
Profiling of Antifungal Activities and In Silico Studies of Natural Polyphenols from Some Plants

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Percent Extract Recovery
2.2. Antifungal Activity
2.3. Antioxidant Activities
2.3.1. Total Antioxidant Capacity and Total Reducing Power
2.3.2. DPPH Scavenging Assay
2.4. Phytochemical Investigation
2.4.1. Total Phenolic and Flavonoid Content
2.4.2. Reversed-Phase HPLC Analysis
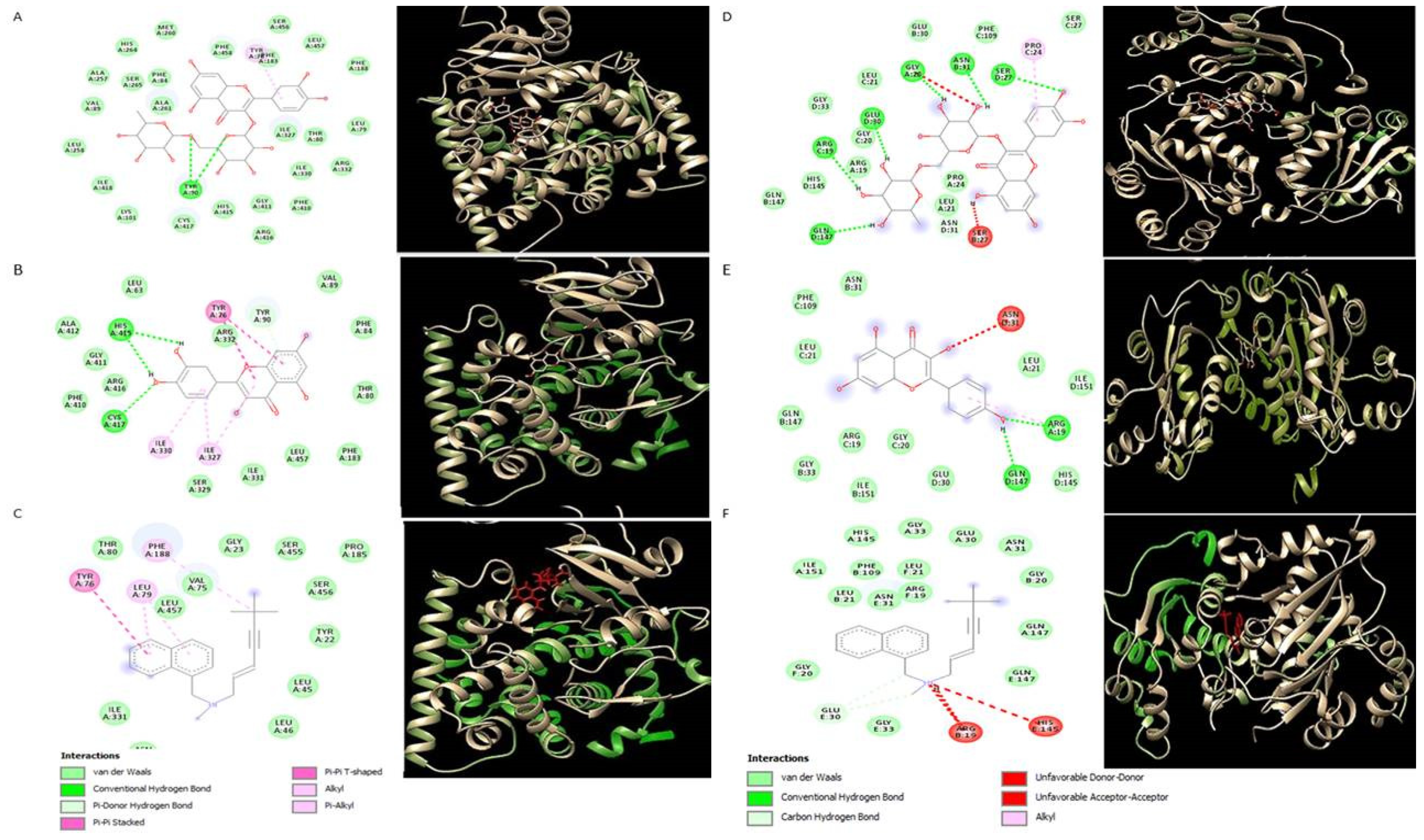
2.5. Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Collection
4.2. Extract Preparation
4.3. Antifungal Activity
4.4. Antioxidant Activities
4.4.1. Total Reducing Power (TRP)
4.4.2. Total Antioxidant Capacity (TAC)
4.4.3. DPPH (2,2-Diphenyl1–1-picryl-hydrazyl radical) Free Radical Scavenging
4.5. Phytochemical Analysis
4.5.1. Total Phenolic Content (TPC)
4.5.2. Total Flavonoid Content (TFC)
4.5.3. High-Performance Liquid Chromatography (HPLC) Analysis
4.6. Molecular Docking
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases—estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Guo, S.; Fayyaz, S.; Zhang, G.; Xu, B. Targeting programmed fusobacterium nucleatum Fap2 for colorectal cancer therapy. Cancers 2019, 11, 1592. [Google Scholar] [CrossRef] [Green Version]
- Thomas, B.; Audonneau, N.C.; Machouart, M.; Debourgogne, A. Fusarium infections: Epidemiological aspects over 10 years in a university hospital in France. J. Infect. Public Health 2020, 13, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Blaize, M.; Mayaux, J.; Nabet, C.; Lampros, A.; Marcelin, A.-G.; Thellier, M.; Piarroux, R.; Demoule, A.; Fekkar, A. Fatal invasive aspergillosis and coronavirus disease in an immunocompetent patient. Emerg. Infect. Dis. 2020, 26, 1636. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, D.; Dandagi, S.; Chandrappa, P.R.; Hema, K. Mucormycosis in immunocompetent patient resulting in extensive maxillary sequestration. J. Oral Maxillofac. Pathol. JOMFP 2018, 22, S112. [Google Scholar] [CrossRef]
- Aboody, M.S.A.; Mickymaray, S. Anti-fungal efficacy and mechanisms of flavonoids. Antibiotics 2020, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Taghipour, S.; Shamsizadeh, F.; Pchelin, I.M.; Rezaei-Matehhkolaei, A.; Mahmoudabadi, A.Z.; Valadan, R.; Ansari, S.; Katiraee, F.; Pakshir, K.; Zomorodian, K. Emergence of terbinafine resistant Trichophyton mentagrophytes in Iran, harboring mutations in the squalene epoxidase (SQLE) gene. Infect. Drug Resist. 2020, 13, 845. [Google Scholar] [CrossRef] [Green Version]
