
Protease Produced by Endophytic Fungi: A Systematic Review

 ,
,  ,
,  and
and
Abstract
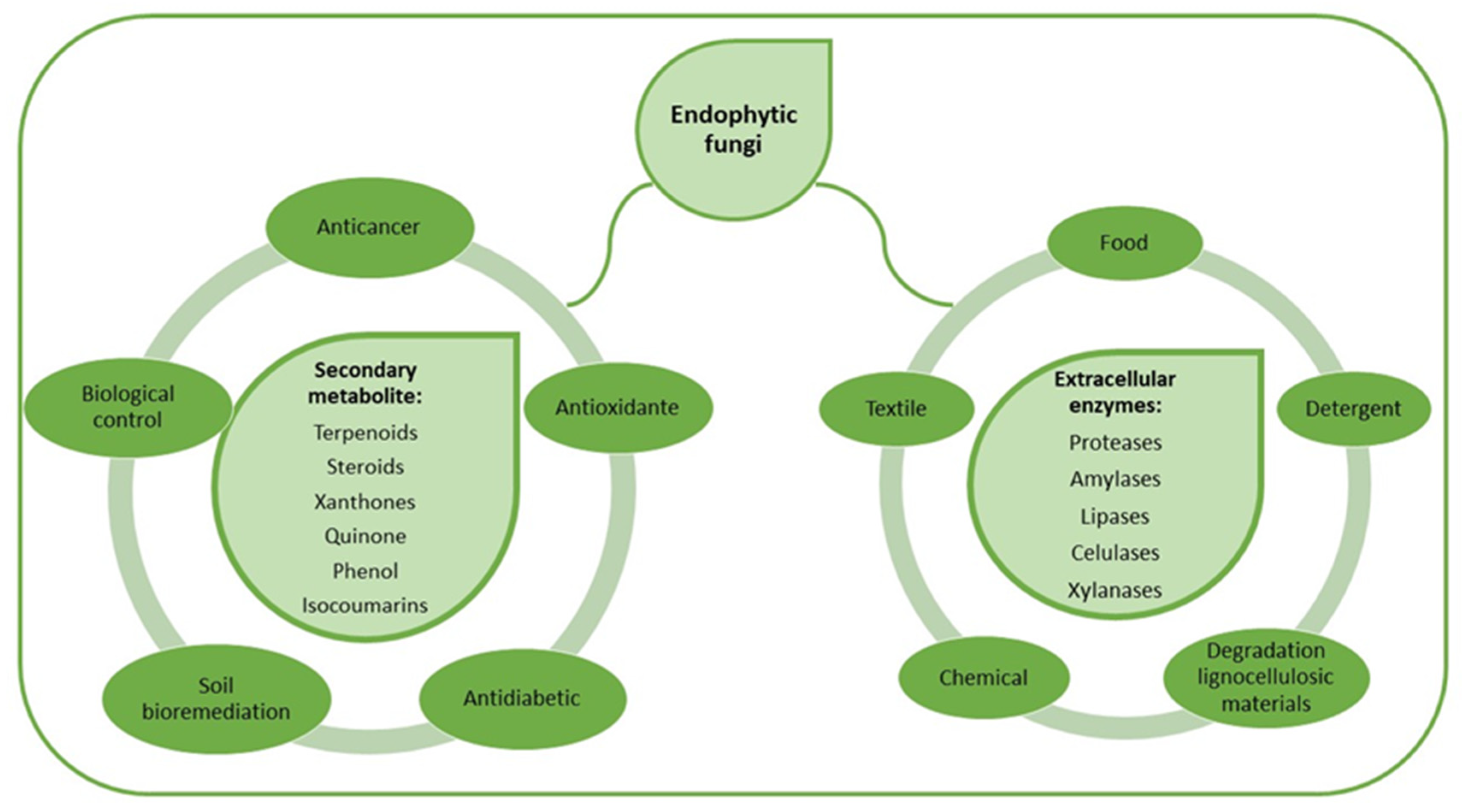
1. Introduction
2. Results
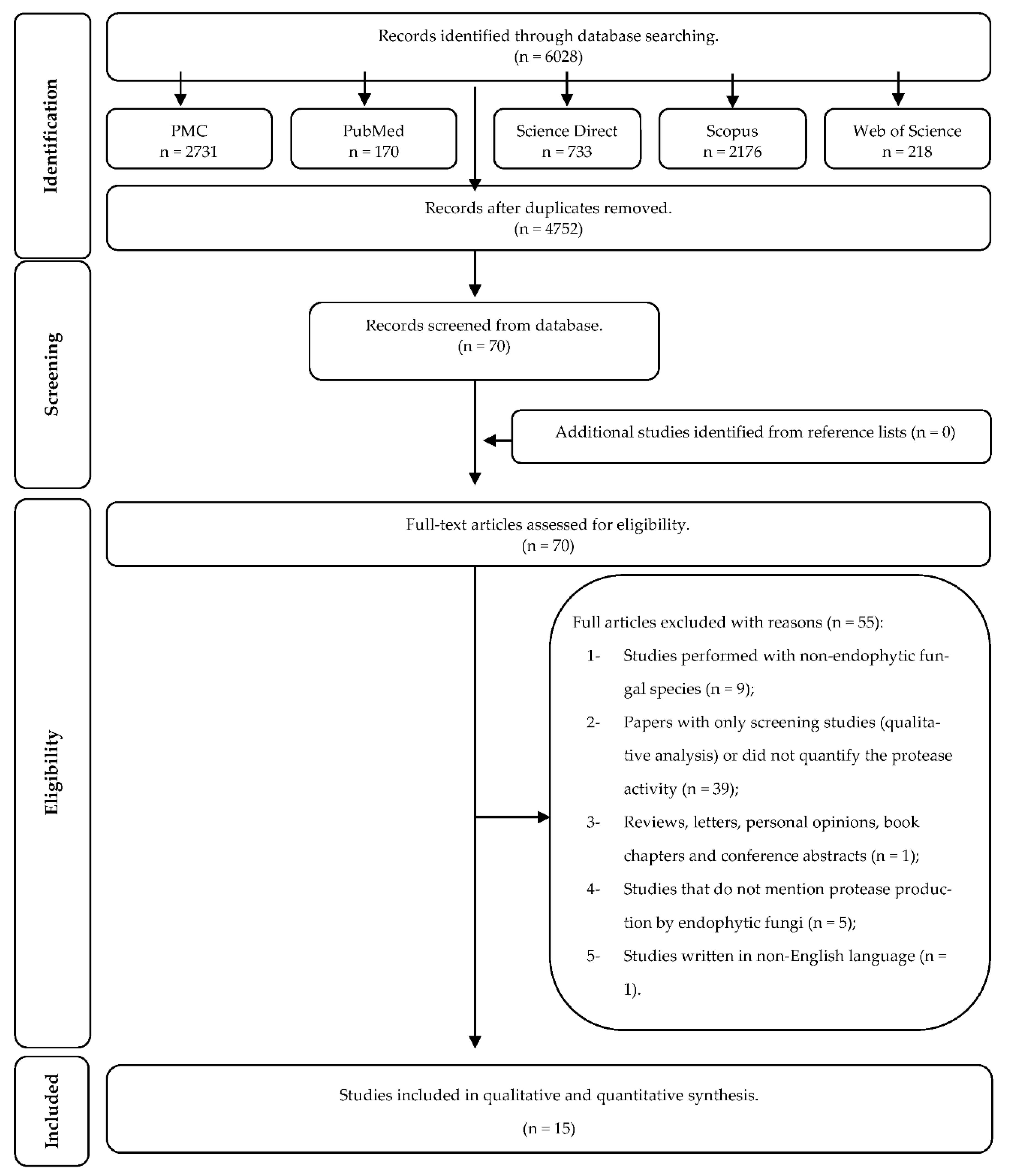
2.1. Study Selection
2.2. Study Characteristics
2.3. Synthesis of Results
2.3.1. Microorganism
2.3.2. Optimization of Protease Production
2.3.3. Growth Conditions
2.3.4. Assay for Protease Activity
2.3.5. Enzyme Characterization
2.3.6. Purification
2.4. Risk of Bias
3. Discussion
4. Materials and Methods
4.1. Information Sources and Search Strategy
4.2. Study Selection
4.3. Eligibility Criteria
4.3.1. Inclusion Criteria
4.3.2. Exclusion Criteria
4.4. Data Collection Process and Data Items
4.5. Risk of Bias in Individual Studies
4.6. Risk of Bias in Individual Studies
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| References | Reason for Exclusion |
|---|---|
| Abou El-Kassem et al. [49] | 4 |
| Alberto et al. [50] | 2 |
| Amobonye et al. [51] | 3 |
| Ayob and Simarani [42] | 2 |
| Baazeem et al. [43] | 2 |
| Bajwa et al. [52] | 1 |
| Bastos et al. [53] | 4 |
| Bensaci et al. [54] | 2 |
| Bezerra et al. [55] | 2 |
| Bezerra et al. [56] | 2 |
| Borgi et al. [57] | 1 |
| Bryant et al. [58] | 4 |
| Cairney and Burke [59] | 1 |
| da Silva et al. [60] | 2 |
| da Silva et al. [61] | 1 |
| De Azevedo Silva et al. [62] | 2 |
| Devi et al. [63] | 2 |
| El-Gendy [64] | 1 |
| Fouda et al. [38] | 2 |
