Treatments and Side-Effects. Int. J. Clin. Exp. Med. 2014, 5, 393–406. [Google Scholar] [CrossRef]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Curtin, N.J. DNA repair dysregulation from cancer driver to therapeutic target. Nat. Rev. Cancer 2012, 12, 801–817. [Google Scholar] [CrossRef] [PubMed]
- Brookman, K.W.; Lamerdin, J.E.; Thelen, M.P.; Hwang, M.; Reardon, J.T.; Sancar, A.; Zhou, Z.Q.; Walter, C.A.; Parris, C.N.; Thompson, L.H. ERCC4 (XPF) encodes a human nucleotide excision repair protein with eukaryotic recombination homologs. Mol. Cell. Biol. 1996, 16, 6553–6562. [Google Scholar] [CrossRef] [PubMed]
- Facista, A.; Nguyen, H.; Lewis, C.; Prasad, A.R.; Ramsey, L.; Zaitlin, B.; Nfonsam, V.; Krouse, R.S.; Bernstein, H.; Payne, C.M.; et al. Deficient expression of DNA repair enzymes in early progression to sporadic colon cancer. Genome Integr. 2012. [Google Scholar] [CrossRef]
- Ahmad, A.; Robinson, A.R.; Duensing, A.; van Drunen, E.; Beverloo, H.B.; Weisberg, D.B.; Hasty, P.; Hoeijmakers, J.H.; Niedernhofer, L.J. ERCC1-XPF endonuclease facilitates DNA double-strand break repair. Mol. Cell. Biol. 2008, 28, 5082–5092. [Google Scholar] [CrossRef]
- Bardwell, A.J.; Bardwell, L.; Tomkinson, A.E.; Friedberg, E.C. Specific cleavage of model recombination and repair intermediates by the yeast Rad1-Rad10 DNA endonuclease. Science 1994, 265, 2082–2085. [Google Scholar] [CrossRef]
- Aung, H.S.; Huangteerakul, C.; Jensen, A.N.; Sukrong, S.; Jensen, L.T. Interrogation of ethnomedicinal plants for synthetic lethality effects in combination with deficiency in the DNA repair endonuclease RAD1 using a yeast cell-based assay. J. Ethnopharmacol. 2018, 223, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Harwansh, R.K.; Bahadur, S.; Banerjee, S.; Kar, A.; Chanda, J.; Biswas, S.; Ahmmed, S.M.; Katiyar, C.K. Development of Ayurveda—Tradition to trend. J. Ethnopharmacol. 2017, 197, 10–24. [Google Scholar] [CrossRef]
- Pase, M.P.; Kean, J.; Sarris, J.; Neale, C.; Scholey, A.B.; Stough, C. The cognitive-enhancing effects of Bacopa monnieri: A systematic review of randomized, controlled human clinical trials. J. Altern. Complement. Med. 2012, 18, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Limpeanchob, N.; Jaipan, S.; Rattanakaruna, S.; Phrompittayarat, W.; Ingkaninan, K. Neuroprotective effect of Bacopa monnieri on beta-amyloid-induced cell death in primary cortical culture. J. Ethnopharmacol. 2008, 120, 112–117. [Google Scholar] [CrossRef]
- Rohini, G.; Devi, C.S. Bacopa monniera extract induces apoptosis in murine sarcoma cells (S-180). Phytother. Res. 2008, 22, 1595–1598. [Google Scholar] [CrossRef]
- Peng, L.; Zhou, Y.; Kong de, Y.; Zhang, W.D. Antitumor activities of dammarane triterpene saponins from Bacopa monniera. Phytother. Res. 2010, 24, 864–868. [Google Scholar] [CrossRef]
- Garai, S.; Mahato, S.B.; Ohtani, K.; Yamasaki, K. Dammarane-type triterpenoid saponins from Bacopa monniera. Phytochemistry 1996, 42, 815–820. [Google Scholar] [CrossRef]
- Chakravarty, A.K.; Garai, S.; Masuda, K.; Nakane, T.; Kawahara, N. Bacopasides III-V: Three new triterpenoid glycosides from Bacopa monniera. Chem. Pharm. Bull. (Tokyo) 2003, 51, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishna, C.; Rao, C.V.; Trimurtulu, G.; Vanisree, M.; Subbaraju, G.V. Triterpenoid glycosides from Bacopa monnieri. Phytochemistry 2005, 66, 2719–2728. [Google Scholar] [CrossRef]