- Mickymaray, S.; Alturaiki, W. Antifungal efficacy of marine macroalgae against fungal isolates from bronchial asthmatic cases. Molecules 2018, 23, 3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duong, T.M.N.; Nguyen, P.T.; Le, T.V.; Nguyen, H.L.P.; Nguyen, B.N.T.; Nguyen, B.P.T.; Nguyen, T.A.; Chen, S.C.-A.; Barrs, V.R.; Halliday, C.L. Drug-Resistant Aspergillus flavus Is Highly Prevalent in the Environment of Vietnam: A new challenge for the management of aspergillosis? J. Fungi 2020, 6, 296. [Google Scholar] [CrossRef] [PubMed]
- Schauwvlieghe, A.F.; Buil, J.B.; Verweij, P.E.; Hoek, R.A.; Cornelissen, J.J.; Blijlevens, N.M.; Henriet, S.S.; Rijnders, B.J.; Brüggemann, R.J. High-dose posaconazole for azole-resistant aspergillosis and other difficult to treat mould infections. Mycoses 2020, 63, 122–130. [Google Scholar] [CrossRef]
- Giordani, C.; Simonetti, G.; Natsagdorj, D.; Choijamts, G.; Ghirga, F.; Calcaterra, A.; Quaglio, D.; De Angelis, G.; Toniolo, C.; Pasqua, G. Antifungal activity of Mongolian medicinal plant extracts. Nat. Prod. Res. 2020, 34, 449–455. [Google Scholar] [CrossRef]
- de Andrade Monteiro, C.; Ribeiro Alves dos Santos, J. Phytochemicals and their antifungal potential against pathogenic yeasts. Phytochem. Hum. Health 2019, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Keikha, N.; Shafaghat, M.; Mousavia, S.M.; Moudi, M.; Keshavarzi, F. Antifungal effects of ethanolic and aqueous extracts of Vitex agnus-castus against vaginal isolates of Candida albicans. Curr. Med. Mycol. 2018, 4, 1. [Google Scholar] [CrossRef]
- Li, K.; Xing, S.; Wang, M.; Peng, Y.; Dong, Y.; Li, X. Anticomplement and antimicrobial activities of flavonoids from Entada phaseoloides. Nat. Prod. Commun. 2012, 7, 1934578X1200700715. [Google Scholar] [CrossRef] [Green Version]
- Sangta, J.; Wongkaew, M.; Tangpao, T.; Withee, P.; Haituk, S.; Arjin, C.; Sringarm, K.; Hongsibsong, S.; Sutan, K.; Pusadee, T. Recovery of Polyphenolic Fraction from Arabica Coffee Pulp and Its Antifungal Applications. Plants 2021, 10, 1422. [Google Scholar] [CrossRef]
- Rocha, O.B.; do Carmo Silva, L.; de Carvalho Júnior, M.A.B.; de Oliveira, A.A.; de Almeida Soares, C.M.; Pereira, M. In vitro and in silico analysis reveals antifungal activity and potential targets of curcumin on Paracoccidioides spp. Braz. J. Microbiol. 2021, 52, 1–15. [Google Scholar] [CrossRef]
- Rampone, S.; Pagliarulo, C.; Marena, C.; Orsillo, A.; Iannaccone, M.; Trionfo, C.; Sateriale, D.; Paolucci, M. In silico analysis of the antimicrobial activity of phytochemicals: Towards a technological breakthrough. Comput. Methods Programs Biomed. 2021, 200, 105820. [Google Scholar] [CrossRef]
