
The Antiproliferative and Apoptosis-Inducing Effects of the Red Macroalgae Gelidium latifolium Extract against Melanoma Cells

 , , ,
, , , 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Macroalgal Material
2.2. Chemicals and Reagents
2.3. Preparation of Gelidium Latifolium Ethanol Extracts
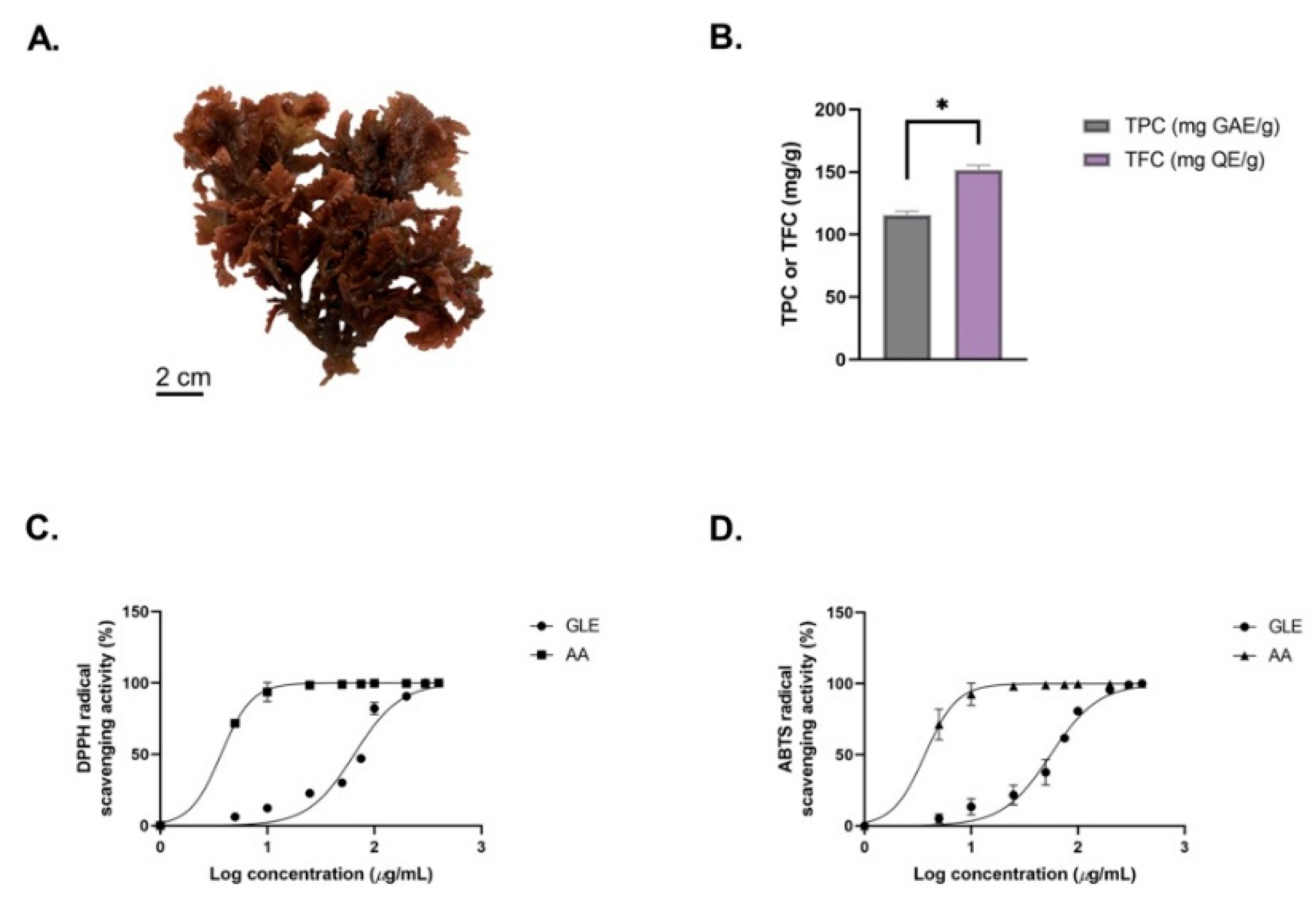
2.4. Determination of GLE’s Total Phenolic Content and Total Flavonoid Content
2.5. Antioxidant Activity of GLE (DPPH and ABTS Assays)
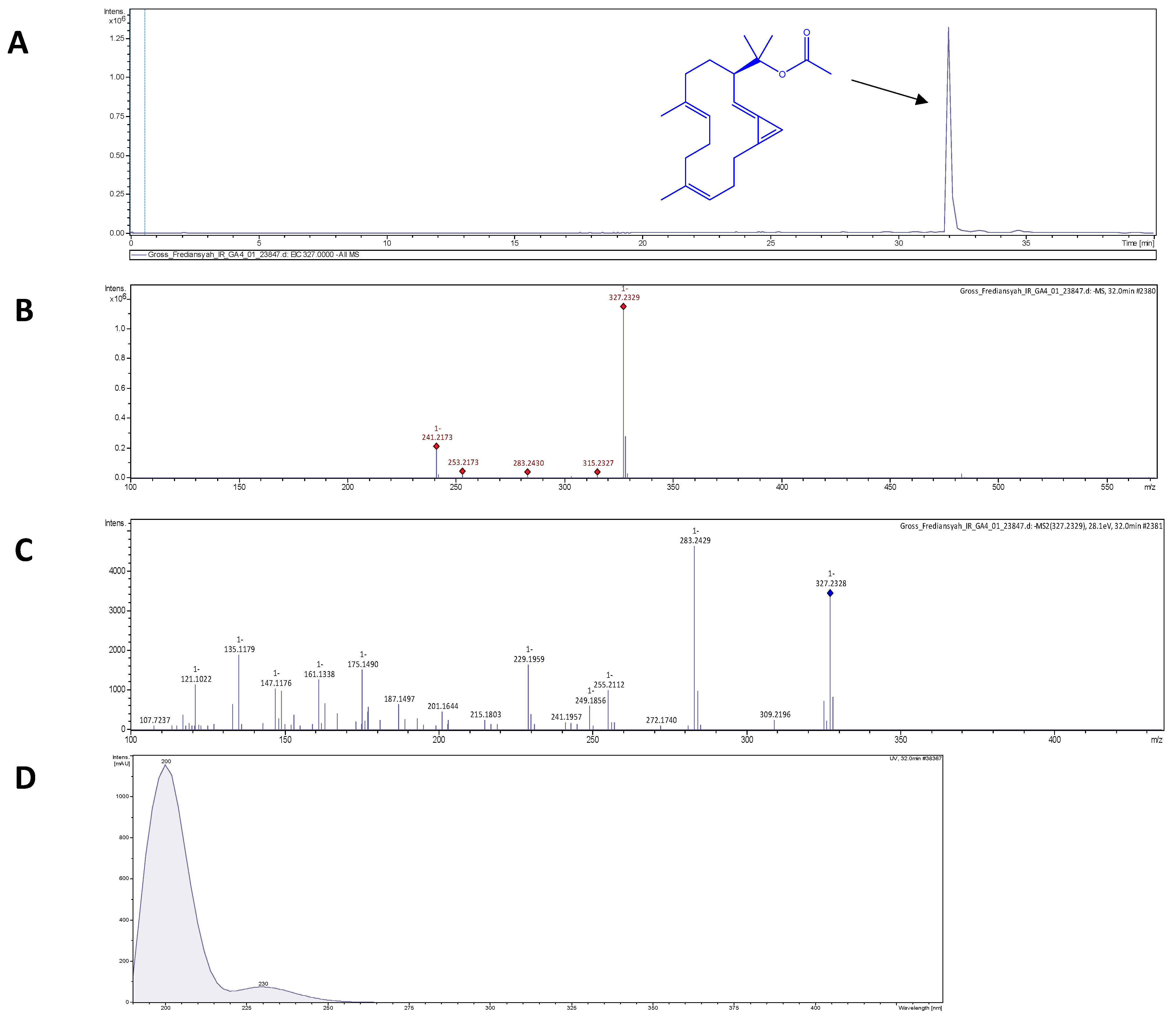
2.6. Electrospray Ionization High-Resolution Time-of-Flight Mass Spectrometry (ESI-HR-TOF-MS)
2.7. B16-F10 Murine Melanoma Cell Culture
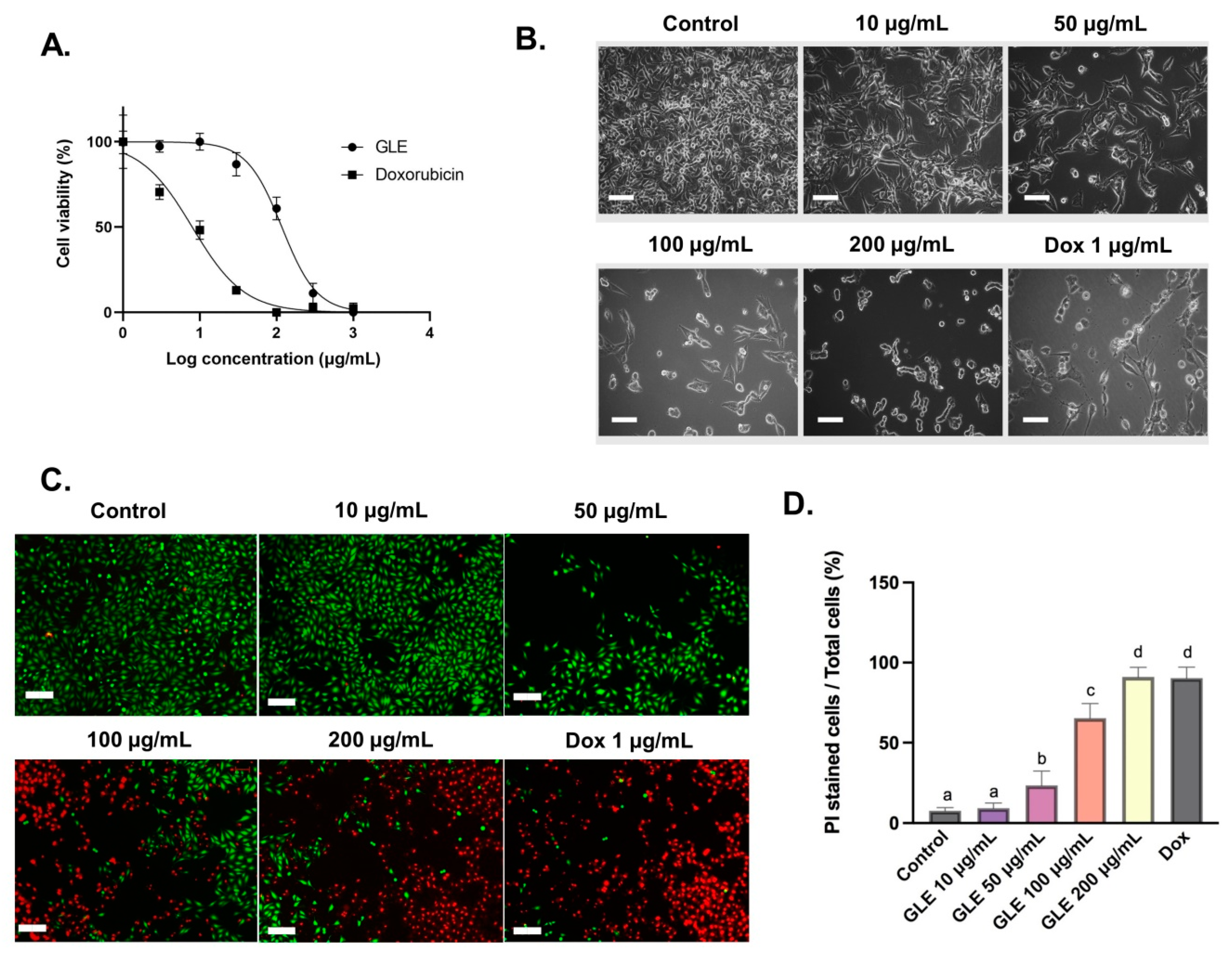
2.8. Cytotoxicity Assay
2.9. Calcein-AM and Propidium Iodide Staining (Viability Analyses)
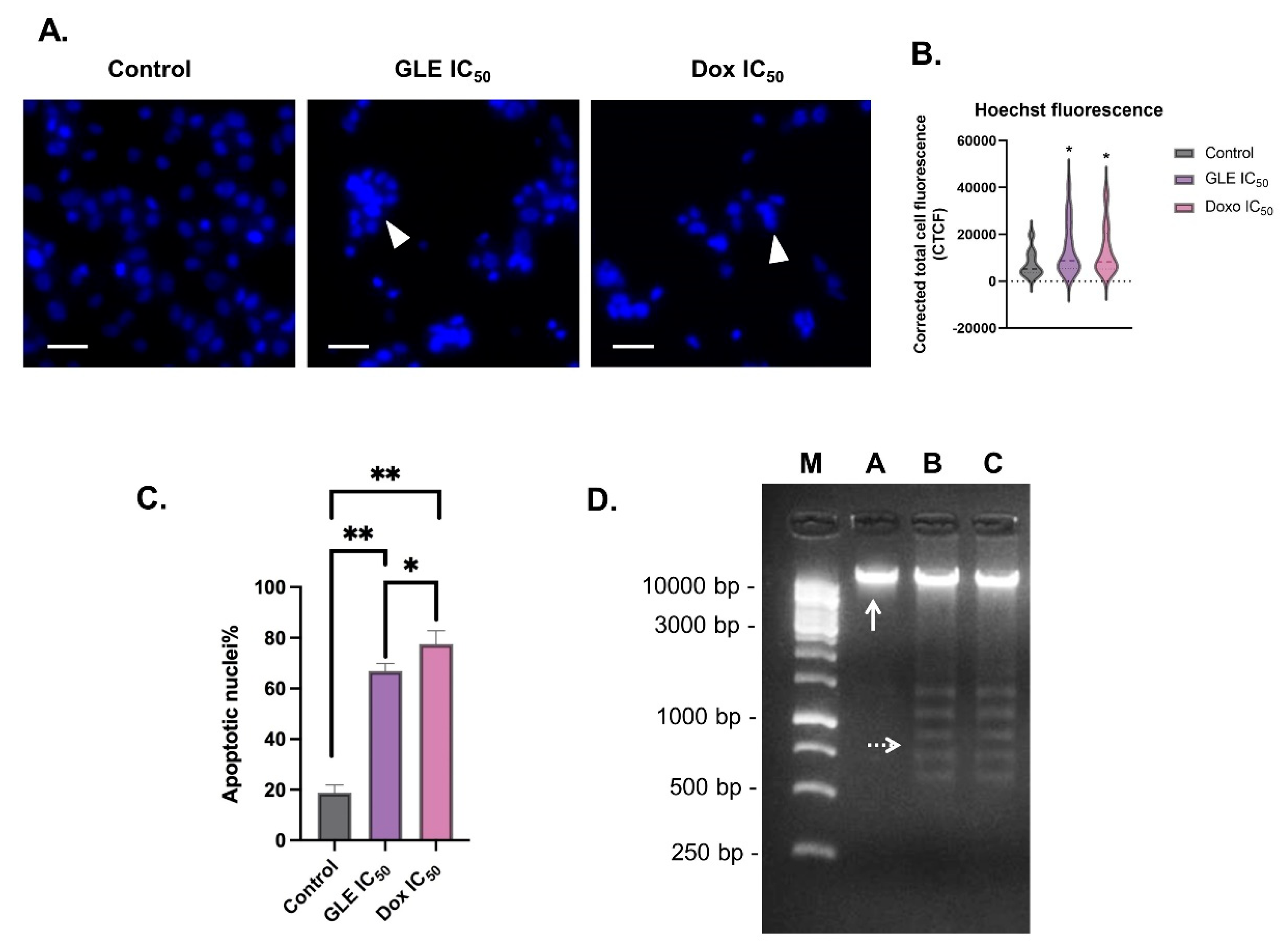
2.10. GLE’s Effect on DNA Condensation (Hoechst33342 Nuclear Staining)
2.11. GLE’s Effect on DNA Fragmentation
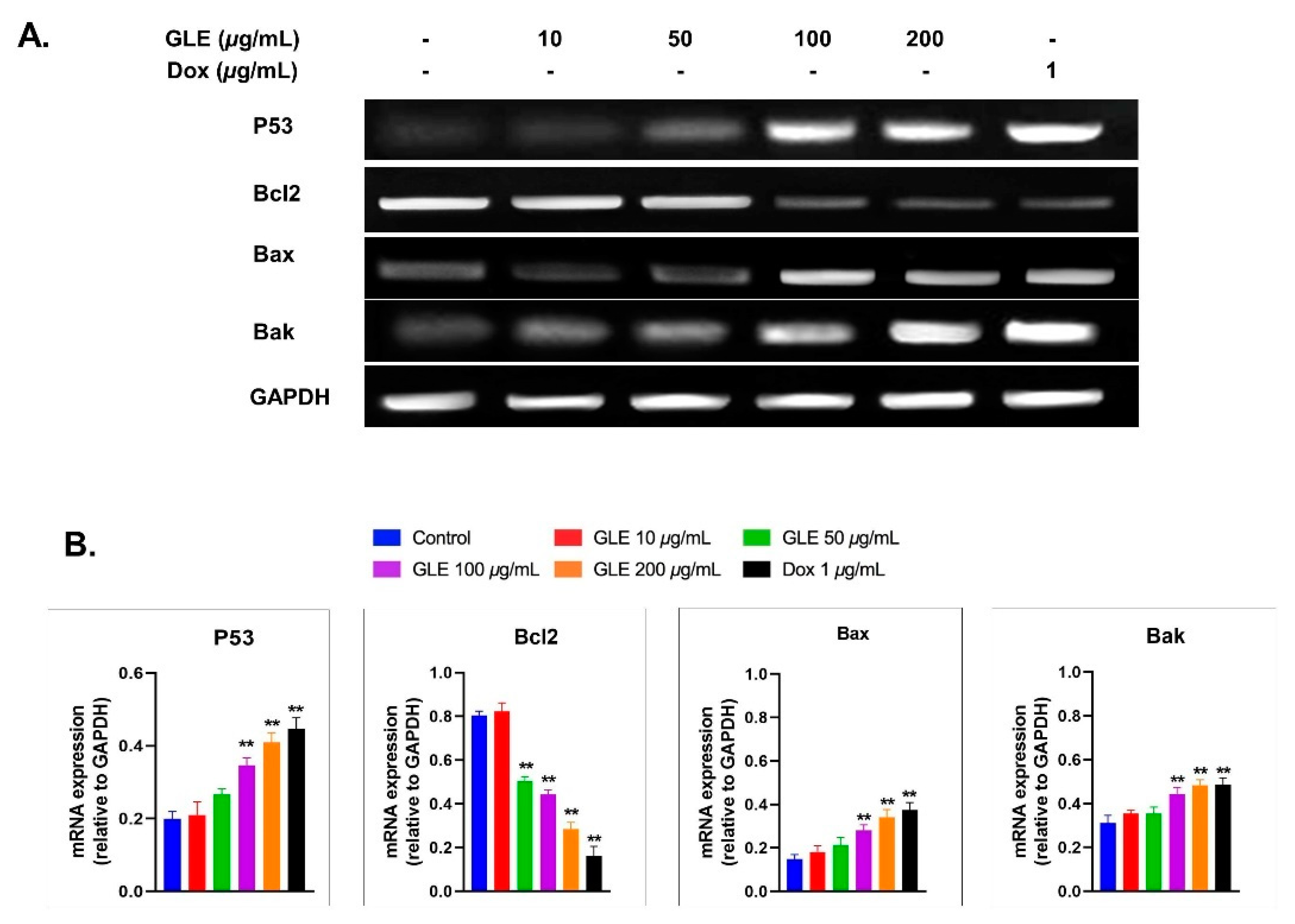
2.12. RNA Isolation and Semi-Quantitative PCR Analysis
2.13. Statistical Analyses
3. Results
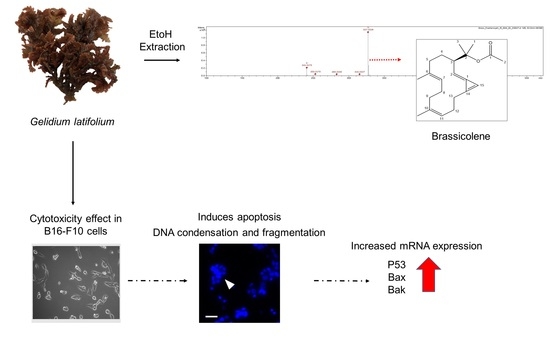
3.1. The Morphology and Phytochemical Analysis of GLE
3.2. Brassicolene Detected in GLE by ESI-HR-TOF-MS
3.3. Cytotoxic Activity of GLE on B16-F10 Melanoma Cells
3.4. Cell Viability Analyses with Fluorescence Double Staining (Calcein-AM/PI)
3.5. Effect of GLE on B16-F10 Apoptosis