- Rastogi, S.; Pal, R.; Kulshreshtha, D.K. Bacoside A3—A triterpenoid saponin from Bacopa monniera. Phytochemistry 1994, 36, 133–137. [Google Scholar] [CrossRef]
- Rohini, G.; Sabitha, K.E.; Devi, C.S. Bacopa monniera Linn. extract modulates antioxidant and marker enzyme status in fibrosarcoma bearing rats. Indian J. Exp. Biol. 2004, 42, 776–780. [Google Scholar] [PubMed]
- Janani, P.; Sivakumari, K.; Geetha, A.; Ravisankar, B.; Parthasarathy, C. Chemopreventive effect of bacoside A on N-nitrosodiethylamine-induced hepatocarcinogenesis in rats. J. Cancer Res. Clin. Oncol. 2010, 136, 759–770. [Google Scholar] [CrossRef]
- Mallick, M.N.; Akhtar, M.S.; Najm, M.Z.; Tamboli, E.T.; Ahmad, S.; Husain, S.A. Evaluation of anticancer potential of Bacopa monnieri L. against MCF-7 and MDA-MB 231 cell line. J. Pharm. Bioallied. Sci. 2015, 7, 325–328. [Google Scholar] [CrossRef]
- Xiao, W.; Chow, B.L. Synergism between yeast nucleotide and base excision repair pathways in the protection against DNA methylation damage. Curr. Genet. 1998, 33, 92–99. [Google Scholar] [CrossRef]
- Jones, E.W. Three proteolytic systems in the yeast saccharomyces cerevisiae. J. Biol. Chem. 1991, 266, 7963–7966. [Google Scholar] [CrossRef]
- Knop, M.; Schiffer, H.H.; Rupp, S.; Wolf, D.H. Vacuolar/lysosomal proteolysis: Proteases, substrates, mechanisms. Curr. Opin. Cell Biol. 1993, 5, 990–996. [Google Scholar] [CrossRef]
- Yamashima, T.; Kohda, Y.; Tsuchiya, K.; Ueno, T.; Yamashita, J.; Yoshioka, T.; Kominami, E. Inhibition of ischaemic hippocampal neuronal death in primates with cathepsin B inhibitor CA-074: A novel strategy for neuroprotection based on ‘calpain-cathepsin hypothesis’. Eur. J. Neurosci. 1998, 10, 1723–1733. [Google Scholar] [CrossRef]
- Foghsgaard, L.; Wissing, D.; Mauch, D.; Lademann, U.; Bastholm, L.; Boes, M.; Elling, F.; Leist, M.; Jaattela, M. Cathepsin B acts as a dominant execution protease in tumor cell apoptosis induced by tumor necrosis factor. J. Cell. Biol. 2001, 153, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.M.; Parker, R. The RNase Rny1p cleaves tRNAs and promotes cell death during oxidative stress in Saccharomyces cerevisiae. J. Cell. Biol. 2009, 185, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Boya, P.; Kroemer, G. Lysosomal membrane permeabilization in cell death. Oncogene 2008, 27, 6434–6451. [Google Scholar] [CrossRef]
- Deepak, M.; Sangli, G.K.; Arun, P.C.; Amit, A. Quantitative determination of the major saponin mixture bacoside A in Bacopa monnieri by HPLC. Phytochem. Anal. 2005, 16, 24–29. [Google Scholar] [CrossRef]
- Wassler, M.; Jonasson, I.; Persson, R.; Fries, E. Differential permeabilization of membranes by saponin treatment of isolated rat hepatocytes. Release of secretory proteins. Biochem. J. 1987, 247, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.C.; Favre, M.; Bensa, J.C. Membrane cell permeabilization with saponin and multiparametric analysis by flow cytometry. Cytometry 1991, 12, 550–558. [Google Scholar] [CrossRef]
- Roberg, K.; Ollinger, K. Oxidative stress causes relocation of the lysosomal enzyme cathepsin D with ensuing apoptosis in neonatal rat cardiomyocytes. Am. J. Pathol. 1998, 152, 1151–1156. [Google Scholar]
- Liu, N.; Raja, S.M.; Zazzeroni, F.; Metkar, S.S.; Shah, R.; Zhang, M.; Wang, Y.; Bromme, D.; Russin, W.A.; Lee, J.C.; et al. NF-kappaB protects from the lysosomal pathway of cell death. EMBO J. 2003, 22, 5313–5322. [Google Scholar] [CrossRef]