- Dahiya, P.; Purkayastha, S. Phytochemical screening and antimicrobial activity of some medicinal plants against multi-drug resistant bacteria from clinical isolates. Indian J. Pharm. Sci. 2012, 74, 443. [Google Scholar] [PubMed] [Green Version]
- Ahmed, M.; Fatima, H.; Qasim, M.; Gul, B. Polarity directed optimization of phytochemical and in vitro biological potential of an indigenous folklore: Quercus dilatata Lindl. ex Royle. Bmc Complementary Altern. Med. 2017, 17, 1–16. [Google Scholar] [CrossRef]
- Kowalska, J.; Tyburski, J.; Krzymińska, J.; Jakubowska, M. Cinnamon powder: An in vitro and in vivo evaluation of antifungal and plant growth promoting activity. Eur. J. Plant Pathol. 2020, 156, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Yeole, G.; Teli, N.; Kotkar, H.; Mendki, P. Cinnamomum zeylanicum extracts and their formulations control early blight of tomato. J. Biopestic. 2014, 7, 110. [Google Scholar]
- Agnihotri, S.; Wakode, S. Antimicrobial activity of essential oil and various extracts of fruits of greater cardamom. Indian J. Pharm. Sci. 2010, 72, 657. [Google Scholar] [CrossRef] [Green Version]
- Omezzine, F.; Bouaziz, M.; Daami-Remadi, M.; Simmonds, M.S.J.; Haouala, R. Chemical composition and antifungal activity of Trigonella foenum-graecum L. varied with plant ploidy level and developmental stage. Arab. J. Chem. 2017, 10, S3622–S3631. [Google Scholar] [CrossRef] [Green Version]
- Skrinjar, M.; Mandic, A.; Misan, A.; Sakac, M.; Saric, L.C.; Zec, M. Effect of Mint (Mentha piperita L.) and Caraway (Carum carvi L.) on the growth of some toxigenic Aspergillus species and Aflatoxin B1 production. Matica Srp. Proc. Nat. Sci. 2009, 116, 131–139. [Google Scholar] [CrossRef]
- Abeysekera, W.; Premakumara, G.; Ratnasooriya, W. In vitro antioxidant properties of leaf and bark extracts of ceylon cinnamon (Cinnamomum zeylanicum Blume). Trop. Agric. Res. 2013, 24, 128–138. [Google Scholar]
- Prasad, K.N.; Yang, B.; Dong, X.; Jiang, G.; Zhang, H.; Xie, H.; Jiang, Y. Flavonoid contents and antioxidant activities from Cinnamomum species. Innov. Food Sci. Emerg. Technol. 2009, 10, 627–632. [Google Scholar] [CrossRef]
- Lee, H.; Woo, E.-R.; Lee, D.G. Apigenin induces cell shrinkage in Candida albicans by membrane perturbation. Fems Yeast Res. 2018, 18, foy003. [Google Scholar] [CrossRef]
