
Type-3 Hyaluronan Synthase Attenuates Tumor Cells Invasion in Human Mammary Parenchymal Tissues

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Primary Culture of HB-TDFs
2.3. Cell Proliferation Assay
2.4. HAS3 Gene RNA Interference Experiments
2.5. Immunohistochemistry (IHC) Analysis
2.6. Measurement of Serum HA Concentrations
2.7. Determination of the Molecular Mass of HA by Sandwich ELISA-like Assay in Conditioned Medium Harvested from MEF and MEF-Has3-KO Cells
2.8. In Vitro Cell Migration (Transwell) Assay
2.9. RNA Extraction and Quantitative Reverse Transcription PCR
2.10. Protein Extraction and Western Blotting
2.11. Immunofluorescence Staining and Confocal Microscopy
2.12. Transfection and Electroporation of Cells
2.13. Transmission Electron Microscopy (TEM)
2.14. Flow Cytometry Cell Cycle Analysis
2.15. Fluorescence Lifetime Imaging (FLIM) Microscopy
2.16. Generation of Has3-KO Mice
2.17. Preparation of Mouse Embryonic Fibroblasts (MEFs)
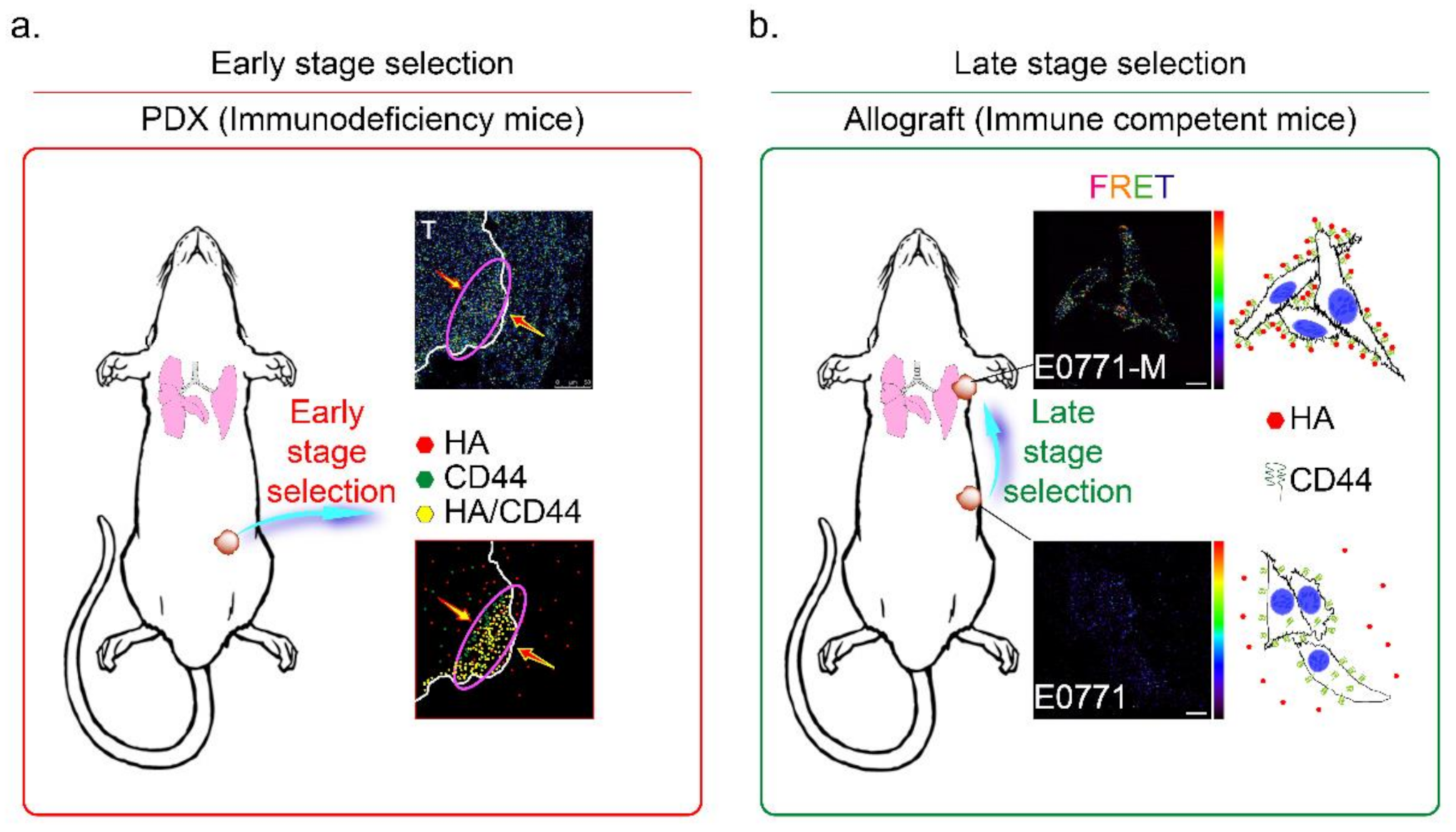
2.18. In Vivo Human Breast Cancer Xenograft Mouse Model
2.19. In Vivo Breast Cancer Allograft C57B6/J-Has3-KO Mouse Model
2.20. Ex Vivo Tumors: Isolation and Processing
2.21. In Vivo Breast Cancer Patient-Derived TNBC Tumor Xenograft Mouse Model
2.22. Fluorescent Immunohistochemistry (IHC) Staining
2.23. Statistical Analysis
3. Results
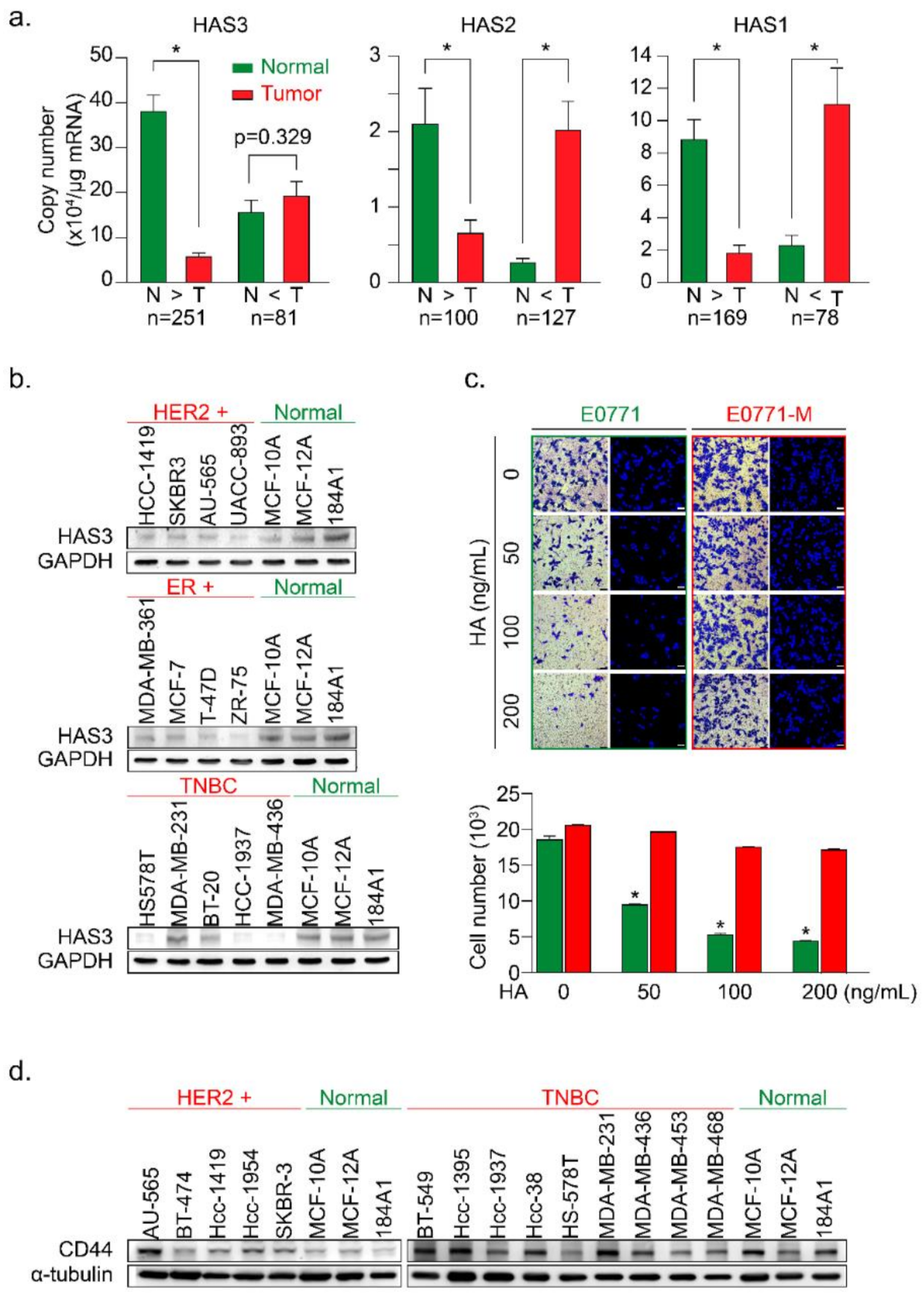
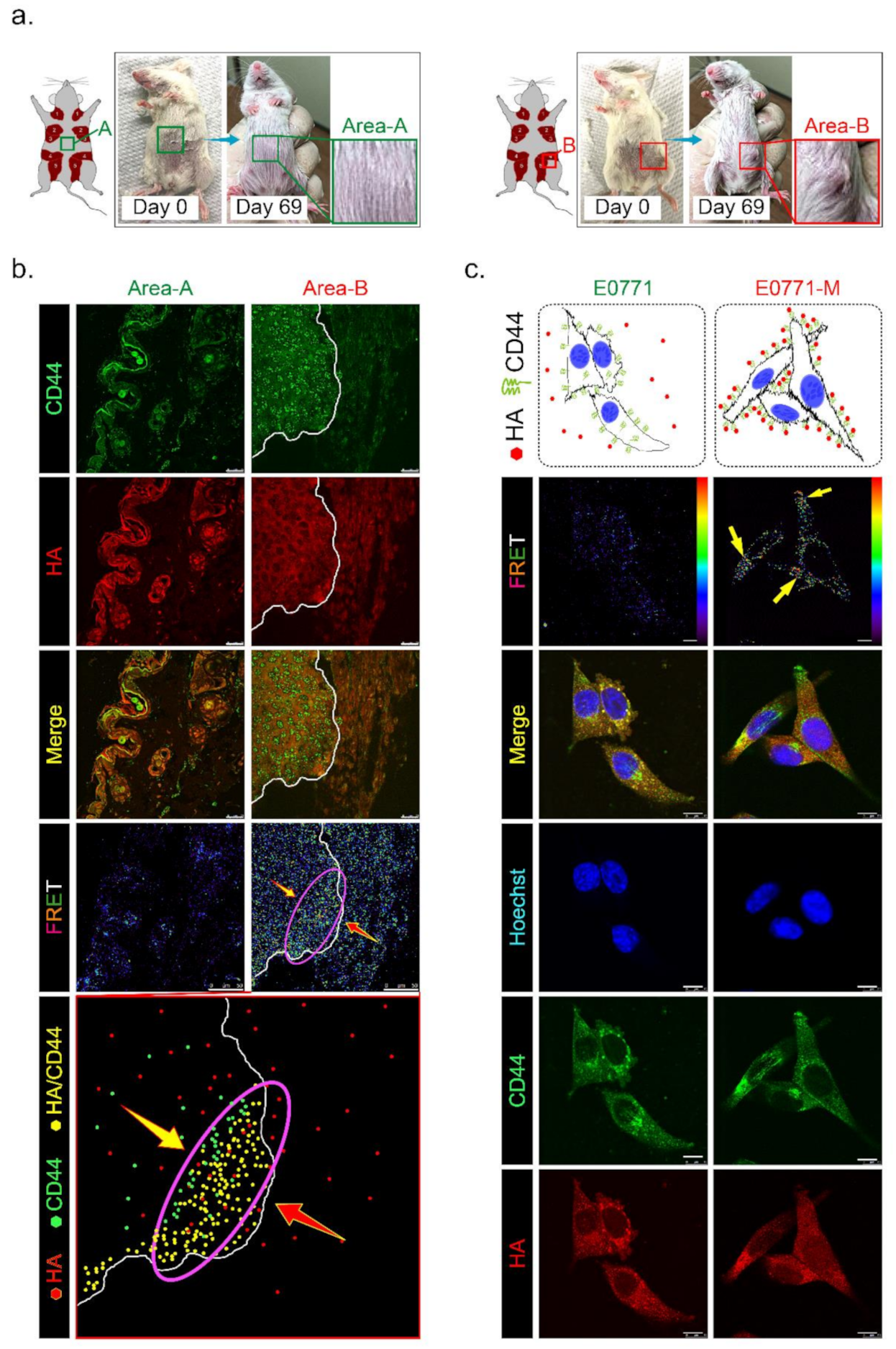
3.1. Higher HAS3 Protein Expression was Detected in the Stromal ECM of Breast Tumor Tissue
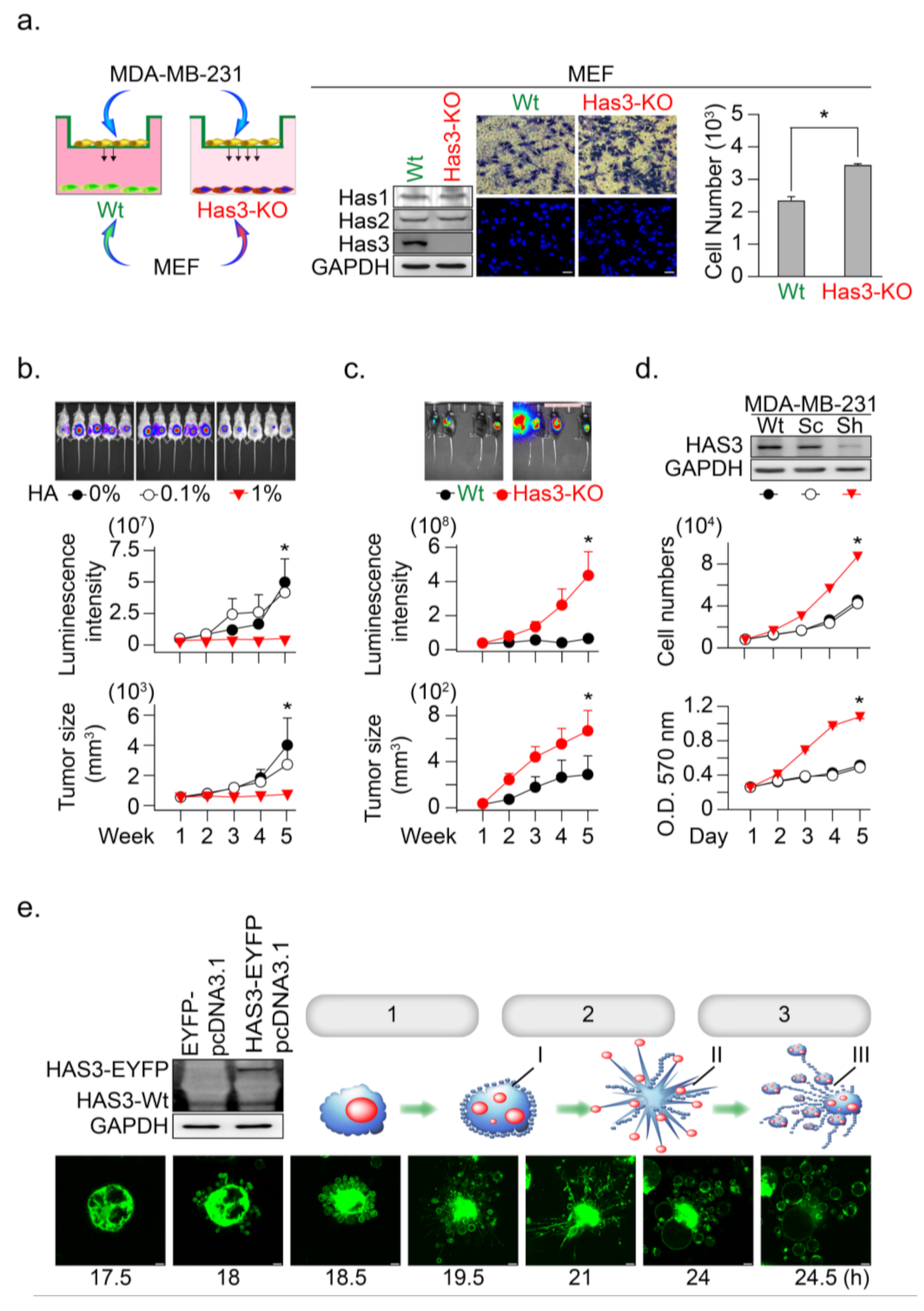
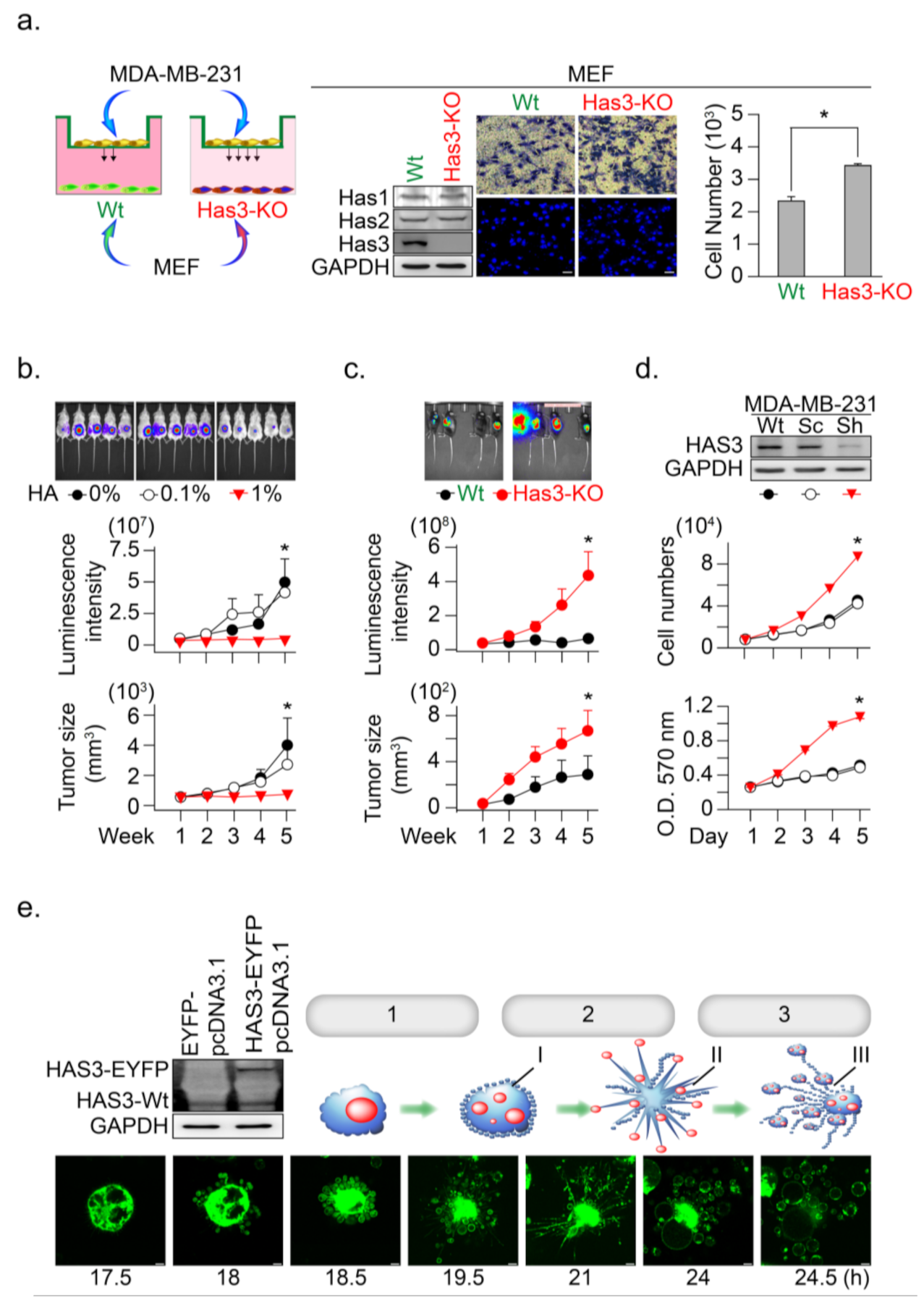
3.2. HAS3 Deficiency in Normal Stroma Tissue Promoted Breast Cancer Tumorigenesis
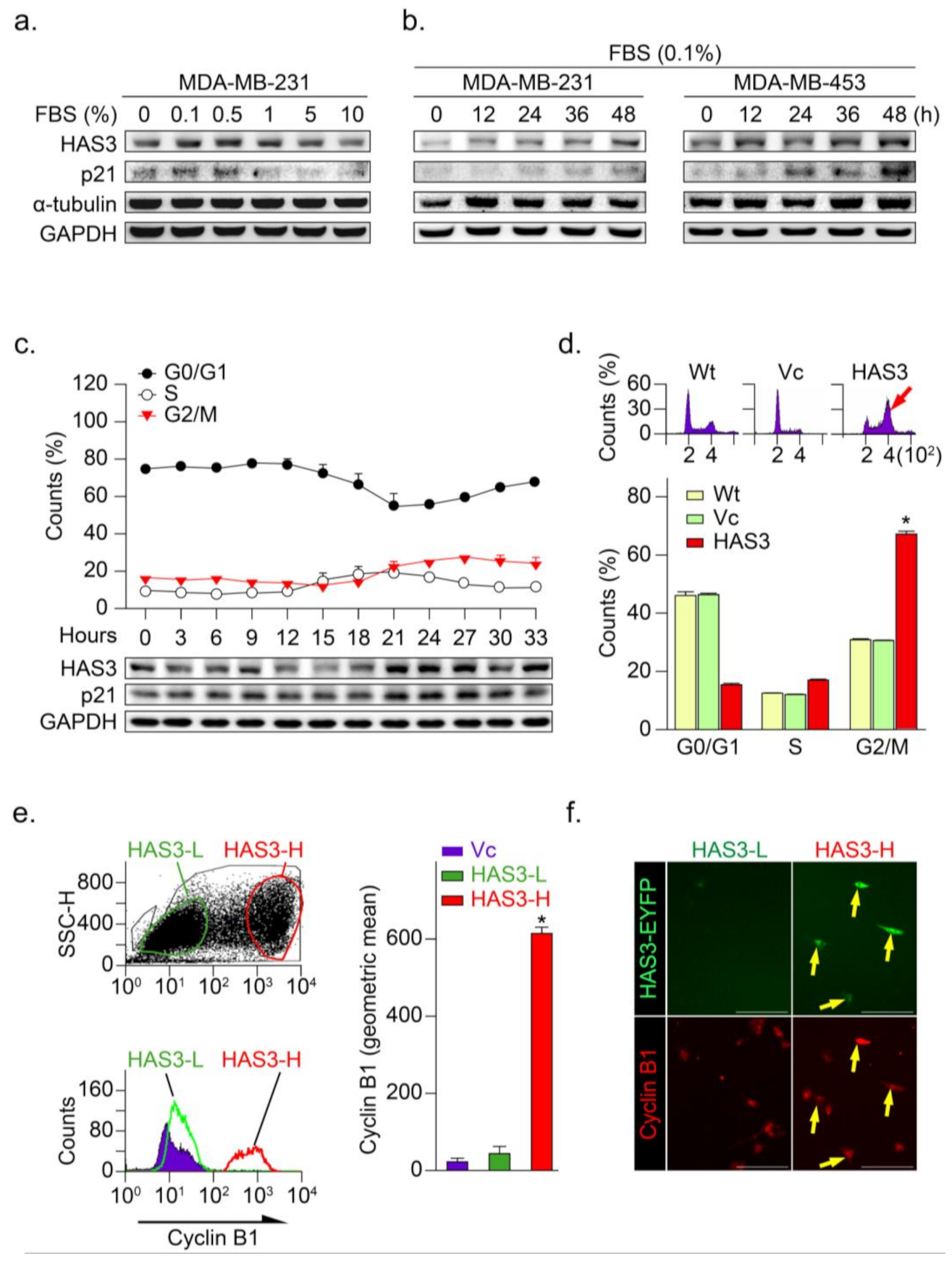
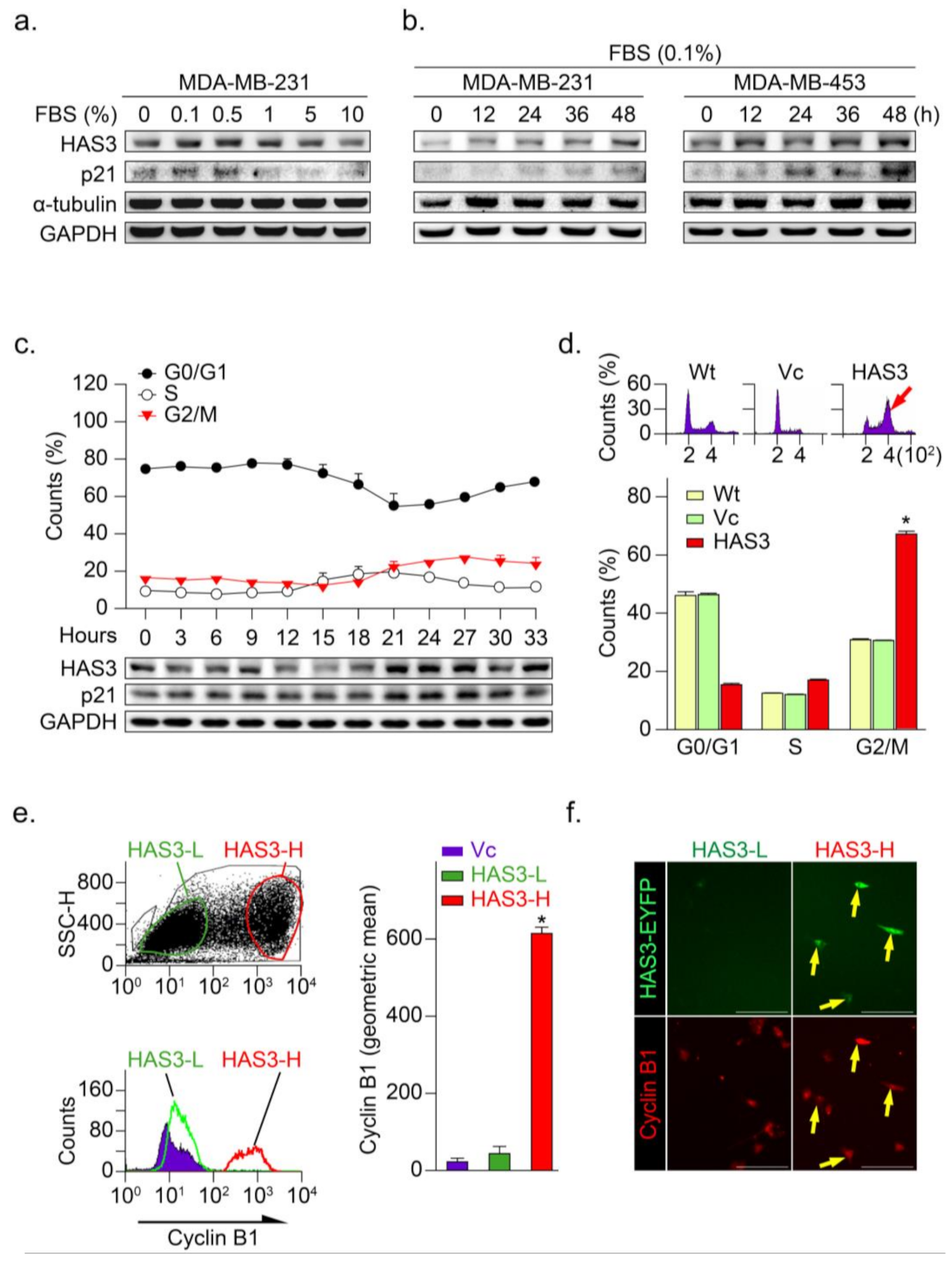
3.3. HAS3 Overexpression in Human Breast Cancer Cells Arrests the Cancer Cell Cycle at the G2/M Phase
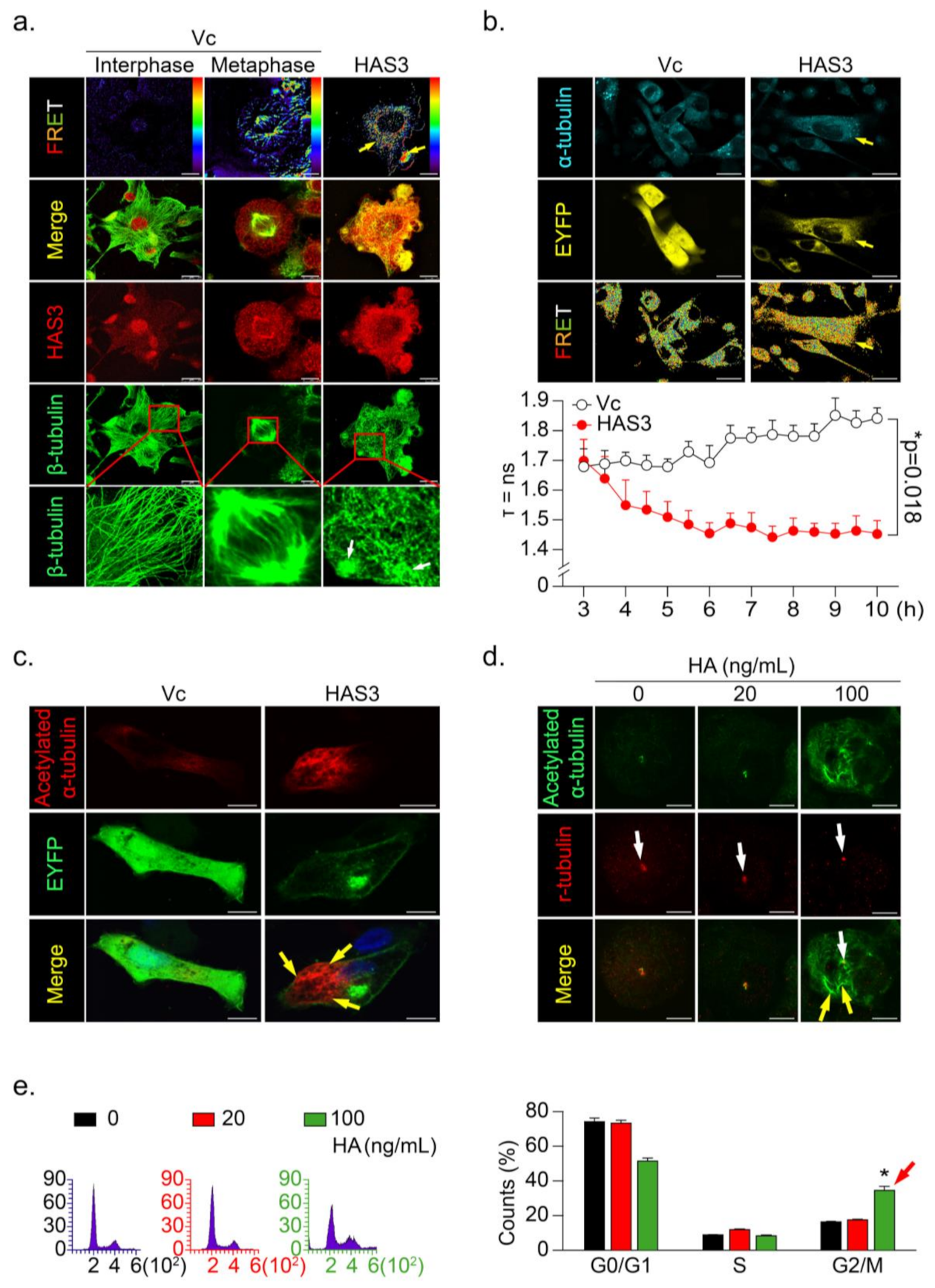
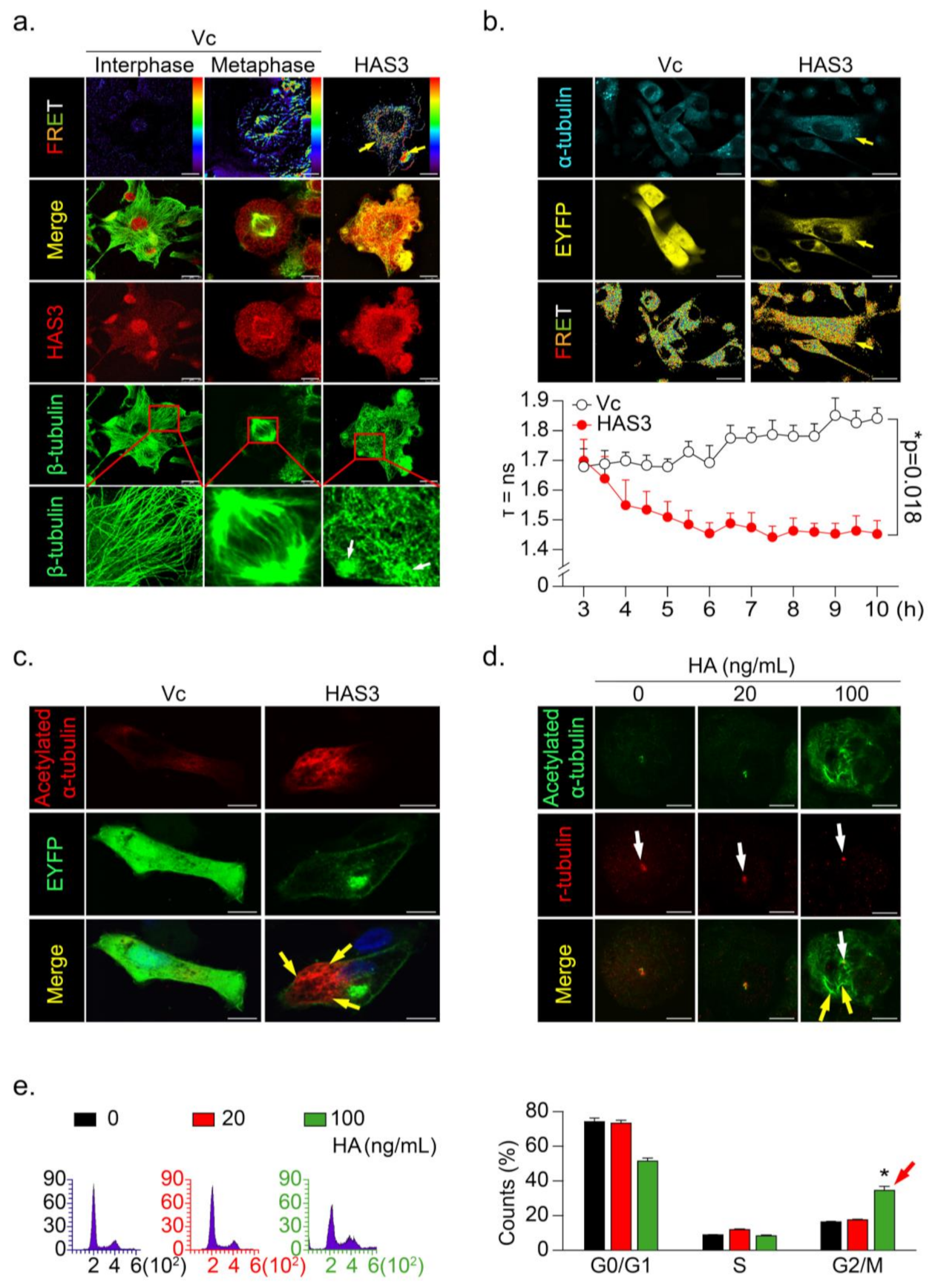
3.4. HAS3 Induces G2/M Phase Cell Cycle Arrest in Cancer Cells through Activation of Microtubule Hyperacetylation
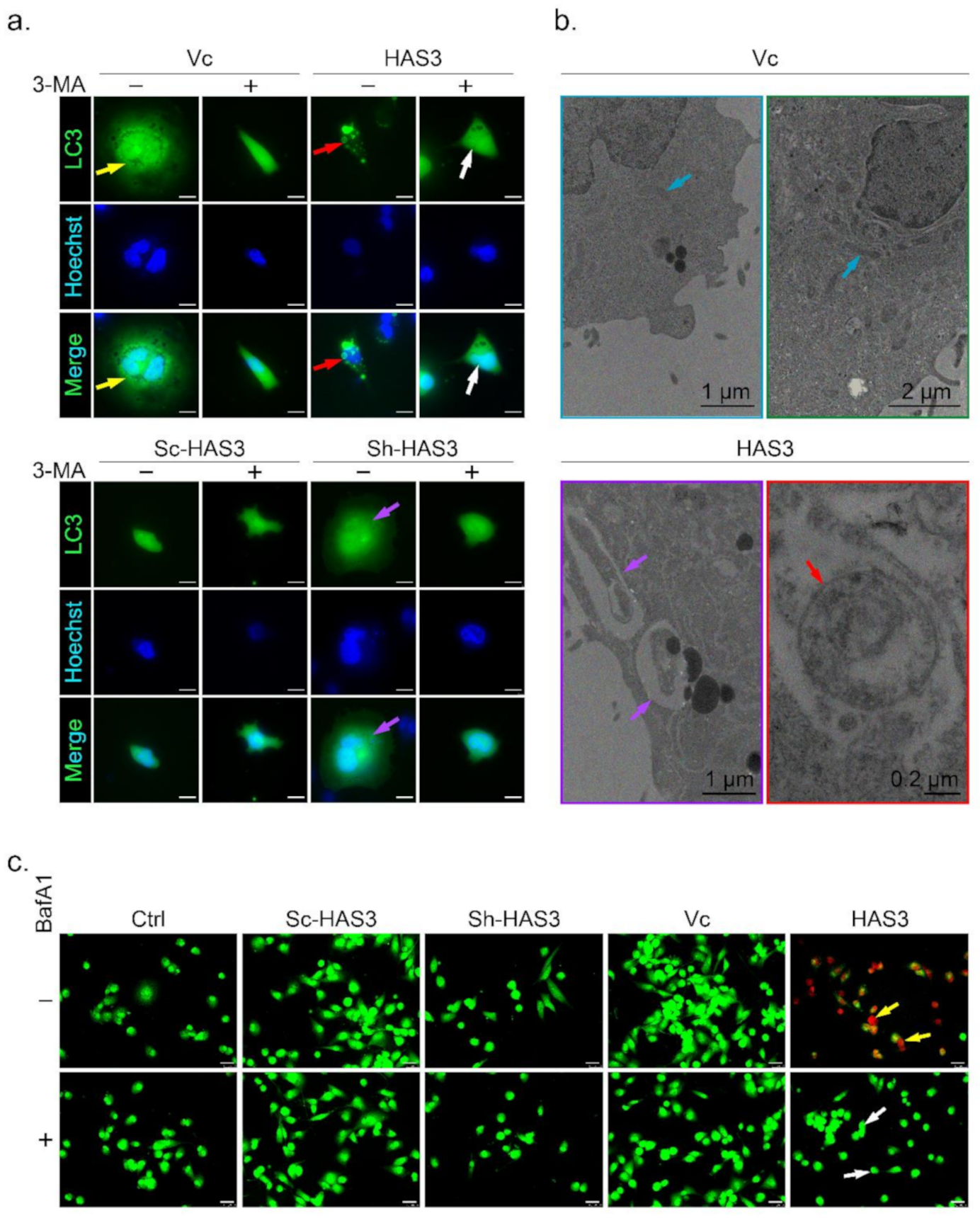
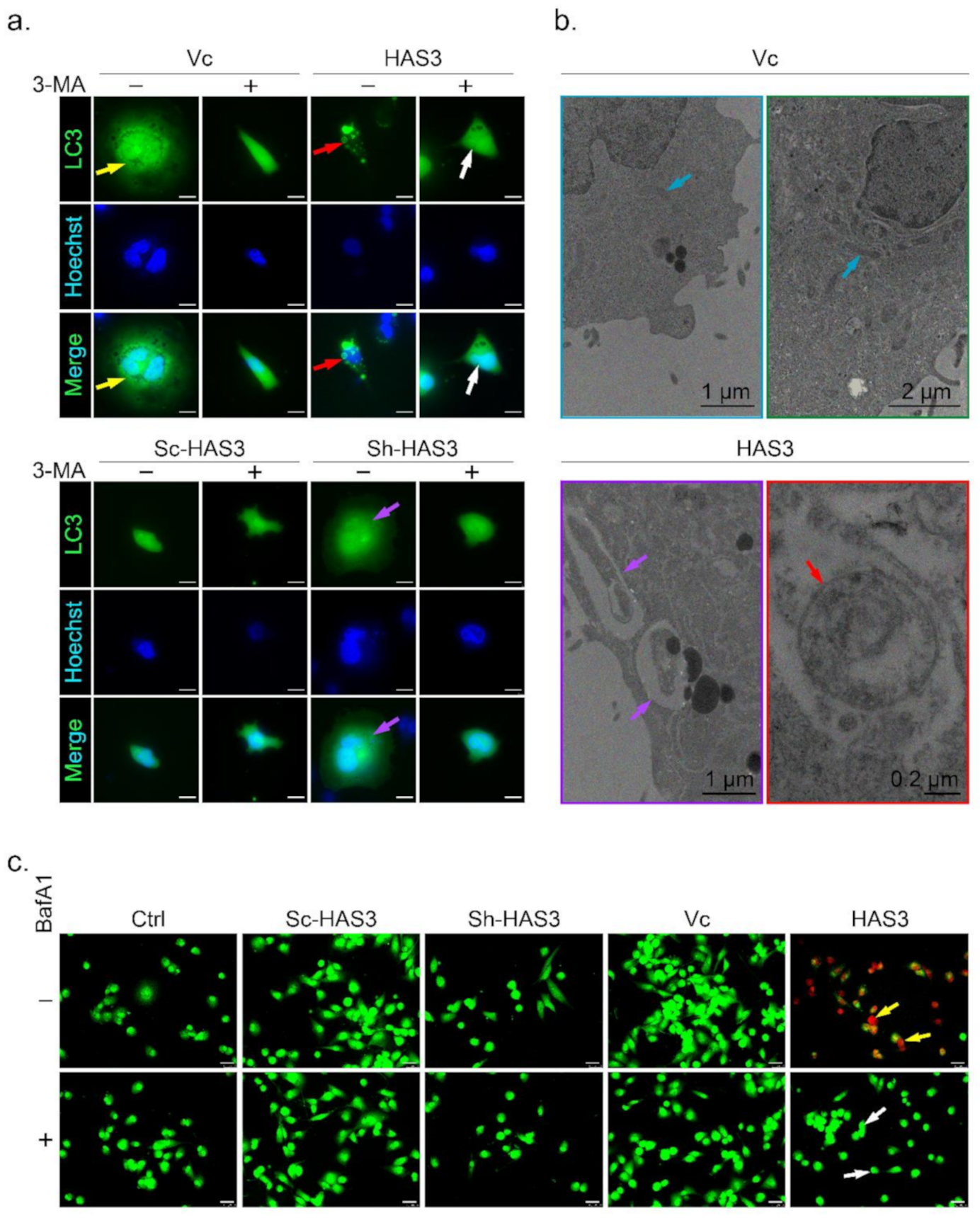
3.5. HAS3 Overexpression Promotes Breast Cancer Cell Autophagy
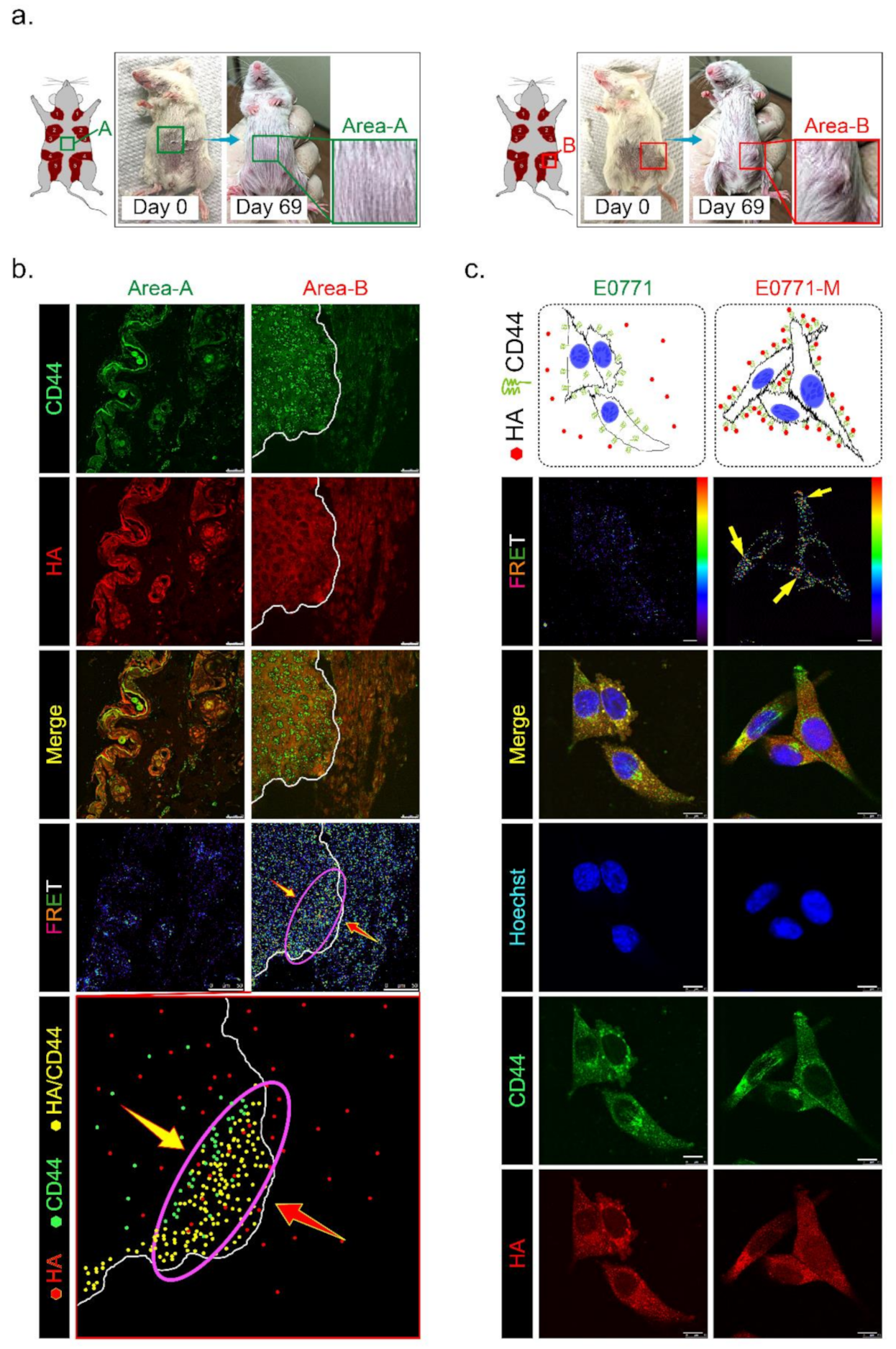
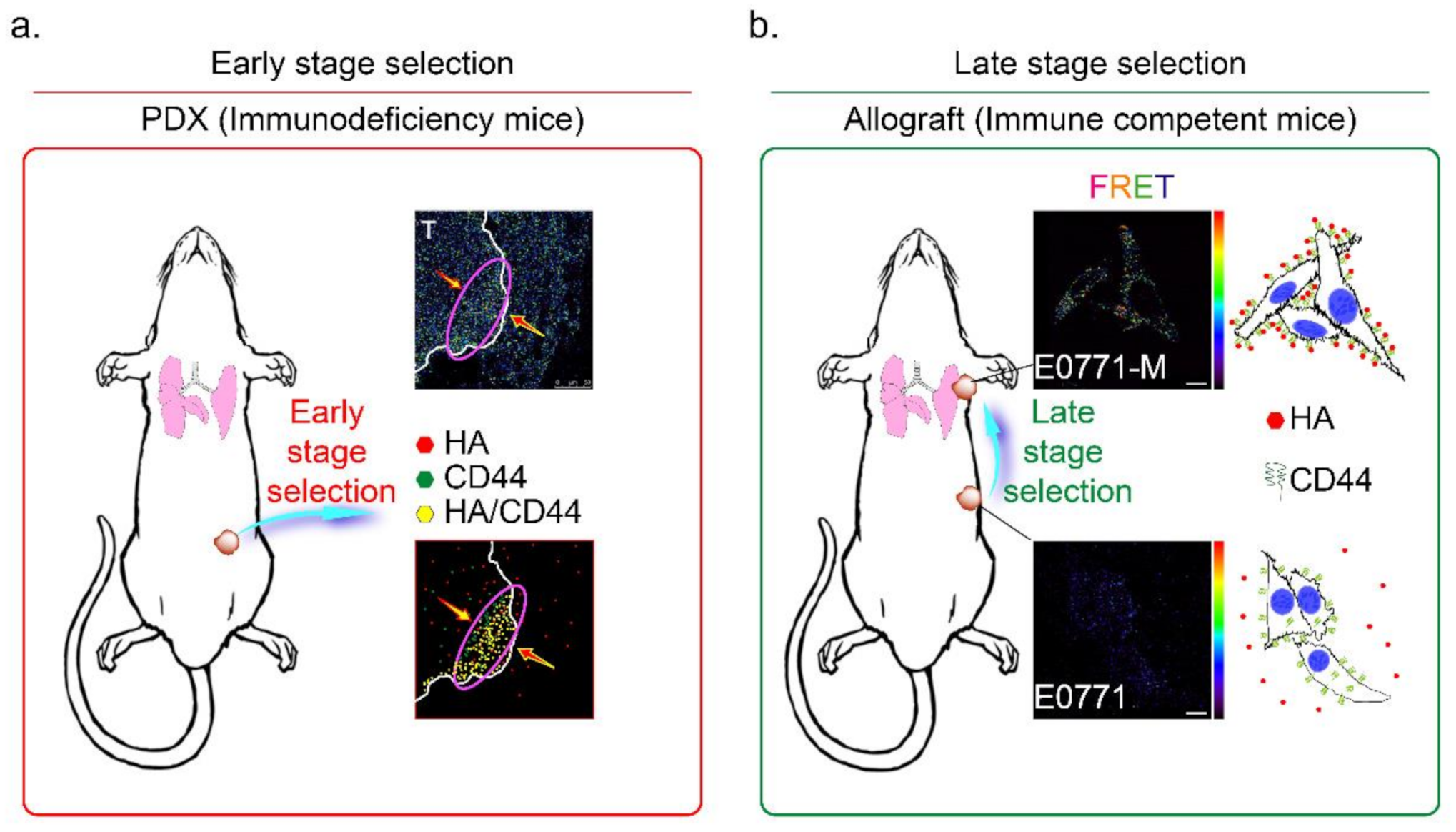
3.6. HAS3 Expression in Cutaneous Tissue Inhibits TNBC-PDX Tumor Growth in Immunodeficient Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| ECM | extracellular matrix |
| HA | hyaluronic acid |
| HAS1 | hyaluronan synthases 1 |
| HAS2 | hyaluronan synthases 2 |
| HAS3 | hyaluronan synthases 3 |
| ABC | ATP-binding cassette |
| MW | molecular weight |
| ALL | acute lymphoblastic leukemia |
| MLL | mixed lineage leukemia |
| ER | endoplasmic reticulum |
| TMZ | temozolomide |
| RIPK1 | receptor-interacting protein kinase 1 |
| ROS | reactive oxygen species |
| HB-TDFs | human breast tumor-derived fibroblasts |
| TNBC | triple-negative breast cancer |