- de Duve, C.; de Barsy, T.; Poole, B.; Trouet, A.; Tulkens, P.; Van Hoof, F. Lysosomotropic agents. Biochem. Pharmacol. 1974, 23, 2495–2531. [Google Scholar] [CrossRef]
- Ammerer, G.; Hunter, C.P.; Rothman, J.H.; Saari, G.C.; Valls, L.A.; Stevens, T.H. PEP4 gene of Saccharomyces cerevisiae encodes proteinase A, a vacuolar enzyme required for processing of vacuolar precursors. Mol. Cell. Biol. 1986, 6, 2490–2499. [Google Scholar] [CrossRef]
- Hu, J.; Dong, L.; Outten, C.E. The redox environment in the mitochondrial intermembrane space is maintained separately from the cytosol and matrix. J. Biol. Chem. 2008, 283, 29126–29134. [Google Scholar] [CrossRef]
- Armah, C.N.; Mackie, A.R.; Roy, C.; Price, K.; Osbourn, A.E.; Bowyer, P.; Ladha, S. The membrane-permeabilizing effect of avenacin A-1 involves the reorganization of bilayer cholesterol. Biophys. J. 1999, 76, 281–290. [Google Scholar] [CrossRef]
- Lorent, J.; Le Duff, C.S.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. Induction of highly curved structures in relation to membrane permeabilization and budding by the triterpenoid saponins, alpha- and delta-Hederin. J. Biol. Chem. 2013, 288, 14000–14017. [Google Scholar] [CrossRef] [PubMed]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef]
- Madeo, F.; Herker, E.; Maldener, C.; Wissing, S.; Lachelt, S.; Herlan, M.; Fehr, M.; Lauber, K.; Sigrist, S.J.; Wesselborg, S.; et al. A caspase-related protease regulates apoptosis in yeast. Mol. Cell. 2002, 9, 911–917. [Google Scholar] [CrossRef]
- Bauerle, C.; Ho, M.N.; Lindorfer, M.A.; Stevens, T.H. The Saccharomyces cerevisiae VMA6 gene encodes the 36-kDa subunit of the vacuolar H(+)-ATPase membrane sector. J. Biol. Chem. 1993, 268, 12749–12757. [Google Scholar] [CrossRef]
- Muris, D.F.; Bezzubova, O.; Buerstedde, J.M.; Vreeken, K.; Balajee, A.S.; Osgood, C.J.; Troelstra, C.; Hoeijmakers, J.H.; Ostermann, K.; Schmidt, H.; et al. Cloning of human and mouse genes homologous to RAD52, a yeast gene involved in DNA repair and recombination. Mutat. Res. 1994, 315, 295–305. [Google Scholar] [CrossRef][Green Version]
- Symington, L.S. Role of RAD52 epistasis group genes in homologous recombination and double-strand break repair. Microbiol Mol. Biol. Rev. 2002, 66, 630–670. [Google Scholar] [CrossRef] [PubMed]
- Gan, G.N.; Wittschieben, J.P.; Wittschieben, B.O.; Wood, R.D. DNA polymerase zeta (pol zeta) in higher eukaryotes. Cell Res. 2008, 18, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Paques, F.; Haber, J.E. Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1999, 63, 349–404. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, R.H.; Prakash, S. RAD1, an excision repair gene of Saccharomyces cerevisiae, is also involved in recombination. Mol. Cell. Biol. 1988, 8, 3619–3626. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.E.; Washington, M.T.; Haracska, L.; Prakash, S.; Prakash, L. Eukaryotic polymerases iota and zeta act sequentially to bypass DNA lesions. Nature 2000, 406, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Ganjewala, D.; Srivastava, A.K. Recent Progress on Chemical Composition and Bioactivities of Bacopa monnieri (Linn.) a Plant of Ayurveda. Med. Aromat. Plant. Sci. Biotechnol. 2011, 5, 102–108. [Google Scholar]
- De Zio, D.; Cianfanelli, V.; Cecconi, F. New insights into the link between DNA damage and apoptosis. Antioxid. Redox. Signal. 2013, 19, 559–571. [Google Scholar] [CrossRef]
