
Towards the Use of Adsorption Methods for the Removal of Purines from Beer



Abstract
:1. Fermented Alcoholic Beverages
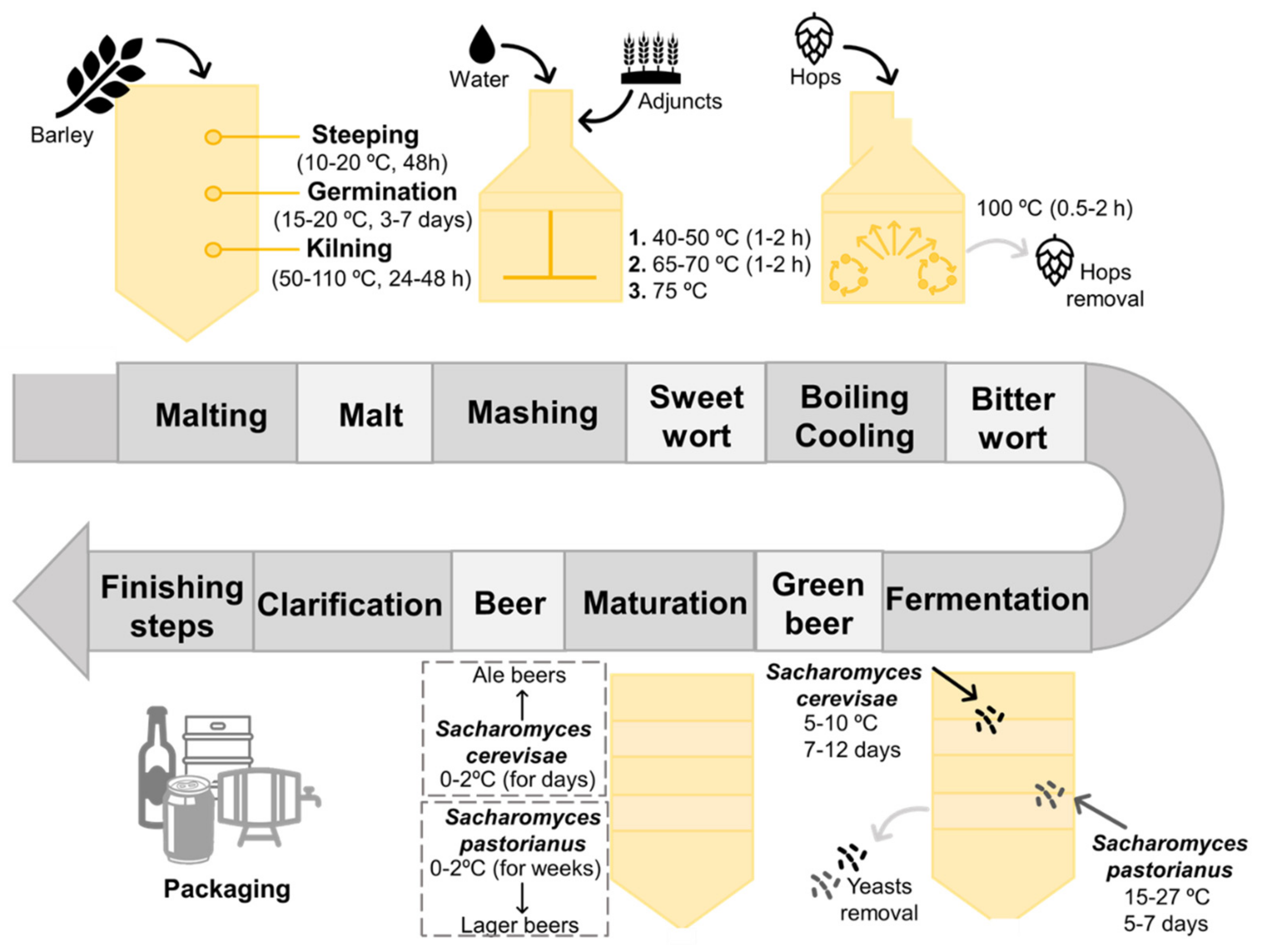
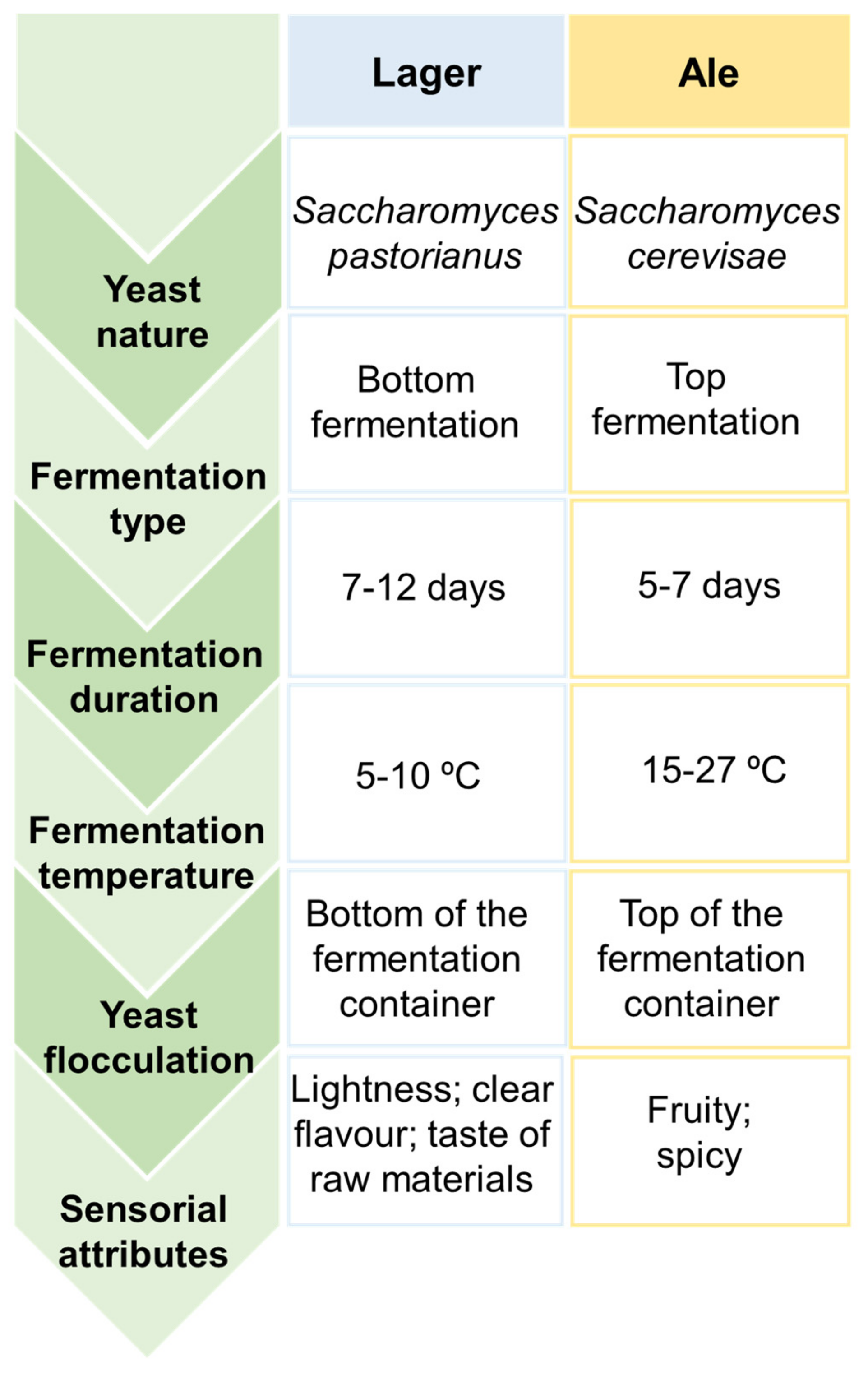
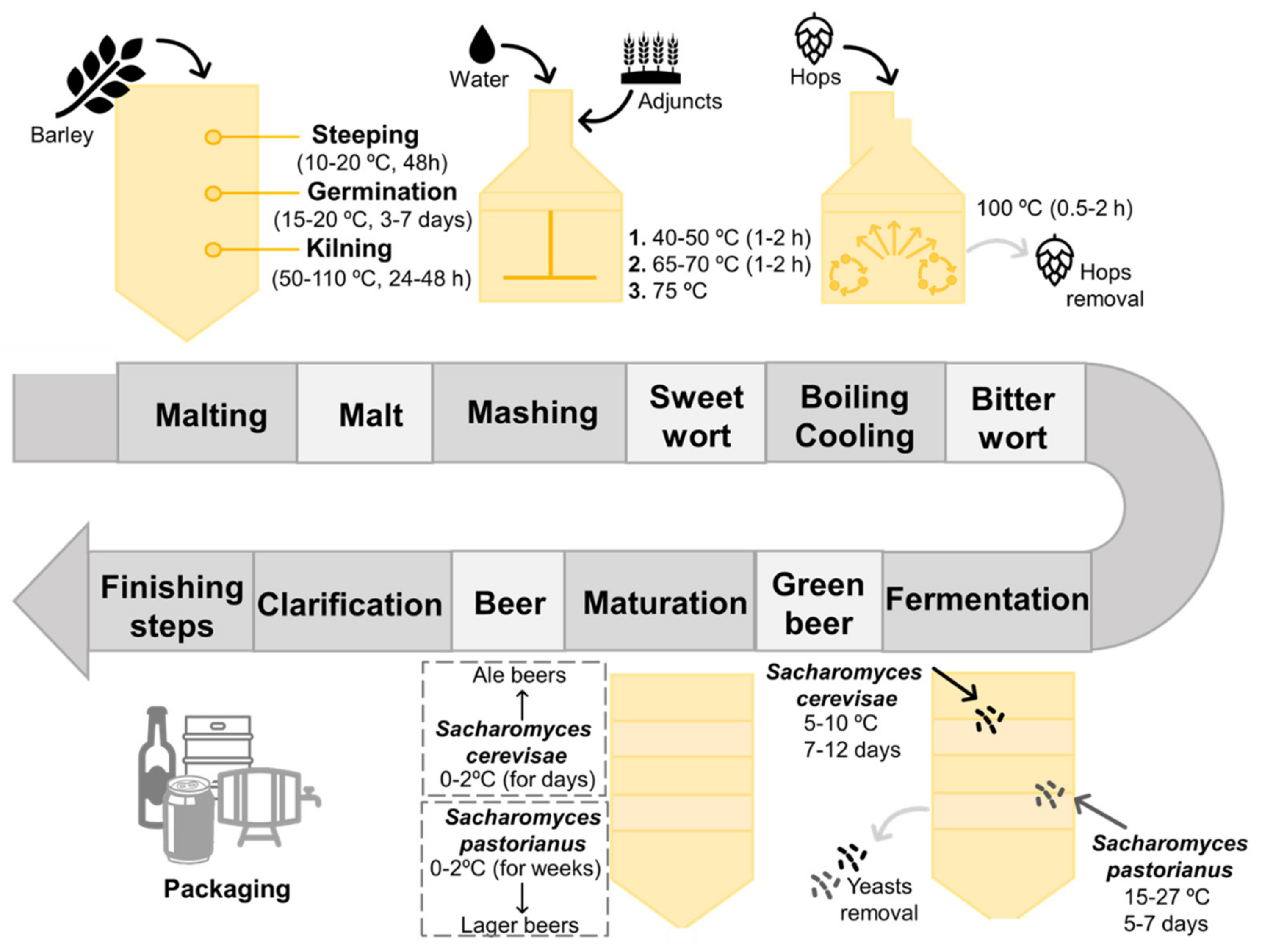
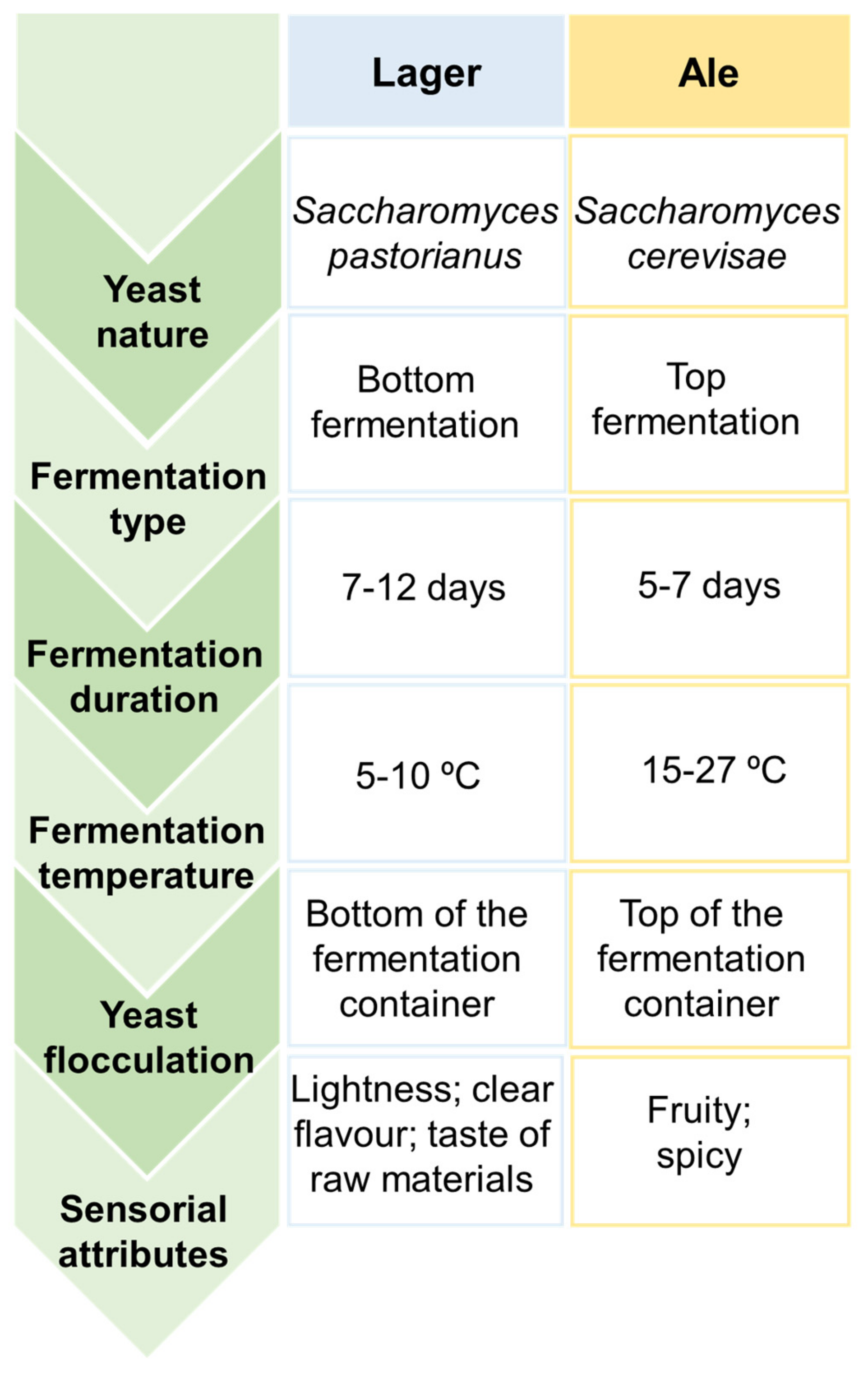
2. Beer, a Fermented Alcoholic Beverage
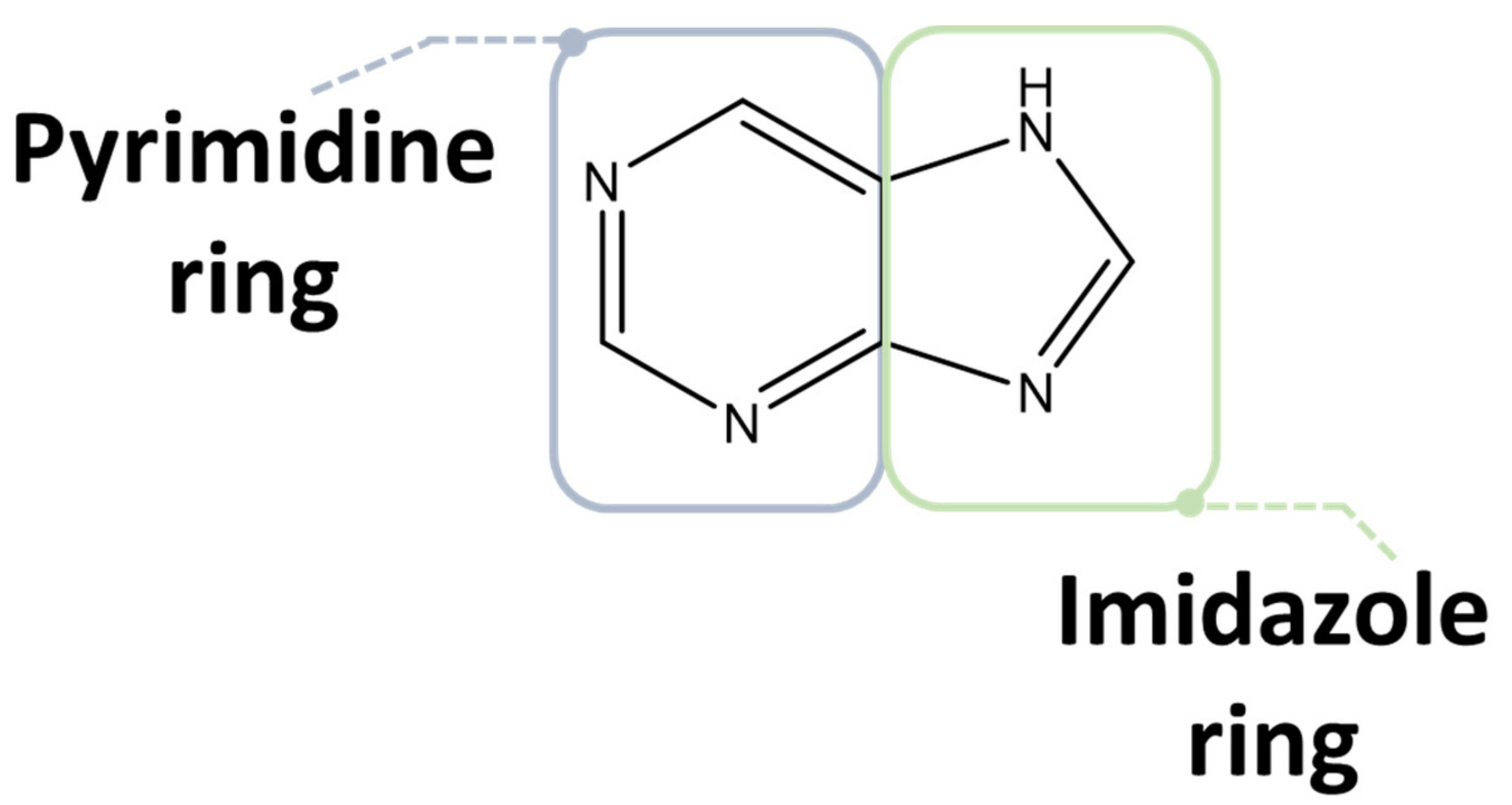
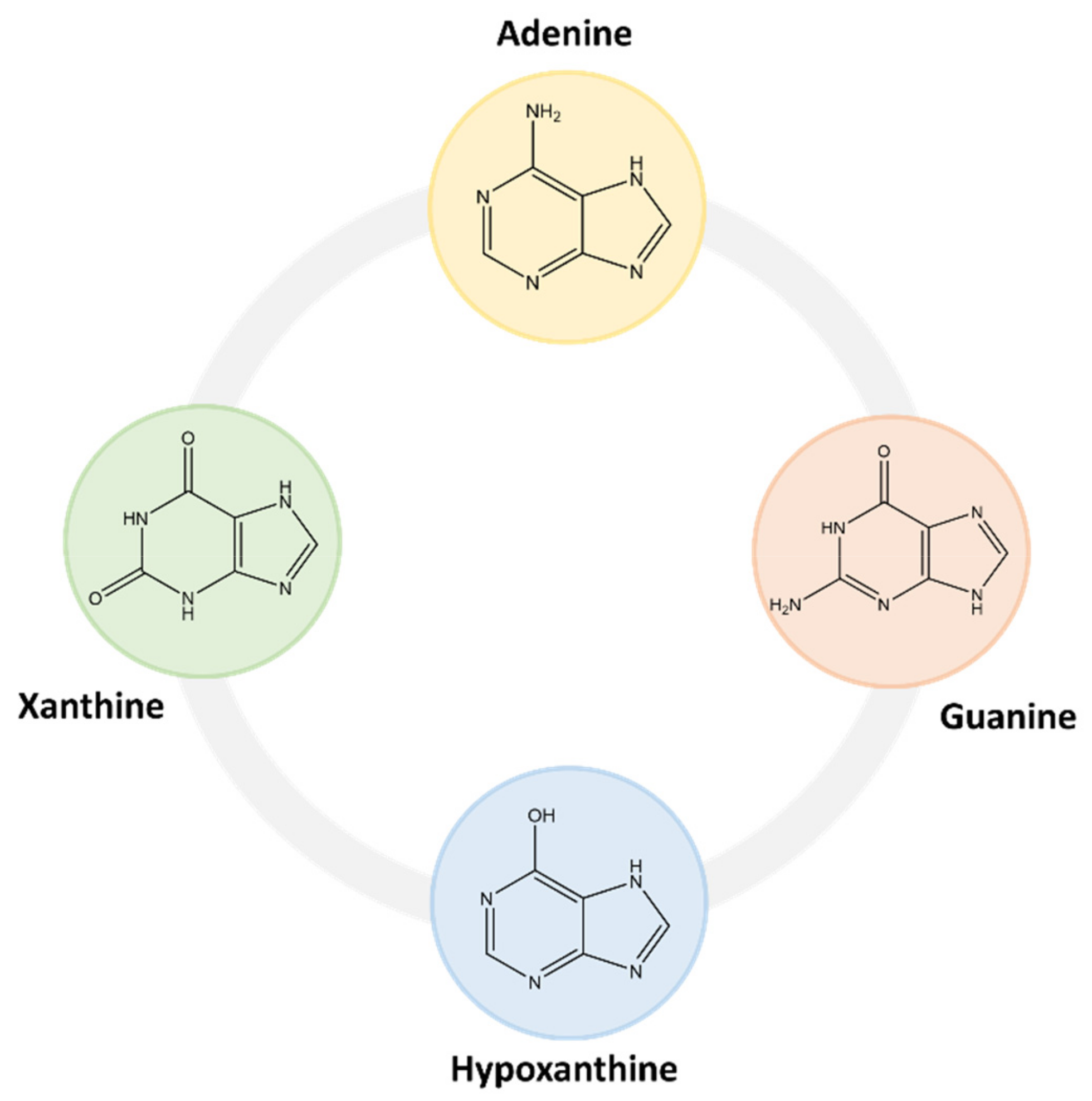
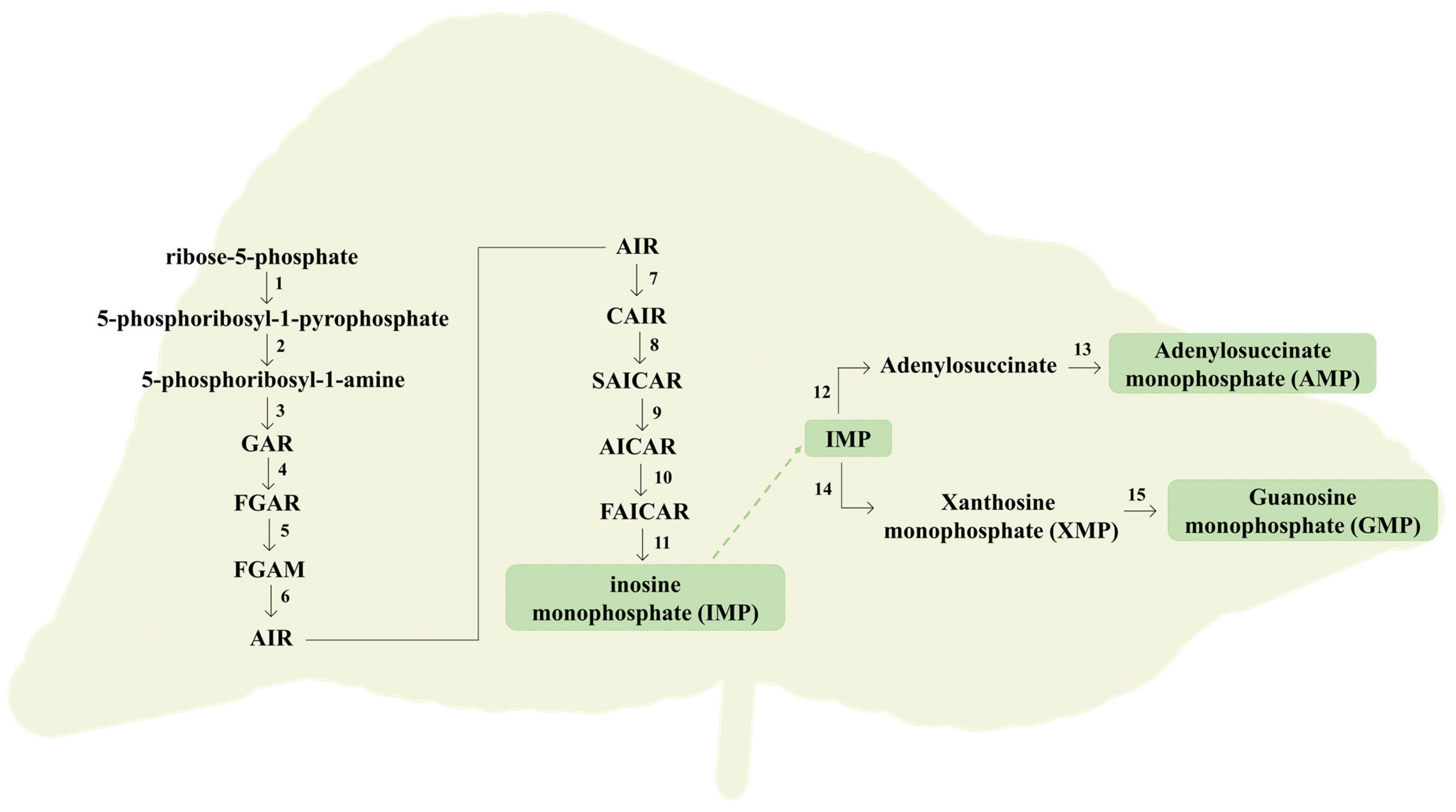
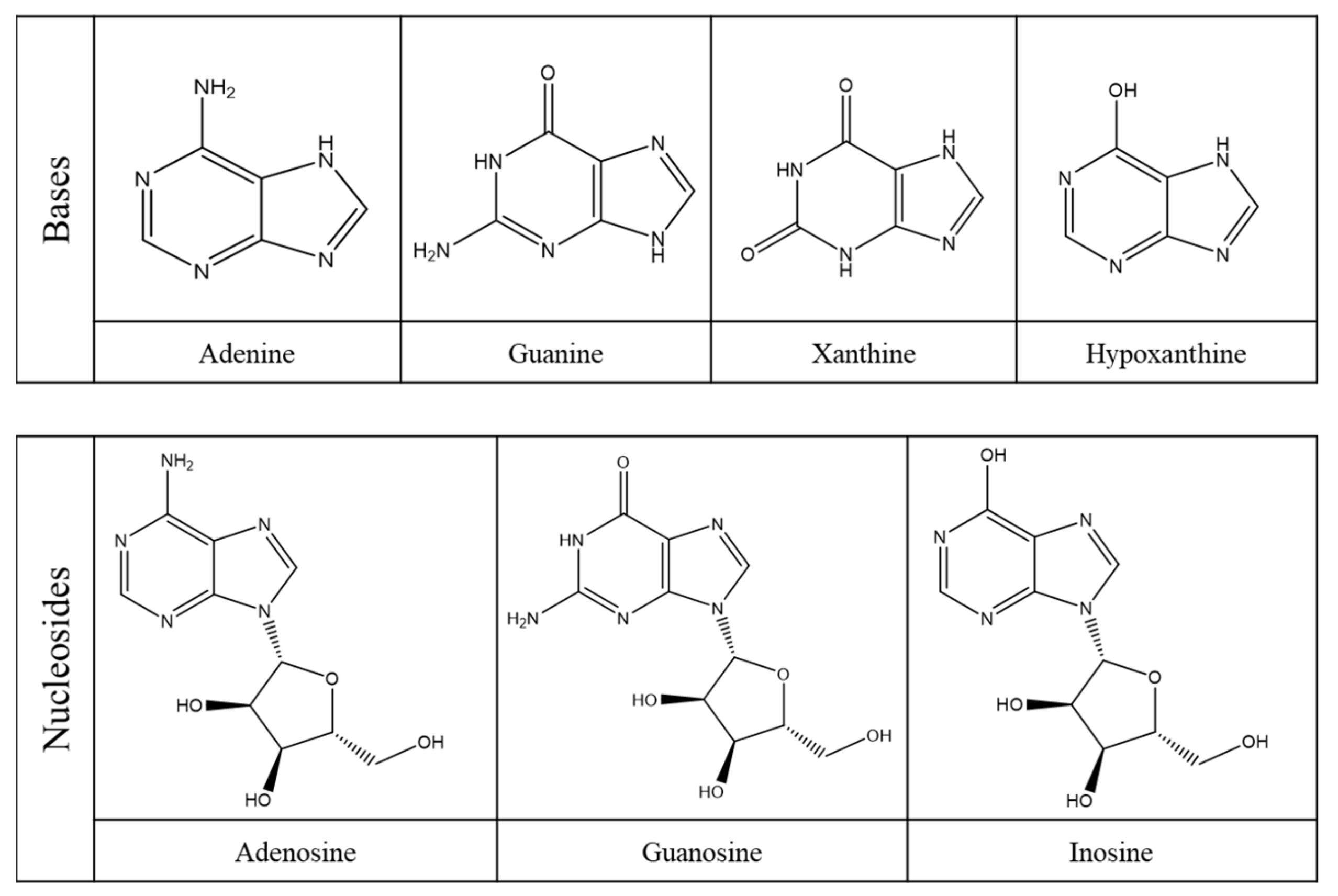
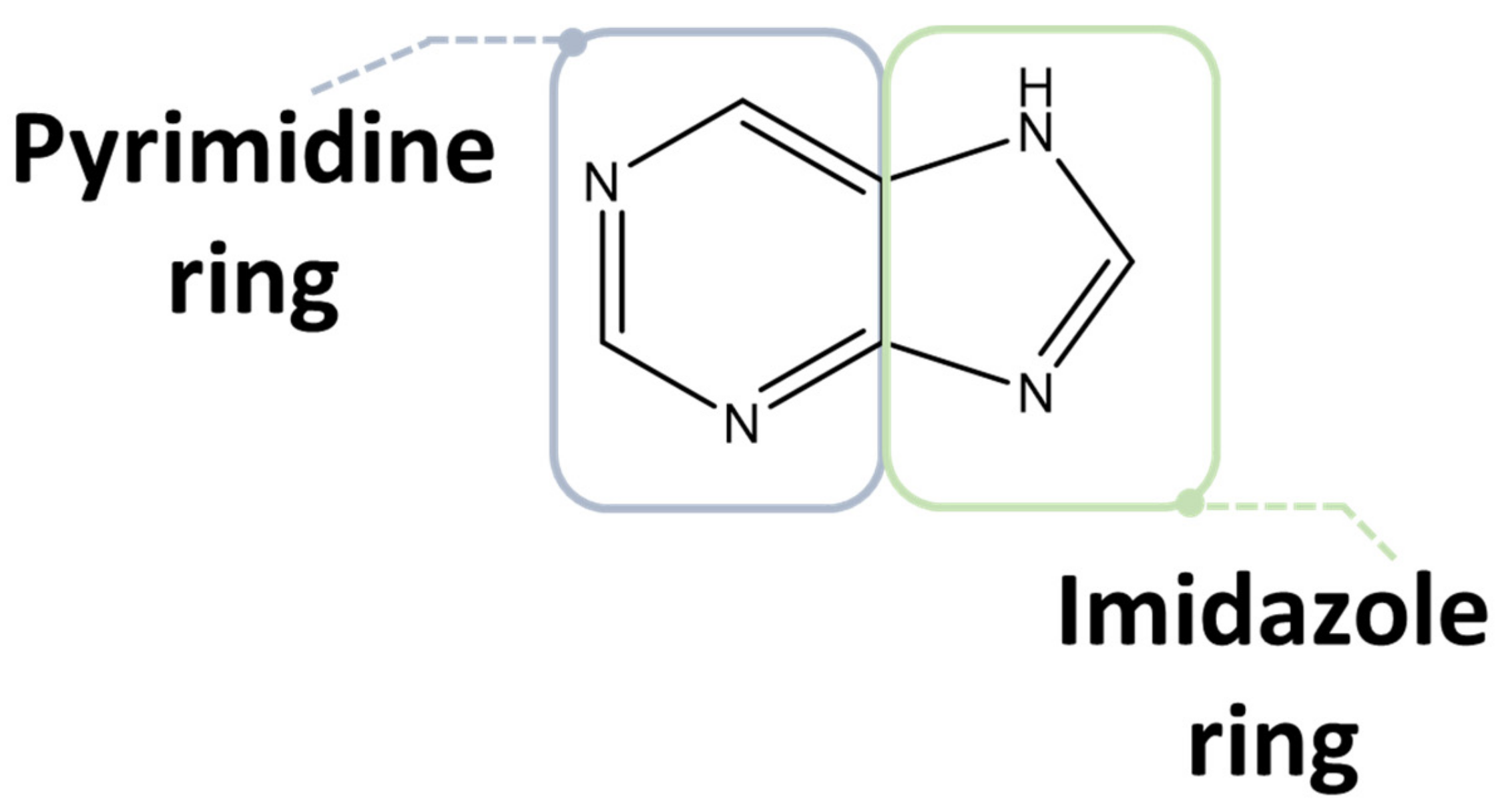
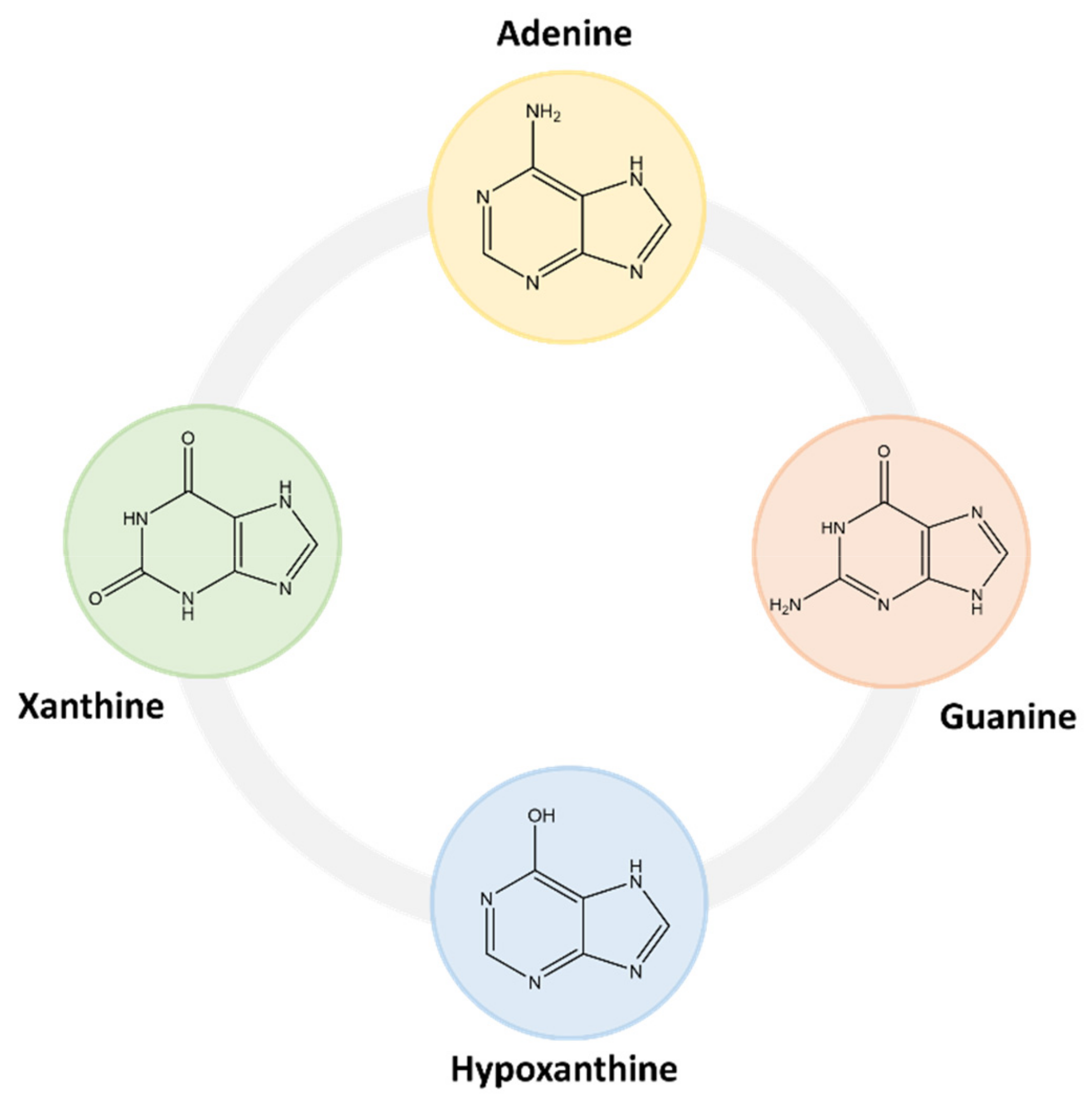
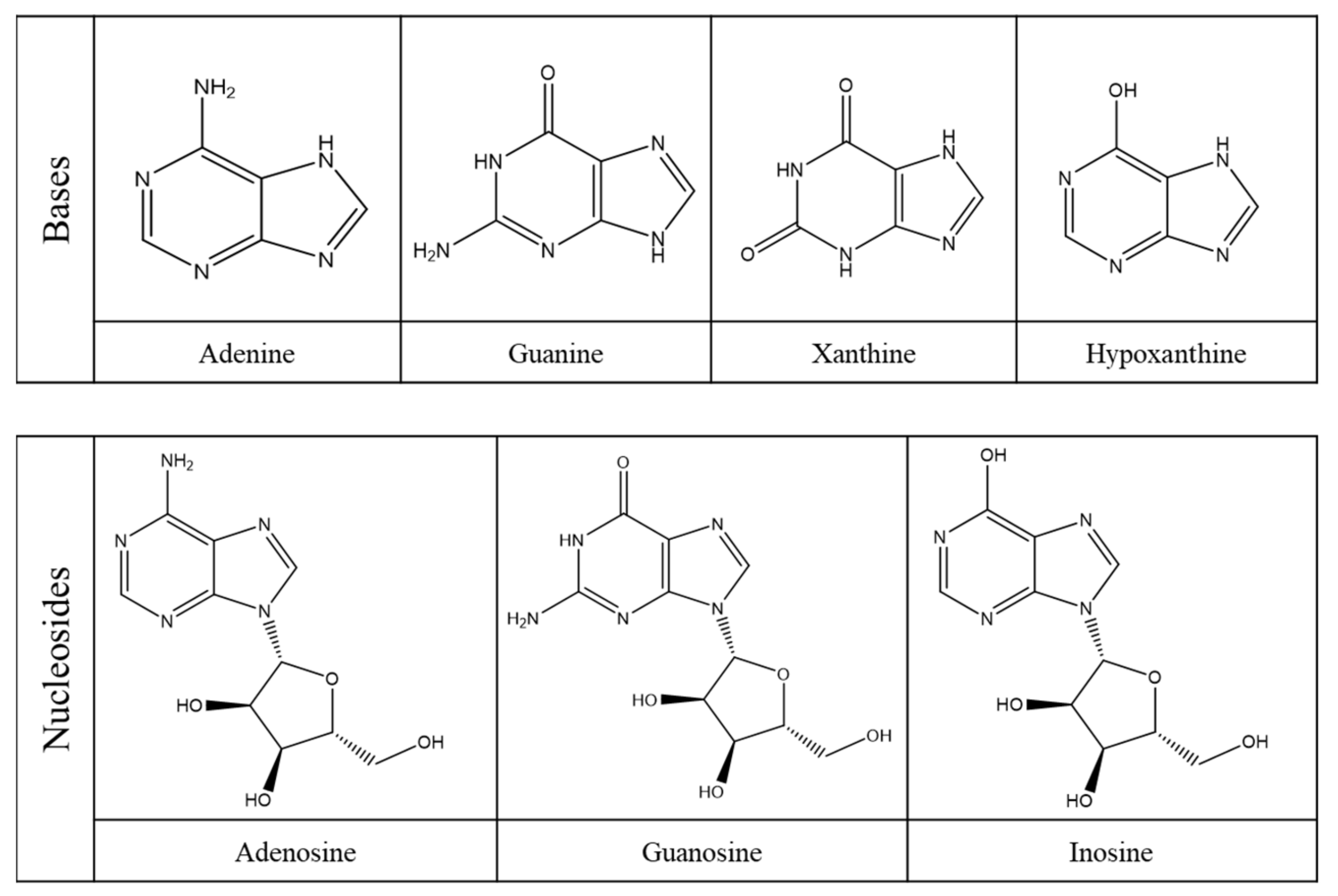
2.1. Purine Compounds in Beer
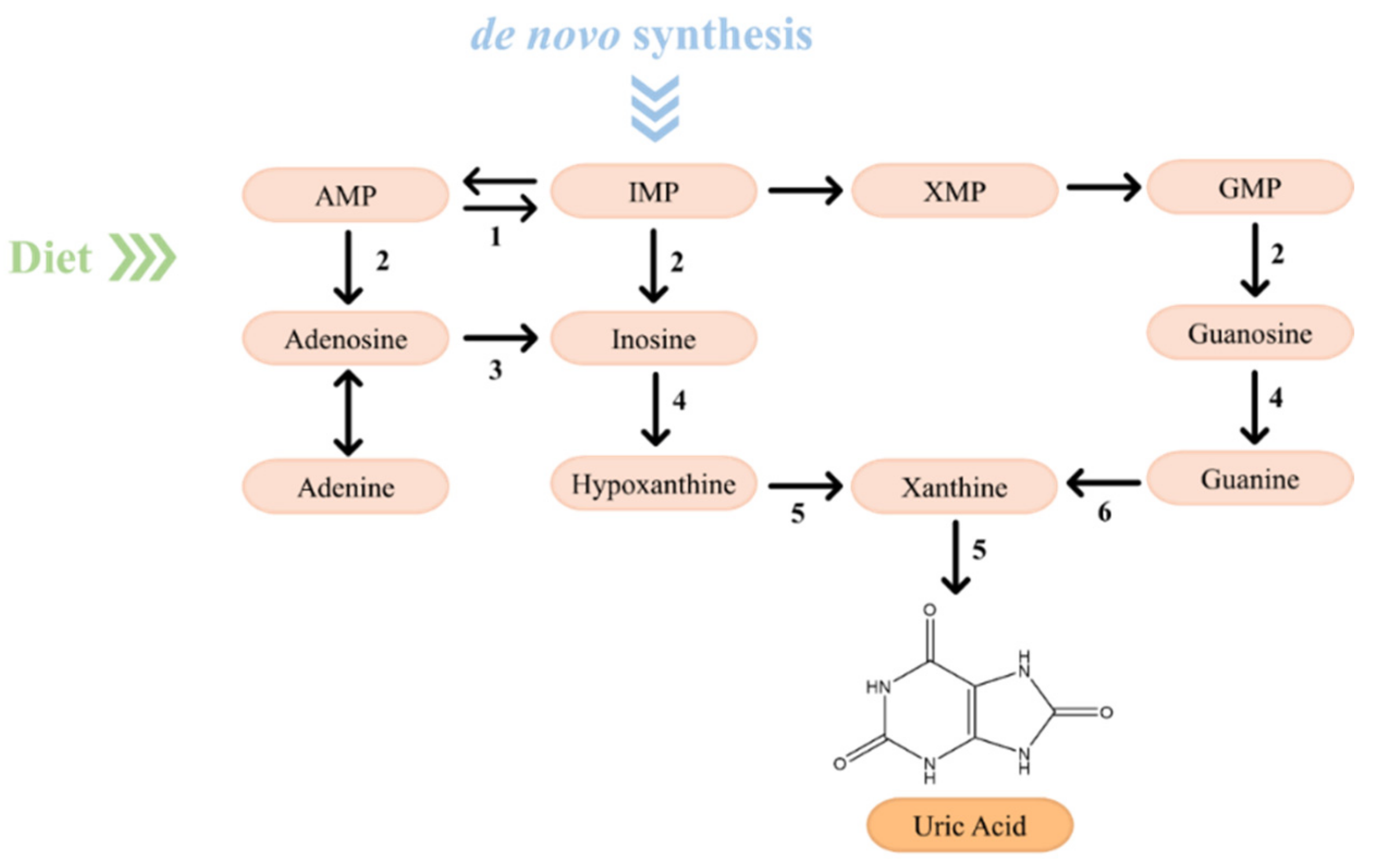
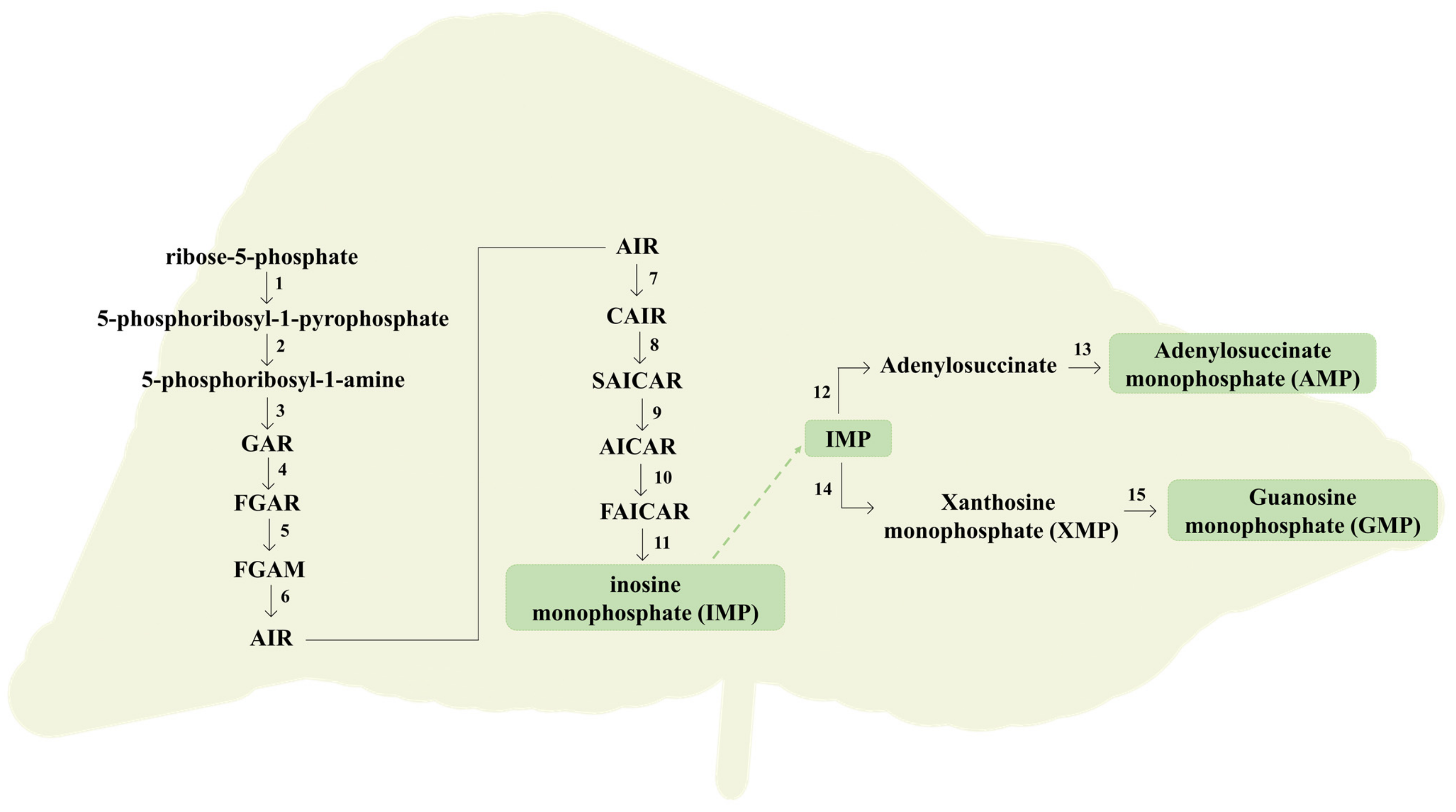
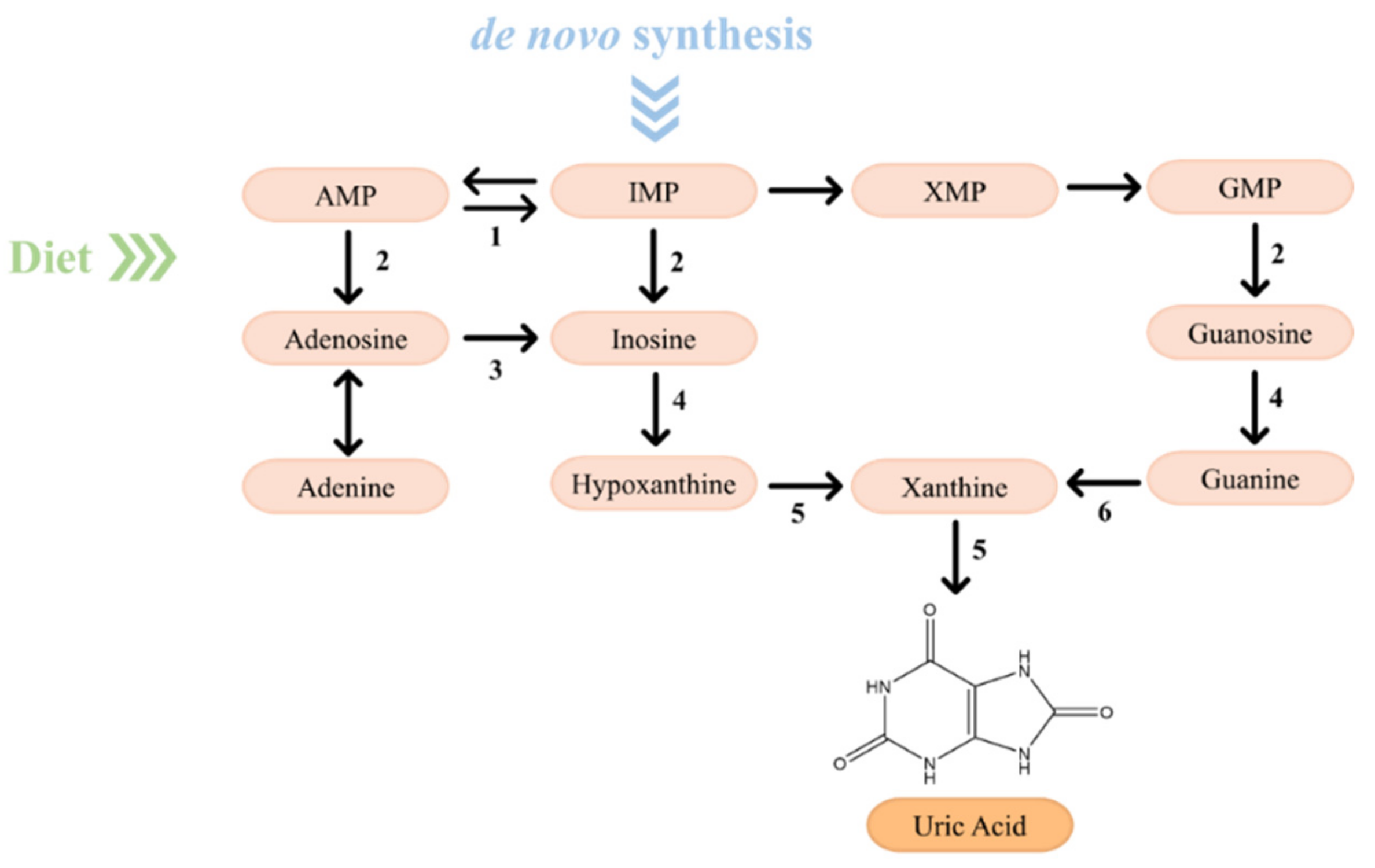
