
Peptide Regulation of Gene Expression: A Systematic Review

 ,
,  ,
,  and
and
Abstract
:1. Introduction
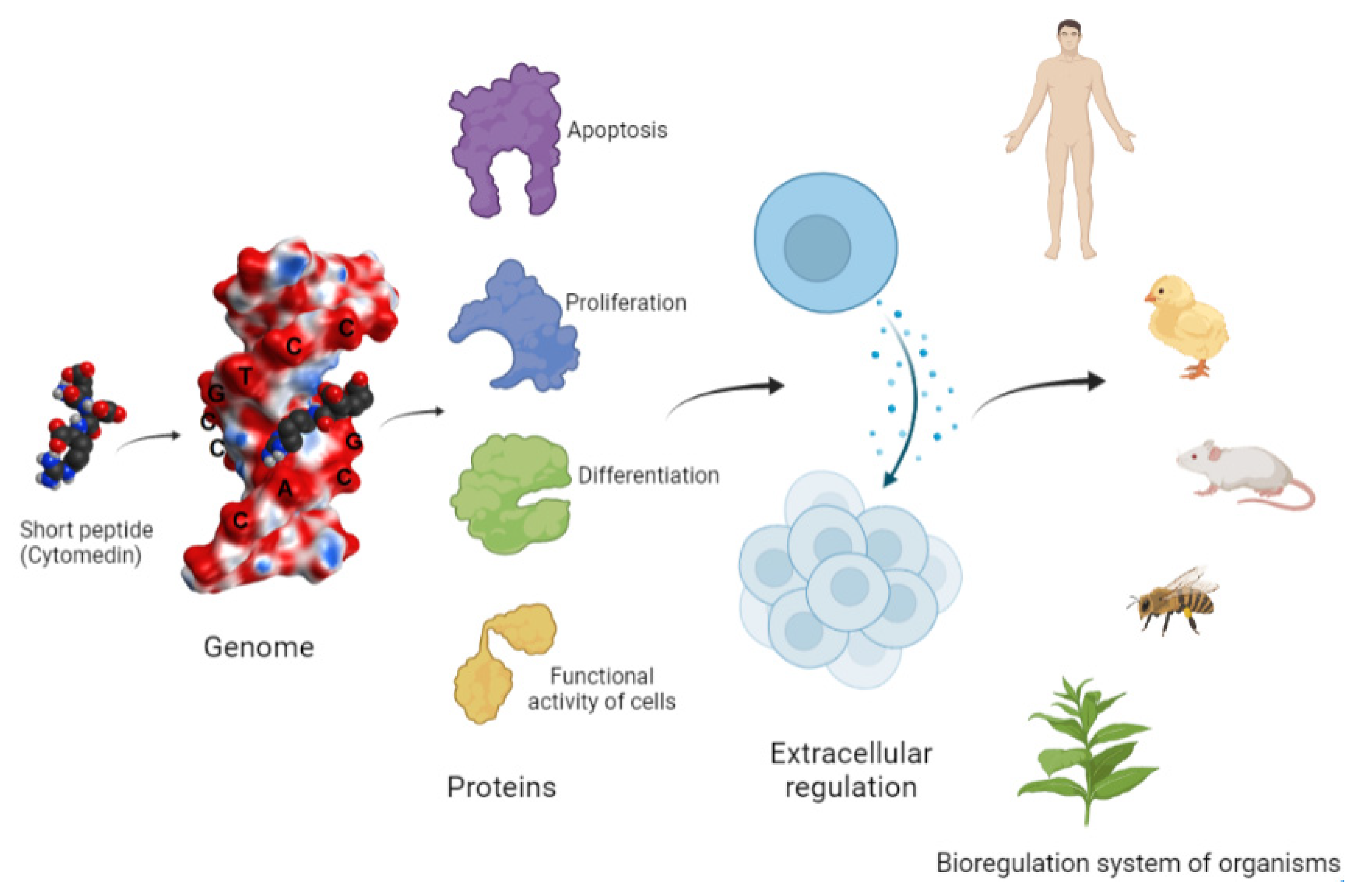
2. Biological Effects of Peptides
3. Peptide Regulation of Gene Expression and Protein Synthesis
3.1. Peptide Regulation of Cell Differentiation
3.2. Peptide Regulation of the Functional Activity of Cells
3.3. Peptide Regulation of Senescence and Apoptosis
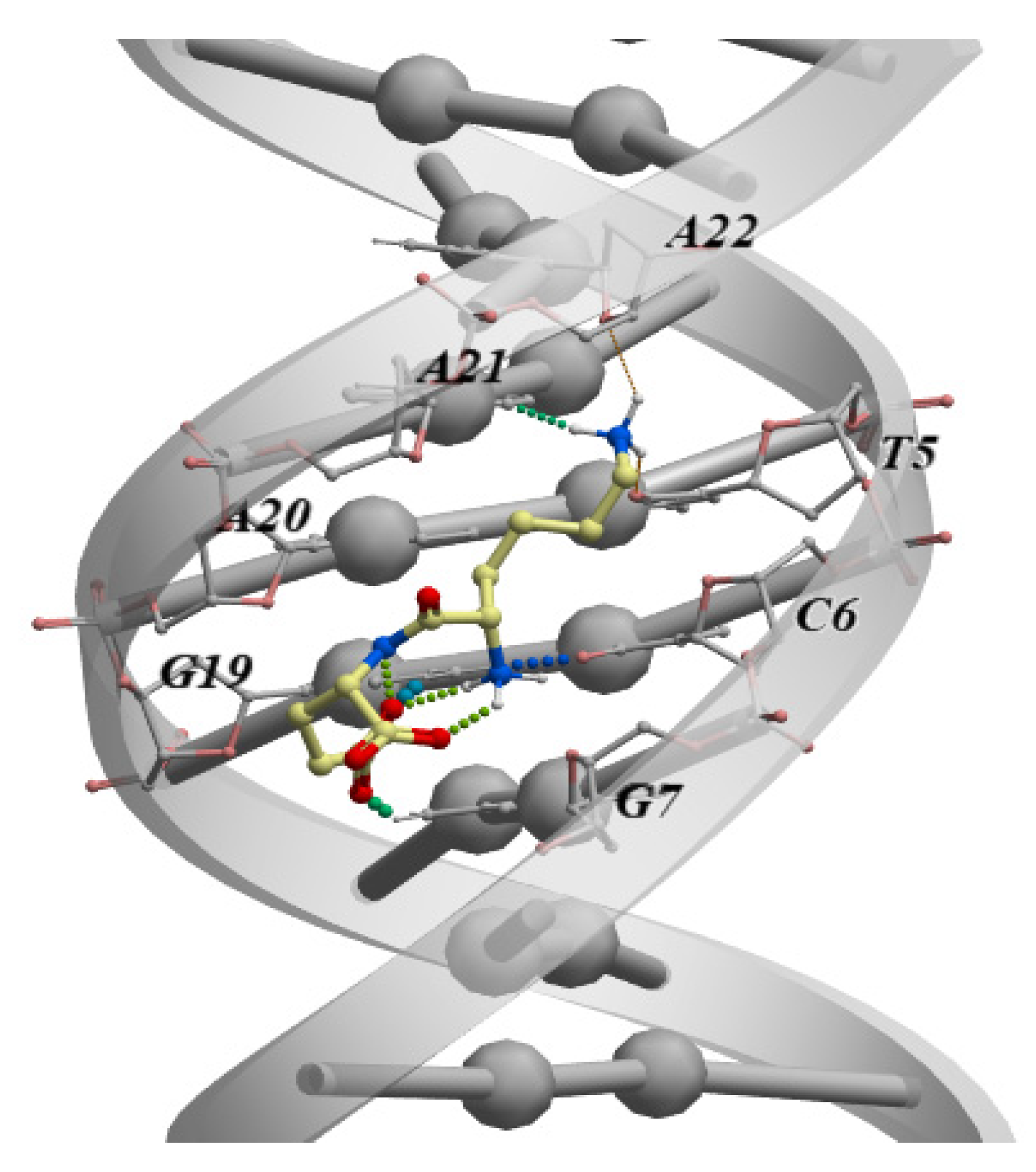
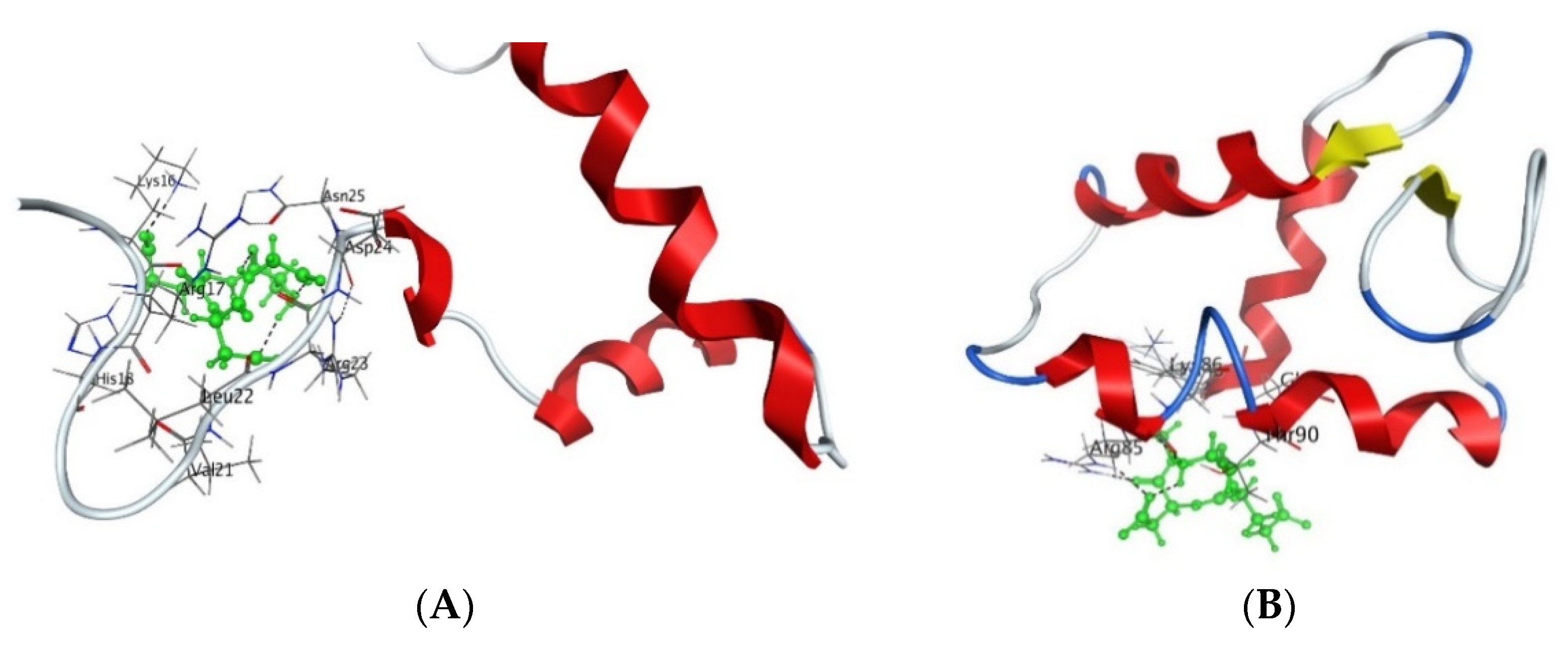
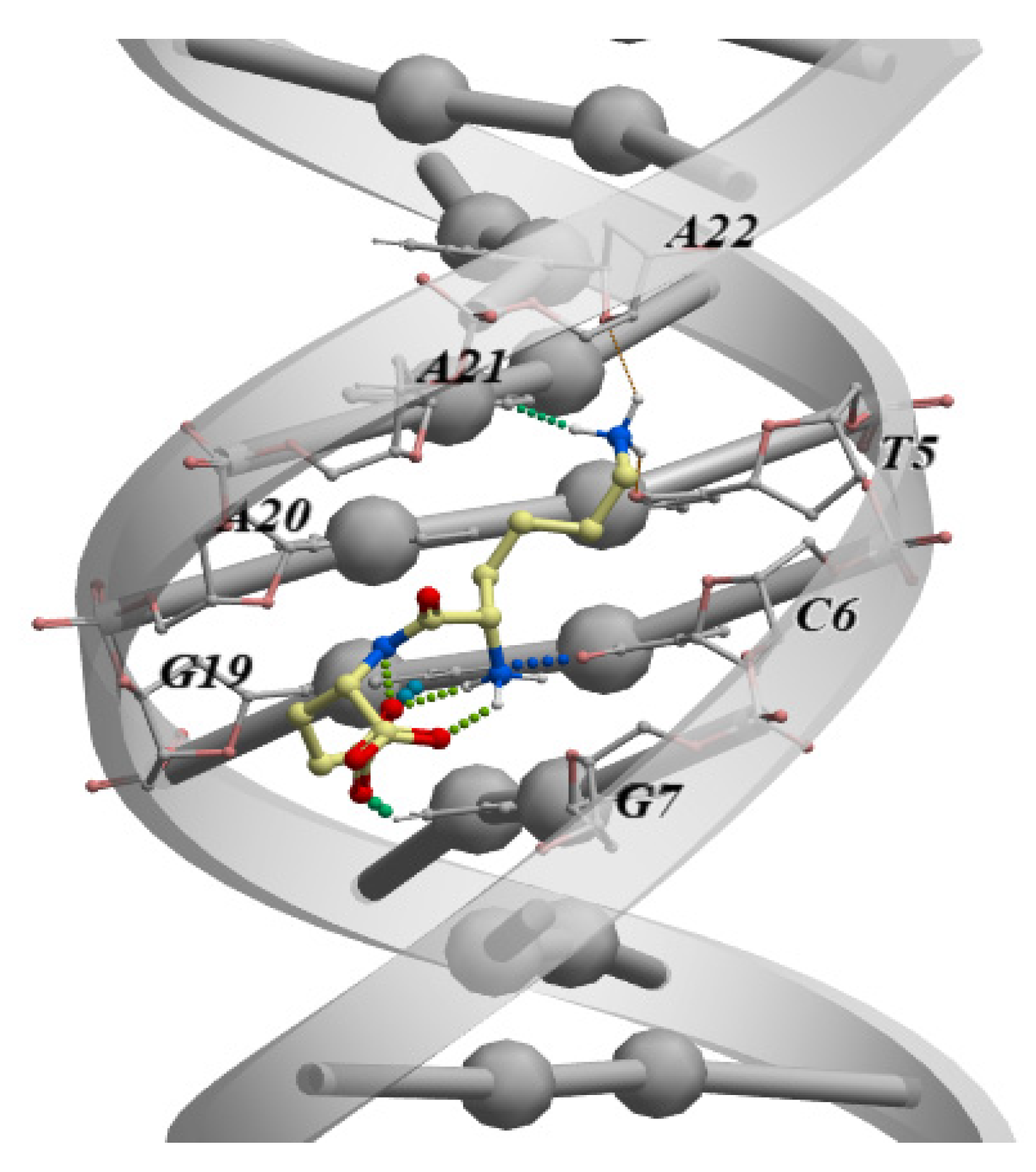
4. Interaction of Short Peptides with DNA and Histone Proteins
4.1. Short Peptides + DNA
4.2. Short Peptides + Histone Proteins
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Apostolopoulos, V.; Bojarska, J.; Chai, T.T.; Elnagdy, S.; Kaczmarek, K.; Matsoukas, J.; New, R.; Parang, K.; Lopez, O.P.; Parhiz, H.; et al. A Global Review on Short Peptides: Frontiers and Perspectives. Molecules 2021, 26, 430. [Google Scholar] [CrossRef]
- IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature and Symbolism for Amino Acids and Peptides. Eur. J. Biochem. 1984, 138, 9–37. [Google Scholar] [CrossRef]
- Nelson, D.; Cox, M. Lehninger Principles of Biochemistry, 4th ed.; Freeman and Company: New York, NY, USA, 2005; p. 1216. [Google Scholar]
- Hoffknecht, B.C.; Worm, D.J.; Bobersky, S.; Prochnow, P.; Bandow, J.E.; Metzler-Nolte, N. Influence of the Multivalency of Ultrashort Arg-Trp-Based Antimicrobial Peptides (AMP) on Their Antibacterial Activity. ChemMedChem 2015, 10, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Tresset, G.; Iliescu, C.; Hauser, C.A.E. Ultrashort Peptide Theranostic Nanoparticles by Microfluidic-Assisted Rapid Solvent Exchange. IEEE Trans NanoBiosci. 2020, 19, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Abuine, R.; Rathnayake, A.U.; Byun, H.G. Biological activity of peptides purified from fish skin hydrolysates. Fish Aquat. Sci. 2019, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- Ciociola, T.; Giovati, L.; Conti, S.; Magliani, W.; Santinoli, C.; Polonelli, L. Natural and synthetic peptides with antifungal activity. Future Med. Chem. 2016, 8, 1413–1433. [Google Scholar] [CrossRef]
- Khavinson, V.K. Peptide medicines: Past, present, future. Klin. Med. 2020, 98, 165–177. [Google Scholar] [CrossRef]
- Lee, A.C.; Harris, J.L.; Khanna, K.K.; Hong, J.H. A Comprehensive Review on Current Advances in Peptide Drug Development and Design. Int. J. Mol. Sci. 2019, 20, 2383. [Google Scholar] [CrossRef] [Green Version]
- Morozov, V.G.; Khavinson, V.K. Natural and synthetic thymic peptides as therapeutics for immune dysfunction. Int. J. Immunopharmacol. 1997, 19, 501–505. [Google Scholar] [CrossRef]
- Morozov, V.G.; Khavinson, V.K. The new class of biological regulators of multicellular systems: Cytomedins. Biol. Bull. Rev. 1983, 96, 339–352. (In Russian) [Google Scholar]
- Morozov, V.G.; Khavinson, V.K. Role of cellular mediators (cytomedins) in regulation genetic activity. Izv. Akad. Nauk SSSR Biol. 1985, 4, 581–587. (In Russian) [Google Scholar]
- Khavinson, V.K.; Malinin, V.V.; Chalisova, N.I.; Grigor’ev, E.I. Tissue-specific action of peptides in tissue culture of rats of various ages. Adv. Gerontol. 2002, 9, 95–100. [Google Scholar] [PubMed]
- Anisimov, V.N.; Khavinson, V.K. Peptide bioregulation of aging: Results and prospects. Biogerontology 2010, 11, 139–149. [Google Scholar] [CrossRef]
- Fedoreyeva, L.I.; Smirnova, T.A.; Kolomijtseva, G.Y.; Khavinson, V.K.; Vanyushin, B.F. Interaction of short peptides with FITC-labeled wheat histones and their complexes with deoxyribooligonucleotides. Biochemistry 2013, 78, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Fedoreyeva, L.I.; Dilovarova, T.A.; Ashapkin, V.V.; Martirosyan, Y.T.; Khavinson, V.K.; Kharchenko, P.N.; Vanyushin, B.F. Short Exogenous Peptides Regulate Expression of CLE, KNOX1, and GRF Family Genes in Nicotiana tabacum. Biochemistry 2017, 82, 521–528. [Google Scholar] [CrossRef]
