
Crosstalk of Cancer Signaling Pathways by Cyclic Hexapeptides and Anthraquinones from Rubia cordifolia

 ,
,  ,
,  ,
,  ,
, 
Abstract
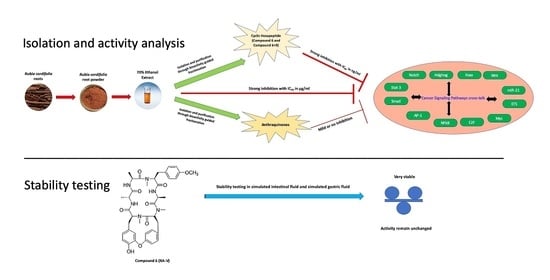
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extraction and Isolation
3.4. UHPLC/APCI-MS Analysis
3.5. Transfection and Luciferase Assays
3.6. Cytotoxicity Assay
3.7. SGF Digestion Stability Assay
3.8. SIF Digestion Stability Assay
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Shan, M.; Yu, S.; Yan, H.; Chen, P.; Zhang, L.; Ding, A. A review of the botany, phytochemistry, pharmacology and toxicology of rubiae radix et rhizoma. Molecules 2016, 21, 1747. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Yang, J.; Wang, Z.; Yang, B.; Kuang, H.; Liu, L.; Wang, L.; Yang, C. Simultaneous determination of purpurin, munjistin and mollugin in rat plasma by ultra high performance liquid chromatography-tandem mass spectrometry: Application to a pharmacokinetic study after oral administration of Rubia cordifolia L. extract. Molecules 2016, 21, 717. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China Part I; Chinese Medical Science Press: Beijing, China, 2015; Volume 1, pp. 234–235. [Google Scholar]
- Do, M.T.; Hwang, Y.P.; Kim, H.G.; Na, M.; Jeong, H.G. Mollugin inhibits proliferation and induces apoptosis by suppressing fatty acid synthase in HER2-overexpressing cancer cells. J. Cell. Physiol. 2013, 228, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Son, J.K.; Jung, S.J.; Jung, J.H.; Fang, Z.; Lee, C.S.; Seo, C.S.; Moon, D.C.; Min, B.S.; Kim, M.R.; Woo, M.H. Anticancer Constituents from the Roots of Rubia cordifolia L. Chem. Pharm. Bull. 2008, 56, 213–216. [Google Scholar] [CrossRef]
- Jolad, S.D.; Hoffman, J.J.; Torrance, S.J.; Wiedhopf, R.M.; Cole, J.R.; Arora, S.K.; Bates, R.B.; Gargiulo, R.L.; Kriek, G.R. Bouvardin and deoxybouvardin, antitumor cyclic hexapeptides from Bouvardia ternifolia (Rubiaceae). J. Am. Chem. Soc. 1977, 99, 8040–8044. [Google Scholar] [CrossRef]
- Itokawa, H.; Takeya, K.; Mori, N.; Sonobe, T.; Mihashi, S.; Hamanaka, T. Studies on antitumor cyclic hexapeptides RA obtained from Rubiae radix, Rubiaceae. VI.: Minor antitumor constituents. Chem. Pharm. Bull. 1986, 34, 3762–3768. [Google Scholar] [CrossRef]
- Itokawa, H.; Takeya, K.; Mihara, K.; Mori, N.; Hamanaka, T.; Sonobe, T.; Iitaka, Y. Studies on the antitumor cyclic hexapeptides obtained from Rubiae Radix. Chem. Pharm. Bull. 1983, 31, 1424–1427. [Google Scholar] [CrossRef]
- Tan, N.-H.; Zhou, J. Plant cyclopeptides. Chem. Rev. 2006, 106, 840–895. [Google Scholar] [CrossRef]
- Shilpa, P.; Venkatabalasubramanian, S.; Devaraj, S.N. Ameliorative effect of methanol extract of Rubia cordifolia in N-nitrosodiethylamine-induced hepatocellular carcinoma. Pharm. Biol. 2012, 50, 376–383. [Google Scholar] [CrossRef]
- Itokawa, H.; Takeya, K.; Mori, N.; Hamanaka, T.; Sonobe, T.; Mihara, K. Isolation and antitumor activity of cyclic hexapeptides isolated from Rubiae Radix. Chem. Pharm. Bull. 1984, 32, 284–290. [Google Scholar] [CrossRef]
