
Synergistic Effect of Biphasic Calcium Phosphate and Platelet-Rich Fibrin Attenuate Markers for Inflammation and Osteoclast Differentiation by Suppressing NF-κB/MAPK Signaling Pathway in Chronic Periodontitis

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Study Design
2.3. Inclusion and Exclusion Criteria
2.4. Preparation of PRF and BCP
2.5. Monocytes Isolation from Whole Blood
2.6. Osteoclasts Generation and Differentiation
2.7. TRAP Activity Assay
2.8. Measurement of Proinflammatory Cytokines by ELISA
2.9. Western Blot Analysis
2.10. RNA Isolation and Reverse Transcription-Polymerase Chain Reaction
2.11. Statistical Analysis
3. Results
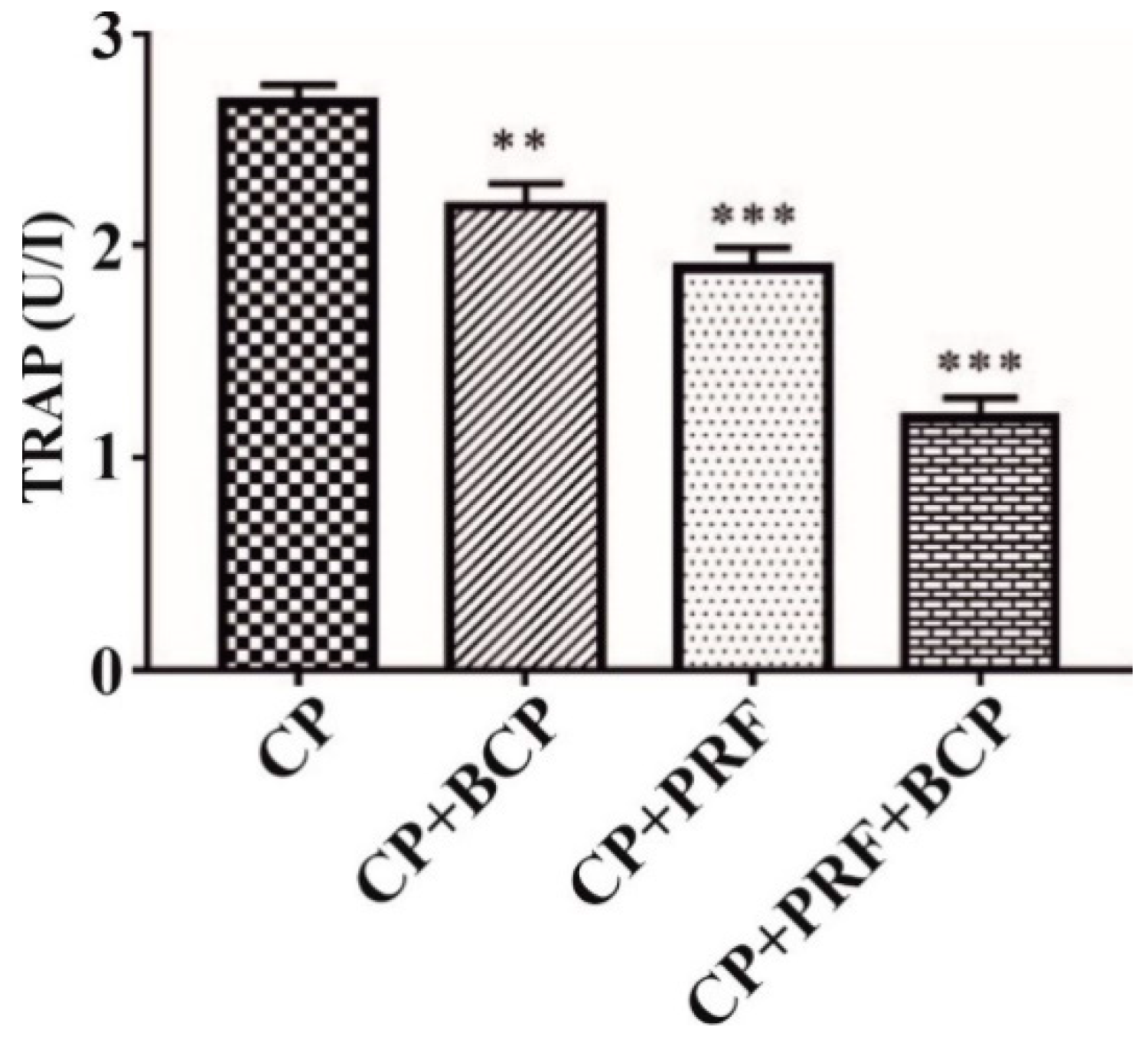
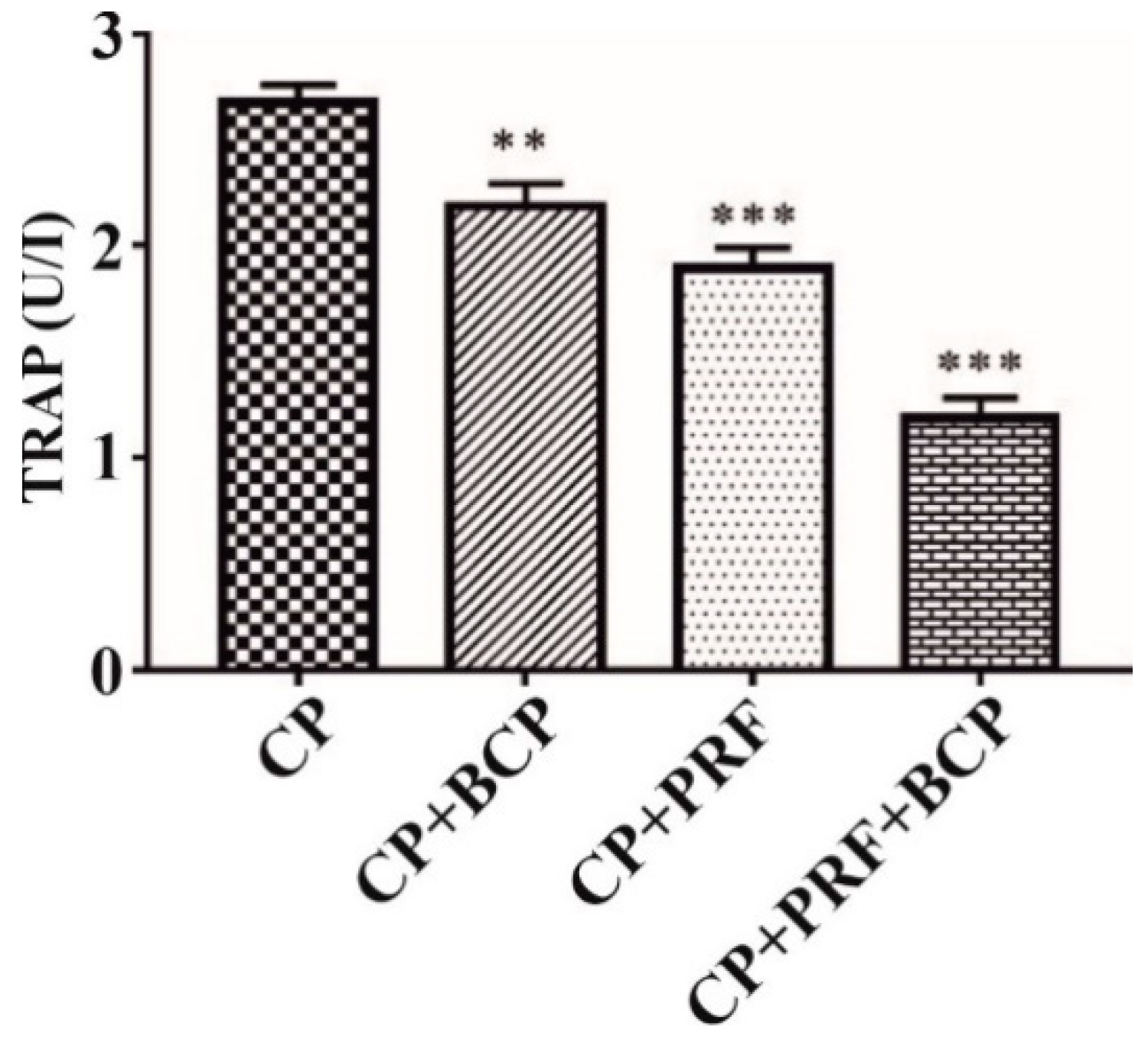
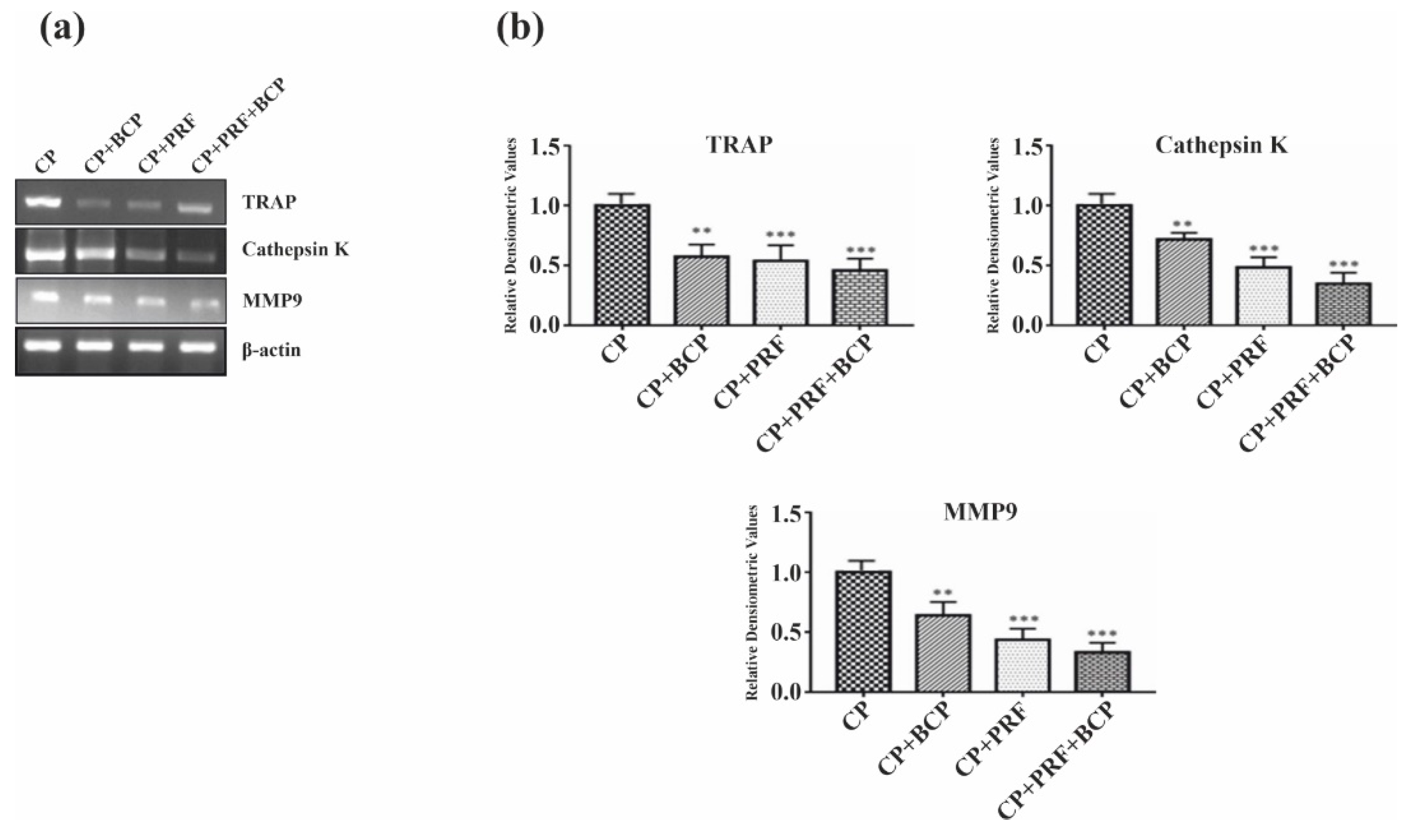
3.1. PRF/BCP Decreases TRAP Activity
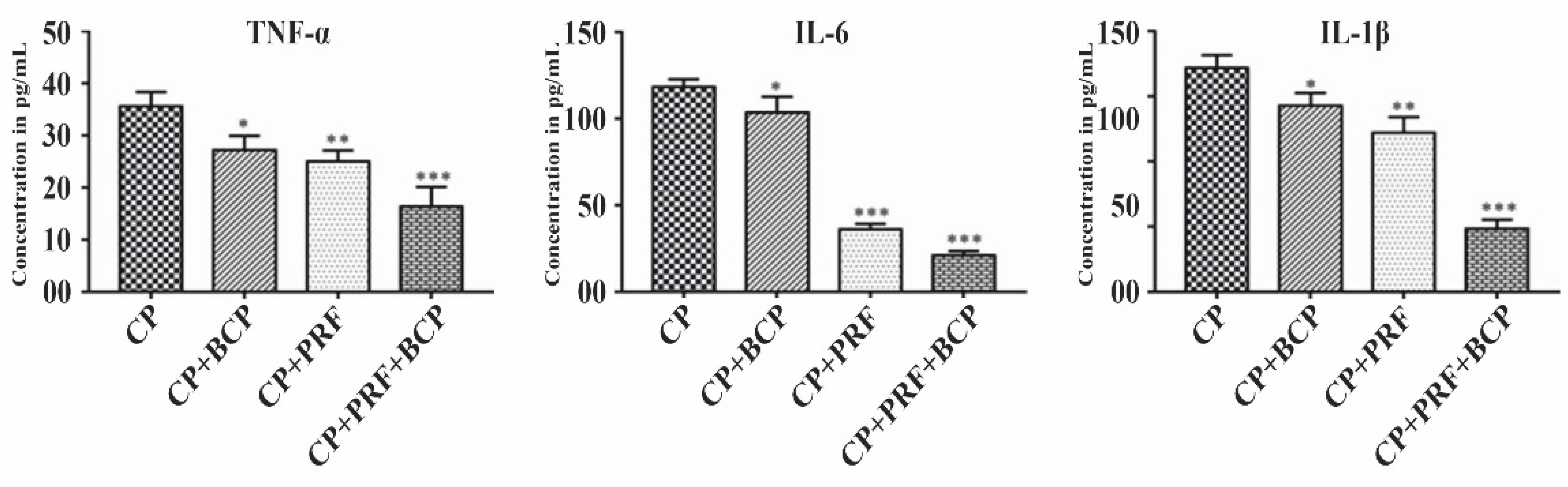
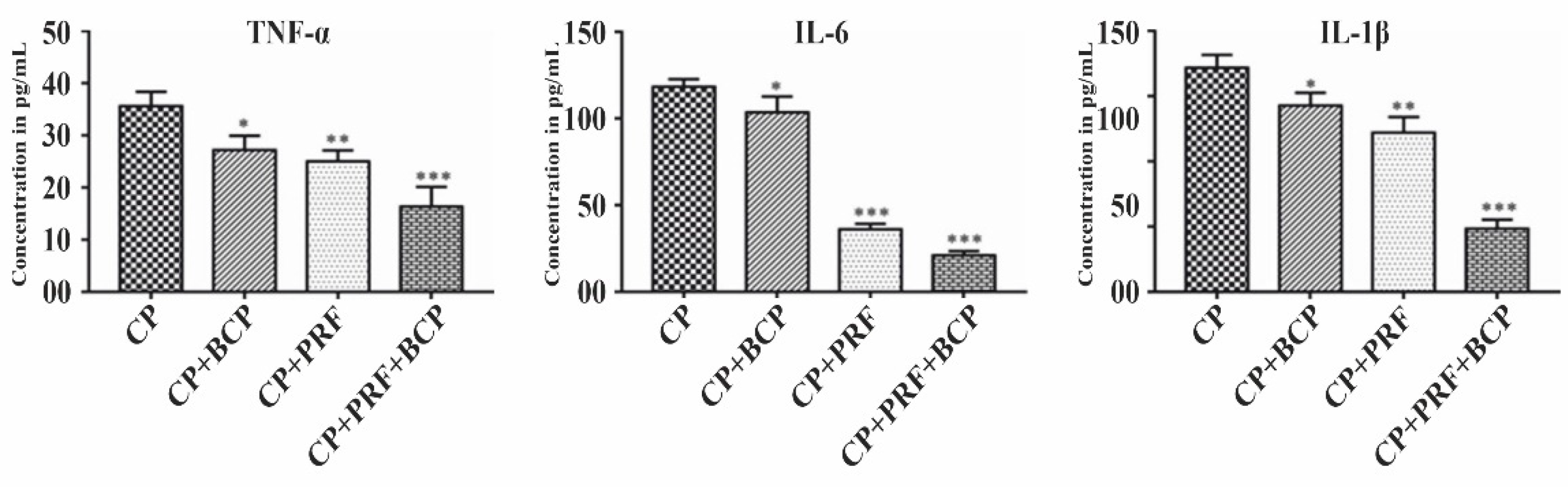
3.2. PRF/BCP Attenuates Proinflammatory Cytokines
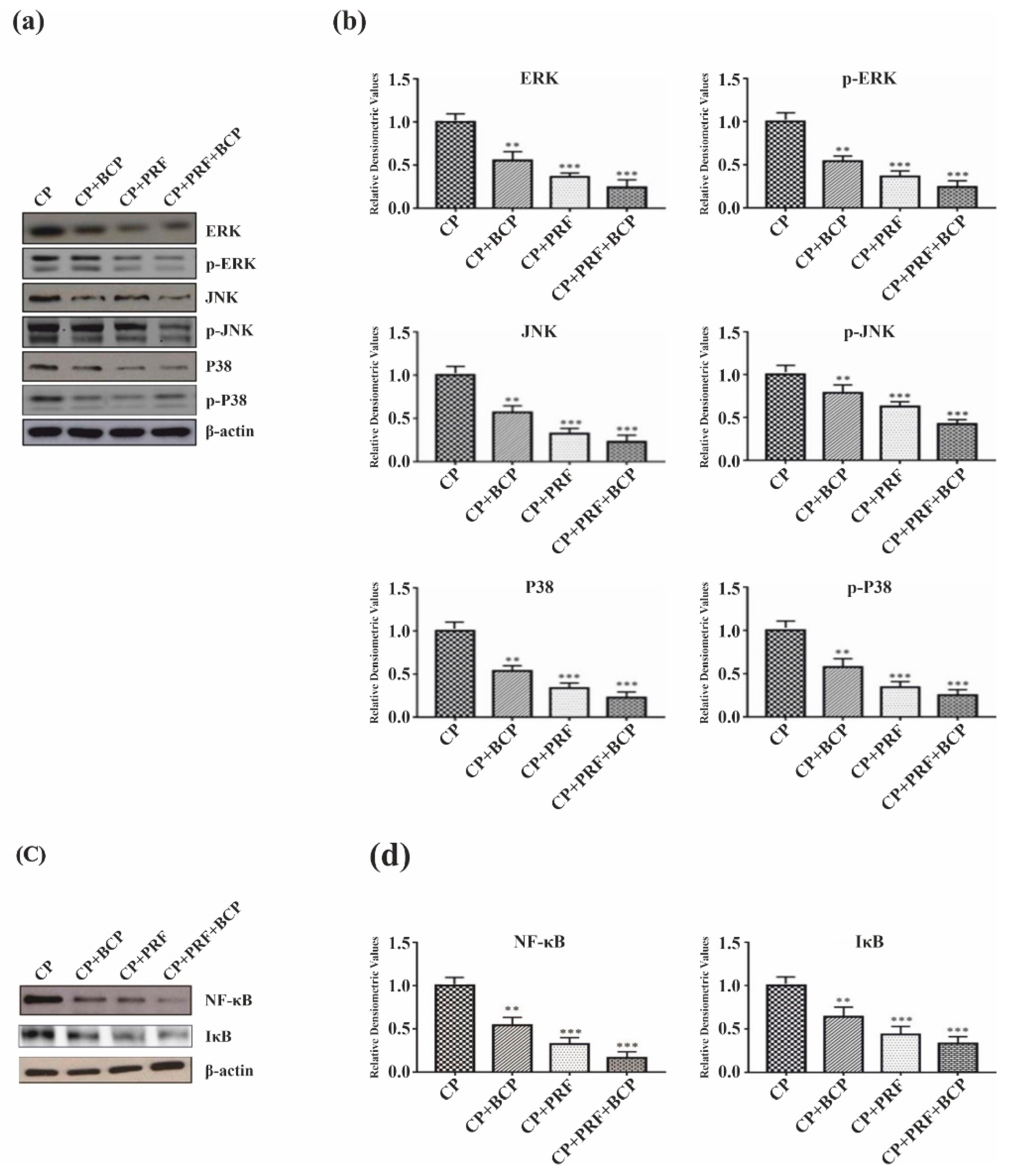
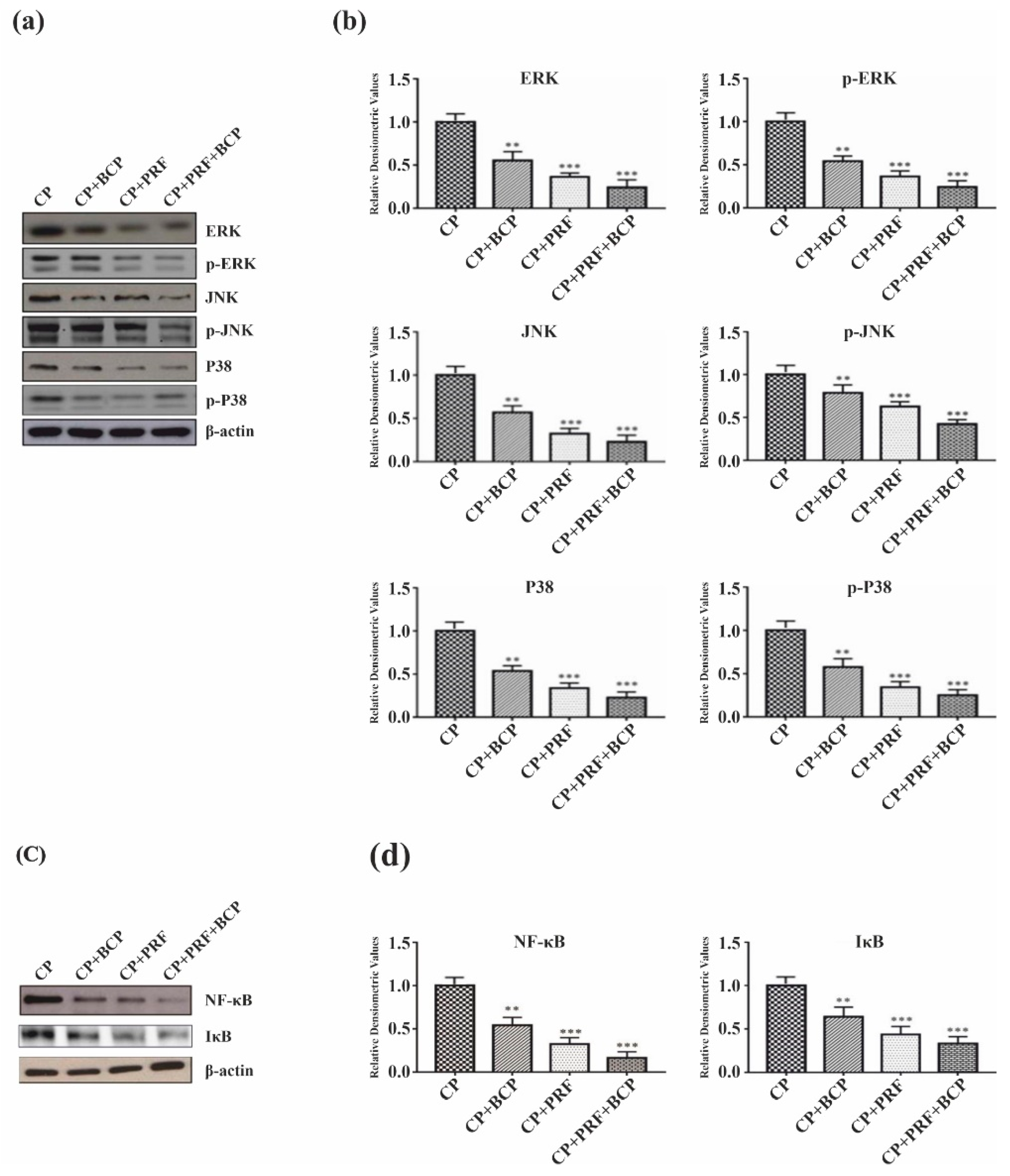
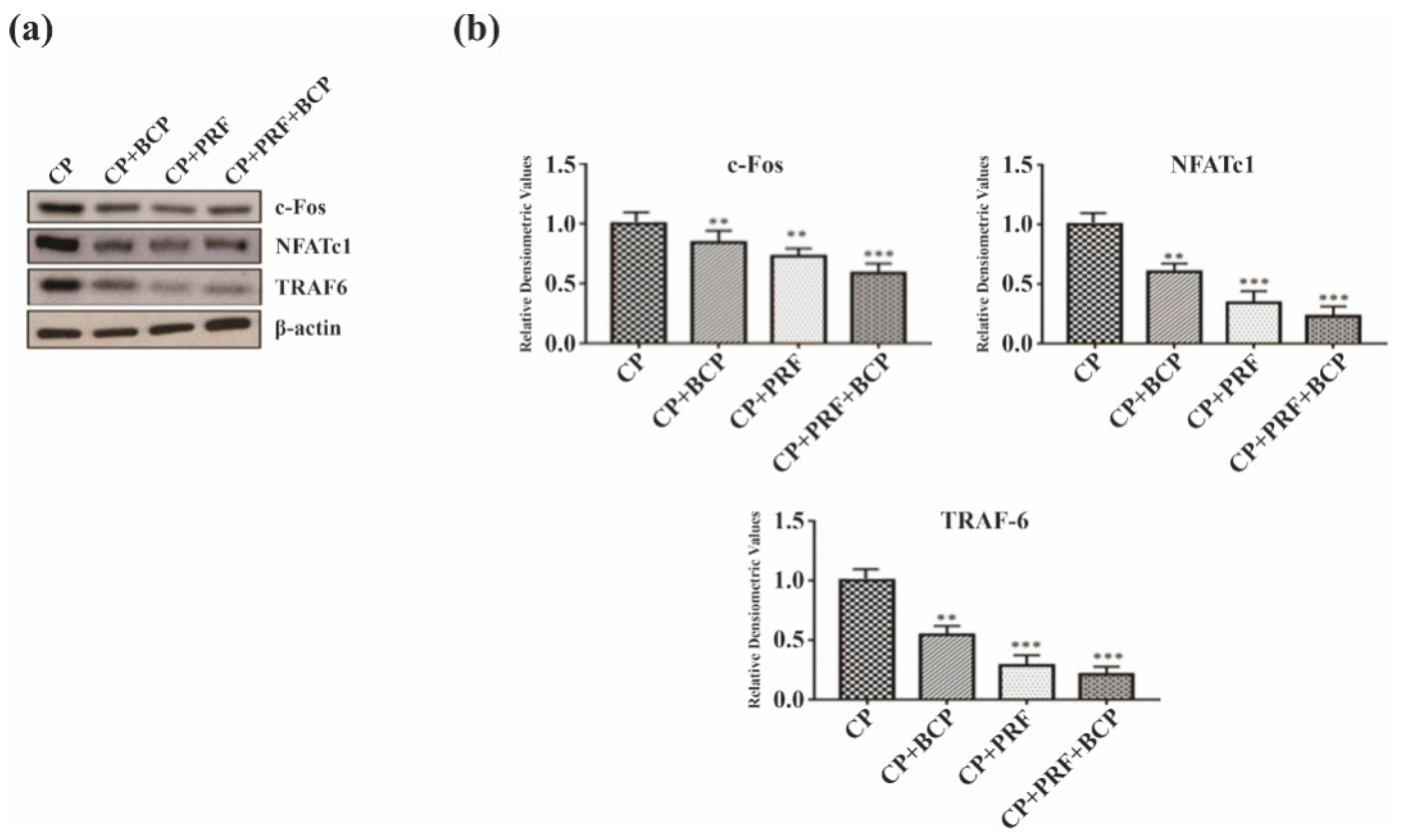
3.3. PRF/BCP Regulates Periodontal Inflammation and Osteoclastogenesis by Inhibiting MAPK Signaling and NF-κB Pathways
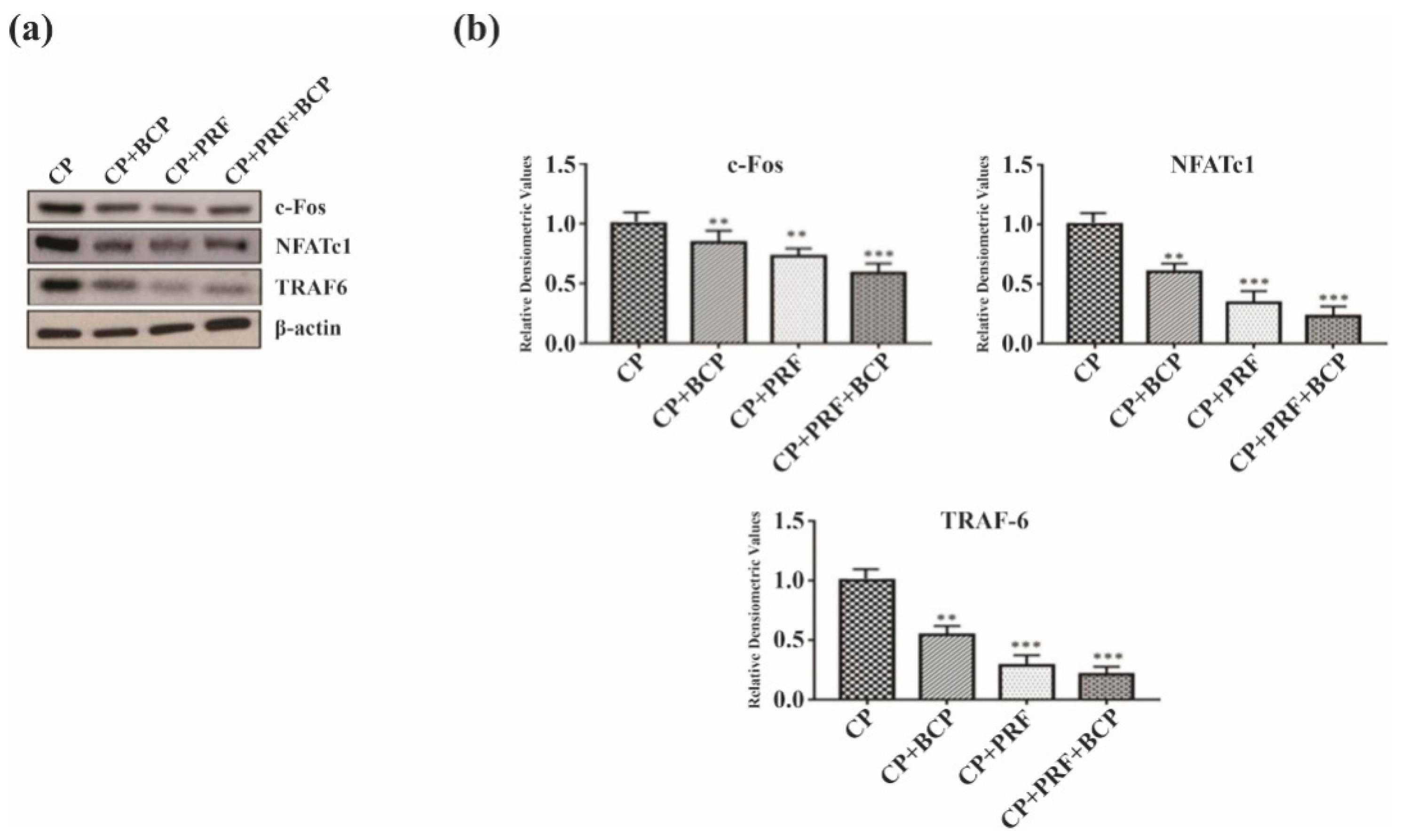
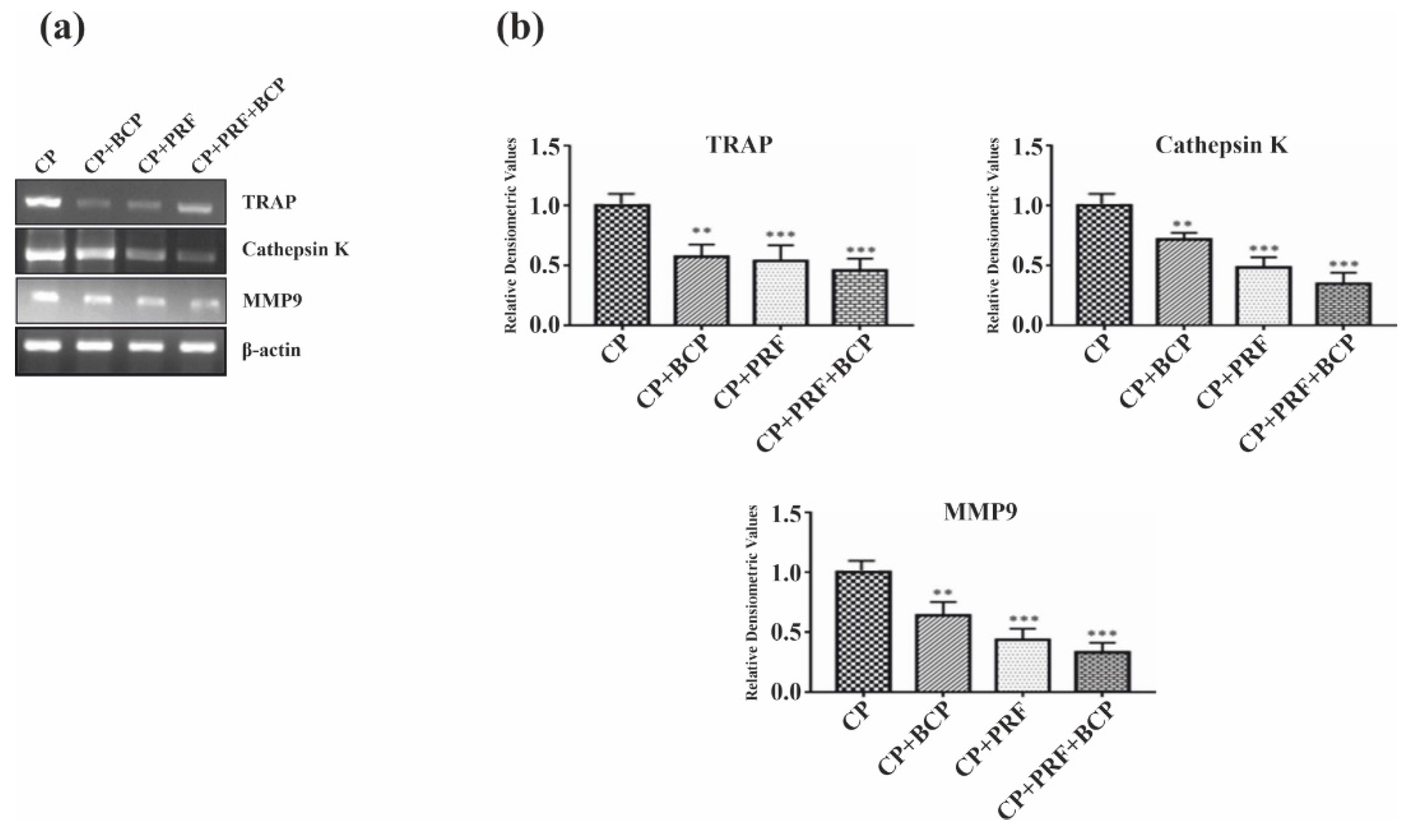
3.4. PRF/BCP Inhibits Osteoclastogenic Transcription Factors and Osteoclast Marker Genes
4. Discussion
Limitations of the Current Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [Green Version]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef] [PubMed]