- Caputi, S.; Trubiani, O.; Sinjari, B.; Trofimova, S.; Diomede, F.; Linkova, N.; Diatlova, A.; Khavinson, V. Effect of short peptides on neuronal differentiation of stem cells. Int. J. Immunapathol. Pharmacol. 2019, 33, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashapkin, V.; Khavinson, V.; Shilovsky, G.; Linkova, N.; Vanuyshin, B. Gene expression in human mesenchymal stem cell aging cultures: Modulation by short peptides. Mol. Biol. Rep. 2020, 47, 4323–4329. [Google Scholar] [CrossRef]
- Ivko, O.M.; Linkova, N.S.; Ilina, A.R.; Sharova, A.A.; Ryzhak, G.A. AEDG peptide regulates human circadian rhythms genes expression during pineal gland accelerated aging. Adv. Gerontol. 2020, 33, 429–435. [Google Scholar]
- Khavinson, V.; Diomede, F.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S.; Sinjari, B. AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism. Molecules 2020, 25, 609. [Google Scholar] [CrossRef] [Green Version]
- Sinjari, B.; Diomede, F.; Khavinson, V.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S. Short peptides protect oral stem cells from ageing. Stem Cell Rev. Reps. 2020, 16, 159–166. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Linkova, N.S.; Umnov, R.S. Peptide KED: Molecular-Genetic Aspects of Neurogenesis Regulation in Alzheimer’s Disease. Bull. Exp. Biol. Med. 2021, 171, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Malinin, V.V. Gerontological Aspects of Genome Peptide Regulation; Karger AG: Basel, Switzerland, 2005; pp. 1–104. [Google Scholar]
- Vanyushin, B.F.; Khavinson, V.K. Epigenetics—A Different Way of Looking at Genetics; Walter, D., Bohm, P., Eds.; Springer: Cham, Switzerland, 2016; Chapter 5; p. 69. [Google Scholar]
- Gianfranceschi, G.L.; Amici, D.; Guglielmi, L. Evidence for the presence in calf thymus of a peptidic factor controlling DNA transcription in vitro. Biochim. Biophys. Acta 1975, 414, 9–19. [Google Scholar] [CrossRef]
- Guglielmi, L.; Gianfranceschi, G.L.; Venanzi, F.; Polzonetti, A.; Amici, D. Specific thymic peptides-DNA interaction. Correlation with the possible stereochemical kinking scheme of DNA. Mol. Biol. Rep. 1979, 4, 195–201. [Google Scholar] [CrossRef]
- Vakhitova, Y.V.; Sadovnikov, S.V.; Borisevich, S.S.; Ostrovskaya, R.U.; Gudasheva, T.A.; Seredenin, S.B. Molecular Mechanism Underlying the Action of Substituted Pro-Gly Dipeptide Noopept. Acta Nat. 2016, 8, 82–89. [Google Scholar] [CrossRef]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine Peptides: Bioactivities and Applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef]
- Pavlicevic, M.; Maestri, E.; Marmiroli, M. Marine Bioactive Peptides—An Overview of Generation, Structure and Application with a Focus on Food Sources. Mar. Drugs 2020, 18, 424. [Google Scholar] [CrossRef]
- Maestri, E.; Pavlicevic, M.; Montorsi, M.; Marmiroli, N. Meta-Analysis for Correlating Structure of Bioactive Peptides in Foods of Animal Origin with Regard to Effect and Stability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 3–30. [Google Scholar] [CrossRef] [Green Version]
- Khavinson, V.K.; Kuznik, B.I.; Ryzhak, G.A. Peptide Geroprotectors—Epigenetic Regulators of Physiological Function of Organism; RGPU named after A.I. Hertsen: Saint Petersburg, Russia, 2014; pp. 1–271. [Google Scholar]
- Vigneault, F.; Guérin, S.L. Regulation of gene expression: Probing DNA-protein interactions in vivo and in vitro. Expert Rev. Proteom. 2005, 2, 705–718. [Google Scholar] [CrossRef]
- Fedoreyeva, L.I.; Kireev, I.I.; Khavinson, V.K.; Vanyushin, B.F. Penetration of Short Fluorescence-Labeled Peptides into the Nucleus in HeLa Cells and in vitro Specific Interaction of the Peptides with Deoxyribooligonucleotides and DNA. Biochemistry 2011, 76, 1210–1219. [Google Scholar] [CrossRef]
- Khavinson, V.; Linkova, N.; Diatlova, A.; Trofimova, S. Peptide Regulation of Cell Differentiation. Stem Cell Rev. Rep. 2020, 16, 118–125. [Google Scholar] [CrossRef]