- Wang, J.Y. DNA damage and apoptosis. Cell Death Differ. 2001, 8, 1047–1048. [Google Scholar] [CrossRef]
- Falcone, C.; Mazzoni, C. External and internal triggers of cell death in yeast. Cell Mol. Life Sci. 2016, 73, 2237–2250. [Google Scholar] [CrossRef]
- Carmona-Gutierrez, D.; Eisenberg, T.; Buttner, S.; Meisinger, C.; Kroemer, G.; Madeo, F. Apoptosis in yeast: Triggers, pathways, subroutines. Cell Death Differ. 2010, 17, 763–773. [Google Scholar] [CrossRef]
- Lang, G.I.; Murray, A.W. Estimating the per-base-pair mutation rate in the yeast Saccharomyces cerevisiae. Genetics 2008, 178, 67–82. [Google Scholar] [CrossRef]
- Novoa, C.A.; Ang, J.S.; Stirling, P.C. The A-Like Faker Assay for Measuring Yeast Chromosome III Stability. Methods Mol. Biol 2018, 1672, 1–9. [Google Scholar] [CrossRef]
- Ludovico, P.; Sousa, M.J.; Silva, M.T.; Leao, C.L.; Corte-Real, M. Saccharomyces cerevisiae commits to a programmed cell death process in response to acetic acid. Microbiology (Reading) 2001, 147, 2409–2415. [Google Scholar] [CrossRef]
- Mason, D.A.; Shulga, N.; Undavai, S.; Ferrando-May, E.; Rexach, M.F.; Goldfarb, D.S. Increased nuclear envelope permeability and Pep4p-dependent degradation of nucleoporins during hydrogen peroxide-induced cell death. FEMS Yeast Res. 2005, 5, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Boya, P.; Andreau, K.; Poncet, D.; Zamzami, N.; Perfettini, J.L.; Metivier, D.; Ojcius, D.M.; Jaattela, M.; Kroemer, G. Lysosomal membrane permeabilization induces cell death in a mitochondrion-dependent fashion. J. Exp. Med. 2003, 197, 1323–1334. [Google Scholar] [CrossRef]
- Stark, C.; Breitkreutz, B.J.; Chatr-Aryamontri, A.; Boucher, L.; Oughtred, R.; Livstone, M.S.; Nixon, J.; Van Auken, K.; Wang, X.; Shi, X.; et al. The BioGRID Interaction Database: 2011 update. Nucleic Acids Res. 2011, 39, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Hu, B.; Arno, M.J.; Panaretou, B. Genomic screening in vivo reveals the role played by vacuolar H+ ATPase and cytosolic acidification in sensitivity to DNA-damaging agents such as cisplatin. Mol. Pharmacol 2007, 71, 416–425. [Google Scholar] [CrossRef]
- Dhanasekaran, M.; Tharakan, B.; Holcomb, L.A.; Hitt, A.R.; Young, K.A.; Manyam, B.V. Neuroprotective mechanisms of ayurvedic antidementia botanical Bacopa monniera. Phytother Res. 2007, 21, 965–969. [Google Scholar] [CrossRef]
- Liu, X.; Yue, R.; Zhang, J.; Shan, L.; Wang, R.; Zhang, W. Neuroprotective effects of bacopaside I in ischemic brain injury. Restor. Neurol. Neurosci. 2013, 31, 109–123. [Google Scholar] [CrossRef]
- Pei, J.V.; Kourghi, M.; De Ieso, M.L.; Campbell, E.M.; Dorward, H.S.; Hardingham, J.E.; Yool, A.J. Differential Inhibition of Water and Ion Channel Activities of Mammalian Aquaporin-1 by Two Structurally Related Bacopaside Compounds Derived from the Medicinal Plant Bacopa monnieri. Mol. Pharmacol. 2016, 90, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Chin, S.P.; Sukumaran, S.D.; Buckle, M.J.; Kiew, L.V.; Chung, L.Y. In Silico and In Vitro Analysis of Bacoside A Aglycones and Its Derivatives as the Constituents Responsible for the Cognitive Effects of Bacopa monnieri. PLoS ONE 2015, 10, e0126565. [Google Scholar] [CrossRef] [PubMed]
- Phrompittayarat, W.; Putalun, W.; Tanaka, H.; Jetiyanon, K.; Wittaya-areekul, S.; Ingkaninan, K. Comparison of Various Extraction Methods of Bacopa monnieri. Naresuan Univ. J. 2007, 15, 29–34. [Google Scholar]