| PDX | patient derived xenograft |
| DMEM/F12 | Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 |
| RPMI 1640 | Roswell Park Memorial Institute 1640 |
| FBS | fetal bovine serum |
| α-SMA | alpha-smooth muscle actin |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium |
| siRNA | small interfering RNA |
| scRNA | scrambled siRNA |
| BLAST | basic local alignment search tool |
| IHC | immunohistochemistry |
| PBS | phosphate-buffered saline |
| DPX | distyrene, plasticizer, xylene |
| ELISA | enzyme-linked immunosorbent assay |
| MEF | mouse embryonic fibroblasts |
| MEF-Has3-KO | HAS3 knockout mouse embryonic fibroblasts |
| PCR | polymerase chain reaction |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| HCl | hydrochloric acid |
| NaCl | sodium chloride |
| EDTA | ethylenediaminetetraacetic acid |
| EGTA | ethylene glycol tetraacetic acid |
| NaF | sodium fluoride |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| BSA | bovine serum albumin |
| TEM | transmission electron microscopy |
| ASW | artificial seawater |
| PI | propidium iodide |
| FLIM | fluorescence lifetime imaging |
| EYFP | enhanced yellow fluorescent protein |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| Cas9 | CRISPR-associated protein 9 |
| sgRNAs | single guide RNAs |
| tracrRNA | transactivating RNA |
| RNP | ribonucleoprotein |
| SCID | severe combined immunodeficient |
| IVIS | in vivo imaging system |
| NSG | non-obese diabetic–severe combined immunodeficiency–iL2Rgammanull |
| PR | progesterone receptor |
| HER2 | human epidermal growth factor receptor 2 |
References
- Costa, A.; Kieffer, Y.; Scholer-Dahirel, A.; Pelon, F.; Bourachot, B.; Cardon, M.; Sirven, P.; Magagna, I.; Fuhrmann, L.; Bernard, C.; et al. Fibroblast Heterogeneity and Immunosuppressive Environment in Human Breast Cancer. Cancer Cell 2018, 33, 463–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Schulz, T.; Schumacher, U.; Prehm, P. Hyaluronan export by the ABC transporter MRP5 and its modulation by intracellular cGMP. J. Biol. Chem. 2007, 282, 20999–21004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takabe, P.; Bart, G.; Ropponen, A.; Rilla, K.; Tammi, M.; Tammi, R.; Pasonen-Seppanen, S. Hyaluronan synthase 3 (HAS3) overexpression downregulates MV3 melanoma cell proliferation, migration and adhesion. Exp. Cell Res. 2015, 337, 1–15. [Google Scholar] [CrossRef]
- Masarwah, A.; Tammi, M.; Sudah, M.; Sutela, A.; Oikari, S.; Kosma, V.M.; Tammi, R.; Vanninen, R.; Auvinen, P. The reciprocal association between mammographic breast density, hyaluronan synthesis and patient outcome. Breast Cancer Res. Treat. 2015, 153, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Vasvani, S.; Kulkarni, P.; Rawtani, D. Hyaluronic acid: A review on its biology, aspects of drug delivery, route of administrations and a special emphasis on its approved marketed products and recent clinical studies. Int. J. Biol. Macromol. 2020, 151, 1012–1029. [Google Scholar] [CrossRef]