- Chen, X.-Q.; Zhao, S.-M.; Wang, Z.; Zeng, G.-Z.; Huang, M.-B.; Tan, N.-H. Rubicordins A–C, new cyclopeptides from Rubia cordifolia with cytotoxicity and inhibiting NF-κB signaling pathway. Tetrahedron 2015, 71, 9673–9678. [Google Scholar] [CrossRef]
- Sever, R.; Brugge, J.S. Signal transduction in cancer. Cold Spring Harb. Perspect. Med. 2015, 5, a006098. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.A. The jak/stat pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011205. [Google Scholar] [CrossRef] [PubMed]
- Clara, J.A.; Monge, C.; Yang, Y.; Takebe, N. Targeting signalling pathways and the immune microenvironment of cancer stem cells—A clinical update. Nat. Rev. Clin. Oncol. 2020, 17, 204–232. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S. Oncogenic signaling pathways in the cancer genome atlas. Cell 2018, 173, 321–337.e310. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E. Transcription factors as targets for cancer therapy. Nat. Rev. Cancer 2002, 2, 740–749. [Google Scholar] [CrossRef]
- Zaki, M.A.; Balachandran, P.; Khan, S.; Wang, M.; Mohammed, R.; Hetta, M.H.; Pasco, D.S.; Muhammad, I. Cytotoxicity and modulation of cancer-related signaling by (Z)-and (E)-3,4,3′,5′-tetramethoxystilbene isolated from Eugenia rigida. J. Nat. Prod. 2013, 76, 679–684. [Google Scholar] [CrossRef]
- Danielsen, K. Substituent-effect additivity in 1-X-, 2-X-and 1, 2-di-X-9, 10-anthraquinone series, a tool for 13C NMR chemical shift assignment. Magn. Reson. Chem. 1995, 33, 823–826. [Google Scholar] [CrossRef]
- Bhuyan, R.; Saikia, C. Isolation of colour components from native dye-bearing plants in northeastern India. Bioresour. Technol. 2005, 96, 363–372. [Google Scholar] [CrossRef]
- Danielsen, K.; Aksnes, D.W.; Francis, G.W. NMR study of some anthraquinones from rhubarb. Magn. Reson. Chem. 1992, 30, 359–360. [Google Scholar] [CrossRef]
- Raimundo, J.M.; Trindade, A.P.F.; Velozo, L.S.M.; Kaplan, M.A.C.; Sudo, R.T.; Zapata-Sudo, G. The lignan eudesmin extracted from Piper truncatum induced vascular relaxation via activation of endothelial histamine H1 receptors. Eur. J. Pharmacol. 2009, 606, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Koyama, J.; Morita, I.; Kobayashi, N.; Hirai, K.; Simamura, E.; Nobukawa, T.; Kadota, S. Antiallergic activity of aqueous extracts and constituents of Taxus yunnanensis. Biol. Pharm. Bull. 2006, 29, 2310–2312. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Gauri, S.S.; Mukhopadhyay, S.K.; Chatterjee, S.; Das, S.S.; Mandal, S.M.; Dey, S. Identification and structural characterization of a new pro-apoptotic cyclic octapeptide cyclosaplin from somatic seedlings of Santalum album L. Peptides 2014, 54, 148–158. [Google Scholar] [CrossRef]