3.6. Effects of GLE on Bak, Bax, and Bcl2 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dimitriou, F.; Krattinger, R.; Ramelyte, E.; Barysch, M.J.; Micaletto, S.; Dummer, R.; Goldinger, S.M. The World of Melanoma: Epidemiologic, Genetic, and Anatomic Differences of Melanoma Across the Globe. Curr. Oncol. Rep. 2018, 20, 87. [Google Scholar] [CrossRef] [PubMed]
- Savoye, I.; Olsen, C.M.; Whiteman, D.C.; Bijon, A.; Wald, L.; Dartois, L.; Clavel-Chapelon, F.; Boutron-Ruault, M.-C.; Kvaskoff, M. Patterns of Ultraviolet Radiation Exposure and Skin Cancer Risk: The E3N-SunExp Study. J. Epidemiol. 2018, 28, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.; Holman, D.M.; Maguire-Eisen, M. Ultraviolet Radiation Exposure and Its Impact on Skin Cancer Risk. Semin. Oncol. Nurs. 2016, 32, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Ashraf, M.A. Phytochemicals as Potential Anticancer Drugs: Time to Ponder Nature’s Bounty. BioMed Res. Int. 2020, 2020, 8602879. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Identification of Bioactive Peptides with α-Amylase Inhibitory Potential from Enzymatic Protein Hydrolysates of Red Seaweed (Porphyra Spp). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef] [PubMed]
- Carson, M.A.; Clarke, S.A. Bioactive Compounds from Marine Organisms: Potential for Bone Growth and Healing. Mar. Drugs 2018, 16, 340. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine Macroalgae as Sources of Protein and Bioactive Compounds in Feed for Monogastric Animals. J. Sci. Food Agric. 2018, 99, 13–24. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Miyake, M.; Kobayashi, D.; Hazama, A. Carrageenan Delays Cell Cycle Progression in Human Cancer Cells in Vitro Demonstrated by FUCCI Imaging. BMC Complement. Altern. Med. 2016, 16, 270. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Syafitri, S.M.; Geraldine, B.A.F.D.; Hamdin, C.D.; Frediansyah, A.; Miyake, M.; Kobayashi, D.; Hazama, A.; Sunarpi, H. UVA Photoprotective Activity of Brown Macroalgae Sargassum Cristafolium. Biomedicines 2019, 7, 77. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Martyasari, N.W.R.; Abidin, A.S.; Pebriani, S.A.; Ilhami, B.T.K.; Frediansyah, A.; Sunarwidhi, A.L.; Widyastuti, S.; Sunarpi, H. Macroalgae Sargassum Cristaefolium Extract Inhibits Proinflammatory Cytokine Expression in BALB/C Mice. Scientifica 2020, 2020, 9769454. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef] [PubMed]
- Ruan, B.-F.; Ge, W.-W.; Lin, M.-X.; Li, Q.-S. A Review of the Components of Seaweeds as Potential Candidates in Cancer Therapy. Anticancer Agents Med. Chem. 2018, 18, 354–366. [Google Scholar] [CrossRef]
- Ghannam, A.; Murad, H.; Jazzara, M.; Odeh, A.; Allaf, A.W. Isolation, Structural Characterization, and Antiproliferative Activity of Phycocolloids from the Red Seaweed Laurencia Papillosa on MCF-7 Human Breast Cancer Cells. Int. J. Biol. Macromol. 2018, 108, 916–926. [Google Scholar] [CrossRef]