- Zhao, Y.; Qiao, S.; Hou, X.; Tian, H.; Deng, S.; Ye, K.; Nie, Y.; Chen, X.; Yan, H.; Tian, W. Bioengineered tumor microenvironments with naked mole rats high-molecular-weight hyaluronan induces apoptosis in breast cancer cells. Oncogene 2019, 38, 4297–4309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.F.; Qiao, S.P.; Shi, S.L.; Yao, L.F.; Hou, X.L.; Li, C.F.; Lin, F.H.; Guo, K.; Acharya, A.; Chen, X.B.; et al. Modulating Three-Dimensional Microenvironment with Hyaluronan of Different Molecular Weights Alters Breast Cancer Cell Invasion Behavior. ACS Appl. Mater. Interfaces 2017, 9, 9327–9338. [Google Scholar] [CrossRef]
- Lee, W.J.; Chen, L.C.; Lin, J.H.; Cheng, T.C.; Kuo, C.C.; Wu, C.H.; Chang, H.W.; Tu, S.H.; Ho, Y.S. Melatonin promotes neuroblastoma cell differentiation by activating hyaluronan synthase 3-induced mitophagy. Cancer Med. 2019, 8, 4821–4835. [Google Scholar] [CrossRef]
- Kim, J.E.; Kim, B.G.; Jang, Y.; Kang, S.; Lee, J.H.; Cho, N.H. The stromal loss of miR-4516 promotes the FOSL1-dependent proliferation and malignancy of triple negative breast cancer. Cancer Lett. 2020, 469, 256–265. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, G.Z.; Wen, Z.S.; Zhou, Y.C.; Huang, Y.C.; Chen, Y.; Zhou, G.B. Somatic Mutations and Splicing Variants of Focal Adhesion Kinase in Non-Small Cell Lung Cancer. J. Natl. Cancer Inst. 2018, 110, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Hellman, U.; Engstrom-Laurent, A.; Larsson, A.; Lindqvist, U. Hyaluronan concentration and molecular mass in psoriatic arthritis: Biomarkers of disease severity, resistance to treatment, and outcome. Scand. J. Rheumatol. 2019, 48, 284–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Tank, M.; Alsofyani, A.; Shah, N.; Talati, N.; LoBello, J.C.; Kim, J.R.; Oonuki, Y.; de la Motte, C.A.; Cowman, M.K. Molecular mass dependence of hyaluronan detection by sandwich ELISA-like assay and membrane blotting using biotinylated hyaluronan binding protein. Glycobiology 2013, 23, 1270–1280. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Lee, S.; Shin, S.G.; Hwang, S. Real-time PCR determination of rRNA gene copy number: Absolute and relative quantification assays with Escherichia coli. Appl. Microbiol. Biotechnol. 2008, 78, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Akash, M.S.; Rehman, K. Recent progress in biomedical applications of Pluronic (PF127): Pharmaceutical perspectives. J. Control. Release 2015, 209, 120–138. [Google Scholar] [CrossRef] [PubMed]