- Anisimov, S.V.; Bokeler, K.R.; Khavinson, V.K.; Anisimov, V.N. Studies of the Effects of Vilon and Epitalon on Gene Expression in Mouse Heart using DNA-Microarray Technology. Bull. Exp. Biol. Med. 2002, 133, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Tatman, P.D.; Muhonen, E.G.; Wickers, S.T.; Gee, A.O.; Kim, E.S.; Kim, D.H. Self-assembling peptides for stem cell and tissue engineering. Biomater. Sci. 2016, 4, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boopathy, A.V.; Che, P.L.; Somasuntharam, I.; Fiore, V.F.; Cabigas, E.B.; Ban, K.; Brown, M.E.; Narui, Y.; Barker, T.H.; Yoon, Y.S.; et al. The modulation of cardiac progenitor cell function by hydrogel-dependent Notch1 activation. Biomaterials 2014, 35, 8103–8112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennessy, K.M.; Pollot, B.E.; Clem, W.C.; Phipps, M.C.; Sawyer, A.A.; Culpepper, B.K.; Bellis, S.L. The effect of collagen I mimetic peptides on mesenchymal stem cell adhesion and differentiation, and on bone formation at hydroxyapatite surfaces. Biomaterials 2009, 30, 1898–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneda, T.; Yoshida, H.; Nakajima, Y.; Toishi, M.; Nugroho, A.E.; Morita, H. Cyclolinopeptides, cyclic peptides from flaxseed with osteoclast differentiation inhibitory activity. Bioorg. Med. Chem. Lett. 2016, 1, 1760–1761. [Google Scholar] [CrossRef]
- Ma, W.; Jin, G.W.; Gehret, P.M.; Chada, N.C.; Suh, W.H. A novel cell penetrating peptide for the differentiation of human neural stem cells. Biomolecules 2018, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Khavinson, V.K.; Tendler, S.M.; Vanyushin, B.F.; Kasyanenko, N.A.; Kvetnoy, I.M.; Linkova, N.S.; Ashapkin, V.V.; Polyakova, V.O.; Basharina, V.S.; Bernadotte, A. Peptide regulation of gene expression and protein synthesis in bronchial epithelium. Lung 2014, 192, 781–791. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Tendler, S.M.; Kasyanenko, N.A.; Tarnovskaya, S.I.; Linkova, N.S.; Ashapkin, V.V.; Yakutseni, P.P.; Vanyushin, B.F. Tetrapeptide KEDW Interacts with DNA and Regulates Gene Expression. Am. J. Biomed. Sci. 2015, 7, 156–169. [Google Scholar] [CrossRef]
- Shang, N.; Bhullar, K.S.; Hubbard, B.P.; Wu, J. Tripeptide IRW initiates differentiation in osteoblasts via the RUNX2 pathway. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1138–1146. [Google Scholar] [CrossRef]
- Yao, C.; Slamovich, E.B.; Webster, T.J. Enhanced osteoblast functions on anodized titanium with nanotube-like structures. J. Biomed. Mater. Res. 2008, 85, 157–166. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, L.S.; Park, S.W.; Lee, K.; Yun, K.D.; Kim, H.S.; Oh, G.J.; Ji, M.K.; Lim, H.P. Evaluation of osteoblast-like cell viability and differentiation on the Gly-Arg-Gly-Asp-Ser peptide immobilized titanium dioxide nanotube via chemical grafting. J. Nanosci. Nanotechnol. 2016, 16, 1396–1399. [Google Scholar] [CrossRef] [PubMed]
- Otsuki, Y.; Ii, M.; Moriwaki, K.; Okada, M.; Ueda, K.; Asahi, M. W9 peptide enhanced osteogenic differentiation of human adipose-derived stem cells. Biochem. Biophys. Res. Commun. 2018, 495, 904–910. [Google Scholar] [CrossRef]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y. Mechanisms Inspired Targeting Peptides. Adv. Exp. Med. Biol. 2020, 1248, 531–546. [Google Scholar] [CrossRef]
- Kim, K.H.; Son, J.M.; Benayoun, B.A.; Lee, C. The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress. Cell Metab. 2018, 28, 516–524.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, F.; Jin, T.; Drucker, D.J. Proglucagon gene expression is induced by gastrin-releasing peptide in a mouse enteroendocrine cell line. Endocrinology 1996, 137, 3710–3716. [Google Scholar] [CrossRef] [Green Version]
- Khavinson, V.; Popovich, I. Anti-Aging Drugs: From Basic Research to Clinical Practice; Vaiserman, A.M., Ed.; RSC Drug Discovery Series No. 57; RSC Publishing: London UK, 2017; Chapter 20; p. 496. [Google Scholar]
- Calcagno, A.M.; Fostel, J.M.; Reyner, E.L.; Sinaga, E.; Alston, J.T.; Mattes, W.B.; Siahaan, T.J.; Ware, J.A. Effects of an E-cadherin-derived peptide on the gene expression of Caco-2 cells. Pharm. Res. 2004, 21, 2085–2094. [Google Scholar] [CrossRef]
- Stavchanskiĭ, V.V.; Tvorogova, T.V.; Botsina, A.I.; Limborskaia, S.A.; Skvortsova, V.I.; Miasoedov, N.F.; Dergunova, L.V. The effect of semax and its C-end peptide PGP on expression of the neurotrophins and their receptors in the rat brain during incomplete global ischemia. Mol. Biol. 2011, 45, 1026–1035. [Google Scholar]
- Medvedeva, E.V.; Dmitrieva, V.G.; Povarova, O.V.; Limborska, S.A.; Skvortsova, V.I.; Myasoedov, N.F.; Dergunova, L.V. Effect of semax and its C-terminal fragment Pro-Gly-Pro on the expression of VEGF family genes and their receptors in experimental focal ischemia of the rat brain. J. Mol. Neurosci. 2013, 49, 328–333. [Google Scholar] [CrossRef]