| George et al. [65] | 2 |
| Gupta et al. [36] | 2 |
| Hassan [66] | 2 |
| Indarmawan et al. [67] | 1 |
| Jagannath et al. [68] | 2 |
| Jalili et al. [69] | 2 |
| Kapoor et al. [70] | 2 |
| Katoch et al. [71] | 2 |
| Katoch et al. [72] | 2 |
| Kudryavtseva et al. [73] | 5 |
| Kumar et al. [74] | 4 |
| Kuzhalvaymani et al. [75] | 1 |
| Leake and Read [76] | 1 |
| Lindstrom et al. [77] | 2 |
| Liu et al. [78] | 1 |
| Lopez-Llorca et al. [79] | 4 |
| Lumyong et al. [80] | 2 |
| Maccheroni et al. [81] | 2 |
| Martins et al. [82] | 2 |
| Meshram et al. [39] | 2 |
| Mishra et al. [83] | 2 |
| Monteiro et al. [84] | 2 |
| Orlandelli et al. [85] | 2 |
| Prathyusha et al. [86] | 2 |
| Rajagopal et al. [87] | 2 |
| Rajesh and Ravishankar Rai [88] | 2 |
| Reddy et al. [89] | 2 |
| Santos et al. [90] | 2 |
| Seshagiri and Tallapragada [91] | 2 |
| Sharma et al. [92] | 2 |
| Silva et al. [93] | 2 |
| Sopalun and Iamtham [94] | 2 |
| Sopalun et al. [95] | 2 |
| Swetha et al. [96] | 2 |
| Wu et al. [97] | 2 |
| Zaferanloo et al. [98] | 2 |
- Studies performed with non-endophytic fungal species (n = 9);
- Papers with only screening studies (qualitative analysis) or did not quantify the protease activity (n = 39);
- Reviews, letters, personal opinions, book chapters, and conference abstracts (n = 1);
- Studies that do not mention protease production by endophytic fungi (n = 5); and
- Studies written in non-English language (n = 1).