2.2. From Purine Compounds Catabolism to Hyperuricemia and Gout
3. Removal of Purine Compounds from Beer
3.1. Enzymatic, Biological and Processing Methods
3.2. Adsorption Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nogueira, L.C.; Couri, S.; Trugo, N.F.; Lollo, P.C.B. The effect of different alcoholic beverages on blood alcohol levels, plasma insulin and plasma glucose in humans. Food Chem. 2014, 158, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, J.; Rosenberg, D.; Zhao, H.; Lengyel, G.; Nadel, D. Fermented beverage and food storage in 13,000 y-old stone mortars at Raqefet Cave, Israel: Investigating Natufian ritual feasting. J. Archaeol. Sci. Rep. 2018, 21, 783–793. [Google Scholar] [CrossRef]
- Monteiro Vieira, I.M.; Passos Santos, B.L.; Santos Ruzene, D.; Brányik, T.; Teixeira, J.A.; de Almeida, E.; Silva, J.B.; Pereira Silva, D. Alcohol and health: Standards of consumption, benefits and harm—A review. Czech J. Food Sci. 2018, 36, 427–440. [Google Scholar] [CrossRef]
- Gschaedler, A. Contribution of non-conventional yeasts in alcoholic beverages. Curr. Opin. Food Sci. 2017, 13, 73–77. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Kabak, B.; Dobson, A.D.W. An introduction to the traditional fermented foods and beverages of Turkey. Crit. Rev. Food Sci. Nutr. 2011, 51, 248–260. [Google Scholar] [CrossRef]
- Lopes, A.C.A.; Eda, S.H.; Andrade, R.P.; Amorim, J.C.; Duarte, W.F. New alcoholic fermented beverages-potentials and challenges. In Fermented Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Elsevier: New York, NY, USA, 2019; pp. 577–603. [Google Scholar]