- Shadrina, M.; Kolomin, T.; Agapova, T.; Agniullin, Y.; Shram, S.; Slominsky, P.; Lymborska, S.; Myasoedov, N. Comparison of the temporary dynamics of NGF and BDNF gene expression in rat hippocampus, frontal cortex, and retina under Semax action. J. Mol. Neurosci. 2010, 41, 30–35. [Google Scholar] [CrossRef]
- Ostrovskaia, R.U.; Vakhitova, I.V.; Salimgareeva, M.K.; Iamidanov, R.S.; Sadovnikov, S.V.; Kapitsa, I.G.; Seredenin, S.B. On the mechanism of noopept action: Decrease in activity of stress-induced kinases and increase in expression of neutrophines. Eksp. Klin. Farmakol. 2010, 73, 2–5. [Google Scholar]
- Umriukhin, P.E.; Koplik, E.V.; Grivennikov, I.A.; Miasoedov, N.F.; Sudakov, K.V. Gene c-Fos expression in brain of rats resistant and predisposed to emotional stress after intraperitoneal injection of the ACTH(4-10)analog-semax. Zh. Vyssh. Nerv. Deiat. Im. I. P. Pavlov. 2001, 51, 220–227. [Google Scholar]
- Volkova, A.; Shadrina, M.; Kolomin, T.; Andreeva, L.; Limborska, S.; Myasoedov, N.; Slominsky, P. Selank Administration Affects the Expression of Some Genes Involved in GABAergic Neurotransmission. Front. Pharmacol. 2016, 7, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filatova, E.; Kasian, A.; Kolomin, T.; Rybalkina, E.; Alieva, A.; Andreeva, L.; Limborska, S.; Myasoedov, N.; Pavlova, G.; Slominsky, P.; et al. GABA, Selank, and Olanzapine Affect the Expression of Genes Involved in GABAergic Neurotransmission in IMR-32 Cells. Front. Pharmacol. 2017, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, K.L.; Bolotov, I.I.; Linkova, N.S.; Drobintseva, A.O.; Khavinson, V.K.; Dyakonov, M.M.; Kozina, L.S. Molecular aspects of vasoprotective peptide KED activity during atherosclerosis and restenosis. Adv. Gerontol. 2016, 29, 646–650. [Google Scholar]
- Khavinson, V.; Ilina, A.; Kraskovskaya, N.; Linkova, N.; Kolchina, N.; Mironova, E.; Erofeev, A.; Petukhov, M. Neuroprotective Effects of Tripeptides—Epigenetic Regulators in Mouse Model of Alzheimer’s Disease. Pharmaceuticals 2021, 14, 515. [Google Scholar] [CrossRef]
- Kraskovskaya, N.A.; Kukanova, E.O.; Lin’kova, N.S.; Popugaeva, E.A.; Khavinson, V.K. Tripeptides Restore the Number of Neuronal Spines under Conditions of In Vitro Modeled Alzheimer’s Disease. Bull. Exp. Biol. Med. 2017, 163, 550–553. [Google Scholar] [CrossRef]
- Kolomin, T.; Shadrina, M.; Andreeva, L.; Limborska, S.; Myasoedov, N. Expression of inflammation-related genes in mouse spleen under tuftsin analog Selank. Regul. Pept. 2011, 170, 18–23. [Google Scholar] [CrossRef]
- Kolomin, T.A.; Shadrina, M.I.; Slominskiĭ, P.A.; Limborska, S.A.; Myasoedov, N.F. Changes in expression of the genes for chemokines, cytokines, and their receptors in response to selank and its fragments. Genetika 2011, 47, 711–714. [Google Scholar] [CrossRef]
- Medvedeva, E.V.; Dmitrieva, V.G.; Povarova, O.V.; Limborska, S.A.; Skvortsova, V.I.; Myasoedov, N.F.; Dergunova, L.V. The peptide semax affects the expression of genes related to the immune and vascular systems in rat brain focal ischemia: Genome-wide transcriptional analysis. BMC Genom. 2014, 15, 228. [Google Scholar] [CrossRef] [Green Version]
- Medvedeva, E.V.; Dmitrieva, V.G.; Limborska, S.A.; Myasoedov, N.F.; Dergunova, L.V. Semax, an analog of ACTH(4–7), regulates expression of immune response genes during ischemic brain injury in rats. Mol. Genet. Genom. 2017, 292, 635–653. [Google Scholar] [CrossRef]
- Sun, Y.; Coppé, J.P.; Lam, E.W. Cellular Senescence: The Sought or the Unwanted? Trends Mol. Med. 2018, 24, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Miller, B.; Kumagai, H.; Silverstein, A.R.; Flores, M.; Yen, K. Mitochondrial-derived peptides in aging and age-related diseases. Geroscience 2021, 43, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Guichard, G.; Benkirane, N.; Zeder-Lutz, G.; van Regenmortel, M.H.; Briand, J.P.; Muller, S. Antigenic mimicry of natural L-peptides with retro-inverso-peptidomimetics. Proc. Natl. Acad. Sci. USA 1994, 91, 9765–9769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Xu, B. Inspiration from the mirror: D-amino acid containing peptides in biomedical approaches. Biomol. Concepts 2016, 7, 179–187. [Google Scholar] [CrossRef]