- Appell, M.; Tu, Y.-S.; Compton, D.L.; Evans, K.O.; Wang, L.C. Quantitative structure-activity relationship study for prediction of antifungal properties of phenolic compounds. Struct. Chem. 2020, 31, 1621–1630. [Google Scholar] [CrossRef]
- Samapundo, S.; De Meulenaer, B.; Osei-Nimoh, D.; Lamboni, Y.; Debevere, J.; Devlieghere, F. Can phenolic compounds be used for the protection of corn from fungal invasion and mycotoxin contamination during storage? Food Microbiol. 2007, 24, 465–473. [Google Scholar] [CrossRef]
- Palumbo, J.D.; O’Keeffe, T.L.; Mahoney, N.E. Inhibition of ochratoxin A production and growth of Aspergillus species by phenolic antioxidant compounds. Mycopathologia 2007, 164, 241–248. [Google Scholar] [CrossRef]
- Hussain, Z.; Khan, J.A.; Arshad, M.I.; Muhammad, F.; Abbas, R.Z.; Moreno, M.; Moreno Murillo, B. Comparative characterization of cinnamon, cinnamaldehyde and kaempferol for phytochemical, antioxidant and pharmacological properties using acetaminophen-induced oxidative stress mouse model. Boletín Latinoam. Caribe Plantas Med. Aromáticas 2021, 20, 339–350. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W.d.A.; de Aguiar Cordeiro, R.; Castelo-Branco, D.d.S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal effects of the flavonoids kaempferol and quercetin: A possible alternative for the control of fungal biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef]
- Oliveira, V.; Carraro, E.; Auler, M.; Khalil, N. Quercetin and rutin as potential agents antifungal against Cryptococcus spp. Braz. J. Biol. 2016, 76, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Bitencourt, T.A.; Komoto, T.T.; Massaroto, B.G.; Miranda, C.E.S.; Beleboni, R.O.; Marins, M.; Fachin, A.L. Trans-chalcone and quercetin down-regulate fatty acid synthase gene expression and reduce ergosterol content in the human pathogenic dermatophyte Trichophyton rubrum. BMC Complementary Altern. Med. 2013, 13, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Pinho, F.V.; da Cruz, L.C.; Rodrigues, N.R.; Waczuk, E.P.; Souza, C.E.; Coutinho, H.D. Phytochemical Composition, Antifungal and Antioxidant Activity of Duguetia furfuracea A. St.-Hill. 2016, 2016, 7821051. [Google Scholar]
- Ngolong Ngea, G.L.; Qian, X.; Yang, Q.; Dhanasekaran, S.; Ianiri, G.; Ballester, A.R.; Zhang, X.; Castoria, R.; Zhang, H. Securing fruit production: Opportunities from the elucidation of the molecular mechanisms of postharvest fungal infections. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2508–2533. [Google Scholar] [CrossRef]