- Wardencki, W. Alcoholic Beverages. In Encyclopedia of Analytical Science; Worsfold, P., Poole, C., Townshend, A., Miró, M., Eds.; Elsevier Inc.: London, UK, 2019; pp. 67–76. [Google Scholar]
- Dasgupta, A.; Klein, K. Alcoholic Beverages: Antioxidant and Other Health Benefits of Moderate Consumption. In Antioxidants in Food, Vitamins and Supplements; Dasgupta, A., Klein, K., Eds.; Elsevier: New York, NY, USA, 2014; pp. 259–276. [Google Scholar]
- Walker, G.; Stewart, G. Saccharomyces cerevisiae in the Production of Fermented Beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Foster, R.K.; Marriott, H.E. Alcohol consumption in the new millennium—Weighing up the risks and benefits for our health. Br. Nutr. Found. 2006, 31, 286–331. [Google Scholar] [CrossRef]
- Day, L.; McSweeney, P.L.H. Nanoengineering in the Beverage Industry. Ref. Modul. Food Sci. 2016, 20, 1–5. [Google Scholar]
- Rodhouse, L.; Carbonero, F. Overview of craft brewing specificities and potentially associated microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 462–473. [Google Scholar] [CrossRef]
- Ryan, R. Safety of Food and Beverages: Alcoholic Beverages. Encycl. Food Saf. 2013, 3, 364–370. [Google Scholar]
- World Health Organization (WHO). Global Status Report on Alcohol 2004. Available online: https://www.drugsandalcohol.ie/6373/1/4183_global_status_report_2004_overview.pdf (accessed on 6 September 2006).