- Beydoun, T.; Deloche, C.; Perino, J.; Kirwan, B.A.; Combette, J.M.; Behar-Cohen, F. Subconjunctival injection of XG-102, a JNK inhibitor peptide, in patients with intraocular inflammation: A safety and tolerability study. J. Ocul. Pharmacol. Ther. 2015, 31, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warso, M.A.; Richards, J.M.; Mehta, D.; Christov, K.; Schaeffer, C.; Rae Bressler, L.; Yamada, T.; Majumdar, D.; Kennedy, S.A.; Beattie, C.W.; et al. A first-in-class, first-in-human, phase I trial of p28, a non-HDM2-mediated peptide inhibitor of p53 ubiquitination in patients with advanced solid tumours. Br. J. Cancer 2013, 108, 1061–1070. [Google Scholar] [CrossRef]
- Baar, M.P.; Brandt, R.M.C.; Putavet, D.A.; Klein, J.D.D.; Derks, K.W.J.; Bourgeois, B.R.M.; Stryeck, S.; Rijksen, Y.; van Willigenburg, H.; Feijtel, D.A.; et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 2017, 169, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Khavinson, V.K.; Lin’kova, N.S.; Polyakova, V.O.; Durnova, A.O.; Nichik, T.E.; Kvetnoi, I.M. Peptides regulate expression of signaling molecules in kidney cell cultures during in vitro aging. Bull. Exp. Biol. Med. 2014, 157, 261–264. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Linkova, N.S.; Diatlova, A.S.; Gutop, E.O.; Orlova, O.A. Short peptides: Regulation of skin function during aging. Adv. Gerontol. 2020, 33, 46–54. [Google Scholar]
- Lin’kova, N.S.; Polyakova, V.O.; Trofimov, A.V.; Kvetnoy, I.M.; Khavinson, V.K. Peptidergic regulation of thymocyte differentiation, proliferation, and apoptosis during aging of the thymus. Bull. Exp. Biol. Med. 2011, 151, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.T.; Lo, V.; Chen, J.; Chen, W.C.; Lin, C.Y.; Lin, H.C.; Yang, C.H.; Sheh, L. Synthesis and DNA nicking studies of a novel cyclic peptide: Cyclo[Lys-Trp-Lys-Ahx-]. Bioorg. Med. Chem. 2001, 9, 1493–1498. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Solovyov, A.Y.; Shataeva, L.K. Melting of DNA double strand after binding to geroprotective tetrapeptide. Bull. Exp. Biol. Med. 2008, 146, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Sidorova, N.I.; Nikolaev, V.A.; Surovaia, A.N.; Zhuze, A.L.; Gurskiĭ, G.V. Interaction of a cysteine-containing peptide with DNA. Mol. Biol. 1991, 25, 706–717. [Google Scholar]
- Kolchina, N.; Khavinson, V.; Linkova, N.; Yakimov, A.; Baitin, D.; Afanasyeva, A.; Petukhov, M. Systematic search for structural motifs of peptide binding to double-stranded DNA. Nucleic Acids Res. 2019, 47, 10553–10563. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Tarnovskaya, S.I.; Linkova, N.S.; Chervyakova, N.A.; Nichik, T.E.; Elashkina, E.V.; Chalisova, N.I. Role of peptide bond in the realization of biological activity of short peptides. Bull. Exp. Biol. Med. 2015, 158, 551–554. [Google Scholar] [CrossRef]
- Solovyev, A.Y.; Tarnovskaya, S.I.; Chernova, I.A.; Shataeva, L.K.; Skorik, Y.A. The interaction of amino acids, peptides, and proteins with DNA. Int. J. Biol. Macromol. 2015, 78, 39–45. [Google Scholar] [CrossRef]
- Silanteva, I.A.; Komolkin, A.V.; Morozova, E.A.; Vorontsov-Velyaminov, P.N.; Kasyanenko, N.A. Role of Mono- and Divalent Ions in Peptide Glu-Asp-Arg-DNA Interaction. J. Phys. Chem. B 2019, 123, 1896–1902. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Fedoreyeva, L.I.; Vanyushin, B.F. Site-specific binding of short peptides with DNA modulated eukaryotic endonuclease activity. Bull. Exp. Biol. Med. 2011, 151, 66–70. [Google Scholar] [CrossRef]
- Flock, S.; Bailly, F.; Bailly, C.; Waring, M.J.; Hénichart, J.P.; Colson, P.; Houssier, C. Interaction of two peptide-acridine conjugates containing the SPKK peptide motif with DNA and chromatin. Biomol. Struct. Dyn. 1994, 11, 881–900. [Google Scholar] [CrossRef]
- Bourdouxhe-Housiaux, C.; Colson, P.; Houssier, C.; Bailly, C. Interaction of DNA-threading peptide-amsacrine conjugates with DNA and chromatin. Anticancer Drug Des. 1996, 11, 509–525. [Google Scholar]