- Snowden, E.; Porter, W.; Hahn, F.; Ferguson, M.; Tong, F.; Parker, J.S.; Middlebrook, A.; Ghanekar, S.; Dillmore, W.S.; Blaesius, R. Immunophenotyping and Transcriptomic Outcomes in PDX-Derived TNBC Tissue. Mol. Cancer Res. 2017, 15, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, S.G.; Li, S.; Siddappa, C.M.; Ellis, M.J.; Watson, M.A.; Aft, R. Identifying biomarkers of breast cancer micrometastatic disease in bone marrow using a patient-derived xenograft mouse model. Breast Cancer Res. 2018, 20, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Punzi, S.; Meliksetian, M.; Riva, L.; Marocchi, F.; Pruneri, G.; Criscitiello, C.; Orsi, F.; Spaggiari, L.; Casiraghi, M.; Della Vigna, P.; et al. Development of Personalized Therapeutic Strategies by Targeting Actionable Vulnerabilities in Metastatic and Chemotherapy-Resistant Breast Cancer PDXs. Cells 2019, 8, 605. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Bourguignon, L.Y. Hyaluronan-CD44 interaction promotes c-Jun signaling and miRNA21 expression leading to Bcl-2 expression and chemoresistance in breast cancer cells. Mol. Cancer 2014, 13, 52. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Tolg, C.; Turley, E. Dissecting the Dual Nature of Hyaluronan in the Tumor Microenvironment. Front. Immunol. 2019, 10, 947. [Google Scholar] [CrossRef]
- Wu, R.L.; Sedlmeier, G.; Kyjacova, L.; Schmaus, A.; Philipp, J.; Thiele, W.; Garvalov, B.K.; Sleeman, J.P. Hyaluronic acid-CD44 interactions promote BMP4/7-dependent Id1/3 expression in melanoma cells. Sci. Rep. 2018, 8, 14913. [Google Scholar] [CrossRef] [Green Version]
- Bacus, S.S.; Kiguchi, K.; Chin, D.; King, C.R.; Huberman, E. Differentiation of cultured human breast cancer cells (AU-565 and MCF-7) associated with loss of cell surface HER-2/neu antigen. Mol. Carcinog. 1990, 3, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Jeng, J.H.; Wang, Y.J.; Tseng, C.J.; Liang, Y.C.; Chen, C.H.; Lee, H.M.; Lin, J.K.; Lin, C.H.; Lin, S.Y.; et al. Antitumor effects of miconazole on human colon carcinoma xenografts in nude mice through induction of apoptosis and G0/G1 cell cycle arrest. Toxicol. Appl. Pharmacol. 2002, 180, 22–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.Y.; Lee, C.H.; Chuang, Y.H.; Lee, J.Y.; Chiu, Y.Y.; Wu Lee, Y.H.; Jong, Y.J.; Hwang, J.K.; Huang, S.H.; Chen, L.C.; et al. Membrane protein-regulated networks across human cancers. Nat. Commun. 2019, 10, 3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, J.F.; Peng, Y.F.; Chen, S.; Gao, W.J.; Yang, Q.X.; Zhu, P.; Guo, J.; Tao, J.; Luo, L.; Zhang, Y.; et al. A novel harmine derivative, N-(4-(hydroxycarbamoyl)benzyl)-1-(4-methoxyphenyl)-9H-pyrido[3,4-b]indole-3-carboxamide (HBC), as histone deacetylase inhibitor: In vitro antiproliferation, apoptosis induction, cell cycle arrest, and antimetastatic effects. Eur. J. Pharmacol. 2018, 824, 78–88. [Google Scholar] [CrossRef]
- Bao, L.; Diao, H.; Dong, N.; Su, X.; Wang, B.; Mo, Q.; Yu, H.; Wang, X.; Chen, C. Histone deacetylase inhibitor induces cell apoptosis and cycle arrest in lung cancer cells via mitochondrial injury and p53 up-acetylation. Cell Biol. Toxicol. 2016, 32, 469–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahaddalkar, T.; Mehta, S.; Cheriyamundath, S.; Muthurajan, H.; Lopus, M. Tryptone-stabilized gold nanoparticles target tubulin and inhibit cell viability by inducing an unusual form of cell cycle arrest. Exp. Cell Res. 2017, 360, 163–170. [Google Scholar] [CrossRef]