Appendix B. PRISMA

Appendix C
| Database | Search |
|---|---|
| PMC (10 September 2021) | ((“peptide hydrolases”[MeSH Terms] OR (“peptide”[All Fields] AND “hydrolases”[All Fields]) OR “peptide hydrolases”[All Fields] OR “protease”[All Fields]) OR (“peptide hydrolases”[MeSH Terms] OR (“peptide”[All Fields] AND “hydrolases”[All Fields]) OR “peptide hydrolases”[All Fields] OR (“proteolytic”[All Fields] AND “enzyme”[All Fields]) OR “proteolytic enzyme”[All Fields]) OR (“peptide hydrolases”[MeSH Terms] OR (“peptide”[All Fields] AND “hydrolases”[All Fields]) OR “peptide hydrolases”[All Fields] OR “peptidase”[All Fields]) OR (“peptide hydrolases”[MeSH Terms] OR (“peptide”[All Fields] AND “hydrolases”[All Fields]) OR “peptide hydrolases”[All Fields] OR “proteinase”[All Fields])) AND ((endophytic[All Fields] AND (“fungi”[MeSH Terms] OR “fungi”[All Fields] OR “fungus”[All Fields])) OR (endophytic[All Fields] AND (“microbiology”[Subheading] OR “microbiology”[All Fields] OR “fungi”[All Fields] OR “fungi”[MeSH Terms])) OR mycoendophyte[ All Fields]) |
| PubMed (10 September 2021) | (protease OR proteolytic enzyme OR peptidase OR proteinase) AND (endophytic fungus OR endophytic fungi OR mycoendophyte) |
| Scopus (10 September 2021) | ( ( ( ( ( protease ) OR proteinase ) OR peptidase ) OR proteolytic AND enzyme ) ) AND ( ( ( endophytic AND fungi ) OR endophytic AND fungus ) OR mycoendophytics ) AND ( LIMIT-TO ( DOCTYPE, “ar” ) OR LIMIT-TO ( DOCTYPE, “sh” ) ) |
| Science Direct (10 September 2021) | (protease OR proteolytic enzyme OR peptidase OR proteinase) AND (endophytic fungus OR endophytic fungi OR mycoendophyte)—Limited to: Research articles, Discussion, News, Short communications and Other |
| Web of Science (10 September 2021) | #1 TS = (protease OR proteolytic enzyme OR peptidase OR proteinase) AND #2 TS = (endophytic fungus OR endophytic fungi OR mycoendophyte) COMBINE #1 and #2 |
| Google Scholar (10 September 2021) | (protease OR proteolytic enzyme OR peptidase OR proteinase) AND (endophytic fungus OR endophytic fungi OR mycoendophyte) |
References
- Ahuja, S.K.; Ferreira, G.M.; Moreira, A.R. Utilization of enzymes for environmental applications. Crit. Rev. Biotechnol. 2004, 24, 125–154. [Google Scholar] [CrossRef]
- Kumar, V.; Sangwan, P.; Singh, D.; Kaur Gill, P. Global Scenario of Industrial Enzyme Market.; Nova Science Publishers: New York, NY, USA, 2014; pp. 173–196. [Google Scholar]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial proteases applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar] [CrossRef] [PubMed]
- Vojcic, L.; Pitzler, C.; Körfer, G.; Jakob, F.; Martinez, R.; Maurer, K.-H.; Schwaneberg, U. Advances in protease engineering for laundry detergents. New Biotechnol. 2015, 32, 629–634. [Google Scholar] [CrossRef]
- Nirmal, N. Fungal proteases: An overview. Int. J. Biotechnol. Biosci. 2011, 1, 1–40. [Google Scholar]
- Savitha, S.; Sadhasivam, S.; Swaminathan, K.; Lin, F.H. Fungal protease: Production, purification and compatibility with laundry detergents and their wash performance. J. Taiwan Inst. Chem. Eng. 2011, 42, 298–304. [Google Scholar] [CrossRef]
- Tavano, O.L. Protein hydrolysis using proteases: An important tool for food biotechnology. J. Mol. Catal. B Enzym. 2013, 90, 1–11. [Google Scholar] [CrossRef]
- Veloorvalappil, N.J.; Robinson, B.S.; Selvanesan, P.; Sasidharan, S.; Kizhakkepawothail, N.U.; Sreedharan, S.; Prakasan, P.; Moolakkariyil, S.J.; Sailas, B. Versatility of microbial proteases. Adv. Enzym. Res. 2013, 1, 39–51. [Google Scholar]
- Haddar, A.; Fakhfakh-Zouari, N.; Hmidet, N.; Frikha, F.; Nasri, M.; Kamoun, A.S. Low-cost fermentation medium for alkaline protease production by Bacillus mojavensis A21 using hulled grain of wheat and sardinella peptone. J. Biosci. Bioeng. 2010, 110, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, C.; Sumantha, A.; Szakacs, G.; Pandey, A. Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Process. Biochem. 2005, 40, 2689–2694. [Google Scholar] [CrossRef]
- Mandal, S.; Banerjee, D. Proteases from endophytic fungi with potential industrial applications. In Recent Advancement in White Biotechnology Through Fungi; Springer: Cham, Switzerland, 2019; pp. 319–359. [Google Scholar]
- Gurumallesh, P.; Alagu, K.; Ramakrishnan, B.; Muthusamy, S. A systematic reconsideration on proteases. Int. J. Biol. Macromol. 2019, 128, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.-S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef]
- Bacon, C.W.; White, J. Microbial Endophytes; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Rana, K.; Kour, D.; Yadav, A.; Kumar, V.; Dhaliwal, H. Endophytic microbes from wheat: Diversity and biotechnological applications for sustainable agriculture. In Proceeding of the 57th Association of Microbiologist of India & International Symposium on “Microbes and Biosphere: What’s New What’s Next”, Assam, India, 24–27 November 2016. [Google Scholar]
- Rana, K.L.; Kour, D.; Sheikh, I.; Dhiman, A.; Yadav, N.; Yadav, A.N.; Rastegari, A.A.; Singh, K.; Saxena, A.K. Endophytic fungi: Biodiversity, ecological significance, and potential industrial applications. In Recent Advancement in White Biotechnology through Fungi; Springer: Cham, Switzerland, 2019; pp. 1–62. [Google Scholar]