- Basson, M.A. Signaling in Cell Differentiation and Morphogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corre, I.; Paris, F.; Huot, J. The p38 pathway, a major pleiotropic cascade that transduces stress and metastatic signals in endothelial cells. Oncotarget 2017, 8, 55684–55714. [Google Scholar] [CrossRef]
- Dhanasekaran, D.N.; Reddy, E.P. JNK-signaling: A multiplexing hub in programmed cell death. Genes Cancer 2017, 8, 682–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Yang, J.; Song, P. Correlation of ERK/MAPK signaling pathway with proliferation and apoptosis of colon cancer cells. Oncol. Lett. 2019, 17, 2266–2270. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current Understanding of RANK Signaling in osteoclast differentiation and Maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, N. Regulation of NFATc1 in osteoclast differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Ausenda, F.; Rasperini, G.; Acunzo, R.; Gorbunkova, A.; Pagni, G. New Perspectives in the Use of Biomaterials for Periodontal Regeneration. Materials 2019, 12, 2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ruan, Z.; Shen, M.; Tan, L.; Huang, W.; Wang, L.; Huang, Y. Clinical effect of platelet-rich fibrin on the preservation of the alveolar ridge following tooth extraction. Exp. Ther. Med. 2018, 15, 2277–2286. [Google Scholar] [CrossRef] [Green Version]
