
Potential Vasculoprotective Effects of Blackcurrant (Ribes nigrum) Extract in Diabetic KK-Ay Mice



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
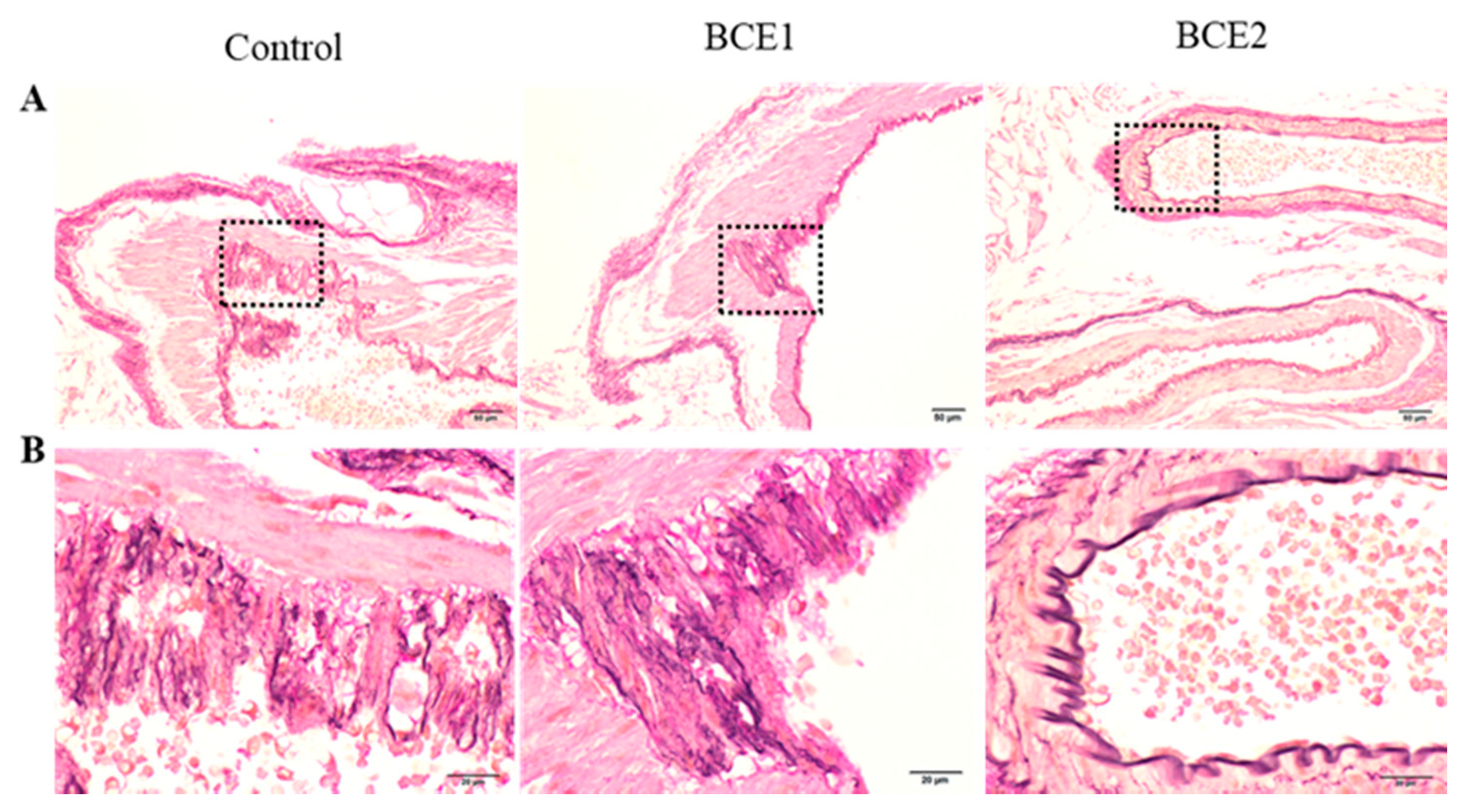
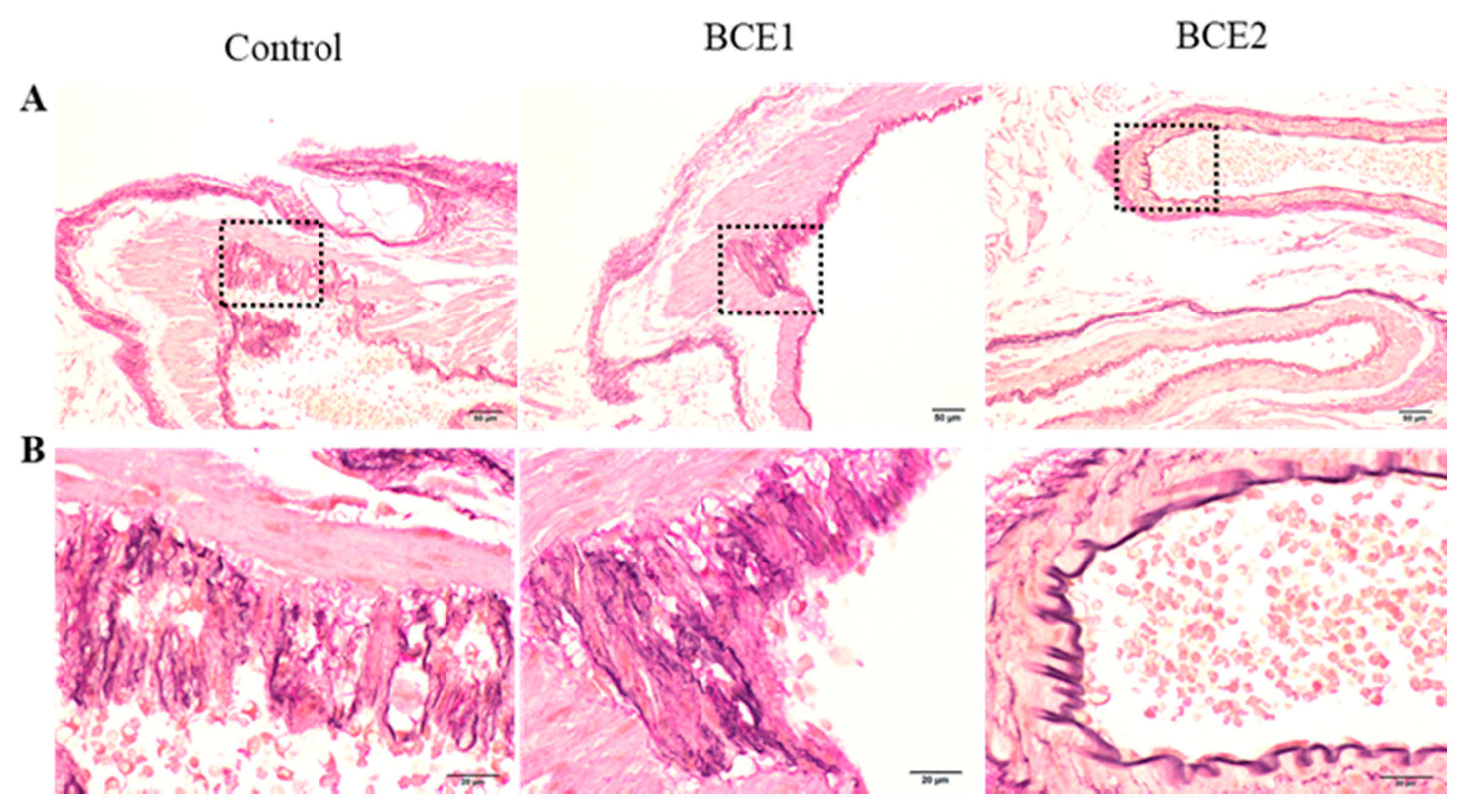