- Zhao, S.M.; Kuang, B.; Fan, J.T.; Yan, H.; Xu, W.Y.; Tan, N.H. Antitumor cyclic hexapeptides from Rubia plants: History, chemistry, and mechanism (2005–2011). CHIMIA Int. J. Chem. 2011, 65, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, S.-M.; Zhao, L.-M.; Chen, X.-Q.; Zeng, G.-Z.; Tan, N.-H. Rubipodanin A, the first natural N-desmonomethyl Rubiaceae-type cyclopeptide from Rubia podantha, indicating an important role of the N 9-methyl group in the conformation and bioactivity. PLoS ONE 2015, 10, e0144950. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, S.; Song, L.; Pu, Y.; Wang, Q.; Zeng, G.; Liu, X.; Bai, M.; Li, S.; Gao, F. Natural cyclopeptide RA-V inhibits the NF-κB signaling pathway by targeting TAK1. Cell Death Dis. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Fang, X.-Y.; Chen, W.; Fan, J.-T.; Song, R.; Wang, L.; Gu, Y.-H.; Zeng, G.-Z.; Shen, Y.; Wu, X.-F.; Tan, N.-H. Plant cyclopeptide RA-V kills human breast cancer cells by inducing mitochondria-mediated apoptosis through blocking PDK1–AKT interaction. Toxicol. Appl. Pharmacol. 2013, 267, 95–103. [Google Scholar] [CrossRef]
- Yue, G.G.; Fan, J.T.; Lee, J.K.; Zeng, G.Z.; Ho, T.W.; Fung, K.P.; Leung, P.C.; Tan, N.H.; Lau, C.B. Cyclopeptide RA-V inhibits angiogenesis by down-regulating ERK1/2 phosphorylation in HUVEC and HMEC-1 endothelial cells. Br. J. Pharmacol. 2011, 164, 1883–1898. [Google Scholar] [CrossRef]
- Ji, X.; Song, L.; Sheng, L.; Gao, A.; Zhao, Y.; Han, S.; Zhang, Y.; Zhu, C.; Zhao, S.; Wang, Z. Cyclopeptide RA-V Inhibits Organ Enlargement and Tumorigenesis Induced by YAP Activation. Cancers 2018, 10, 449. [Google Scholar] [CrossRef]
- Badouel, C.; McNeill, H. SnapShot: The hippo signaling pathway. Cell 2011, 145, 484–484.e481. [Google Scholar] [CrossRef]
- Jiang, L.; Li, J.; Zhang, C.; Shang, Y.; Lin, J. YAP-mediated crosstalk between the Wnt and Hippo signaling pathways. Mol. Med. Rep. 2020, 22, 4101–4106. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.L.; Sheehan, S.A.; Retzbach, E.P.; Timmerman, C.A.; Gianneschi, G.B.; Tempera, P.J.; Balachandran, P.; Goldberg, G.S. Effects of Maackia amurensis seed lectin (MASL) on oral squamous cell carcinoma (OSCC) gene expression and transcriptional signaling pathways. J. Cancer Res. Clin. Oncol. 2021, 147, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, M.; Li, K.; Sun, S.; Xing, G.; Gao, R.; Lei, Z.; Zhang, Z. Anti-tumor and anti-metastasis activities of honey bee larvae powder by suppressing the expression of EZH2. Biomed. Pharmacother. 2018, 105, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. The United States Pharmacopeia 23, The National Formulary 18. In United States Pharmacopeial Convention; US Pharmacopeial Convention. Inc.: Rockville, MD, USA, 1995. [Google Scholar]
- Imajo, M.; Miyatake, K.; Iimura, A.; Miyamoto, A.; Nishida, E. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/β-catenin signalling. EMBO J. 2012, 31, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; Kim, Y.C.; Yu, B.; Moroishi, T.; Mo, J.-S.; Plouffe, S.W.; Meng, Z.; Lin, K.C.; Yu, F.-X.; Alexander, C.M. Alternative Wnt signaling activates YAP/TAZ. Cell 2015, 162, 780–794. [Google Scholar] [CrossRef]
- Kuroda, M.; Wada, H.; Kimura, Y.; Ueda, K.; Kioka, N. Vinculin promotes nuclear localization of TAZ to inhibit ECM stiffness-dependent differentiation into adipocytes. J. Cell Sci. 2017, 130, 989–1002. [Google Scholar] [CrossRef]