- Nguyen, S.; Jovcevski, B.; Pukala, T.L.; Bruning, J.B. Nucleoside selectivity of Aspergillus fumigatus nucleoside-diphosphate kinase. FEBS J. 2021, 288, 2398–2417. [Google Scholar] [CrossRef]
- Monk, B.C.; Sagatova, A.A.; Hosseini, P.; Ruma, Y.N.; Wilson, R.K.; Keniya, M.V. Fungal Lanosterol 14α-demethylase: A target for next-generation antifungal design. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2020, 1868, 140206. [Google Scholar] [CrossRef]
- Jadhav, A.K.; Khan, P.K.; Karuppayil, S.M. Phytochemicals as potential inhibitors of lanosterol 14 α-demethylase (CYP51) enzyme: An in silico study on sixty molecules. Int. J. Appl. Pharm. 2020, 18–30. [Google Scholar] [CrossRef]
- Pérez, A. Terbinafine: Broad new spectrum of indications in several subcutaneous and systemic and parasitic diseases. Mycoses 1999, 42, 111–114. [Google Scholar] [CrossRef]
- Monod, M.; Feuermann, M.; Yamada, T. Terbinafine and Itraconazole Resistance in Dermatophytes. In Dermatophytes and Dermatophytoses; Springer Nature: Cham, Switzerland, 2021; pp. 415–429. [Google Scholar] [CrossRef]
- Mhatre, S.; Patravale, V. Drug repurposing of triazoles against mucormycosis using molecular docking: A short communication. Comput. Biol. Med. 2021, 136, 104722. [Google Scholar] [CrossRef]
- Godamudunage, M.P.; Lampe, J.N.; Scott, E.E. Comparison of Cytochrome P450 3A4 and 3A7 with Azole Inhibitors. FASEB J. 2017, 31, 669.5. [Google Scholar]
- Wang, Y.; Wang, S.; Nie, X.; Yang, K.; Xu, P.; Wang, X.; Liu, M.; Yang, Y.; Chen, Z.; Wang, S. Molecular and structural basis of nucleoside diphosphate kinase–mediated regulation of spore and sclerotia development in the fungus Aspergillus flavus. J. Biol. Chem. 2019, 294, 12415–12431. [Google Scholar] [CrossRef]
- de Jesús Joaquín-Ramos, A.; López-Palestina, C.U.; Pinedo-Espinoza, J.M.; Altamirano-Romo, S.E.; Santiago-Saenz, Y.O.; Aguirre-Mancilla, C.L.; Gutiérrez-Tlahque, J. Phenolic compounds, antioxidant properties and antifungal activity of jarilla (Barkleyanthus salicifolius [Kunth] H. Rob & Brettell). Chil. J. Agric. Res. 2020, 80, 352–360. [Google Scholar]
- Younus, I.; Ismail, H.; Rizvi, C.B.; Dilshad, E.; Saba, K.; Mirza, B.; Tahir, M. Antioxidant, analgesic and anti-inflammatory activities of in vitro and field-grown Iceberg lettuce extracts. J. Pharm. Pharmacogn. Res. 2019, 7, 343–355. [Google Scholar]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.; Chan, K.W.; Ebrahimi, M. Antioxidant capacities of fucoxanthin-producing algae as influenced by their carotenoid and phenolic contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef]