- Majora, M.; Sondenheimer, K.; Knechten, M.; Uthe, I.; Esser, C.; Schiavi, A.; Ventura, N.; Krutmann, J. HDAC inhibition improves autophagic and lysosomal function to prevent loss of subcutaneous fat in a mouse model of Cockayne syndrome. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLendon, P.M.; Ferguson, B.S.; Osinska, H.; Bhuiyan, M.S.; James, J.; McKinsey, T.A.; Robbins, J. Tubulin hyperacetylation is adaptive in cardiac proteotoxicity by promoting autophagy. Proc. Natl. Acad. Sci. USA 2014, 111, E5178–E5186. [Google Scholar] [CrossRef] [Green Version]
- Cui, D.; Huo, S.; Wang, X.; Zheng, Z.; Zhang, Y.; Zhang, J.; Zhong, F. Establishment of canine macrophages stably expressing GFP-tagged canine LC3 protein for effectively detecting autophagy. Mol. Cell Probes. 2020, 49, 101493. [Google Scholar] [CrossRef]
- He, S.S.; Zhou, Q.; Luo, B.Y.; Chen, B.; Li, L.J.; Yan, F.H. Chloroquine and 3-Methyladenine Attenuates Periodontal Inflammation and Bone Loss in Experimental Periodontitis. Inflammation 2020, 43, 220–230. [Google Scholar] [CrossRef]
- Eskelinen, E.L. Maturation of autophagic vacuoles in mammalian cells. Autophagy 2005, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ren, J.Q.; He, X.L.; Chen, H.; Wei, T.T.; Feng, W. YWHA/14-3-3 proteins recognize phosphorylated TFEB by a noncanonical mode for controlling TFEB cytoplasmic localization. Autophagy 2019, 15, 1017–1030. [Google Scholar] [CrossRef]
- Mauffrey, P.; Tchitchek, N.; Barroca, V.; Bemelmans, A.; Firlej, V.; Allory, Y.; Romeo, P.H.; Magnon, C. Progenitors from the central nervous system drive neurogenesis in cancer. Nature 2019, 569, 672–678. [Google Scholar] [CrossRef]
- Elia, I.; Rossi, M.; Stegen, S.; Broekaert, D.; Doglioni, G.; van Gorsel, M.; Boon, R.; Escalona-Noguero, C.; Torrekens, S.; Verfaillie, C.; et al. Breast cancer cells rely on environmental pyruvate to shape the metastatic niche. Nature 2019, 568, 117–121. [Google Scholar] [CrossRef]
- Ligorio, M.; Sil, S.; Malagon-Lopez, J.; Nieman, L.T.; Misale, S.; Di Pilato, M.; Ebright, R.Y.; Karabacak, M.N.; Kulkarni, A.S.; Liu, A.; et al. Stromal Microenvironment Shapes the Intratumoral Architecture of Pancreatic Cancer. Cell 2019, 178, 160–175. [Google Scholar] [CrossRef] [PubMed]
- Corte, M.D.; Gonzalez, L.O.; Lamelas, M.L.; Alvarez, A.; Junquera, S.; Allende, M.T.; Garcia-Muniz, J.L.; Arguelles, J.; Vizoso, F.J. Expression and clinical signification of cytosolic hyaluronan levels in invasive breast cancer. Breast Cancer Res. Treat. 2006, 97, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Tavianatou, A.G.; Caon, I.; Franchi, M.; Piperigkou, Z.; Galesso, D.; Karamanos, N.K. Hyaluronan: Molecular size-dependent signaling and biological functions in inflammation and cancer. FEBS J. 2019, 286, 2883–2908. [Google Scholar] [CrossRef] [PubMed]