- Brachmann, C.B.; Davies, A.; Cost, G.J.; Caputo, E.; Li, J.; Hieter, P.; Boeke, J.D. Designer deletion strains derived from Saccharomyces cerevisiae S288C: A useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 1998, 14, 115–132. [Google Scholar] [CrossRef]
- Liu, X.F.; Elashvili, I.; Gralla, E.B.; Valentine, J.S.; Lapinskas, P.; Culotta, V.C. Yeast lacking superoxide dismutase: Isolation of genetic suppressors. J. Biol. Chem. 1992, 267, 18298–18302. [Google Scholar] [CrossRef]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. Applications of high efficiency lithium acetate transformation of intact yeast cells using single-stranded nucleic acids as carrier. Yeast 1991, 7, 253–263. [Google Scholar] [CrossRef]
- Sherman, F.; Fink, G.R.; Lawrence, C.W. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1978; pp. 178–179. [Google Scholar]
- Jensen, L.T.; Ajua-Alemanji, M.; Culotta, V.C. The Saccharomyces cerevisiae high affinity phosphate transporter encoded by PHO84 also functions in manganese homeostasis. J. Biol. Chem. 2003, 278, 42036–42040. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Sundin, B.A.; Chiu, C.H.; Riffle, M.; Davis, T.N.; Muller, E.G. Localization of proteins that are coordinately expressed with Cln2 during the cell cycle. Yeast 2004, 21, 793–800. [Google Scholar] [CrossRef]
- Jensen, L.T.; Carroll, M.C.; Hall, M.D.; Harvey, C.J.; Beese, S.E.; Culotta, V.C. Down-regulation of a manganese transporter in the face of metal toxicity. Mol. Biol. Cell 2009, 20, 2810–2819. [Google Scholar] [CrossRef] [PubMed]
- Indge, K.J. The isolation and properties of the yeast cell vacuole. J. Gen. Microbiol. 1968, 51, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.T.; Culotta, V.C. Role of Saccharomyces cerevisiae ISA1 and ISA2 in iron homeostasis. Mol. Cell Biol. 2000, 20, 3918–3927. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huangteerakul, C.; Aung, H.M.; Thosapornvichai, T.; Duangkaew, M.; Jensen, A.N.; Sukrong, S.; Ingkaninan, K.; Jensen, L.T. Chemical-Genetic Interactions of Bacopa monnieri Constituents in Cells Deficient for the DNA Repair Endonuclease RAD1 Appear Linked to Vacuolar Disruption. Molecules 2021, 26, 1207. https://doi.org/10.3390/molecules26051207
Huangteerakul C, Aung HM, Thosapornvichai T, Duangkaew M, Jensen AN, Sukrong S, Ingkaninan K, Jensen LT. Chemical-Genetic Interactions of Bacopa monnieri Constituents in Cells Deficient for the DNA Repair Endonuclease RAD1 Appear Linked to Vacuolar Disruption. Molecules. 2021; 26(5):1207. https://doi.org/10.3390/molecules26051207
Chicago/Turabian StyleHuangteerakul, Chananya, Hsu Mon Aung, Thitipa Thosapornvichai, Marisa Duangkaew, Amornrat Naranuntarat Jensen, Suchada Sukrong, Kornkanok Ingkaninan, and Laran T. Jensen. 2021. "Chemical-Genetic Interactions of Bacopa monnieri Constituents in Cells Deficient for the DNA Repair Endonuclease RAD1 Appear Linked to Vacuolar Disruption" Molecules 26, no. 5: 1207. https://doi.org/10.3390/molecules26051207
APA StyleHuangteerakul, C., Aung, H. M., Thosapornvichai, T., Duangkaew, M., Jensen, A. N., Sukrong, S., Ingkaninan, K., & Jensen, L. T. (2021). Chemical-Genetic Interactions of Bacopa monnieri Constituents in Cells Deficient for the DNA Repair Endonuclease RAD1 Appear Linked to Vacuolar Disruption. Molecules, 26(5), 1207. https://doi.org/10.3390/molecules26051207