- Corrêa, R.C.G.; Rhoden, S.A.; Mota, T.R.; Azevedo, J.L.; Pamphile, J.A.; de Souza, C.G.M.; Polizeli, M.d.L.T.d.M.; Bracht, A.; Peralta, R.M. Endophytic fungi: Expanding the arsenal of industrial enzyme producers. J. Ind. Microbiol. Biotechnol. 2014, 41, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Mefteh, F.; Frikha, F.; Daoud, A.; Chenari Bouket, A.; Luptakova, L.; Alenezi, F.N.; Al-Anzi, B.S.; Oszako, T.; Gharsallah, N.; Belbahri, L. Response surface methodology optimization of an acidic protease produced by Penicillium bilaiae isolate TDPEF30, a newly recovered endophytic fungus from healthy roots of date palm trees (Phoenix dactylifera L.). Microorganisms 2019, 7, 74. [Google Scholar] [CrossRef]
- Bhagobaty, R.K.; Joshi, S.R. Enzymatic activity of fungi endophytic on five medicinal plant species of the pristine sacred forests of Meghalaya, India. Biotechnol. Bioprocess. Eng. 2012, 17, 33–40. [Google Scholar] [CrossRef]
- Elgammal, E.W.; El-Khonezy, M.I.; Ahmed, E.F.; Abd-Elaziz, A.M. Enhanced production, partial purification, and characterization of alkaline thermophilic protease from the endophytic fungus Aspergillus ochraceus BT21. Egypt. Pharm. J. 2020, 19, 338–349. [Google Scholar]
- Galeano, R.M.S.; Franco, D.G.; Chaves, P.O.; Giannesi, G.C.; Masui, D.C.; Ruller, R.; Corrêa, B.O.; da Silva Brasil, M.; Zanoelo, F.F. Plant growth promoting potential of endophytic Aspergillus niger 9-p isolated from native forage grass in Pantanal of Nhecolândia region, Brazil. Rhizosphere 2021, 18, 100332. [Google Scholar] [CrossRef]
- Li, Y.; Shuang, J.L.; Yuan, W.W.; Huang, W.Y.; Tan, R.X. Verticase: A fibrinolytic enzyme produced by Verticillium sp. Tj33, an endophyte of Trachelospermum jasminoides. J. Integr. Plant. Biol. 2007, 49, 1548–1554. [Google Scholar] [CrossRef]
- Lindstrom, J.T.; Belanger, F.C. Purification and characterization of an endophytic fungal proteinase that is abundantly expressed in the infected host grass. Plant. Physiol. 1994, 106, 7–16. [Google Scholar] [CrossRef]
- Matias, R.R.; Sepúlveda, A.M.G.; Batista, B.N.; de Lucena, J.; Albuquerque, P.M. Degradation of Staphylococcus aureus biofilm using hydrolytic enzymes produced by amazonian endophytic fungi. Appl. Biochem. Biotechnol. 2021, 193, 2145–2161. [Google Scholar] [CrossRef]
- Meshram, V.; Saxena, S. Potential fibrinolytic activity of an endophytic Lasiodiplodia pseudotheobromae species. 3 Biotech. 2016, 6, 114. [Google Scholar] [CrossRef]
- Meshram, V.; Saxena, S.; Paul, K. Xylarinase: A novel clot busting enzyme from an endophytic fungus Xylaria curta. J. Enzym. Inhib. Med. Chem. 2016, 31, 1502–1511. [Google Scholar] [CrossRef]
- Meshram, V.; Saxena, S.; Paul, K.; Gupta, M.; Kapoor, N. Production, purification and characterisation of a potential fibrinolytic protease from endophytic Xylaria curta by solid substrate fermentation. Appl. Biochem. Biotechnol. 2017, 181, 1496–1512. [Google Scholar] [CrossRef] [PubMed]
- Noor, Z.M.; Ahmad, M.S.; Ariffin, Z.Z. Purification and characterisation of fibrinolytic enzymes from endophytic fungi and Lignosus rhinocerus. J. Teknol. 2016, 78, 53–57. [Google Scholar]
- Rajput, K.; Chanyal, S.; Agrawal, P.K. Optimization of protease production by endophytic fungus, Alternaria alternata isolated from gymnosperm tree-Cupressus torulosa D Don. World J. Pharm. Pharmaceut. Sci. 2016, 5, 1034–1054. [Google Scholar]
- Wu, B.; Wu, L.; Chen, D.; Yang, Z.; Luo, M. Purification and characterization of a novel fibrinolytic protease from Fusarium sp. CPCC 480097. J. Ind. Microbiol. Biotechnol. 2009, 36, 451–459. [Google Scholar] [CrossRef]
- Zaferanloo, B.; Quang, T.D.; Daumoo, S.; Ghorbani, M.M.; Mahon, P.J.; Palombo, E.A. Optimization of protease production by endophytic fungus, Alternaria alternata, isolated from an Australian native plant. World J. Microbiol. Biotechnol. 2014, 30, 1755–1762. [Google Scholar] [CrossRef]
- El-Khonezy, M.I.; Elgammal, E.W.; Ahmed, E.F.; Abd-Elaziz, A.M. Detergent stable thiol-dependant alkaline protease produced from the endophytic fungus Aspergillus ochraceus BT21: Purification and kinetics. Biocatal. Agric. Biotechnol. 2021, 35, 102046. [Google Scholar] [CrossRef]
- Qiuling, X.; Jian, L.; Ling, Z. A new assay of fibrinolytic enzymes. Prog. Biochem. Biophys. 2001, 28, 582–583. [Google Scholar]
- Gupta, S.; Chaturvedi, P. Research, Phytochemical screening and extracellular enzymatic enumeration of foliar endophytic fungal isolates of Centella asiatica (L.) Urban. Int. J. Pharm. Sci. Rev. Res. 2015, 35, 21–24. [Google Scholar]
- Sunitha, V.; Nirmala Devi, D.; Srinivas, C. Extracellular enzymatic activity of endophytic fungal strains isolated from medicinal plants. World J. Agric. Sci. 2013, 9, 1–9. [Google Scholar]
- Fouda, A.H.; Hassan, S.E.-D.; Eid, A.M.; Ewais, E.E.-D. Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Ann. Agric. Sci. 2015, 60, 95–104. [Google Scholar] [CrossRef]
- Meshram, V.; Kapoor, N.; Saxena, S. Endophytic Fusarium isolates from Aegle marmelos in Western Ghats of India and their fibrinolytic ability. Sydowia 2016, 68, 119–130. [Google Scholar]
- Zhu, M.J.; Cheng, J.R.; Chen, H.T.; Deng, M.C.; Xie, W.H. Optimization of neutral protease production from Bacillus subtilis: Using agroindustrial residues as substrates and response surface methodology. Biotechnol. Appl. Biochem. 2013, 60, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Beg, Q.; Khan, S.; Chauhan, B. An overview on fermentation, downstream processing and properties of microbial alkaline proteases. Appl. Microbiol. Biotechnol. 2002, 60, 381–395. [Google Scholar]
- Ayob, F.W.; Simarani, K. Endophytic filamentous fungi from a Catharanthus roseus: Identification and its hydrolytic enzymes. Saudi Pharm. J. 2016, 24, 273–278. [Google Scholar] [CrossRef]
- Baazeem, A.; Almanea, A.; Manikandan, P.; Alorabi, M.; Vijayaraghavan, P.; Abdel-Hadi, A. In Vitro Antibacterial, Antifungal, Nematocidal and Growth Promoting Activities of Trichoderma hamatum FB10 and Its Secondary Metabolites. J. Fungi 2021, 7, 331. [Google Scholar] [CrossRef]