- Azzolin, L.; Panciera, T.; Soligo, S.; Enzo, E.; Bicciato, S.; Dupont, S.; Bresolin, S.; Frasson, C.; Basso, G.; Guzzardo, V. YAP/TAZ incorporation in the β-catenin destruction complex orchestrates the Wnt response. Cell 2014, 158, 157–170. [Google Scholar] [CrossRef]
- Wang, J.; Martin, J. Hippo pathway: An emerging regulator of craniofacial and dental development. J. Dent. Res. 2017, 96, 1229–1237. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stat3/IL-6 | Smad/TGF-beta | Ap-1/PMA | NF-κB/PMA | E2F/PMA | Myc/PMA | Ets/PMA | Notch/PMA | FoxO | Wnt/m-wnt 3a | Hdghog/PMA | miR-21 | pTK | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rubia 70% ETOH extract | 8.9 | 11.9 | 5.2 | 5.9 | 16.9 | 3.7 | 8.4 | 25.6 | ND | 2.5 | 5.3 | 25.4 | ND |
| Compound 6 | 0.18 | 0.3 | 0.22 | 0.17 | 0.36 | 0.075 | 0.18 | 0.093 | 0.1 | 0.05 | 0.2 | ND | 0.9 |
| Compound 6+9 | 0.05 | 0.11 | 0.05 | 0.05 | 0.19 | 0.012 | 0.062 | 0.13 | 0.47 | 0.0013 | 0.046 | 0.7 | 0.47 |
| Pure Compounds | Stat3/IL-6 | Smad/TGF-β | Ap-1/PMA | NF-kB/PMA | E2F/PMA | Myc/PMA | Ets/PMA | Notch/PMA | FoxO/No Induction | Wnt/m-wnt 3a | Hdghog/PMA | mi-R21/No Induction | pTK/No Induction |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound 1 (Alizarine red) (30µM) | 81 | 109 | 87 | 60 | 172 | 61 | 66 | 106 | 69 | 46 | 57 | 141 | 116 |
| Compound 2 (Purpurine) (30µM) | 102 | 163 | 73 | 65 | 190 | 61 | 103 | 194 | 94 | 77 | 152 | 259 | 198 |
| Compound 3 (Emodin ) (30µM) | 48 | 46 | 45 | 45 | 144 | 46 | 53 | 424 | 112 | 11 | 42 | 309 | 144 |
| Compound 11 (Chrysophanol) (30µM) | 171 | 160 | 115 | 90 | 251 | 144 | 130 | 195 | 102 | 114 | 214 | 280 | 183 |
| Compound 12 (Danthron) (30µM) | 137 | 101 | 122 | 62 | 328 | 115 | 120 | 126 | 85 | 122 | 168 | 156 | 101 |
| Compound 13 (Quinizarin) (30µM) | 164 | 159 | 107 | 62 | 282 | 81 | 127 | 198 | 101 | 97 | 142 | 171 | 134 |
| Compound 14 (Aloe-Emodin) (30µM) | 132 | 78 | 138 | 70 | 178 | 95 | 128 | 157 | 109 | 129 | 99 | 167 | 136 |
| Compound 15 (Dehydro-α-lapachone) (DAL) (5µM) | 22 | 27 | 88 | 44 | 82 | 44 | 45 | 85 | 100 | 53 | 86 | 127 | 97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balachandran, P.; Ibrahim, M.A.; Zhang, J.; Wang, M.; Pasco, D.S.; Muhammad, I. Crosstalk of Cancer Signaling Pathways by Cyclic Hexapeptides and Anthraquinones from Rubia cordifolia. Molecules 2021, 26, 735. https://doi.org/10.3390/molecules26030735
Balachandran P, Ibrahim MA, Zhang J, Wang M, Pasco DS, Muhammad I. Crosstalk of Cancer Signaling Pathways by Cyclic Hexapeptides and Anthraquinones from Rubia cordifolia. Molecules. 2021; 26(3):735. https://doi.org/10.3390/molecules26030735
Chicago/Turabian StyleBalachandran, Premalatha, Mohamed Ali Ibrahim, Jin Zhang, Mei Wang, David S. Pasco, and Ilias Muhammad. 2021. "Crosstalk of Cancer Signaling Pathways by Cyclic Hexapeptides and Anthraquinones from Rubia cordifolia" Molecules 26, no. 3: 735. https://doi.org/10.3390/molecules26030735
APA StyleBalachandran, P., Ibrahim, M. A., Zhang, J., Wang, M., Pasco, D. S., & Muhammad, I. (2021). Crosstalk of Cancer Signaling Pathways by Cyclic Hexapeptides and Anthraquinones from Rubia cordifolia. Molecules, 26(3), 735. https://doi.org/10.3390/molecules26030735