- Saberianpour, S.; Heidarzadeh, M.; Geranmayeh, M.H.; Hosseinkhani, H.; Rahbarghazi, R.; Nouri, M. Tissue engineering strategies for the induction of angiogenesis using biomaterials. J. Biol. Eng. 2018, 12, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witek, L.; Tian, H.; Tovar, N.; Torroni, A.; Neiva, R.; Gil, L.F.; Coelho, P.G. The effect of platelet-rich fibrin exudate addition to porous poly(lactic-co-glycolic acid) scaffold in bone healing: An in vivo study. J. Biomed. Mater. Res. B Appl. Biomater. 2020, 108, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Strauss, F.-J.; Nasirzade, J.; Kargarpoor, Z.; Stähli, A.; Gruber, R. Effect of platelet-rich fibrin on cell proliferation, migration, differentiation, inflammation, and osteoclastogenesis: A systematic review of in vitro studies. Clin. Oral Investig. 2020, 24, 569–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kargarpour, Z.; Nasirzade, J.; Strauss, F.J.; Di Summa, F.; Hasannia, S.; Müller, H.-D.; Gruber, R. Platelet-rich fibrin suppresses in vitro osteoclastogenesis. J. Periodontol. 2020, 91, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.N.; Pereira, M.M.; Goes, A.M.; Leite, M.F. Effect of biphasic calcium phosphate on human macrophage functions in vitro. J. Biomed. Mater. Res. A 2003, 65, 475–481. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, S.; Zhou, W.; Fu, H.; Qian, L.; Miron, R.J. Addition of a Synthetically Fabricated Osteoinductive Biphasic Calcium Phosphate Bone Graft to BMP2 Improves New Bone Formation. Clin. Implant Dent. Relat. Res. 2016, 18, 1238–1247. [Google Scholar] [CrossRef]