- De Keukeleire, D. Fundamentals Of Beer And Hop Chemistry. Quim. Nova 2000, 23, 108–112. [Google Scholar] [CrossRef]
- Preedy, V.R. Beer in Health and Disease Prevention; Academic Press: Cambridge, UK, 2008. [Google Scholar]
- Cortacero-Ramírez, S.; De Castro, H.B.M.; Segura-Carretero, A.; Cruces-Blanco, C.; Fernández-Gutiérrez, A. Analysis of beer components by capillary electrophoretic methods. Trends Anal. Chem. 2003, 22, 440–445. [Google Scholar] [CrossRef]
- Tamang, J.P.; Kailasapathy, K. Fermented Foods and Beverages of the World, 1st ed.; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781420094954. [Google Scholar]
- Willaert, R. Biochemistry of Beer Fermentation. In Food Biochemistry and Food Processing: Second Edition; Simpson, B.K., Nollet, L.M.L., Toldrá, F., Benjakul, S., Paliyath, G., Hui, Y.H., Eds.; John Wiley & Sons, Inc.: Ames, IA, USA, 2012; pp. 627–653. [Google Scholar] [CrossRef]
- Harrison, M.A.; Albanese, J.B. Beer/Brewing. In Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–11. ISBN 9780128117378. [Google Scholar]
- Buglass, A.J. Handbook of Alcoholic Beverages: Technical, Analytical and Nutritional Aspects; Wiley: Hoboken, NJ, USA, 2010; Volume 1–2, ISBN 9780470512029. [Google Scholar]
- Bettenhausen, H.M.; Barr, L.; Broeckling, C.D.; Chaparro, J.M.; Holbrook, C.; Sedin, D.; Heuberger, A.L. Influence of malt source on beer chemistry, flavor, and flavor stability. Food Res. Int. 2018, 113, 487–504. [Google Scholar] [CrossRef]
- Mathias, T.; Mello, P.; Sérvulo, E. Nitrogen compounds in brewing wort and beer: A review. J. Brew. Distill. 2014, 5, 10–17. [Google Scholar]
- Cozzolino, D.; Dambergs, R. Wine and Beer. In Infrared Spectroscopy for Food Quality Analysis and Control; Sun, D.-W., Ed.; Elsevier Academic Press: San Diego, CA, USA, 2009; pp. 377–397. [Google Scholar]
- Li, Q.; Wang, J.; Liu, C. Beers. In Current Developments in Biotechnology and Bioengineering: Food and Beverages Industry; Pandey, A., Du, G., Sanromán, M., Soccol, C.R., Dussap, C.-G., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 305–351. ISBN 9780444636775. [Google Scholar]
- Bokulich, N.A.; Bamforth, C.W. The Microbiology of Malting and Brewing. Microbiol. Mol. Biol. Rev. 2013, 77, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Anderson, H.E.; Santos, I.C.; Hildenbrand, Z.L.; Schug, K.A. A review of the analytical methods used for beer ingredient and finished product analysis and quality control. Anal. Chim. Acta 2019, 1085, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Macwilliam, I.C. Wort composition—A Review. J. Inst. Brew. 1968, 74, 38–54. [Google Scholar] [CrossRef]
- Keenan, R.T.; Krasnokutsky, S.; Pillinger, M.H. Etiology and Pathogenesis of Hyperuricemia and Gout, 10th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Yamamoto, T.; Moriwaki, Y. Purines in beer. In Beer in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: Oxford, UK, 2009; pp. 285–290. [Google Scholar]
- Fukuuchi, T.; Yasuda, M.; Inazawa, K.; Ota, T.; Yamaoka, N.; Mawatari, K.; Nakagomi, K.; Kaneko, K. A Simple HPLC Method for Determining the Purine Content of Beer and Beer-like Alcoholic Beverages. Anal. Sci. 2013, 29, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Moriwaki, Y.; Takahashi, S.; Tsutsumi, Z.; Ka, T.; Fukuchi, M.; Hada, T. Effect of beer on the plasma concentrations of uridine and purine bases. Metabolism 2002, 51, 1317–1323. [Google Scholar] [CrossRef]
- Cameron, J.S.; Moro, F.; Simmonds, A. Gout, uric acid and purine metabolsim in paediatric nephrology. Pediatr. Nephrol. 1993, 7, 105–118. [Google Scholar] [CrossRef]
- Haskó, G.; Linden, J.; Cronstein, B.; Pacher, P. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N. Fundamentals of Biomedical Science-Clinical Biochemistry, 1st ed.; Ahmed, N., Ed.; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; Macmillan Learning: New York, NY, USA, 2017. [Google Scholar]
- Li, H.; Liu, F.; Hao, J.; Liu, C. Determination of purines in beer by HPLC using a simple and rapid sample pretreatment. J. Am. Soc. Brew. Chem. 2015, 73, 137–142. [Google Scholar] [CrossRef]
- Kaneko, K.; Aoyagi, Y.; Fukuuchi, T.; Inazawa, K.; Yamaoka, N. Total Purine and Purine Base Content of Common Foodstuffs for Facilitating Nutritional Therapy for Gout and Hyperuricemia. Biol. Pharm. Bull. 2014, 37, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Sayuti, K.; Yenrina, R.; Refdi, C.W.; Fajri, P.Y. Research Article Adenine, Guanine, Xanthine and Hypoxanthine Content in Various Indonesian Foods. Pak. J. Nutr. 2019, 18, 260–263. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Roseland, J.M.; Haytowitz, D.B.; Pehrsson, P.R.; Ershow, A.G. Availability and quality of published data on the purine content of foods, alcoholic beverages, and dietary supplements. J. Food Compos. Anal. 2019, 84, 103281. [Google Scholar] [CrossRef]
- Ragab, G.; Elshahaly, M.; Bardin, T. Gout: An old disease in new perspective—A review. J. Adv. Res. 2017, 8, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Suwa, Y. Purine Metabolism and the Influence of Alcoholic Beverages. In Beer in Health and Disease Prevention; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; Volume 2, pp. 1069–1081. [Google Scholar]
- Dale, C.J.; Lyddiatt, A. Quantitative analysis of purine nucleosides and free bases in wort and beer. J. Inst. Brw. 1994, 100, 173–178. [Google Scholar] [CrossRef]
- Klein, H.; Leubolt, R. Ion-exchange high-performance the brewing industry liquid chromatography in the brewing industry. J. Chromatogr. 1993, 640, 259–270. [Google Scholar] [CrossRef]
- Kaneko, K.; Yamanobe, T.; Fujimori, S. Detersamination of purine contents of alcoholic beverages using high performance liquid chromatography. Biomed. Chromatogr. 2009, 23, 858–864. [Google Scholar] [CrossRef]
- Klampfl, C.W.; Himmelsbach, M.; Buchberger, W.; Klein, H. Determination of purines and pyrimidines in beer samples by capillary zone electrophoresis. Anal. Chim. Acta 2002, 454, 185–191. [Google Scholar] [CrossRef]
- Cortacero-Ramírez, S.; Segura-Carretero, A.; Cruces-Blanco, C.; Hernáinz-Bermúdez de Castro, M.; Fernández-Gutiérrez, A. Direct multicomponent analysis of beer samples constituents using micellar electrokinetic capillary chromatography. Electrophoresis 2004, 25, 1867–1871. [Google Scholar] [CrossRef] [PubMed]
- Harris, G. Brewing Industry Research Foundation and Brewing. J. Inst. Brew. 1958, 64, 290–303. [Google Scholar] [CrossRef]
- Gibson, T.; Rodgers, A.V.; Simmonds, H.A.; Toseland, P. Beer drinking and its effect on uric acid. Rheumatology 1984, 23, 203–209. [Google Scholar] [CrossRef]
- Cortacero-Ramírez, S.; Segura-Carretero, A.; Cruces-Blanco, C.; Romero-Romero, M.L.; Fernández-Gutiérrez, A. Simultaneous determination of multiple constituents in real beer samples of different origins by capillary zone electrophoresis. Anal. Bioanal. Chem. 2004, 380, 831–837. [Google Scholar] [CrossRef]
- Markelj, J.; Zupančić, T.; Pihlar, B. Optimization of high performance liquid chromatography method for simultaneous determination of some purine and pyrimidine bases. Acta Chim. Slov. 2016, 63, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiuolo, J.; Oppedisano, F.; Gratteri, S.; Muscoli, C.; Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 2016, 213, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Grassi, D.; Ferri, L.; Desideri, G.; Di Giosia, P.; Cheli, P.; Del Pinto, R.; Ferri, C. Chronic Hyperuricemia, Uric Acid Deposit and Cardiovascular Risk. Curr. Pharm. Des. 2013, 19, 2432–2438. [Google Scholar] [CrossRef] [Green Version]
- Stompór, T. Gout, hyperuricemia and chronic kidney disease: New treatment possibilities. Pol. Ann. Med. 2016, 23, 195–201. [Google Scholar]
- De Oliveira, E.P.; Burini, R.C. High plasma uric acid concentration: Causes and consequences. Diabetol. Metab. Syndr. 2012, 4, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gliozzi, M.; Malara, N.; Muscoli, S.; Mollace, V. The treatment of hyperuricemia. Int. J. Cardiol. 2016, 213, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Yang, F.; Yang, I.; Yin, Y.; Luo, J.J.; Wang, H.; Yang, X.-F. Uric Acid, Hyperuricemia and Vascular Diseases. Front Biosci. 2012, 17, 656–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafez, R.M.; Abdel-rahman, T.M.; Naguib, R.M. A review on uric acid in plants and microorganisms: Biological applications and genetics. J. Adv. Res. 2017, 8, 475–486. [Google Scholar] [CrossRef]
- Qiu, L.; Cheng, X.; Wu, J.; Liu, J.; Xu, T.; Ding, H.; Liu, Y.; Ge, Z. Prevalence of hyperuricemia and its related risk factors in healthy adults from Northern and Northeastern Chinese provinces. BMC Public Health 2013, 13, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Pandya, B.J.; Choi, H.K. Prevalence of Gout and Hyperuricemia in the US General Population The National Health and Nutrition Examination Survey 2007–2008. Arthritis Rheum. 2011, 63, 3136–3141. [Google Scholar] [CrossRef]
- Trifi, G.; Morabito, P.; Cavagna, L.; Ferrajolo, C.; Pecchioli, S.; Simonetti, M.; Bianchini, E.; Medea, G.; Cricelli, C.; Caputi, A.P.; et al. Epidemiology of gout and hyperuricaemia in Italy during the years 2005–2009: A nationwide population-based study. Ann. Rheum. Dis. 2013, 72, 694–700. [Google Scholar] [CrossRef]
- Krishnan, E. Reduced Glomerular Function and Prevalence of Gout: NHANES 2009–10. PLoS ONE 2012, 7, e50046. [Google Scholar] [CrossRef] [Green Version]
- Miao, Z.; Li, C.; Chen, Y.; Zhao, S.; Wang, Y.; Wang, Z.; Chen, X.; Xu, F.; Wang, F.; Sun, R.; et al. Dietary and lifestyle changes associated with high prevalence of hyperuricemia and gout in the Shandong coastal cities of Eastern China. J. Rheumatol. 2008, 35, 1859–1864. [Google Scholar]