- Urakawa, H.; Nishida, Y.; Knudson, W.; Knudson, C.B.; Arai, E.; Kozawa, E.; Futamura, N.; Wasa, J.; Ishiguro, N. Therapeutic potential of hyaluronan oligosaccharides for bone metastasis of breast cancer. J. Orthop. Res. 2012, 30, 662–672. [Google Scholar] [CrossRef]
- Al-Othman, N.; Alhendi, A.; Ihbaisha, M.; Barahmeh, M.; Alqaraleh, M.; Al-Momany, B.Z. Role of CD44 in breast cancer. Breast Dis. 2020, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Yang, J.; Feng, X.; Pandey, A.; Barhoumi, R.; Zhang, D.; Bell, S.L.; Liu, Y.; da Costa, L.F.; Rice-Ficht, A.; et al. Interactions between fungal hyaluronic acid and host CD44 promote internalization by recruiting host autophagy proteins to forming phagosomes. iScience 2021, 24, 102192. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Green, D.R. Autophagy-Independent Functions of the Autophagy Machinery. Cell 2019, 177, 1682–1699. [Google Scholar] [CrossRef]
- Hu, J.B.; Li, S.J.; Kang, X.Q.; Qi, J.; Wu, J.H.; Wang, X.J.; Xu, X.L.; Ying, X.Y.; Jiang, S.P.; You, J.; et al. CD44-targeted hyaluronic acid-curcumin prodrug protects renal tubular epithelial cell survival from oxidative stress damage. Carbohydr. Polym. 2018, 193, 268–280. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Total No. of Patients | HAS3 (N > T) | HAS3 (N < T) | ||||
|---|---|---|---|---|---|---|---|
| No. | Mean Fold Change and 95% CI | P | No. | Mean Fold Change and 95% CI | P | ||
| Age | 0.75 | 0.28 | |||||
| <45 y | 73 | 53 | 89.49 (16.0 to 163.0) | 20 | 0.26 (0.1 to 0.4) | ||
| ≥45y | 258 | 197 | 109.14 (50.4 to 167.9) | 61 | 0.33 (0.3 to 0.4) | ||
| Nodal status | 0.38 | 0.99 | |||||
| N0 | 152 | 119 | 150.77 (55.1 to 246.4) | 33 | 0.31 (0.2 to 0.4) | ||
| N1 | 91 | 69 | 57.08 (8.7 to 105.4) | 22 | 0.3 (0.2 to 0.4) | ||
| N2 | 42 | 33 | 66.95 (11.7 to 122.2) | 9 | 0.33 (0.1 to 0.6) | ||
| N3 | 36 | 25 | 78.12 (−12.8 to 169.0) | 11 | 0.32 (0.1 to 0.5) | ||
| Stage | 0.85 | 0.03 | |||||
| 0 | 7 | 5 | 8.45 (−2.5 to 19.4) | 2 | 0.62 (−0.7 to 1.9) | ||
| I | 71 | 56 | 104.39 (29.4 to 179.4) | 15 | 0.39 (0.3 to 0.5) | ||
| II | 159 | 119 | 126.43 (33.4 to 219.4) | 40 | 0.27 (0.2 to 0.3) | ||
| III | 91 | 69 | 70.25 (24.7 to 115.8) | 22 | 0.29 (0.2 to 0.4) | ||
| IV | 2 | 1 | 265.1 | 1 | 0.89 | ||
| ER status | 0.76 | 0.46 | |||||
| Negative | 78 | 60 | 91.27 (16.5 to 166.1) | 18 | 0.36 (0.2 to 0.5) | ||
| Positive | 252 | 190 | 109.28 (49.5 to 169.1) | 62 | 0.3 (0.2 to 0.4) | ||
| PR status | 0.19 | 0.75 | |||||
| Negative | 137 | 103 | 143.78 (34.3 to 253.3) | 34 | 0.33 (0.2 to 0.4) | ||
| Positive | 190 | 145 | 78.63 (46.1 to 111.2) | 45 | 0.31 (0.2 to 0.4) | ||
| Her-2 status | 0.99 | 0.69 | |||||
| Negative | 237 | 180 | 105.8 (42.9 to 168.6) | 57 | 0.31 (0.2 to 0.4) | ||
| Positive | 77 | 56 | 105.54 (24.7 to 186.4) | 21 | 0.34 (0.2 to 0.5) | ||
| Path | 0.98 | 0.22 | |||||
| DCIS | 12 | 8 | 11.27 (−1.6 to 23.9) | 4 | 0.57 (0.3 to 0.9) | ||