- Cordonnier, T.; Langonné, A.; Sohier, J.; Layrolle, P.; Rosset, P.; Sensébé, L.; Deschaseaux, F. Consistent osteoblastic differentiation of human mesenchymal stem cells with bone morphogenetic protein 4 and low serum. Tissue Eng. Part C Methods 2011, 17, 249–259. [Google Scholar] [CrossRef]
- Jeong, J.; Kim, J.H.; Shim, J.H.; Hwang, N.S.; Heo, C.Y. Bioactive calcium phosphate materials and applications in bone regeneration. Biomater. Res. 2019, 23, 4. [Google Scholar] [CrossRef] [Green Version]
- Balaguer, T.; Boukhechba, F.; Clavé, A.; Bouvet-Gerbettaz, S.; Trojani, C.; Michiels, J.-F.; Laugier, J.-P.; Bouler, J.-M.; Carle, G.F.; Scimeca, J.-C.; et al. Biphasic calcium phosphate microparticles for bone formation: Benefits of combination with blood clot. Tissue Eng. Part A 2010, 16, 3495–3505. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Mahendra, J.; Samuel, S.; Govindraj, J.; Loganathan, T.; Vashum, Y.; Mahendra, L.; Krishnamoorthy, T. Platelet-rich fibrin/biphasic calcium phosphate impairs osteoclast differentiation and promotes apoptosis by the intrinsic mitochondrial pathway in chronic periodontitis. J. Periodontol. 2019, 90, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, M.-B.; Kim, C.; Hwang, J.-K. Inhibitory Effects of Panduratin A on Periodontitis-Induced Inflammation and Osteoclastogenesis through Inhibition of MAPK Pathways In Vitro. J. Microbiol. Biotechnol. 2018, 28, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouler, J.M.; Pilet, P.; Gauthier, O.; Verron, E. Biphasic calcium phosphate ceramics for bone reconstruction: A review of biological response. Acta Biomater. 2017, 53, 1–12. [Google Scholar] [CrossRef]