- Souza, P.M.; Aliakbarian, B.; Ferreira Filho, E.X.; Magalhães, P.O.; Junior, A.P.; Converti, A.; Perego, P. Kinetic and thermodynamic studies of a novel acid protease from Aspergillus foetidus. Int. J. Biol. Macromol. 2015, 81, 17–21. [Google Scholar] [CrossRef]
- Luo, Y.; Matejic, T.; Ng, C.-K.; Nunnally, B.; Porter, T.; Raso, S.; Rouse, J.; Shang, T.; Steckert, J. Technology, Characterization and analysis of biopharmaceutical proteins. Sep. Sci. Technol. 2011, 10, 283–359. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C. Understanding the roles of multifunctional mycorrhizal and endophytic fungi. In Microbial Root Endophytes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 281–298. [Google Scholar]
- Huguet, A.; Hayden, J.A.; Stinson, J.; McGrath, P.J.; Chambers, C.T.; Tougas, M.E.; Wozney, L. Judging the quality of evidence in reviews of prognostic factor research: Adapting the GRADE framework. Syst. Rev. 2013, 2, 1–12. [Google Scholar] [CrossRef]
- Abou El-Kassem, L.; Hawas, U.W.; El-Souda, S.; Ahmed, E.F.; El-Khateeb, W.; Fayad, W. Anti-HCV protease potential of endophytic fungi and cytotoxic activity. Biocatal. Agric. Biotechnol. 2019, 19, 101170. [Google Scholar] [CrossRef]
- Alberto, R.N.; Costa, A.T.; Polonio, J.C.; Santos, M.S.; Rhoden, S.A.; Azevedo, J.L.; Pamphile, J.A. Extracellular enzymatic profiles and taxonomic identification of endophytic fungi isolated from four plant species. Genet. Mol. Res. 2016, 15, gmr15049016. [Google Scholar] [CrossRef] [PubMed]
- Amobonye, A.; Bhagwat, P.; Pandey, A.; Singh, S.; Pillai, S. Biotechnological potential of Beauveria bassiana as a source of novel biocatalysts and metabolites. Crit. Rev. Biotechnol. 2020, 40, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, R.; Abuarghub, S.; Read, D.J. The biology of mycorrhiza in the Ericaceae: X. The utilization of proteins and the production of proteolytic enzymes by the mycorrhizal endophyte and by mycorrhizal plants. New Phytol. 1985, 101, 469–486. [Google Scholar] [CrossRef]
- Bastos, A.; Cardoso, P.G.; Santos, Í.A.F.M.; Trento, M.V.C.; Porto, L.C.J.; Marcussi, S. Enzymatic Modulators from Induratia spp. Curr. Microbiol. 2020, 77, 3603–3611. [Google Scholar] [CrossRef]
- Bensaci, O.A.; Daoud, H.; Lombarkia, N.; Rouabah, K. Formulation of the endophytic fungus Cladosporium oxysporum Berk. & M.A. Curtis, isolated from Euphorbia bupleuroides subsp. luteola, as a new biocontrol tool against the black bean aphid (Aphis fabae Scop.). J. Plant. Prot. Res. 2015, 55, 80–87. [Google Scholar]
- Bezerra, J.D.; Nascimento, C.C.; Barbosa Rdo, N.; da Silva, D.C.; Svedese, V.M.; Silva-Nogueira, E.B.; Gomes, B.S.; Paiva, L.M.; Souza-Motta, C.M. Endophytic fungi from medicinal plant Bauhinia forficata: Diversity and biotechnological potential. Braz. J. Microbiol. 2015, 46, 49–57. [Google Scholar] [CrossRef]
- Bezerra, J.D.; Santos, M.G.; Svedese, V.M.; Lima, D.M.; Fernandes, M.J.; Paiva, L.M.; Souza-Motta, C.M. Richness of endophytic fungi isolated from Opuntia ficus-indica Mill. (Cactaceae) and preliminary screening for enzyme production. World J. Microbiol. Biotechnol. 2012, 28, 1989–1995. [Google Scholar] [CrossRef]
- Borgi, I.; Dupuy, J.W.; Blibech, I.; Lapaillerie, D.; Lomenech, A.M.; Rebai, A.; Ksantini, M.; Bonneu, M.; Gargouri, A. Hyper-proteolytic mutant of Beauveria bassiana, a new biological control agent against the tomato borer. Agron. Sustain. Dev. 2016, 36, 60. [Google Scholar] [CrossRef][Green Version]
- Bryant, M.K.; Schardl, C.L.; Hesse, U.; Scott, B. Evolution of a subtilisin-like protease gene family in the grass endophytic fungus Epichloë festucae. BMC Evol. Biol. 2009, 9, 168. [Google Scholar] [CrossRef]
- Cairney, J.W.G.; Burke, R.M. Extracellular enzyme activities of the ericoid mycorrhizal endophyte Hymenoscyphus ericae (Read) Korf and Kernan: Their likely roles in decomposition of dead plant tissue in soil. Plant. Soil 1998, 205, 181–192. [Google Scholar] [CrossRef]
- Da Silva, M.A.; Santos, C.; Pérez-Nevado, F.; Lima, N.; da Silva Bentes, J.L. Enzymatic activity and process of initial infection of guarana plant (Paullinia Cupana) by pathogenic and endophytic strains of Colletotrichum guaranicola. Rev. Fitotec. Mex. 2021, 44, 67–75. [Google Scholar]
- Da Silva, R.R.; da Rosa, N.G.; Goncalves de Oliveira, L.C.; Juliano, M.A.; Juliano, L.; Rosa, J.C.; Cabral, H. Biochemical properties and catalytic specificity of a novel neutral serine peptidase secreted by fungus Pyrenochaetopsis sp. Appl. Biochem. Biotechnol. 2019, 187, 1158–1172. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo Silva, F.; Liotti, R.G.; Ana Paula deAraújo, B.; De Melo Reis, É.; Passos, M.B.S.; Dos Santos, E.L.; Sampaio, O.M.; Januário, A.H.; Branco, C.L.B.; Da Silva, G.F.; et al. Diversity of cultivable fungal endophytes in Paullinia cupana (Mart.) Ducke and bioactivity of their secondary metabolites. PLoS ONE 2018, 13, e0195874. [Google Scholar] [CrossRef]
- Devi, N.N.; Prabakaran, J.J.; Wahab, F. Phytochemical analysis and enzyme analysis of endophytic fungi from Centella asiatica. Asian Pac. J. Trop. Biomed. 2012, 2 (Suppl. S3), S1280–S1284. [Google Scholar] [CrossRef]
- El-Gendy, M.M. Keratinase production by endophytic Penicillium spp. Morsy1 under solid-state fermentation using rice straw. Appl. Biochem. Biotechnol. 2010, 162, 780–794. [Google Scholar] [CrossRef]
- George, T.K.; SubaidaBeevi, S.; Asok, A.K.; Shaikmoideen, J.M. Plant growth promoting endophytic yeast Geotrichum candidum (JX 477426) from roots of Bruguiera cylindrica. J. Microbiol. Biotechnol. Food Sci. 2019, 9, 267–272. [Google Scholar]