- Mahor, D.; Prasad, G.S. Biochemical characterization of Kluyveromyces lactis adenine deaminase and guanine deaminase and their potential application in lowering purine content in beer. Front. Bioeng. Biotechnol. 2018, 6, 180. [Google Scholar] [CrossRef]
- Lv, Q.; Meng, X.; He, F.; Chen, S.; Su, H.; Xiong, J.; Gao, P.; Tian, J.; Liu, J.; Zhu, Z.; et al. High Serum Uric Acid and Increased Risk of Type 2 Diabetes: A Systemic Review and Meta-Analysis of Prospective Cohort Studies. PLoS ONE 2013, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Que, S.; Zhou, L.; Zheng, S. Dose-response Relationship of Serum Uric Acid with Metabolic Syndrome and Non-alcoholic Fatty Liver Disease Incidence: A Meta- analysis of Prospective Studies. Sci. Rep. 2015, 5, 14325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Qin, T.; Chen, J.; Li, Y.; Wang, L.; Huang, H. Hyperuricemia and Risk of Incident Hypertension: A Systematic Review and Meta-Analysis of Observational Studies. PLoS ONE 2014, 9, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, J.G.; Juzwishin, K.D.M.; Eiriksdottir, G.; Gudnason, V.; Danesh, J. Serum uric acid and coronary heart disease in 9458 incident cases and 155,084 controls: Prospective study and meta-analysis. PLoS Med. 2005, 2, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Huang, B.; Li, Y.; Huang, Y.; Li, J.; Yao, H.; Jing, X.; Chen, J.; Wang, J. Uric acid and risk of heart failure: A systematic review and meta-analysis. Eur. J. Heart Fail. 2014, 16, 15–24. [Google Scholar] [CrossRef]
- Li, M.; Hou, W.; Zhang, X.; Hu, L.; Tang, Z. Hyperuricemia and risk of stroke: A systematic review and meta-analysis of prospective studies. Atherosclerosis 2014, 232, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Thangaratinam, S.; Sharp, S.; Coomarasamy, A. Accuracy of serum uric acid in predicting complications of pre-eclampsia: A systematic review. Int. J. Obstet. Gynaecol. 2006, 113, 369–378. [Google Scholar] [CrossRef]
- Campion, E.W.; Glynn, R.J.; Sc, D.; Delabry, L. Asymptomatic Hyperuricemia Risks and Consequences in the Normative Aging Study. Am. J. Med. 1987, 82, 421–426. [Google Scholar] [CrossRef]
- Kuo, C.F.; Grainge, M.J.; Zhang, W.; Doherty, M. Global epidemiology of gout: Prevalence, incidence and risk factors. Nat. Rev. Rheumatol. 2015, 11, 649–662. [Google Scholar] [CrossRef]
- Dehlin, M.; Jacobsson, L.; Roddy, E. Global epidemiology of gout: Prevalence, incidence, treatment patterns and risk factors. Nat. Rev. Rheumatol. 2020, 16, 380–390. [Google Scholar] [CrossRef]
- Mai, S.; Zobbe, K.; Erik, L.; Christensen, R. Nutritional recommendations for gout: An update from clinical epidemiology. Autoimmun. Rev. 2018, 17, 1090–1096. [Google Scholar]
- Langford, H.G.; Blaufox, M.D.; Borhani, N.O.; Curb, D.; Molteni, A.; Schneider, K.A.; Pressel, S. Is Thiazide-Produced Uric Acid Elevation Harmful ? Analysis of Data From the Hypertension Detection and Follow-up Program. Arch. Intern. Med. 1987, 147, 645–649. [Google Scholar] [CrossRef]
- Paul, B.J.; Anoopkumar, K.; Krishnan, V. Asymptomatic hyperuricemia: Is it time to intervene ? Clin. Rheumatol. 2017, 36, 2637–2644. [Google Scholar] [CrossRef]
- Chen-xu, M.; Yokose, C.; Rai, S.K.; Pillinger, M.H.; Choi, H.K. Contemporary Prevalence of Gout and Hyperuricemia in the United States and Decadal Trends: The National Health and Nutrition Examination Survey, 2007–2016. Arthritis Rheumatol. 2019, 71, 991–999. [Google Scholar] [CrossRef]
- Kim, J.; Kwak, S.G.; Lee, H.; Kim, S.; Choe, J.; Park, S. Prevalence and incidence of gout in Korea: Data from the national health claims database 2007–2015. Rheumatol. Int. 2017, 37, 1499–1506. [Google Scholar] [CrossRef]
- González-chica, D.A.; Vanlint, S.; Hoon, E.; Stocks, N. Epidemiology of arthritis, chronic back pain, gout, osteoporosis, spondyloarthropathies and rheumatoid arthritis among 1.5 million patients in Australian general practice: NPS MedicineWise MedicineInsight dataset. BMC Musculoskelet. Disord. 2018, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.; Grainge, M.J.; Mallen, C.; Zhang, W.; Doherty, M. Rising burden of gout in the UK but continuing suboptimal management: A nationwide population study. Ann. Rheum. Dis. 2015, 74, 661–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branco, J.C.; Rodrigues, A.M.; Gouveia, N.; Eusébio, M.; Ramiro, S.; Machado, P.M.; Pereira, L.; Mourão, A.F.; Silva, I.; Laires, P. Prevalence of rheumatic and musculoskeletal diseases and their impact on health-related quality of life, physical function and mental health in Portugal: Results from EpiReumaPt–a national health survey. RMD Open 2016, 2, e000166. [Google Scholar] [CrossRef] [Green Version]
- Riches, P.L.; Wright, A.F.; Ralston, S.H. Recent insights into the pathogenesis of hyperuricaemia and gout. Hum. Mol. Genet. 2009, 18, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Roddy, E.; Zhang, W.; Doherty, M. The changing epidemiology of gout. Nat. Clin. Pract. Rheumatol. 2007, 3, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Akizuki, S. Serum uric acid levels among thirty-four thousand people in Japan. Ann. Rheum. Dis. 1982, 41, 272–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, T.C.; Keenan, R.T. Unusual clinical presentations of gout. Curr. Opin. Rheumatol. 2010, 22, 181–187. [Google Scholar] [CrossRef]
- Singh, J.A.; Gaffo, A. Gout epidemiology and comorbidities. Semin. Arthritis Rheum. 2020, 50, 11–16. [Google Scholar] [CrossRef]
- Curhan, G.; Sc, D. Purine-Rich Foods, Dairy and Protein Intake, and the Risk of Gout in Men. N. Engl. J. Med. 2004, 350, 1093–1103. [Google Scholar]
- Nishioka, K.; Sumida, T.; Iwatani, M.; Kusumoto, A.; Ishikura, Y.; Hatanaka, H.; Yomo, H.; Kohda, H.; Ashikari, T.; Shibano, Y.; et al. Influence of Moderate Drinking on Purine and Carbohydrate Metabolism. Alcohol. Clin. Exp. Res. 2002, 26, 20s–25s. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Sakurai, M.; Miura, K.; Morikawa, Y.; Yoshita, K.; Ishizaki, M.; Kido, T.; Naruse, Y.; Suwazono, Y.; Nakagawa, H. Alcohol intake and the risk of hyperuricaemia: A 6-year prospective study in Japanese men. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 989–996. [Google Scholar] [CrossRef]
- Nakanishi, N.; Tatara, K.; Nakamura, K.; Suzuki, K. Risk factors for the incidence of hyperuricaemia: A 6-year longitudinal study of middle-aged Japanese men. Int. J. Epidemiol. 1999, 28, 888–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of microbial enzymes in food industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Ozatay, S. Recent Applications of Enzymes in Food Industry. J. Curr. Res. Eng. Sci. Technol. 2020, 6, 17–30. [Google Scholar] [CrossRef]
- Jankowska, D.A.; Trautwein-Schult, A.; Cordes, A.; Bode, R.; Baronian, K.; Kunze, G. A novel enzymatic approach in the production of food with low purine content using Arxula adeninivorans endogenous and recombinant purine degradative enzymes. Bioengineered 2015, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Shibano, Y.; Yomo, H.; Matsumoto, T.; Koda, H.; Suwa, Y.; Amachi, T.; Hatanaka, H.; Shimizu, S. Process for Manufacturing Beer. U.S. Patent 6,013,288, 11 January 2000. [Google Scholar]
- Chen, J.; Wen, C.; City, H.; Sheu, F.; Tsai, M. Method of Reducing the Purine Content of an Edible Material. U.S. Patent 8,460,724, 21 April 2011. [Google Scholar]
- Fujino, S.; Sakuma, S. Process for Producing Fermented Malt Drink with Reduced Purine Content. EU Patent Application No. EP1437398B1, 1 September 2010. [Google Scholar]
- Mahor, D.; Priyanka, A.; Prasad, G.S.; Thakur, K.G. Functional and structural characterization of purine nucleoside phosphorylase from Kluyveromyces lactis and its potential applications in reducing purine content in food. PLoS ONE 2016, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, D.A.; Faulwasser, K.; Trautwein-Schult, A.; Cordes, A.; Hoferichter, P.; Klein, C.; Bode, R.; Baronian, K.; Kunze, G. Arxula adeninivorans recombinant adenine deaminase and its application in the production of food with low purine content. J. Appl. Microbiol. 2013, 115, 1134–1146. [Google Scholar] [CrossRef]
- Jankowska, D.A.; Trautwein-Schult, A.; Cordes, A.; Hoferichter, P.; Klein, C.; Bode, R.; Baronian, K.; Kunze, G. Arxula adeninivorans xanthine oxidoreductase and its application in the production of food with low purine content. J. Appl. Microbiol. 2013, 115, 796–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Ren, L.; Wang, D. Effect of allicin and its mechanism of action in purine removal in turbot. J. Food Sci. 2020, 85, 3562–3569. [Google Scholar] [CrossRef]
- Li, T.; Ren, L.; Wang, D.; Song, M.; Li, Q.; Li, J. Optimization of extraction conditions and determination of purine content in marine fish during boiling. PeerJ 2019, 2019, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Attila, Z.; Buday; Belleau, G. The Use of Activated Carbon for the Recovery and Concentration of Nucleic Acid Derivatives and other Compounds from Biological Samples. Can. Inst. Food Sci. Technol. J. 1974, 7, 188–198. [Google Scholar] [CrossRef]
- Shibata, J.; Murayama, N.; Tateyama, M. Adsorption of Purine Compounds in Beer with Activated Carbon Prepared from Beer Lees. Resour. Process. 2009, 56, 120–126. [Google Scholar] [CrossRef]
- Marahel, F.; Ghaedi, M.; Ansari, A. Zinc oxide nanoparticles loaded on activated carbon and its application for adsorption removal of uric acid. Synth. React. Inorg. Met. Nano-Met. Chem. 2015, 45, 1387–1395. [Google Scholar] [CrossRef]
- Leshchinskaya, A.P.; Ezhova, N.M.; Pisarev, O.A. Synthesis and characterization of 2-hydroxyethyl methacrylate-ethylene glycol dimethacrylate polymeric granules intended for selective removal of uric acid. React. Funct. Polym. 2016, 102, 101–109. [Google Scholar] [CrossRef]