- Armitage, G.C. Development of a classification system for periodontal diseases and conditions. Ann. Periodontol. 1999, 4, 1–6. [Google Scholar] [CrossRef]
- Kylmäoja, E.; Nakamura, M.; Turunen, S.; Patlaka, C.; Andersson, G.; Lehenkari, P.; Tuukkanen, J. Peripheral blood monocytes show increased osteoclast differentiation potential compared to bone marrow monocytes. Heliyon 2018, 4, e00780. [Google Scholar] [CrossRef] [Green Version]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Irwin, C.R.; Myrillas, T.T. The role of IL-6 in the pathogenesis of periodontal disease. Oral Dis. 1998, 4, 43–47. [Google Scholar] [CrossRef]
- Isola, G.; Lo Giudice, A.; Polizzi, A.; Alibrandi, A.; Murabito, P.; Indelicato, F. Identification of the different salivary Interleukin-6 profiles in patients with periodontitis: A cross-sectional study. Arch. Oral Biol. 2021, 122, 104997. [Google Scholar] [CrossRef]
- Assuma, R.; Oates, T.; Cochran, D.; Amar, S.; Graves, D.T. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J. Immunol. 1998, 160, 403–409. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of osteoclast differentiation by Cytokine Networks. Immune Netw. 2018, 18, e8. [Google Scholar] [CrossRef] [PubMed]
- Matarese, G.; Currò, M.; Isola, G.; Caccamo, D.; Vecchio, M.; Giunta, M.L.; Ramaglia, L.; Cordasco, G.; Williams, R.C.; Ientile, R. Transglutaminase 2 up-regulation is associated with RANKL/OPG pathway in cultured HPDL cells and THP-1-differentiated macrophages. Amino Acids 2015, 47, 2447–2455. [Google Scholar] [CrossRef]