- Hassan, S.E.D. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar] [CrossRef]
- Indarmawan, T.; Mustopa, A.Z.; Budiarto, B.R.; Tarman, K. Antibacterial ativity of extracellular protease isolated from an algicolous fungus Xylaria psidii KT30 against gram-positive bacteria. HAYATI J. Biosci. 2016, 23, 73–78. [Google Scholar] [CrossRef]
- Jagannath, S.; Konappa, N.; Lokesh, A.; Bhuvaneshwari; Dasegowda, T.; Udayashankar, A.C.; Chowdappa, S.; Cheluviah, M.; Satapute, P.; Jogaiah, S. Bioactive compounds guided diversity of endophytic fungi from Baliospermum montanum and their potential extracellular enzymes. Anal. Biochem. 2021, 614, 114024. [Google Scholar] [CrossRef] [PubMed]
- Jalili, B.; Bagheri, H.; Azadi, S.; Soltani, J. Identification and salt tolerance evaluation of endophyte fungi isolates from halophyte plants. Int. J. Environ. Sci. Technol. 2020, 17, 3459–3466. [Google Scholar] [CrossRef]
- Kapoor, N.; Rajput, P.; Abu Mushtaque, M.; Gambhir, L. Bio-prospecting fungal endophytes of high altitude medicinal plants for commercially imperative enzymes. Biosci. Biotechnol. Res. Commun. 2018, 11, 370–375. [Google Scholar]
- Katoch, M.; Salgotra, A.; Singh, G. Endophytic fungi found in association with Bacopa monnieri as potential producers of industrial enzymes and antimicrobial bioactive compounds. Braz. Arch. Biol. Technol. 2014, 57, 714–722. [Google Scholar] [CrossRef]
- Katoch, M.; Singh, A.; Singh, G.; Wazir, P.; Kumar, R. Phylogeny, antimicrobial, antioxidant and enzyme-producing potential of fungal endophytes found in Viola odorata. Ann. Microbiol. 2017, 67, 529–540. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N.; Pobedinskaya, M.A.; Balabko, P.N.; Kokaeva, L.Y.; Zaichik, B.T.; Kutuzova, N.A.; Azarkovich, M.I.; Elansky, S.N.; Valueva, T.A. The proteolytic activity and virulence of Alternaria alternata strains, isolated from tomato. Mikol. I Fitopatol. 2017, 51, 110–116. [Google Scholar]
- Kumar, A.; Jha, P.K.; Kumar, R.; Kumar, K.; Sedolkar, V. Antibacterial activity, phytochemical and enzyme analysis of crude extract of endophytic fungus, Alternaria sp. isolated from an ethanobotanical medicinal plant Tridax procumbens. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 1111–1115. [Google Scholar]
- Kuzhalvaymani, K.; Jacquline, E.L.; Subha, T.S. Production, characterization of proteases by solid state fermentation using sugarcane bagasse by Warcupiella spinulosa. Int. J. Sci. Technol. Res. 2019, 8, 384–393. [Google Scholar]
- Leake, J.R.; Read, D.J. Proteinase activity in mycorrhizal fungi: I. The effect of extracellular pH on the production and activity of proteinase by ericoid endophytes from soils of contrasted pH. New Phytol. 1990, 115, 243–250. [Google Scholar] [CrossRef]
- Lindstrom, J.T.; Sun, S.; Belanger, F.C. A novel fungal protease expressed in endophytic infection of Poa species. Plant. Physiol. 1993, 102, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jia, J.; Atkinson, S.; Camara, M.; Gao, K.; Li, H.; Cao, J. Biocontrol potential of an endophytic Serratia sp G3 and its mode of action. World J. Microbiol. Biotechnol. 2010, 26, 1465–1471. [Google Scholar] [CrossRef]
- Lopez-Llorca, L.V.; Gómez-Vidal, S.; Monfort, E.; Larriba, E.; Casado-Vela, J.; Elortza, F.; Jansson, H.B.; Salinas, J.; Martín-Nieto, J. Expression of serine proteases in egg-parasitic nematophagous fungi during barley root colonization. Fungal Genet. Biol. 2010, 47, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Lumyong, S.; Lumyong, P.; McKenzie, E.H.; Hyde, K.D. Enzymatic activity of endophytic fungi of six native seedling species from Doi Suthep-Pui National Park, Thailand. Can. J. Microbiol. 2002, 48, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Maccheroni, W.; Araujo, W.L.; Azevedo, J.L. Ambient pH-regulated enzyme secretion in endophytic and pathogenic isolates of the fungal genus Colletotrichum. Sci. Agric. 2004, 61, 298–302. [Google Scholar] [CrossRef]
- Martins, J.; Veríssimo, P.; Canhoto, J. Isolation and identification of Arbutus unedo L. fungi endophytes and biological control of Phytophthora cinnamomi in vitro. Protoplasma 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Gond, S.K.; Sharma, V.K.; Verma, S.K.; Kumar, J.; Singh, D.K.; Kumar, A.; Pandey, P.; Kharwar, R.N. Characterization of Pseudofusficoccum adansoniae, an endophytic fungus residing in photosynthetic root of Tinospora cordifolia, a medicinal plant. Proc. Natl. Acad. Sci. India Sect. B—Biol. Sci. 2019, 89, 1319–1326. [Google Scholar] [CrossRef]
- Monteiro, M.C.P.; Tavares, D.G.; Nery, E.M.; de Queiroz, M.V.; Pereira, O.L.; Cardoso, P.G. Enzyme production by Induratia spp. isolated from coffee plants in Brazil. Braz. Arch. Biol. Technol. 2020, 63. [Google Scholar] [CrossRef]
- Orlandelli, R.C.; de Almeida, T.T.; Alberto, R.N.; Polonio, J.C.; Azevedo, J.L.; Pamphile, J.A. Antifungal and proteolytic activities of endophytic fungi isolated from Piper hispidum Sw. Braz. J. Microbiol. 2015, 46, 359–366. [Google Scholar] [CrossRef]
- Prathyusha, P.; Rajitha Sri, A.B.; Satya Prasad, K. Diversity and enzymatic activity of foliar endophytic fungi isolated from medicinal plants of indian dry deciduous forest. Der Pharm. Lett. 2015, 7, 244–251. [Google Scholar]
- Rajagopal, K.; Meenashree, B.; Binika, D.; Joshila, D.; Tulsi, P.S.; Arulmathi, R.; Kathiravan, G.; Tuwar, A. Mycodiversity and biotechnological potential of endophytic fungi isolated from hydrophytes. Curr. Res. Environ. Appl. Mycol. 2018, 8, 172–182. [Google Scholar] [CrossRef]