- Suzuki, M.; Gerstein, M.; Johnson, T. An NMR study on the DNA-binding SPKK motif and a model for its interaction with DNA. Protein Eng. 1993, 6, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churchill, M.E.; Travers, A.A. Protein motifs that recognize structural features of DNA. Trends Biochem. Sci. 1991, 16, 92–97. [Google Scholar] [CrossRef]
- Geierstanger, B.H.; Volkman, B.F.; Kremer, W.; Wemmer, D.E. Short peptide fragments derived from HMG-I/Y proteins bind specifically to the minor groove of DNA. Biochemistry 1994, 33, 5347–5355. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, J.; Sánchez, M.I.; Valero, J.; de Mendoza, J.; Vázquez, M.E.; Mascareñas, J.L. Sequence-selective DNA binding with cell-permeable oligoguanidinium-peptide conjugates. Chem. Commun. Camb. 2015, 51, 4811–4814. [Google Scholar] [CrossRef] [Green Version]
- Etzion-Fuchs, A.; Todd, D.A.; Singh, M. dSPRINT: Predicting DNA, RNA, ion, peptide and small molecule interaction sites within protein domains. Nucleic Acids Res. 2021, 49, e78. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Lezhava, T.A.; Monaselidze, J.R.; Jokhadze, T.A.; Dvalishvili, N.A.; Bablishvili, N.K.; Trofimova, S.V. Peptide Epitalon activates chromatin at the old age. Neuro. Endocrinol. Lett. 2003, 24, 329–333. [Google Scholar]
- Lezhava, T. Human chromosomes and aging. From 80 to 114 Years. In Nova Biomedical; Nova Science Publisher: New York, NY, USA, 2006; pp. 1–177. [Google Scholar]
- Khavinson, V.; Shataeva, L.; Chernova, A. DNA double-helix binds regulatory peptides similarly to transcription factors. Neuro Endocrinol. Lett. 2005, 26, 237–241. [Google Scholar]
- Kuznik, B.I.; Davydov, S.O.; Popravka, E.S.; Lin’kova, N.S.; Kozina, L.S.; Khavinson, V.K. Epigenetic Mechanisms of Peptide-Driven Regulation and Neuroprotective Protein FKBP1b. Mol. Biol. 2019, 53, 339–348. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | Function/Biological Process | Disorders |
|---|---|---|---|
| MT-ATP6 (ATP6) | ATP synthase subunit a | ATP synthesis, hydrogen ion transport | Neuropathy, ataxia, retinitis pigmentosa, Leigh Syndrome |
| MT-ND1 | NADH-ubiquinone oxidoreductase core subunit 1 | electron transport | Leber hereditary optic atrophy, mitochondrial complex I deficiency, MELAS syndrome, Leber hereditary optic neuropathy, dystonia |
| MT-ND4 | NADH-ubiquinone oxidoreductase chain 4 | Leber hereditary optic neuropathy, modifier of mitochondrial myopathy, encephalopathy, lactic acidosis, stroke-like episodes | |
| MT-CO1 | cytochrome c oxidase subunit 1 | Deafness, non-syndromic sensorineural, mitochondrial, and genetic recurrent myoglobinuria | |
| AK2 | adenylate kinase 2 | cellular energy homeostasis, adenine nucleotide metabolism | Reticular dysgenesis, immunoerythromyeloid hypoplasia, atopic dermatitis, severe combined immunodeficiency |
| HBA1,2 | Hemoglobin subunit alpha | oxygen transport | Erythrocytosis, familial 7 and hemoglobin h disease, Alpha-thalassemia |
| COP1 | E3 ubiquitin-protein ligase COP1 | ubiquitination and proteasomal degradation of target proteins | autism |
| PDLIM5 (Enh2) | PDZ and LIM domain protein 5 | regulation of cardiomyocyte expansion, heart development by scaffolding PKC to the Z-disk region | Nail-Patella syndrome, bipolar disorder |
| HSP90AB1 | heat shock protein 90 family | protein folding and degradation, gastric apoptosis, and inflammation | Larynx cancer, Powassan encephalitis |
| HSPBAP1 (Pass1) | 27 KDa Heat Shock Protein-Associated Protein 1 | cellular stress response, cell growth and differentiation | Renal cell carcinoma, nonpapillary, epithelial recurrent erosion dystrophy |
| гeны HLA | human leucocyte antigens | MHC class II receptor activity/adaptive immunity, host–virus interaction, innate immunity | Rheumatoid arthritis, type 1 diabetes mellitus, cardiac sarcoidosis, measles, berylliosis, granulomatosis with polyangiitis, Halo Nevi, polyarticular juvenile idiopathic arthritis, pityriasis rosea, fetal and neonatal alloimmune thrombocytopenia, Graham-Little-Piccardi-Lassueur syndrome, penicillin allergy, human cytomegalovirus infection, asthma, severe pre-eclampsia, celiac disease 1, adult-onset myasthenia gravis, psoriasis 1, human immunodeficiency virus type 1, severe cutaneous adverse reaction, birdshot chorioretinopathy, celiac disease 1, Creutzfeldt-Jakob disease, sarcoidosis 1, multiple sclerosis |