| Infiltration ductal carcinoma | 289 | 218 | 108.98 (54.7 to 163.3) | 71 | 0.29 (0.2 to 0.4) | ||
| Mucinous carcinoma | 8 | 6 | 105.15 (−96.2 to 306.5) | 2 | 0.33 (−3.6 to 4.3) | ||
| ILC | 13 | 9 | 31.75 (0.4 to 63.1) | 4 | 0.35 (−0.2 to 0.9) | ||
| IPC | 1 | 1 | 2.75 | 0 | |||
| Mix | 8 | 8 | 182.74 (−228.4 to 593.9) | 0 | |||
| other | 1 | 1 | 16.44 | 0 | |||
| Characteristic | Total No. of Patients | HAS2 (N > T) | HAS2 (N < T) | ||||
|---|---|---|---|---|---|---|---|
| No. | Mean Fold Change and 95% CI | P | No. | Mean Fold Change and 95% CI | P | ||
| Age | 0.11 | 0.18 | |||||
| <45 y | 48 | 23 | 327.2 (−58.0 to 607.7) | 25 | 32.2 (−0.1 to 0.2) | ||
| ≥45y | 179 | 77 | 52.4 (−356.5 to 906.0) | 102 | 432.8 (−0.1 to 0.2) | ||
| Nodal status | 0.75 | 0.53 | |||||
| N0 | 107 | 51 | 197.0 (−242.3 to 537.5) | 56 | 510.9 (−271.5 to 984.7) | ||
| N1 | 62 | 26 | 49.4 (−137.4 to 432.6) | 36 | 154.3 (−170.0 to 883.2) | ||
| N2 | 24 | 12 | 12.9 (−11.5 to 18.7) | 12 | 53.3 (−716 to 175.2) | ||
| N3 | 25 | 7 | 9.3 (−10.7 to 17.9) | 18 | 323.7 (−644.1 to 103.3) | ||
| Stage | 0.69 | 0.31 | |||||
| 0 | 2 | 0 | 0 | 2 | 46.1 (−3491.5 to 2050.6) | ||
| I | 49 | 20 | 113.9 (−520.2 to 419.9) | 29 | 766.5 (−2050.6 to 3491.5) | ||
| II | 116 | 55 | 164.1 (−419.9 to 520.2) | 61 | 264.4 (−2505.5 to 2942) | ||
| III | 45 | 23 | 10.8 (−653.6 to 447.2) | 22 | 191.0 (−2613.0 to 2902.7) | ||
| IV | 0 | 0 | 0 | 0 | 0 | ||
| ER status | 0.47 | 0.76 | |||||
| Negative | 47 | 21 | 16.0 (−473.9 to 221.6) | 26 | 284.3 (−677.4 to 494.9) | ||
| Positive | 179 | 79 | 142.2 (−305.5 to 53.3) | 100 | 375.5 (−561.7 to 379.2) | ||
| PR status | 0.39 | 0.32 | |||||
| Negative | 86 | 43 | 45.9 (−408.1 to 163.5) | 43 | 193.9 (−753.7 to 249.3) | ||
| Positive | 139 | 57 | 168.3 (−374.4 to 129.7) | 82 | 446.1 (−662.9 to 158.6) | ||
| Her-2 status | 0.59 | 0.26 | |||||
| Negative | 195 | 88 | 130.1 (−315.9 to 556.7) | 107 | 411.6 (−277.2 to 1009.2) | ||
| Positive | 32 | 12 | 9.8 (−40.8 to 281.5) | 20 | 45.5 (82.2 to 649.8) | ||
| Path | 0.99 | 0.96 | |||||
| DCIS | 5 | 0 | 0 | 5 | 26.2 (−1704.6 to 909.1) | ||
| Infiltration ductal carcinoma | 193 | 89 | 125.2 (−689.3 to 817.3) | 104 | 423.9 (−684.9 to −110.5) | ||
| Mucinous carcinoma | 9 | 4 | 61.2 (−130.7 to 258.6) | 5 | 3.7 (−171.4 to 15.2) | ||
| ILC | 12 | 6 | 29.7 (−97.7 to 153.4) | 6 | 81.8 (−173.9 to 17.8) | ||
| IPC | 1 | 0 | 0 | 1 | 10.9 | ||
| Mix | 0 | 0 | 0 | 0 | 0 | ||
| other | 6 | 1 | 1.86 | 5 | 39.9 (−219.4 to 161.4) | ||
| Characteristic | Total No. of Patients | HAS1 (N > T) | HAS1 (N < T) | ||||
|---|---|---|---|---|---|---|---|
| No. | Mean Fold Change and 95% CI | P | No. | Mean Fold Change and 95% CI | P | ||
| Age | 0.75 | 0.29 | |||||
| <45 y | 52 | 36 | 63.9 (−39.7 to 55.1) | 16 | 19.4 (−179.2 to 50.2) | ||
| ≥45y | 175 | 113 | 56.2 (−47.6 to 63.0) | 62 | 83.9 (−124.7 to −4.3) | ||
| Nodal status | 0.18 | 0.59 | |||||