- Rajesh, P.S.; Ravishankar Rai, V. Hydrolytic enzymes and quorum sensing inhibitors from endophytic fungi of Ventilago madraspatana Gaertn. Biocatal. Agric. Biotechnol. 2013, 2, 120–124. [Google Scholar] [CrossRef]
- Reddy, P.V.; Lam, C.K.; Belanger, F.C. Mutualistic fungal endophytes express a proteinase that is homologous to proteases suspected to be important in fungal pathogenicity. Plant. Physiol. 1996, 111, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.R.; Abdel-azeem, A.M.; Mohesien, M.T.; Piekutowska, M.; Sheir, D.H.; da Silva, L.L.; Castro, C.D.S.; Carvalho, D.D.C.; Bezerra, J.D.P.; Saad, H.A.; et al. Insights into the bioprospecting of the endophytic fungi of the medicinal plant Palicourea rigida Kunth (Rubiaceae): Detailed biological activities. J. Fungi 2021, 7, 689. [Google Scholar] [CrossRef] [PubMed]
- Seshagiri, S.; Tallapragada, P. Optimization of process parameters for high biomass, alpha-amylase and protease enzyme by Piriformospora indica using mathematical model. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 916–922. [Google Scholar]
- Sharma, H.; Rai, A.K.; Chettri, R.; Nigam, P.S. Bioactivites of Penicillium citrinum isolated from a medicinal plant Swertia chirayita. Arch. Microbiol. 2021, 203, 5173–5182. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.L.D.O.; Luz, J.S.; Da Silveira, E.B.; Cavalcante, U.M.T. Endophytic fungi of Annona spp.: Isolation, enzymatic characterization of isolates and plant growth promotion in Annona squamosa L. seedlings. Acta Bot. Bras. 2006, 20, 649–655. [Google Scholar] [CrossRef]
- Sopalun, K.; Iamtham, S. Isolation and screening of extracellular enzymatic activity of endophytic fungi isolated from Thai orchids. South. Afr. J. Bot. 2020, 134, 273–279. [Google Scholar] [CrossRef]
- Sopalun, K.; Laosripaiboon, W.; Wachirachaikarn, A.; Iamtham, S. Biological potential and chemical composition of bioactive compounds from endophytic fungi associated with thai mangrove plants. South. Afr. J. Bot. 2021, 141, 66–76. [Google Scholar] [CrossRef]
- Swetha, S.; Varma, A.; Padmavathi, T. Statistical evaluation of the medium components for the production of high biomass, α-amylase and protease enzymes by Piriformospora indica using Plackett–Burman experimental design. 3 Biotech. 2014, 4, 439–445. [Google Scholar] [CrossRef]
- Wu, B.; Wu, L.; Ruan, L.; Ge, M.; Chen, D. Screening of endophytic fungi with antithrombotic activity and identification of a bioactive metabolite from the endophytic fungal strain CPCC 480097. Curr. Microbiol. 2009, 58, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Zaferanloo, B.; Virkar, A.; Mahon, P.J.; Palombo, E.A. Endophytes from an Australian native plant are a promising source of industrially useful enzymes. World J. Microbiol. Biotechnol. 2013, 29, 335–345. [Google Scholar] [CrossRef] [PubMed]

| Author | Country | Host Plant | Microorganisms | Fungal Identification | Growth Conditions | Enzyme Activity | Main Conclusions | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Ph | T (°C) | Agitation (rpm) | Time (days) | |||||||
| Mefteh et al. [20] | Tunisia | Phoenix dactylifera L. | Penicillium bilaiae | ITS-rDNA | 6.26 | 24.5 | 150 | ND | 1086.95 U/mL | Plackett–Burman design and RSM approaches were employed for optimization of culture and environment conditions and were shown to significantly enhance protease production. |
| Bhagobaty and Joshi [21] | India | Potentilla fulgens | Talaromycesflavus | β-tubulin | ND | 25 | 120 | 5 | 34.9 U/h/mL | All the endophytic fungal isolates from medicinal plant showed production of protease. The production of extracellular enzymes was greater in the liquid medium in comparison to the plate-based assays. |
| Elgammal et al. [22] | Egypt | Ruprechita salicifolia | Aspergillus ochraceus | 18S rRNA | 8 | 35 | 150 | ND | 292 U/mL | The protease production increased by about 7.5-fold after applying the final optimized fermentation. The partial purification results showed that the highly recovered fraction was at 60% ethanol concentration. |
| El-Khonezy et. al. [34] | Egypt | Ruprechita salicifolia | Aspergillus ochraceus | 18S rRNA | 8 | 35 | 150 | 6 | 800.1 U/mL | The enzyme was characterized as thiol-dependent serine alkaline protease. Low-cost production medium using different waste sources was applied to produce the enzyme. |
| Galeano et al. [23] | Brazil | Axonopus purpusii | Aspergillus niger | ITS-rDNA | ND | 30 | 110 | 7 | 12.01 U/mL | The ability of the fungus to produce proteases might reflect the fact that these fungi have potential as biocontrol agents. |
| Li et al. [24] | China | Trachelospermum jasminoides | Verticillium sp. | Morphology | ND | 28 | 160 | 14 | 3775 U/mg | Verticase is a direct degrader of fibrin clot, most probably playing a negligible role in the conversion of plasminogen to plasmin. However, for protein-based medicines, special care must be taken for an early awareness of the toxicity. |
| Lindstrom and Belanger [25] | United States | Poa ampla | Acremonium typhinum | ND | ND | ND | ND | ND | 27 U/mL | The regulated nature of proteinase At1 suggest that its function is important in the symbiotic interaction of fungus and plants. |
| Matias et al. [26] | Brazil | Myrcia guianensis | Aspergillus sp. | Morphology a | 5 | 28 | 150 | 7 | 3.63 U/mL | The endophytic fungus with the higher protease activity demonstrated total efficacy in the removal of the consolidated biofilm of S. aureus. |
| Meshram and Saxena [27] | India | Aegle marmelos | Lasiodiplodia pseudotheobromae | ITS-rDNA | ND | 26 | 130 | 7 | 6514 U/mL | The endophytic fungus possesses potential in vitro fibrinolytic potential. |