- Desamero, M.J.; Kakuta, S.; Chambers, J.K.; Uchida, K.; Hachimura, S.; Takamoto, M.; Nakayama, J.; Nakayama, H.; Kyuwa, S. Orally Administered Brown Seaweed-Derived β-Glucan Effectively Restrained Development of Gastric Dysplasia in A4gnt KO Mice That Spontaneously Develop Gastric Adenocarcinoma. Int. Immunopharmacol. 2018, 60, 211–220. [Google Scholar] [CrossRef]
- Rocha, D.H.A.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed Secondary Metabolites In Vitro and In Vivo Anticancer Activity. Mar. Drugs 2018, 16, 410. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-C.; Hou, M.-F.; Huang, H.-W.; Chang, F.-R.; Yeh, C.-C.; Tang, J.-Y.; Chang, H.-W. Marine Algal Natural Products with Anti-Oxidative, Anti-Inflammatory, and Anti-Cancer Properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef]
- Tannoury, M.; Saab, A.; Elia, J.; Harb, N.; Makhlouf, H.; Diab, M. In Vitro Cytotoxic Activity of Laurencia Papillosa, Marine Red Algae from the Lebanese Coast. J. Appl. Pharm. Sci. 2017, 7, 175–179. [Google Scholar]
- Pereira, L. Macroalgae. Encyclopedia 2021, 1, 177–188. [Google Scholar] [CrossRef]
- Khotimchenko, M.; Tiasto, V.; Kalitnik, A.; Begun, M.; Khotimchenko, R.; Leonteva, E.; Bryukhovetskiy, I.; Khotimchenko, Y. Antitumor Potential of Carrageenans from Marine Red Algae. Carbohydr. Polym. 2020, 246, 116568. [Google Scholar] [CrossRef]
- Aziz, E.; Batool, R.; Khan, M.U.; Rauf, A.; Akhtar, W.; Heydari, M.; Rehman, S.; Shahzad, T.; Malik, A.; Mosavat, S.H.; et al. An overview on red algae bioactive compounds and their pharmaceutical applications. J. Complement. Integr. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Yanshin, N.; Kushnareva, A.; Lemesheva, V.; Birkemeyer, C.; Tarakhovskaya, E. Chemical Composition and Potential Practical Application of 15 Red Algal Species from the White Sea Coast (the Arctic Ocean). Molecules 2021, 26, 2489. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Marcone, M.; Baptista, J. Health-Promoting Ingredients from Four Selected Azorean Macroalgae. Food Res. Int. 2016, 89, 432–438. [Google Scholar] [CrossRef]
- Yang, T.-H.; Yao, H.-T.; Chiang, M.-T. Red Algae (Gelidium amansii) Hot-Water Extract Ameliorates Lipid Metabolism in Hamsters Fed a High-Fat Diet. J. Food Drug Anal. 2017, 25, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Pugazhendhi, A.; Prabakar, D.; Jacob, J.M.; Karuppusamy, I.; Saratale, R.G. Synthesis and Characterization of Silver Nanoparticles Using Gelidium amansii and Its Antimicrobial Property against Various Pathogenic Bacteria. Microb. Pathog. 2018, 114, 41–45. [Google Scholar] [CrossRef]
- Choi, J.; Kim, K.-J.; Koh, E.-J.; Lee, B.-Y. Gelidium Elegans Extract Ameliorates Type 2 Diabetes via Regulation of MAPK and PI3K/Akt Signaling. Nutrients 2018, 10, 51. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H.-J.; Lee, M. The Suppressive Effect of Gelidium Amansi-EtOH Extracts on the Adipogenesis with MAPK Signals in Adipocytes with or without Macrophages. Food Sci. Biotechnol. 2017, 26, 1715–1723. [Google Scholar] [CrossRef] [PubMed]
- Algaebase: Listing the World’s Algae. Available online: https://www.algaebase.org/ (accessed on 30 August 2021).