| N0 | 112 | 78 | 66.9 (−42.5 to 87.0) | 34 | 76.5 (−222.7 to 134.5) | ||
| N1 | 71 | 53 | 44.7 (−160.9 to 41.8) | 18 | 120.6 (−181.8 to 258.9) | ||
| N2 | 27 | 17 | 104.3 (−36.9 to 220.6) | 10 | 37.9 (−324.3 to 158.9) | ||
| N3 | 26 | 15 | 12.4 (−157.0 to 47.9) | 11 | 16.3 (−272.7 to 152.3) | ||
| Stage | 0.84 | 0.11 | |||||
| 0 | 3 | 2 | 5.9 (−2.5 to 19.4) | 1 | 1.14 | ||
| I | 53 | 38 | 44.3 (29.4 to 179.4) | 15 | 112.7 (−768.1 to 544.9) | ||
| II | 123 | 88 | 62.2 (33.4 to 219.4) | 35 | 70.1 (−49.4 to 133.9) | ||
| III | 64 | 40 | 54.8 (24.7 to 115.8) | 24 | 27.8 (−36.4 to 120.9) | ||
| IV | 1 | 0 | 0 | 1 | 566.2 | ||
| ER status | 0.68 | 0.77 | |||||
| Negative | 52 | 36 | 50.0 (−57.3 to 37.5) | 16 | 85.3 (−99.1 to 133.9) | ||
| Positive | 194 | 133 | 59.9 (−53.5 to 33.7) | 61 | 67.9 (−83.6 to 118.5) | ||
| PR status | 0.65 | 0.53 | |||||
| Negative | 90 | 65 | 63.7 (−30.8 to 49.4) | 25 | 49.8 (−132.9 to 68.5) | ||
| Positive | 155 | 103 | 54.4 (−32.1 to 50.6) | 52 | 81.9 (−114.4 to 50.1) | ||
| Her-2 status | 0.66 | 0.89 | |||||
| Negative | 209 | 143 | 59.7 (−41.9 to 65.7) | 66 | 69.3 (−138.4 to 120.5) | ||
| Positive | 38 | 26 | 47.7 (−32.0 to 55.9) | 12 | 78.2 (−130.2 to 112.3) | ||
| Path | 0.86 | 0.85 | |||||
| DCIS | 6 | 3 | 4.4 (−212.7 to 94.7) | 3 | 3.3 (0.3 to 0.9) | ||
| Infiltration ductal carcinoma | 212 | 142 | 63.5 (−81.9 to −36.1) | 70 | 73.5 (0.2 to 0.4) | ||
| Mucinous carcinoma | 7 | 7 | 19.0 (−14.9 to 20.1) | 0 | 0 | ||
| ILC | 13 | 8 | 16.5 (−15.2 to 20.3) | 5 | 71.8 (−0.2 to 0.9) | ||
| IPC | 1 | 1 | 7.45 | 0 | |||
| Mix | 0 | 0 | 0 | 0 | 0 | ||
| other | 7 | 7 | 66.8 (−466.7 to 347.9) | 0 | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, W.-J.; Tu, S.-H.; Cheng, T.-C.; Lin, J.-H.; Sheu, M.-T.; Kuo, C.-C.; Changou, C.A.; Wu, C.-H.; Chang, H.-W.; Chang, H.-L.; et al. Type-3 Hyaluronan Synthase Attenuates Tumor Cells Invasion in Human Mammary Parenchymal Tissues. Molecules 2021, 26, 6548. https://doi.org/10.3390/molecules26216548
Lee W-J, Tu S-H, Cheng T-C, Lin J-H, Sheu M-T, Kuo C-C, Changou CA, Wu C-H, Chang H-W, Chang H-L, et al. Type-3 Hyaluronan Synthase Attenuates Tumor Cells Invasion in Human Mammary Parenchymal Tissues. Molecules. 2021; 26(21):6548. https://doi.org/10.3390/molecules26216548
Chicago/Turabian StyleLee, Wen-Jui, Shih-Hsin Tu, Tzu-Chun Cheng, Juo-Han Lin, Ming-Thau Sheu, Ching-Chuan Kuo, Chun A. Changou, Chih-Hsiung Wu, Hui-Wen Chang, Hang-Lung Chang, and et al. 2021. "Type-3 Hyaluronan Synthase Attenuates Tumor Cells Invasion in Human Mammary Parenchymal Tissues" Molecules 26, no. 21: 6548. https://doi.org/10.3390/molecules26216548
APA StyleLee, W.-J., Tu, S.-H., Cheng, T.-C., Lin, J.-H., Sheu, M.-T., Kuo, C.-C., Changou, C. A., Wu, C.-H., Chang, H.-W., Chang, H.-L., Chen, L.-C., & Ho, Y.-S. (2021). Type-3 Hyaluronan Synthase Attenuates Tumor Cells Invasion in Human Mammary Parenchymal Tissues. Molecules, 26(21), 6548. https://doi.org/10.3390/molecules26216548