| Meshram et al. [28] | India | Cathranthus roseus | Xylaria curta | 28s rDNA | ND | 26 | 130 | 7 | 34.11 U/mL | Submerged fermentation was used to produce the fibrinolytic enzyme. This protease is a novel metalloprotease possessing dual activity including direct degradation of fibrin(ogen) or by activating the tissue plasminogen. |
| Meshram et al. [29] | India | Cathranthus roseus | Xylaria curta | ITS-rDNA 28s rDNA | ND | 28 | ND | 15 | 103.56 U/mL | The fibrinolytic enzyme xylarinase was produced by solid substrate fermentation using rice chaff medium. The purified metalloprotease showed in vitro thrombolytic activity and no cytotoxic effect. |
| Noor et al. [30] | Malaysia | Hibiscus | Fusarium sp. | 18S rRNA a | ND | ND | ND | ND | 5284 U/mL | Two fibrinolytic enzymes were purified and characterized based only on molecular weight and effect of pH and temperature. |
| Penicillium citrinum | 18S rRNA a | ND | ND | ND | ND | 2200 U/mL | ||||
| Rajput et al. [31] | India | Cupressus torulosa D. Don | Alternaria alternata | Morphology | 7 | 27 | ND | 2 | 162 U/mL | The fungus can be industrially exploited for the synthesis of protease and strain improvement studies can be carried out to enhance enzyme production. |
| Wu et al. [32] | China | Chrysanthemum | Fusarium sp. | ITS | ND | 28 | 220 | 6 | 137,000 U | The fibrinolytic enzyme, named Fu-P, was purified and identified as a chymotrypsin-like serine metalloprotease. May be a potential candidate for thrombolytic therapy or thrombosis prevention. |
| Zaferanloo et al. [33] | Australia | Eremophilia longifolia. | Alternaria alternata | Morphology ITS | 6.5 | 30 | ND | 7 | 69.86 BAEE units/mg | The protease can be applied to cheese making and in milk-clotting where the fermentation conditions are suitable to the activation of protease |
| Author/Year | Purification Method | Microorganisms | Specific Activity | Purification fold | Molecular Weight (kDa) | Ph Optimum | Temperature Optimum (°C) |
|---|---|---|---|---|---|---|---|
| Elgammal et al. [22] | Partial purification Ethanol fractionation and precipitation | Aspergillus ochraceus | 384.2 UI/mg | 0.11 | ND | 8 | 50 |
| El-Khonezy et. al. [34] | Ammonium sulfate precipitation Sephacryl S-200 DEAE-Sepharose CM-Sepharose | Aspergillus ochraceus | 111,379.5 U/mg protein | 15.3 | 59 | 8 | 50 |
| Li et al. [24] | Ammonium sulfate precipitation DEAE-52 column Sephadex G-75 Octyl Sepharose 4 FF hydrophobic column | Verticillium sp. | 3775 UI/mg | 8.1 | 31 | 9–10 | 50–60 |
| Lindstrom and Belanger [25] | Ultrafiltration 30 Kda, Phenylboronate Column Methanol precipitation | Acremonium typhinum | 710 UI/units/ng | ND | 34 | 10–11 | 37 |
| Meshram and Saxena [27] | Partial purification Ammonium sulphate precipitation and dialyze | Lasiodiplodia pseudotheobromae | 3.56 U/mg | 2.01 | 80 | ND | ND |
| Meshram et al. [28] | Ammonium sulfate precipitation Q-sepharose anion exchange | Xylaria curta | 36.67 U/mg | 9.19 | ~33 | 8 | 35 |
| Meshram et al. [29] | Ammonium sulphate precipitationSephacryl S-300 column | Xylaria curta | 9.22 U/mg | 8.37 | ~33 | 8 | 35 |
| Noor et al. [30] | Ammonium sulfate precipitation Hi-Prep 26/10 Desalting Column Hi-Trap Benzamidine FF Column | Fusarium sp. | 246.92 UI/mg | 11.2 | ~34 | 7 | 30 |
| Noor et al. [30] | Ammonium sulfate precipitation Hi-Prep 26/10 Desalting Column Hi-Trap Benzamidine FF Column | Penicilium citrinum | 198.2 UI/mg | 9.7 | ~34 | 8 | 40 |
| Wu et al. [32] | Ammonium sulfate precipitation MonoQ Column Superdex 75 Column | Fusarium sp. | 76,111 UI/mg | 158.5 | 28 | 8.5 | 45 |
| Author | Study Limitation | Inconsistency | Indirectness | Imprecision | Publication Bias | Overall Quality |
|---|---|---|---|---|---|---|
| Mefteh et al. [20] | √ | √ | √ | √ | √ | ++++ |
| Bhagobaty and Joshi [21] | X | X | √ | √ | X | + |
| Elgammal et al. [22] | √ | √ | √ | √ | √ | +++ |
| El-Khonezy et. al. [34] | √ | √ | √ | √ | √ | ++++ |
| Galeano et al. [23] | √ | √ | X | √ | √ | +++ |
| Li et al. [24] | X | X | √ | X | √ | +++ |
| Lindstrom and Belanger [25] | X | X | √ | X | √ | + |
| Matias et al. [26] | √ | X | X | √ | √ | ++ |
| Meshram and Saxena [27] | √ | X | √ | Unclear | √ | +++ |
| Meshram et al. [28] | √ | X | √ | Unclear | √ | +++ |
| Meshram et al. [29] | √ | X | √ | Unclear | √ | +++ |
| Noor et al. [30] | √ | X | √ | X | X | ++ |
| Noor et al. [30] | √ | X | √ | X | X | ++ |
| Rajput et al. [31] | √ | √ | √ | Unclear | √ | +++ |
| Wu et al. [32] | X | √ | √ | √ | √ | ++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra, V.H.S.; Cardoso, S.L.; Fonseca-Bazzo, Y.; Silveira, D.; Magalhães, P.O.; Souza, P.M. Protease Produced by Endophytic Fungi: A Systematic Review. Molecules 2021, 26, 7062. https://doi.org/10.3390/molecules26227062
Bezerra VHS, Cardoso SL, Fonseca-Bazzo Y, Silveira D, Magalhães PO, Souza PM. Protease Produced by Endophytic Fungi: A Systematic Review. Molecules. 2021; 26(22):7062. https://doi.org/10.3390/molecules26227062
Chicago/Turabian StyleBezerra, Victor Hugo Souto, Samuel Leite Cardoso, Yris Fonseca-Bazzo, Dâmaris Silveira, Pérola Oliveira Magalhães, and Paula Monteiro Souza. 2021. "Protease Produced by Endophytic Fungi: A Systematic Review" Molecules 26, no. 22: 7062. https://doi.org/10.3390/molecules26227062
APA StyleBezerra, V. H. S., Cardoso, S. L., Fonseca-Bazzo, Y., Silveira, D., Magalhães, P. O., & Souza, P. M. (2021). Protease Produced by Endophytic Fungi: A Systematic Review. Molecules, 26(22), 7062. https://doi.org/10.3390/molecules26227062