- Valentina, F.; Pinheiro, C.M. Physicochemical Changes of Air-Dried and Salt-Processed Ulva rigida over Storage Time. Molecules 2019, 24, 2955. [Google Scholar] [CrossRef]
- Hideki Kanda, W. Direct Extraction of Lutein from Wet Macroalgae by Liquefied Dimethyl Ether without Any Pretreatment. ACS Omega 2020, 5, 24005. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.; Schellenberg, I. Comparison of Different Extraction Methods for the Determination of Essential Oils and Related Compounds from Aromatic Plants and Optimization of Solid-Phase Microextraction/Gas Chromatography. Anal. Bioanal. Chem. 2007, 387, 2207–2217. [Google Scholar] [CrossRef]
- Boonchumi, W.; Peerapornpisal, Y.; Kanjanapothi, D.; Pekkoh, J.; Pumas, C.; Jamjai, U.; Amornlerdpison, D.; Noiraksar, T.; Vacharapiyasophon, P. Antioxidant Activity of Some Seaweed from the Gulf of Thailand. Int. J. Agric. Biol. 2011, 13, 1560–8530. [Google Scholar]
- Neto, R.T.; Marçal, C.; Queirós, A.S.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Screening of Ulva Rigida, Gracilaria Sp., Fucus Vesiculosus and Saccharina Latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Viedma, J.; Aguilera, J.M.; Flores, M.; Lemus-Mondaca, R.; Larrazabal, M.J.; Miranda, J.M.; Aubourg, S.P. Protective Effect of Red Algae (Rhodophyta) Extracts on Essential Dietary Components of Heat-Treated Salmon. Antioxidants 2021, 10, 1108. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave Assisted Extraction of Phenolic Compounds from Four Economic Brown Macroalgae Species and Evaluation of Their Antioxidant Activities and Inhibitory Effects on α-Amylase, α-Glucosidase, Pancreatic Lipase and Tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.-J.; Xu, M.-X.; Zhou, B.-H.; Du, Y.-S.; Yao, J.-H.; Zhou, L. Effects of 2-Aryl-1-Cyano-1,2,3,4-Tetrohydroisoquinolines on Apoptosis Induction Mechanism in NB4 and MKN-45 Cells. Toxicol. Vitro Int. 2018, 54, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Bratosin, D.; Mitrofan, L.; Palii, C.; Estaquier, J.; Montreuil, J. Novel fluorescence assay using calcein-AM for the determination of human erythrocyte viability and aging. Cytom. Part A J. Int. Soc. Anal. Cytol. 2005, 66, 78–84. [Google Scholar] [CrossRef]
- Arora, S.; Tandon, S. DNA Fragmentation and Cell Cycle Arrest: A Hallmark of Apoptosis Induced by Ruta Graveolens in Human Colon Cancer Cells. Homeopathy 2015, 104, 36–47. [Google Scholar] [CrossRef]
- Jakic, B.; Buszko, M.; Cappellano, G.; Wick, G. Elevated Sodium Leads to the Increased Expression of HSP60 and Induces Apoptosis in HUVECs. PLoS ONE 2017, 12, e0179383. [Google Scholar] [CrossRef]
- Kasibhatla, S.; Amarante-Mendes, G.P.; Finucane, D.; Brunner, T.; Bossy-Wetzel, E.; Green, D.R. Analysis of DNA Fragmentation Using Agarose Gel Electrophoresis. CSH Protoc. 2006, 2006, pdb.prot4429. [Google Scholar] [CrossRef]
- Zakaria, F.R.; Prangdimurti, E.; Adawiyah, D.R.; Priosoeryanto, B.P.; Huda, N. Chemical Evaluation of a Nori-like Product (Geluring) Made from the Mixture of Gelidium Sp. and Ulva Lactuca Seaweeds. Curr. Res. Nutr. Food Sci. J. 2018, 6, 664–671. [Google Scholar]
- Generalić Mekinić, I.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Popović Perković, Z. Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef]
- Alvarez-Gomez, F. Analysis of Antioxidant Capacity and Bioactive Compounds in Marine Macroalgal and Lichenic Extracts Using Different Solvents and Evaluation Methods. Cienc. Mar. 2016, 42, 271–288. [Google Scholar] [CrossRef]
- Pirian, K.; Jeliani, Z.Z.; Sohrabipour, J.; Arman, M.; Faghihi, M.M.; Yousefzadi, M. Nutritional and Bioactivity Evaluation of Common Seaweed Species from the Persian Gulf. Iran. J. Sci. Technol. Trans. Sci. 2018, 42, 1795–1804. [Google Scholar] [CrossRef]
- Agarwal, P.; Kayala, P.; Chandrasekaran, N.; Mukherjee, A.; Shah, S.; Thomas, J. Antioxidant and Antibacterial Activity of Gelidium Pusillum (Stackhouse) against Aeromonas Caviae and Its Applications in Aquaculture. Aquac. Int. 2021, 29, 845–858. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D.; Praveen, N.K. Antioxidant Activities and Phenolic Contents of Three Red Seaweeds (Division: Rhodophyta) Harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Technol. 2015, 52, 1924–1935. [Google Scholar] [CrossRef]
- Vega, J.; Álvarez-Gómez, F.; Güenaga, L.; Figueroa, F.L.; Gómez-Pinchetti, J.L. Antioxidant Activity of Extracts from Marine Macroalgae, Wild-Collected and Cultivated, in an Integrated Multi-Trophic Aquaculture System. Aquaculture 2020, 522, 735088. [Google Scholar] [CrossRef]