- Baig, M.W.; Nasir, B.; Waseem, D.; Majid, M.; Khan, M.Z.I.; Haq, I.-U. Withametelin: A biologically active withanolide in cancer, inflammation, pain and depression. Saudi Pharm. J. 2020, 28, 1526–1537. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plant Name | Percent Yield (%) |
|---|---|
| C. zeylanicum | 26.51 ± 2.22 |
| C. tamala | 18.79 ± 1.22 |
| A. subulatum | 9.45 ± 1.11 |
| T. foenumgraecum | 5.075 ± 0.52 |
| M. piperita | 10.275 ± 1.11 |
| C. sativum | 5.995 ± 0.91 |
| L. sativa | 6.37 ± 0.81 |
| B. oleraceae | 7.025 ± 1.02 |
| - | Zone of Inhibition (mm) | |||
|---|---|---|---|---|
| - | Fusarium solani | Aspergillus niger | Aspergillus flavus | Mucor Spp. |
| C. zeylanicum | 13.0 ± 0.5 | 10.0 ± 0.1 | 8.1 ± 0.1 | 10.0 ± 0.1 |
| C. tamala | 11.3 ± 1.0 | - | - | 7.1 ± 0.1 |
| A. subulatum | 13.6 ± 0.1 | 7.1 ± 0.1 | 5.0 ± 0.0 | - |
| T. foenumgraecum | 9.3 ± 0.0 | 7.1 ± 0.2 | - | - |
| M. piperita | 10.0 ± 0.1 | 6.0 ± 0.1 | - | - |
| C. sativum | 7.6 ± 0.3 | - | - | - |
| L. sativa | 9.3 ± 0.4 | - | - | - |
| B. oleraceae | 8.3 ± 0.5 | - | - | 9.2 ± 0.0 |
| Terbinafine | 20 ± 0.9 | 22 ± 1.03 | 23 ± 1.0 | 22 ± 1.0 |
| S.No | Plant Name | Total Reducing Power (Vit C Equiv mg/g Extract) | Total Antioxidant Capacity (Vit C Equiv mg/g Extract) | DPPH Free Radical Scavanging (%) | Total Phenolic Content (mg GAE/g Extract) | Total Flavonoid Content (mg QE/g Extract) | |
|---|---|---|---|---|---|---|---|
| - | - | - | - | % Scavanging at 1000 ppm (%) | IC50 (mg/mL) | - | - |
| 1 | C. zeylanicum | 63.08 ± 0.22 | 36.75 ± 0.63 | 78.82 ± 2.52 | 25.4 | 82.42 ± 5.62 | 23.66 ± 0.13 |
| 2 | C. tamala | 57.72 ± 0.41 | 48.47 ± 0.91 | 82.93 ± 2.11 | 8.681 | 176.51 ± 1.52 | 13.69 ± 0.32 |
| 3 | A. subulatum | 8.48 ± 2.42 | 29.97 ± 0.71 | 84.73 ± 3.51 | 283.4 | 24.42 ± 0.07 | 2.45 ± 0.42 |
| 4 | T. foenumgraecum | 9.28 ± 2.31 | 21.20 ± 0.82 | 75 ± 3.32 | 485.9 | 25.51 ± 0.34 | 28.79 ± 0.38 |
| 5 | M. piperita | 65.04 ± 1.11 | 32.73 ± 0.82 | 83.42 ± 3.52 | 124.9 | 136.22 ± 6.41 | 26.78 ± 1.11 |
| 6 | C. sativum | 15.72 ± 0.82 | 5.58 ± 2.12 | 83.52 ± 4.52 | 250.9 | 25.17 ± 0.82 | 11.56 ± 0.57 |
| 7 | L. sativa | 23.22 ± 0.21 | 14.56 ± 1.06 | 80.77 ± 5.52 | 221.6 | 28.63 ± 3.22 | 15.28 ± 1.29 |
| 8 | B. oleraceae | 8.72 ± 2.85 | 21.89 ± 1.11 | 60.99 ± 2.53 | 759.4 | 17.17 ± 3.22 | 2.47 ± 1.11 |
| S.N | Plant | Rutin (R) | Vanillic Acid (VA) | Quercetin (Q) | Ferulic Acid (FA) | Syringic Acid (SA) | Kaempferol (K) | Plumbagin (PL) | Apigenin (A) | Catechin (C) | Luteolin (L) | Emodin (E) | Caffeic Acid (CF) | Coumaric Acid (CA) | Gentisic Acid (GS) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | C. zeylanicum | - | 0.21 ± 0.02 | - | - | - | 0.63 ± 0.02 | - | - | - | - | - | - | - | - |