- Lee, K.; Seo, I.; Choi, M.; Jeong, D. Roles of Mitogen-Activated Protein Kinases in Osteoclast Biology. Int. J. Mol. Sci. 2018, 19, 3004. [Google Scholar] [CrossRef] [Green Version]
- Thouverey, C.; Caverzasio, J. Focus on the p38 MAPK signaling pathway in bone development and maintenance. Bonekey Rep. 2015, 4, 711. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Bal, H.S.; Desta, T.; Behl, Y.; Graves, D.T. Tumor necrosis factor-alpha mediates diabetes-enhanced apoptosis of matrix-producing cells and impairs diabetic healing. Am. J. Pathol. 2006, 168, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Pacios, S.; Andriankaja, O.; Kang, J.; Alnammary, M.; Bae, J.; de Brito Bezerra, B.; Schreiner, H.; Fine, D.H.; Graves, D.T. Bacterial infection increases periodontal bone loss in diabetic rats through enhanced apoptosis. Am. J. Pathol. 2013, 183, 1928–1935. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-W.; Son, Y.-J.; Yun, J.-M.; Kim, S.H. Fisetin Inhibits osteoclast differentiation via Downregulation of p38 and c-Fos-NFATc1 Signaling Pathways. Evid.-Based Complement. Altern. Med. 2012, 2012, 810563. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, G.C.N.; Kajiya, M.; Nakanishi, T.; Ohta, K.; Rosalen, P.L.; Groppo, F.C.; Ernst, C.W.O.; Boyesen, J.L.; Bartlett, J.D.; Stashenko, P.; et al. Inhibition of matrix metalloproteinase-9 activity by doxycycline ameliorates RANK ligand-induced osteoclast differentiation in vitro and in vivo. Exp. Cell Res. 2011, 317, 1454–1464. [Google Scholar] [CrossRef] [Green Version]