- Gao, B.; Jiang, P.; Lei, H. Studies on adsorption property of novel composite adsorption material PEI/SiO2 for uric acid. Mater. Lett. 2006, 60, 3398–3404. [Google Scholar] [CrossRef]
- Liu, C.; Liang, X.; Liu, X.; Wang, Q.; Zhan, L.; Zhang, R.; Qiao, W.; Ling, L. Surface modification of pitch-based spherical activated carbon by CVD of NH 3 to improve its adsorption to uric acid. Appl. Surf. Sci. 2008, 254, 6701–6705. [Google Scholar] [CrossRef]
- Sweetman, L.; Nyhan, W.L. Studies on the mechanism of adsorption of purines in Sephadex G-10 chromatography. J. Chromatogr. A 1971, 59, 349–366. [Google Scholar] [CrossRef]
- Pedreira-Segade, U.; Feuillie, C.; Pelletier, M.; Michot, L.J.; Daniel, I. Adsorption of nucleotides onto ferromagnesian phyllosilicates: Significance for the origin of life. Geochim. Cosmochim. Acta 2016, 176, 81–95. [Google Scholar] [CrossRef]
- Tsurushima, M.; Kokubu, G.; Moriya, K. Method of Separation of Nucleotides and Nucleosides from Each Other. U.S. Patent 4,577,012, 18 March 1986. [Google Scholar]
- Icenhour, C.R.; Loyal, B.V.; Nguyen, L.N.K. Nucleic Acid Extraction From Complex Matrices. U.S. Patent 8,574,890, 11 November 2013. [Google Scholar]
- Erol, O.; Uyan, I.; Hatip, M.; Yilmaz, C.; Tekinay, A.B.; Guler, M.O. Recent advances in bioactive 1D and 2D carbon nanomaterials for biomedical applications. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 2433–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Dong, S. Graphene nanosheet: Synthesis, molecular engineering, thin film, hybrids, and energy and analytical applications. Chem. Soc. Rev. 2011, 40, 2644–2672. [Google Scholar] [CrossRef] [PubMed]
- Nandy, B.; Santosh, M.; Maiti, P.K. Interaction of nucleic acids with carbon nanotubes and dendrimers. J. Biosci. 2012, 37, 457–474. [Google Scholar] [CrossRef] [Green Version]
- Shtogun, Y.V.; Woods, L.M.; Dovbeshko, G.I. Adsorption of adenine and thymine and their radicals on single-wall carbon nanotubes. J. Phys. Chem. C 2007, 111, 18174–18181. [Google Scholar] [CrossRef]
- Novoselov, K.S.; Geim, A.K.; Morozov, S.V.; Jiang, D. Electric Field Effect in Atomically Thin Carbon Films. Science 2004, 306, 666–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Wang, Y.; Zhai, G. Biomedical applications of the graphene-based materials. Mater. Sci. Eng. C 2016, 61, 953–964. [Google Scholar] [CrossRef]
- Perreault, F.; Fonseca De Faria, A.; Elimelech, M. Environmental applications of graphene-based nanomaterials. Chem. Soc. Rev. 2015, 44, 5861–5896. [Google Scholar] [CrossRef]
- Pumera, M. Electrochemistry of Graphene: New Horizons for Sensing and Energy Storage. Chem. Rec. 2009, 9, 211–223. [Google Scholar] [CrossRef]
- Yang, Y.; Asiri, A.M.; Tang, Z.; Du, D.; Lin, Y. Graphene based materials for biomedical applications. Mater. Today 2013, 16, 365–373. [Google Scholar] [CrossRef]
- Zhu, Y.; Murali, S.; Cai, W.; Li, X.; Suk, J.W.; Potts, J.R.; Ruoff, R.S. Graphene and graphene oxide: Synthesis, properties, and applications. Adv. Mater. 2010, 22, 3906–3924. [Google Scholar] [CrossRef]
- Antony, J.; Grimme, S. Structures and interaction energies of stacked graphene–nucleobase complexes. Phys. Chem. Chem. Phys. 2008, 10, 2722–2729. [Google Scholar] [CrossRef] [Green Version]
- Sowerby, S.J.; Cohn, C.A.; Heckl, W.M.; Holm, N.G. Differential adsorption of nucleic acid bases: Relevance to the origin of life. Proc. Natl. Acad. Sci. USA 2001, 98, 820–822. [Google Scholar] [CrossRef] [Green Version]
- Gowtham, S.; Scheicher, R.H.; Ahuja, R.; Pandey, R.; Karna, S.P. Physisorption of nucleobases on graphene: Density-functional calculations. Phys. Rev. B Condens. Matter Mater. Phys. 2007, 76, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Varghese, N.; Mogera, U.; Govindaraj, A.; Das, A.; Maiti, P.K.; Sood, A.K.; Rao, C.N.R. Binding of DNA nucleobases and nucleosides with graphene. ChemPhysChem 2009, 10, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Adsorption of DNA onto gold nanoparticles and graphene oxide: Surface science and applications. Phys. Chem. Chem. Phys. 2012, 14, 10485–10496. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Choi, Y.K.; Kim, H.J.; Scheicher, R.H.; Cho, J.H. Physisorption of DNA nucleobases on h -BN and graphene: VdW-corrected DFT calculations. J. Phys. Chem. C 2013, 117, 13435–13441. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | Source | |
|---|---|---|
| Water | 90% | - |
| Ethanol | 20,000–80,000 mg L−1 | Yeast, malt |
| Carbon Dioxide | 3500–4500 mg L−1 | Yeast, malt |
| Carbohydrates | 3.3–4.4% | Malt |
| Inorganic salts | 500–2000 mg L−1 | Water, malt |
| Total nitrogen compounds | 300–1000 mg L−1 | Yeast, malt |
| Organic acids | 200–500 mg L−1 | Yeast, malt |
| Higher alcohols | 60–100 mg L−1 | Yeast, malt |
| Aldehydes | 10–20 mg L−1 | Yeast, hops |
| Esters | 60–80 mg L−1 | Yeast, malt, hops |
| Sulphur compounds | 1–10 mg L−1 | Yeast, malt, hops |
| Hop derivatives | 20–60 mg L−1 | Hops |
| Complex B vitamins | 5–10 mg L−1 | Yeast, malt |
| Concentration (µmol L−1) * | |
|---|---|
| Guanosine | 174 ± 16.1 |
| Xanthine | 58.5 ± 3.6 |
| Guanine | 42.4 ± 3.6 |
| Adenosine | 42.1 ± 7.9 |
| Inosine | 20.3 ± 2.5 |
| Hypoxanthine | 17.5 ± 1.5 |
| Adenine | 17.2 ± 2.3 |
| Alcoholic Beverage | Total Purine Content (µmol L−1) * |
|---|---|
| Beer | 13.3–1145.7 |
| Regular | 225.0–580.2 |
| Low-malt | 193.4–267.9 |
| Low-malt and low-purine | 13.3 |
| Local | 312.0–1145.7 |
| Low-alcohol | 194.8–898.3 |
| Spirits | 0.7–26.4 |
| Other liquors | 13.1–818.3 |
| Sake | 82.1–110.4 |
| Shoku-shu | 537.4–818.3 |
| Ume liqueur | 13.1 |
| Beer-flavored beverage | 73.5–157.4 |
| Country | Methodology | Year | Prevalence |
|---|---|---|---|
| USA | Third National Health and Nutrition Examination Survey (NHANES-III) 5467 participants/adults; gout definition: self-reported gout | 2007–2016 | 3.9% (9.2 million); men: 5.2% (5.9 million); women: 2.7% (3.3 million) |
| South Korea | National Health Claims Database for specialized care; all ages; gout definition: physician-diagnosed | 2007–2015 | 2007: 3.49/1000 individuals 2015: 7.58/1000 individuals 2025 perspective: increment of 1.66% |
| Australia | National Database of medical records; ≥18 years; Gout definition: diagnosis in medical records | 2013–2016 | 1.6% |
| UK | Clinical Practice Research Datalink | 1997–2012 | 2.49%; increment of 63.9% since 1997 |
| Portugal | Randomly selected 10,661 adult participants; ≥18 years; Gout definition: ACR 1977 criteria | 2011–2013 | 1.3%; men: 2.6%; women: 0.96% |
| Target | Removal Agent | Method | Matrix | Application |
|---|---|---|---|---|
| Adenine, guanine, xanthine, adenosine, guanosine and inosine | Nucleoside phosphorylase isolated from calf spleen | Enzymatic degradation | Wort | Brewing process for the manufacture of a beer reduced in purines |
| Inosine and hypoxanthine | Purine nucleoside phosphosphorylase from K. lactis (KlacPNP) and KlacPNP256E variant | Enzymatic degradation | Beer | Reduction of the purine content of beer |
| Adenine and guanine | Recombinant adenine and guanine deaminases of K. lactis | Enzymatic degradation | Beer | Reduction of the purine content of beer |
| Nucleic acid derivatives (e.g., guanine and guanosine) | Activated carbon | Adsorption | Wort and beer | Recover and concentration of nucleic acids |
| Adenosine, guanosine and guanine | Activated charcoal prepared with beer lees | Adsorption | Beer and low-malt beer | Production of a malt fermented beverage with reduced purine content |
| Target | Matrix | Adsorbent |
|---|---|---|
| Uric acid | Aqueous solutions | 2-hydroxyethyl methacrylate and ethyleneglycol dimethacrylate in the shape of granules |
| Uric acid | Aqueous solutions | Zinc oxide nanoparticles loaded on activated carbon |
| Uric acid | Aqueous solutions | Pitch-based spherical activated carbon (PSAC) modified by CVD of NH3 |
| Uric acid | Aqueous solutions | Polyethyleneimine/SiO2 |
| DNA and RNA | - | Activated carbon coated with polyvinylpyrrolidone, dextran or coconut flours |
| DNA nucleobases (guanine, adenine, cytosine and thymine) | - | Graphene |
| DNA nucleobases (guanine, adenine, cytosine and thymine) | - | Carbon nanotubes, dendrimers and graphene |
| DNA nucleobases (guanine, adenine, cytosine and thymine) | Aqueous solutions | Graphene |
| Adenine, adenosine and AMP | Aqueous solutions and beer | Activated carbon derived from beer lees |
| DNA nucleobases (guanine, adenine, cytosine and thymine) | - | Graphene |
| DNA nucleobases (guanine, adenine, cytosine and thymine) | - | Graphene |
| Adenine, thymine and radicals | Single-wall carbon nanotubes | |
| Adenosine, guanosine and guanine | Wort, beer and low-malt beers | Activated charcoal |
| DNA nucleobases (guanine, adenine, cytosine and thymine) | Graphite | |
| Nucleotides and nucleosides | Activated carbon | |
| Adenine and xanthine | Aqueous solutions | Dextran T40 and Sephadex G-10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, C.; Neves, M.C.; Freire, M.G. Towards the Use of Adsorption Methods for the Removal of Purines from Beer. Molecules 2021, 26, 6460. https://doi.org/10.3390/molecules26216460
Almeida C, Neves MC, Freire MG. Towards the Use of Adsorption Methods for the Removal of Purines from Beer. Molecules. 2021; 26(21):6460. https://doi.org/10.3390/molecules26216460
Chicago/Turabian StyleAlmeida, Catarina, Márcia C. Neves, and Mara G. Freire. 2021. "Towards the Use of Adsorption Methods for the Removal of Purines from Beer" Molecules 26, no. 21: 6460. https://doi.org/10.3390/molecules26216460
APA StyleAlmeida, C., Neves, M. C., & Freire, M. G. (2021). Towards the Use of Adsorption Methods for the Removal of Purines from Beer. Molecules, 26(21), 6460. https://doi.org/10.3390/molecules26216460