| 2 | C. tamala | 8.34 ± 0.26 | 2.35 ± 0.04 | - | - | - | - | 0.11 ± 0.04 | - | - | - | 0.54 ± 0.03 | - | - | - |
| 3 | A. subulatum | - | 0.21 ± 0.01 | 0.85 ± 0.03 | - | 0.2 ± 0.04 | - | - | - | - | - | - | - | - | - |
| 4 | T. foenumgraecum | 6.32 ± 0.03 | - | 1.35 ± 0.04 | - | 0.24 ± 0.06 | - | - | 0.53 ± 0.03 | - | - | 0.05 ± 0.002 | - | -- | - |
| 5 | M. piperita | 4.3 ± 0.05 | - | - | 3.01 ± 0.26 | 0.22 ± 0.07 | - | - | - | - | - | - | 0.43 ± 0.04 | - | - |
| 6 | C. sativum | 3.44 ± 0.06 | - | - | - | - | - | - | - | - | - | - | 0.27 ± 0.02 | - | 1.01 ± 0.22 |
| 7 | L. sativa | 3.07 ± 0.11 | - | - | - | - | - | - | - | 0.55 ± 0.03 | - | - | 0.37 ± 0.04 | - | - |
| 8 | B. oleraceae | 0.84 ± 0.03 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Polyphenol | 14-alphaDemethylase (CYP51) | Nucleoside Diphosphokinase (NDK) |
|---|---|---|
| Rutin | −9.4 | −8.9 |
| Quercetin | −8 | −7.8 |
| Kaempferol | −7.9 | −8.2 |
| Vanillic acid | −5.7 | −5.6 |
| Ferulic acid | −6.1 | −5.9 |
| Catechin | −8.1 | −7.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanzada, B.; Akhtar, N.; Okla, M.K.; Alamri, S.A.; Al-Hashimi, A.; Baig, M.W.; Rubnawaz, S.; AbdElgawad, H.; Hirad, A.H.; Haq, I.-U.; et al. Profiling of Antifungal Activities and In Silico Studies of Natural Polyphenols from Some Plants. Molecules 2021, 26, 7164. https://doi.org/10.3390/molecules26237164
Khanzada B, Akhtar N, Okla MK, Alamri SA, Al-Hashimi A, Baig MW, Rubnawaz S, AbdElgawad H, Hirad AH, Haq I-U, et al. Profiling of Antifungal Activities and In Silico Studies of Natural Polyphenols from Some Plants. Molecules. 2021; 26(23):7164. https://doi.org/10.3390/molecules26237164
Chicago/Turabian StyleKhanzada, Beenish, Nosheen Akhtar, Mohammad K. Okla, Saud A. Alamri, Abdulrahman Al-Hashimi, Muhammad Waleed Baig, Samina Rubnawaz, Hamada AbdElgawad, Abdurahman H. Hirad, Ihsan-Ul Haq, and et al. 2021. "Profiling of Antifungal Activities and In Silico Studies of Natural Polyphenols from Some Plants" Molecules 26, no. 23: 7164. https://doi.org/10.3390/molecules26237164
APA StyleKhanzada, B., Akhtar, N., Okla, M. K., Alamri, S. A., Al-Hashimi, A., Baig, M. W., Rubnawaz, S., AbdElgawad, H., Hirad, A. H., Haq, I.-U., & Mirza, B. (2021). Profiling of Antifungal Activities and In Silico Studies of Natural Polyphenols from Some Plants. Molecules, 26(23), 7164. https://doi.org/10.3390/molecules26237164