| Gene | Protein | Function/Biological Process | Disorders |
|---|---|---|---|
| EPS15 | epidermal growth factor receptor pathway substrate 15 | clathrin-mediated endocytosis and development of HGF signaling pathway. | Vaccinia, cataract 8 multiple types, Menkes disease, autosomal recessive spastic paraplegia type 20 |
| MCM10 | minichromosome Maintenance 10 Replication Initiation Factor | cell proliferation, cellular response to DNA damage, DNA replication | Immunodeficiency 80 with or without congenital cardiomyopathy, Baller-Gerold syndrome, Rapadilino syndrome, Rothmund-Thomson syndrome type 2, Fanconi anemia |
| Cul 5 | cullin-5 (CUL-5, vasopressin-activated calcium-mobilizing receptor 1, VACM-1) | core component of multiple SCF-like ECS (Elongin-Cullin 2/5-SOCS-box protein) E3 ubiquitin-protein ligase complexes, which mediate the ubiquitination and subsequent proteasomal degradation of target proteins. | Molluscum contagiosum, Cockayne syndrome, lung cancer |
| APG5L | autophagy protein 5 | autophagic vesicle formation, mitochondrial quality control after oxidative damage, negative regulation of the innate antiviral immune response, lymphocyte development and proliferation, MHC II antigen presentation, adipocyte differentiation, apoptosis | Spinocerebellar ataxia 25, stomatitis |
| ZNF01 | zinc finger protein 1 homolog | nucleic acid binding, DNA-binding transcription factor activity | Retinoblastoma and neuropathy |
| FLJ12848 fis (TNPO3) | transportin-3 | nuclear import signal receptor activity, small GTPase binding | Muscular dystrophy, limb-girdle, autosomal dominant 2 |
| ITPK1 | inositol-tetrakisphosphate 1-kinase | inositol phosphate metabolism, necroptotic process, neural tube development | Neural tube defects |
| SLC7A6 | Y + L amino acid transporter 2 | amino acid transmembrane transport, leukocyte migration, ornithine transport | Lysinuric protein intolerance, hepatocellular carcinoma cystinuria, persistent fetal circulation syndrome |
| KIAA0699 (BICD2) | protein bicaudal D homolog 2 | Golgi-to-ER retrograde transport | Spinal muscular atrophy, lower extremity-predominant, autosomal dominant |
| FLJ10914 (MRGBP) | MRG/MORF4L-binding protein | acetylation of nucleosomal histones H4 and H2A | Colorectal cancer, colorectal adenoma |
| Gdap1 | Ganglioside-induced differentiation-associated protein 1 | glutathione metabolic process, mitochondrial fission, mitochondrial fusion, protein import into peroxisome membrane, protein targeting to mitochondrion | Charcot-Marie-Tooth disease |
| MSTP028 (KCTD10) | BTB/POZ domain-containing adapter for CUL3-mediated RhoA degradation protein 3 | DNA synthesis and cell proliferation | CBlB type of methylmalonic aciduria, occupational dermatitis |
| Gene | Protein | Function/Biological Process | Disorders |
|---|---|---|---|
| RAD21 | Double-strand-break repair protein rad21 homolog | apoptosis, cell cycle, cell division, chromosome partition, DNA damage, DNA repair, mitosis, transcription, transcription regulation | Cornelia de Lange syndrome 4, Mungan syndrome |
| TOP3B | DNA Topoisomerase III Beta | DNA recombination, cellular aging, and maintenance of genome stability | Chromosome 22Q11.2 Duplication Syndrome Prosopagnosia |
| AK2 | Adenylate kinase 2 | cellular energy homeostasis, adenine nucleotide metabolism | Reticular dysgenesis, immunoerythromyeloid hypoplasia, atopic dermatitis, severe combined immunodeficiency |
| Gene Promoters | Nucleotide Sequence |
|---|---|
| 1 ATP promoter | GGGCGGGGGCAACGGTCACCTGATCTGCGGCTGTCGAGGCCGCTGAGGCAGTGGAGGCTG |
| 2 ATP promoter | CAGCTGTCCCAGCGGAAGCGACGAAGGGACGGGACCCGGGAGCCTGGACGAGTCCGAGCG |
| 1 MT-CO1 promoter | CGGGCGGAGTCTTCCTCGATCCCGTGGTGCTCCGCGGCGCGGCCTTGCTCTCTTCCGGTC; |
| 2 MT-CO1 promoter | CATTAACGGGAACAAATTCTCTTTACACAAAGCTCAGGCACATTCAATCAAGGGGAGCCA |
| 3 MT-CO1 promoter | GCCCCCGCCCGCTCCGGAGCAACCCGCGAGCTTACACCGGCTTCTCTCTGTCCTCAGCCC |
| 4 MT-CO1 promoter | GTGATTGGCCCAGAGAGGGGAGGTGACCCCAGGCCCCAGGAAAGGGAGCGAGGACAGCGC |
| HBA1 promoter | GAGTATGGTGCGGAGGCCCTGGAGAGGTGAGGCTCCCTCCCCTGCTCCGACCCGGGCTCC |