- Molyneux, P. The Use of the Stable Free Radical Diphenylpicrylhydrazyl (DPPH) for Estimating Antioxidant. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Duh, C.-Y.; Wang, S.-K.; Weng, Y.-L. Brassicolene, A Novel Cytotoxic Diterpenoid from the Formosan Soft Coral Nephthea Brassica. Tetrahedron Lett. 2000, 41, 1401–1403. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Ta, Q.T.H.; Pham, Q.T.; Luong, T.N.H.; Phung, V.T.; Duong, T.-H.; Vo, V.G. Anticancer Activity of Novel Plant Extracts and Compounds from Adenosma Bracteosum (Bonati) in Human Lung and Liver Cancer Cells. Molecules 2020, 25, 2912. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Tu, C.-J.; Wu, H.-T. Growth-Inhibitory Effects of the Red Alga Gelidium amansii on Cultured Cells. Biol. Pharm. Bull. 2004, 27, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Noro, T.; Kamei, Y. Selective antitumor activity in vitro from marine algae from Japan coasts. Biol. Pharm. Bull. 1997, 20, 541–546. [Google Scholar] [CrossRef]
- Martins, R.M.; Nedel, F.; Guimarães, V.B.S.; da Silva, A.F.; Colepicolo, P.; de Pereira, C.M.P.; Lund, R.G. Macroalgae Extracts from Antarctica Have Antimicrobial and Anticancer Potential. Front. Microbiol. 2018, 9, 412. [Google Scholar] [CrossRef]
- Shalaby, E. Algae as Promising Organisms for Environment and Health. Plant Signal. Behav. 2011, 6, 1338–1350. [Google Scholar] [CrossRef] [PubMed]
- Shiao, W.-C.; Kuo, C.-H.; Tsai, Y.-H.; Hsieh, S.-L.; Kuan, A.-W.; Hong, Y.-H.; Huang, C.-Y. In Vitro Evaluation of Anti-Colon Cancer Potential of Crude Extracts of Fucoidan Obtained from Sargassum Glaucescens Pretreated by Compressional-Puffing. Appl. Sci. 2020, 10, 3058. [Google Scholar] [CrossRef]
- Yuan, J.; Yankner, B.A. Apoptosis in the Nervous System. Nature 2000, 407, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Pelicci, S.; Diaspro, A.; Lanzanò, L. Chromatin Nanoscale Compaction in Live Cells Visualized by Acceptor-to-Donor Ratio Corrected Förster Resonance Energy Transfer between DNA Dyes. J. Biophotonics 2019, 12, e201900164. [Google Scholar] [CrossRef]
- Walker, P.R.; LeBlanc, J.; Sikorska, M. Evidence That DNA Fragmentation in Apoptosis Is Initiated and Propagated by Single-Strand Breaks. Cell Death Differ. 1997, 4, 506–515. [Google Scholar] [CrossRef]
- Bernardini, G.; Minetti, M.; Polizzotto, G.; Biazzo, M.; Santucci, A. Pro-Apoptotic Activity of French Polynesian Padina Pavonica Extract on Human Osteosarcoma Cells. Mar. Drugs 2018, 16, 504. [Google Scholar] [CrossRef]
- Güner, A.; Nalbantsoy, A.; Sukatar, A.; Karabay Yavaşoğlu, N.Ü. Apoptosis-Inducing Activities of Halopteris Scoparia L. Sauvageau (Brown Algae) on Cancer Cells and Its Biosafety and Antioxidant Properties. Cytotechnology 2019, 71, 687–704. [Google Scholar] [CrossRef]
- Al Monla, R.; Dassouki, Z.; Kouzayha, A.; Salma, Y.; Gali-Muhtasib, H.; Mawlawi, H. The Cytotoxic and Apoptotic Effects of the Brown Algae Colpomenia Sinuosa Are Mediated by the Generation of Reactive Oxygen Species. Molecules 2020, 25, 1993. [Google Scholar] [CrossRef]
- Çelenk, F.G.; Özkaya, A.B.; Sukatar, A. Macroalgae of Izmir Gulf: Dictyotaceae Exhibit High in Vitro Anti-Cancer Activity Independent from Their Antioxidant Capabilities. Cytotechnology 2016, 68, 2667–2676. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef]
- Haq, S.H.; Al-Ruwaished, G.; Al-Mutlaq, M.A.; Naji, S.A.; Al-Mogren, M.; Al-Rashed, S.; Ain, Q.T.; Al-Amro, A.A.; Al-Mussallam, A. Antioxidant, Anticancer Activity and Phytochemical Analysis of Green Algae, Chaetomorpha Collected from the Arabian Gulf. Sci. Rep. 2019, 9, 18906. [Google Scholar] [CrossRef]
- Ismail, M.M.; Alotaibi, B.S.; EL-Sheekh, M.M. Therapeutic Uses of Red Macroalgae. Molecules 2020, 25, 4411. [Google Scholar] [CrossRef]
- Alghazeer, R.; Howell, N.K.; El-Naili, M.B.; Awayn, N. Anticancer and Antioxidant Activities of Some Algae from Western Libyan Coast. Nat. Sci. 2018, 10, 232. [Google Scholar] [CrossRef][Green Version]
- Abdala Díaz, R.T.; Casas Arrojo, V.; Arrojo Agudo, M.A.; Cárdenas, C.; Dobretsov, S.; Figueroa, F.L. Immunomodulatory and Antioxidant Activities of Sulfated Polysaccharides from Laminaria Ochroleuca, Porphyra Umbilicalis, and Gelidium Corneum. Mar. Biotechnol. 2019, 21, 577–587. [Google Scholar] [CrossRef]
- Sithranga Boopathy, N.; Kathiresan, K. Anticancer Drugs from Marine Flora: An Overview. J. Oncol. 2010, 2010, 214186. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.-P. Chapter 9—Phycocolloids from macroalgae. In Natural Bioactive Compounds; Sinha, R.P., Häder, D.-P., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 187–201. ISBN 978-0-12-820655-3. [Google Scholar]
- Kim, M.S.; Kim, J.Y.; Choi, W.H.; Lee, S.S. Effects of Seaweed Supplementation on Blood Glucose Concentration, Lipid Profile, and Antioxidant Enzyme Activities in Patients with Type 2 Diabetes Mellitus. Nutr. Res. Pract. 2008, 2, 62–67. [Google Scholar] [CrossRef]
- Kang, M.-C.; Kang, N.; Kim, S.-Y.; Lima, I.S.; Ko, S.-C.; Kim, Y.-T.; Kim, Y.-B.; Jeung, H.-D.; Choi, K.-S.; Jeon, Y.-J. Popular Edible Seaweed, Gelidium amansii Prevents against Diet-Induced Obesity. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2016, 90, 181–187. [Google Scholar] [CrossRef]