- Klar, R.M.; Duarte, R.; Dix-Peek, T.; Dickens, C.; Ferretti, C.; Ripamonti, U. Calcium ions and osteoclastogenesis initiate the induction of bone formation by coral-derived macroporous constructs. J. Cell. Mol. Med. 2013, 17, 1444–1457. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res. Ther. 2007, 9, S1. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Rotwein, P. Akt promotes BMP2-mediated osteoblast differentiation and bone development. J. Cell Sci. 2009, 122, 716–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katarivas Levy, G.; Birch, M.A.; Brooks, R.A.; Neelakantan, S.; Markaki, A.E. Stimulation of Human Osteoblast Differentiation in Magneto-Mechanically Actuated Ferromagnetic Fiber Networks. J. Clin. Med. 2019, 8, 1522. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of Osteoblast and Odontoblast Differentiation by RUNX2. J. Oral Biosci. 2010, 52, 22–25. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′ to 3′) | Reverse Primer Sequence (5′ to 3′) | Size (bp) |
|---|---|---|---|
| TRAP | CCAATGCCAAAGAGATCGCC | TCTGTGCAGAGACGTTGCCAAG | 216 |
| CAT-K | CCGCAGTAATGACACCCTTT | AAGGCATTGGTCATGTAGCC | 258 |
| MMP-9 | CTCTGGAGGTTCGACGTG | GTCCACCTGGTTCAACTCAC | 183 |
| β-actin | TCACCCACACTGTGCCCATCTACGA | CAGCGGAACCGCTCATTGCCAATGG | 141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; Mahendra, J.; Mahendra, L.; Abdulkarim, H.H.; Sayed, M.; Mugri, M.H.; Ahmad, Z.H.; Bhati, A.K.; Faqehi, H.H.; Algregri, W.O.; et al. Synergistic Effect of Biphasic Calcium Phosphate and Platelet-Rich Fibrin Attenuate Markers for Inflammation and Osteoclast Differentiation by Suppressing NF-κB/MAPK Signaling Pathway in Chronic Periodontitis. Molecules 2021, 26, 6578. https://doi.org/10.3390/molecules26216578