| HSP90 promoter | TTCCAGATGCCTGAGGAAACCCAGACCCAAGACCAACCGATGGAGGAGGAGGAGGTTGAG |
| N | Structure and Name of Peptide | Biological Activity | References |
|---|---|---|---|
| Polyfunctional Peptides | |||
| 1 | AED, Cartalax | regulation of cartilage and skin fibroblasts functions, neuronal cell differentiation | [17,18,21,34,35,74] |
| 2 | AEDG, Epitalon | regulation of neuro-immuno-endocrine function, circadian rhythm regulation, retina-protective effect, antioxidant effect, stress-protective effect, geroprotection, activation of skin fibroblasts’ function, differentiation of plant cells, DNA binder | [15,16,19,21,23,34,40,51,78,82,84] |
| 3 | AEDL, Bronchogen | regulation of lung cells’ function and differentiation, differentiation of plant cells, DNA binding | [15,16,24,35,41,84] |
| 4 | EDL, Ovagen | regulation of renal cells’ function, hepatoprotection, DNA binding | [74] |
| 5 | EDR, Pinealon | neuroprotection, activation of stem cells’ neuronal differentiation, antioxidant effect, DNA binding | [15,61,83,84,95] |
| 6 | EW, Thymogen | drug, regulation of immune system function, antioxidant effect, stress-protective effect, geroprotection, DNA binding | [23,42] |
| 7 | KE, Vilon | regulation of immune system function, antioxidant effect, stress-protective effect, geroprotection, activation of stem cells’ neuronal differentiation, activation of plant cells’ differentiation, DNA binding | [16,17,18,21,23,34,35,81,82] |
| 8 | KED, Vesugen | regulation of cardiovascular system function, neuroprotector, activation of stem cells’ neuronal differentiation, activation of skin fibroblasts’ function, geroprotection, DNA binding | [17,18,21,22,34,35,60,75] |
| 9 | KLDL | osteogenic and chondrogenic differentiation of stem cells | [37] |
| 10 | RADA | osteogenic and chondrogenic differentiation of stem cells | [37] |
| Monofunctional peptides | |||
| 11 | AEDR, Cardiogen | regulation of cardiovascular system function | [15,84] |
| 12 | RADA in combination with the Jagged1 | heart progenitor cell differentiation | [37] |
| 13 | KEDG, Testagen | regulation of male reproductive system function | [15,34,35] |
| 14 | AAAAEKAAAAEKAAAAEK | neuroprotection | [35,40] |
| 15 | MEHFPGP, Semax | drug, neuroprotection | [53,54,55,56,57] |
| 16 | TKPRPGP | neuroprotection | [58,59] |
| 17 | IKVAV | stem cells’ neuronal differentiation | [36] |
| 18 | IRW | osteogenic differentiation of stem cells | [35,43] |
| 19 | GRGDS | osteogenic differentiation of stem cells | [35,44,45] |
| 20 | YCWSQYLCY | osteogenic differentiation of stem cells | [46] |
| 21 | AcSDKP | activation of skin fibroblasts’ function | [75] |
| 22 | TKPRPGP | immunoprotection | [58,59] |
| 23 | Ac-SHAVSS-NH2, HAV | regulation of the gene expression involved in E-cadherin synthesis | [52] |
| Peptides with unknown biological function | |||
| 24 | cyclo[Lys-Trp-Lys-Ahx-] | DNA binding | [77] |
| 25 | peptide dimer KGVCV-N2H2Dns2 | DNA binding | [79] |
| 26 | PRGRP | DNA binding | [89] |
| 27 | PRGRPKK | DNA binding | [89] |
| 28 | RGR | DNA binding | [89] |
| 29 | SPKK | DNA binding | [85,86,87] |
| 30 | SPRKSPRK | DNA binding | [89] |
| 31 | TPKRPRGRPKK | DNA binding | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khavinson, V.K.; Popovich, I.G.; Linkova, N.S.; Mironova, E.S.; Ilina, A.R. Peptide Regulation of Gene Expression: A Systematic Review. Molecules 2021, 26, 7053. https://doi.org/10.3390/molecules26227053
Khavinson VK, Popovich IG, Linkova NS, Mironova ES, Ilina AR. Peptide Regulation of Gene Expression: A Systematic Review. Molecules. 2021; 26(22):7053. https://doi.org/10.3390/molecules26227053
Chicago/Turabian StyleKhavinson, Vladimir Khatskelevich, Irina Grigor’evna Popovich, Natalia Sergeevna Linkova, Ekaterina Sergeevna Mironova, and Anastasiia Romanovna Ilina. 2021. "Peptide Regulation of Gene Expression: A Systematic Review" Molecules 26, no. 22: 7053. https://doi.org/10.3390/molecules26227053
APA StyleKhavinson, V. K., Popovich, I. G., Linkova, N. S., Mironova, E. S., & Ilina, A. R. (2021). Peptide Regulation of Gene Expression: A Systematic Review. Molecules, 26(22), 7053. https://doi.org/10.3390/molecules26227053