- Nursid, M.; Khatulistiani, T.S.; Noviendri, D.; Hapsari, F.; Hardiyati, T. Total Phenolic Content, Antioxidant Activity and Tyrosinase Inhibitor from Marine Red Algae Extract Collected from Kupang, East Nusa Tenggara. IOP Conf. Ser. Earth Environ. Sci. 2020, 493, 012013. [Google Scholar] [CrossRef]
- Sammar, M.; Abu-Farich, B.; Rayan, I.; Falah, M.; Rayan, A. Correlation between Cytotoxicity in Cancer Cells and Free Radical-Scavenging Activity: In Vitro Evaluation of 57 Medicinal and Edible Plant Extracts. Oncol. Lett. 2019, 18, 6563–6571. [Google Scholar] [CrossRef] [PubMed]
- Hassan, L.E.A.; Ahamed, M.B.K.; Majid, A.S.A.; Baharetha, H.M.; Muslim, N.S.; Nassar, Z.D.; Majid, A.M.A. Correlation of Antiangiogenic, Antioxidant and Cytotoxic Activities of Some Sudanese Medicinal Plants with Phenolic and Flavonoid Contents. BMC Complement. Altern. Med. 2014, 14, 406. [Google Scholar] [CrossRef]
- ZiemLewska, A.; Zagórska-Dziok, M.; Nizioł-Łukaszewska, Z. Assessment of Cytotoxicity and Antioxidant Properties of Berry Leaves as By-Products with Potential Application in Cosmetic and Pharmaceutical Products. Sci. Rep. 2021, 11, 3240. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Gustafson, K.R. Marine Pharmacology in 2000: Antitumor and Cytotoxic Compounds. Int. J. Cancer 2003, 105, 291–299. [Google Scholar] [CrossRef] [PubMed]
- El Gamal, A.A. Biological Importance of Marine Algae. Saudi Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.; Murray, J.; Delgoda, R.; Gallimore, W. Two New Oxodolastane Diterpenes from the Jamaican Macroalga Canistrocarpus Cervicornis. Mar. Drugs 2017, 15, 150. [Google Scholar] [CrossRef]
- Altun, İ.; Sonkaya, A. The Most Common Side Effects Experienced by Patients Were Receiving First Cycle of Chemotherapy. Iran. J. Public Health 2018, 47, 1218–1219. [Google Scholar]
- Gambato, G.; Baroni, É.G.; Garcia, C.S.C.; Frassini, R.; Frozza, C.O.S.; Moura, S.; Pereira, C.M.P.; Fujii, M.T.; Colepicolo, P.; Lambert, A.P.F.; et al. Brown Algae Himantothallus Grandifolius (Desmarestiales, Phaeophyceae) Suppresses Proliferation and Promotes Apoptosis-Mediated Cell Death in Tumor Cells. Adv. Biol. Chem. 2014, 04, 98–108. [Google Scholar] [CrossRef]
- Canoy, J.L.; Bitacura, J.G. Cytotoxicity and Antiangiogenic Activity of Turbinaria Ornata Agardh and Padina Australis Hauck Ethanolic Extracts. Anal. Cell. Pathol. 2018, 2018, e3709491. [Google Scholar] [CrossRef]
- Hazekawa, M.; Nishinakagawa, T.; Kawakubo-Yasukochi, T.; Nakashima, M. Evaluation of IC50 Levels Immediately after Treatment with Anticancer Reagents Using a Real-time Cell Monitoring Device. Exp. Ther. Med. 2019, 18, 3197–3205. [Google Scholar] [CrossRef]
- Namvar, F.; Baharara, J.; Mahdi, A.A. Antioxidant and Anticancer Activities of Selected Persian Gulf Algae. Indian J. Clin. Biochem. 2014, 29, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Boise, L.H.; Shanmugam, M. Cancer Metabolism and the Evasion of Apoptotic Cell Death. Cancers 2019, 11, 1144. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Kalimuthu, K.; Park, Y.S.; Luo, X.; Choudry, M.H.A.; Bartlett, D.L.; Lee, Y.J. BAX-Dependent Mitochondrial Pathway Mediates the Crosstalk between Ferroptosis and Apoptosis. Apoptosis 2020, 25, 625–631. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasedya, E.S.; Ardiana, N.; Padmi, H.; Ilhami, B.T.K.; Martyasari, N.W.R.; Sunarwidhi, A.L.; Nikmatullah, A.; Widyastuti, S.; Sunarpi, H.; Frediansyah, A. The Antiproliferative and Apoptosis-Inducing Effects of the Red Macroalgae Gelidium latifolium Extract against Melanoma Cells. Molecules 2021, 26, 6568. https://doi.org/10.3390/molecules26216568
Prasedya ES, Ardiana N, Padmi H, Ilhami BTK, Martyasari NWR, Sunarwidhi AL, Nikmatullah A, Widyastuti S, Sunarpi H, Frediansyah A. The Antiproliferative and Apoptosis-Inducing Effects of the Red Macroalgae Gelidium latifolium Extract against Melanoma Cells. Molecules. 2021; 26(21):6568. https://doi.org/10.3390/molecules26216568
Chicago/Turabian StylePrasedya, Eka Sunarwidhi, Nur Ardiana, Hasriaton Padmi, Bq Tri Khairina Ilhami, Ni Wayan Riyani Martyasari, Anggit Listyacahyani Sunarwidhi, Aluh Nikmatullah, Sri Widyastuti, Haji Sunarpi, and Andri Frediansyah. 2021. "The Antiproliferative and Apoptosis-Inducing Effects of the Red Macroalgae Gelidium latifolium Extract against Melanoma Cells" Molecules 26, no. 21: 6568. https://doi.org/10.3390/molecules26216568
APA StylePrasedya, E. S., Ardiana, N., Padmi, H., Ilhami, B. T. K., Martyasari, N. W. R., Sunarwidhi, A. L., Nikmatullah, A., Widyastuti, S., Sunarpi, H., & Frediansyah, A. (2021). The Antiproliferative and Apoptosis-Inducing Effects of the Red Macroalgae Gelidium latifolium Extract against Melanoma Cells. Molecules, 26(21), 6568. https://doi.org/10.3390/molecules26216568