Kumar A, Mahendra J, Mahendra L, Abdulkarim HH, Sayed M, Mugri MH, Ahmad ZH, Bhati AK, Faqehi HH, Algregri WO, et al. Synergistic Effect of Biphasic Calcium Phosphate and Platelet-Rich Fibrin Attenuate Markers for Inflammation and Osteoclast Differentiation by Suppressing NF-κB/MAPK Signaling Pathway in Chronic Periodontitis. Molecules. 2021; 26(21):6578. https://doi.org/10.3390/molecules26216578
Chicago/Turabian StyleKumar, Anil, Jaideep Mahendra, Little Mahendra, Hesham H. Abdulkarim, Mohammed Sayed, Maryam H. Mugri, Zeeshan Heera Ahmad, Ashok Kumar Bhati, Hadeel Hussain Faqehi, Waleed Omar Algregri, and et al. 2021. "Synergistic Effect of Biphasic Calcium Phosphate and Platelet-Rich Fibrin Attenuate Markers for Inflammation and Osteoclast Differentiation by Suppressing NF-κB/MAPK Signaling Pathway in Chronic Periodontitis" Molecules 26, no. 21: 6578. https://doi.org/10.3390/molecules26216578
APA StyleKumar, A., Mahendra, J., Mahendra, L., Abdulkarim, H. H., Sayed, M., Mugri, M. H., Ahmad, Z. H., Bhati, A. K., Faqehi, H. H., Algregri, W. O., Varadarajan, S., Balaji, T. M., Baeshen, H. A., & Patil, S. (2021). Synergistic Effect of Biphasic Calcium Phosphate and Platelet-Rich Fibrin Attenuate Markers for Inflammation and Osteoclast Differentiation by Suppressing NF-κB/MAPK Signaling Pathway in Chronic Periodontitis. Molecules, 26(21), 6578. https://doi.org/10.3390/molecules26216578