2.1. Histological Analyses of the Abdominal Aorta of BCE-Treated KK-Ay Mice
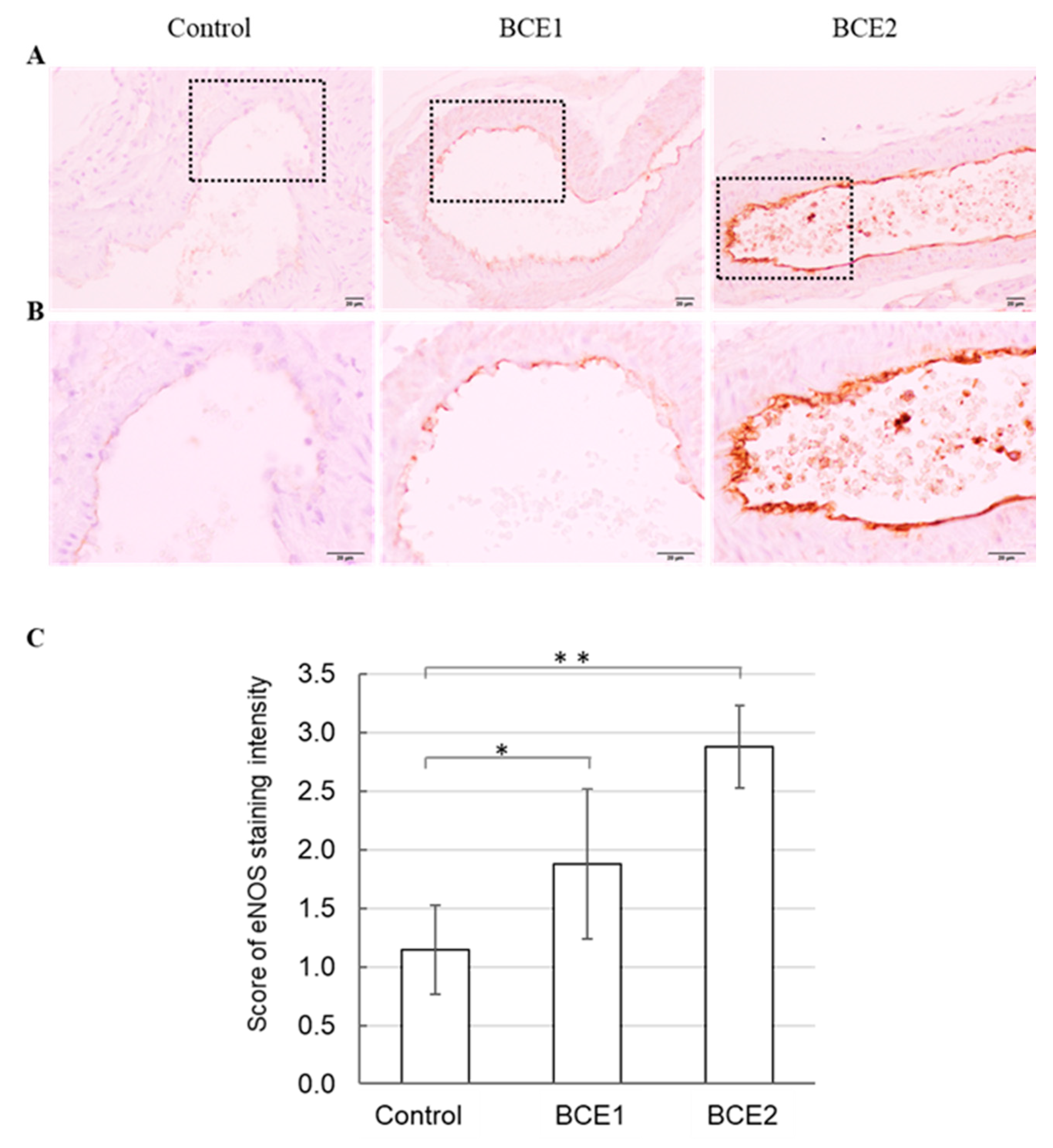
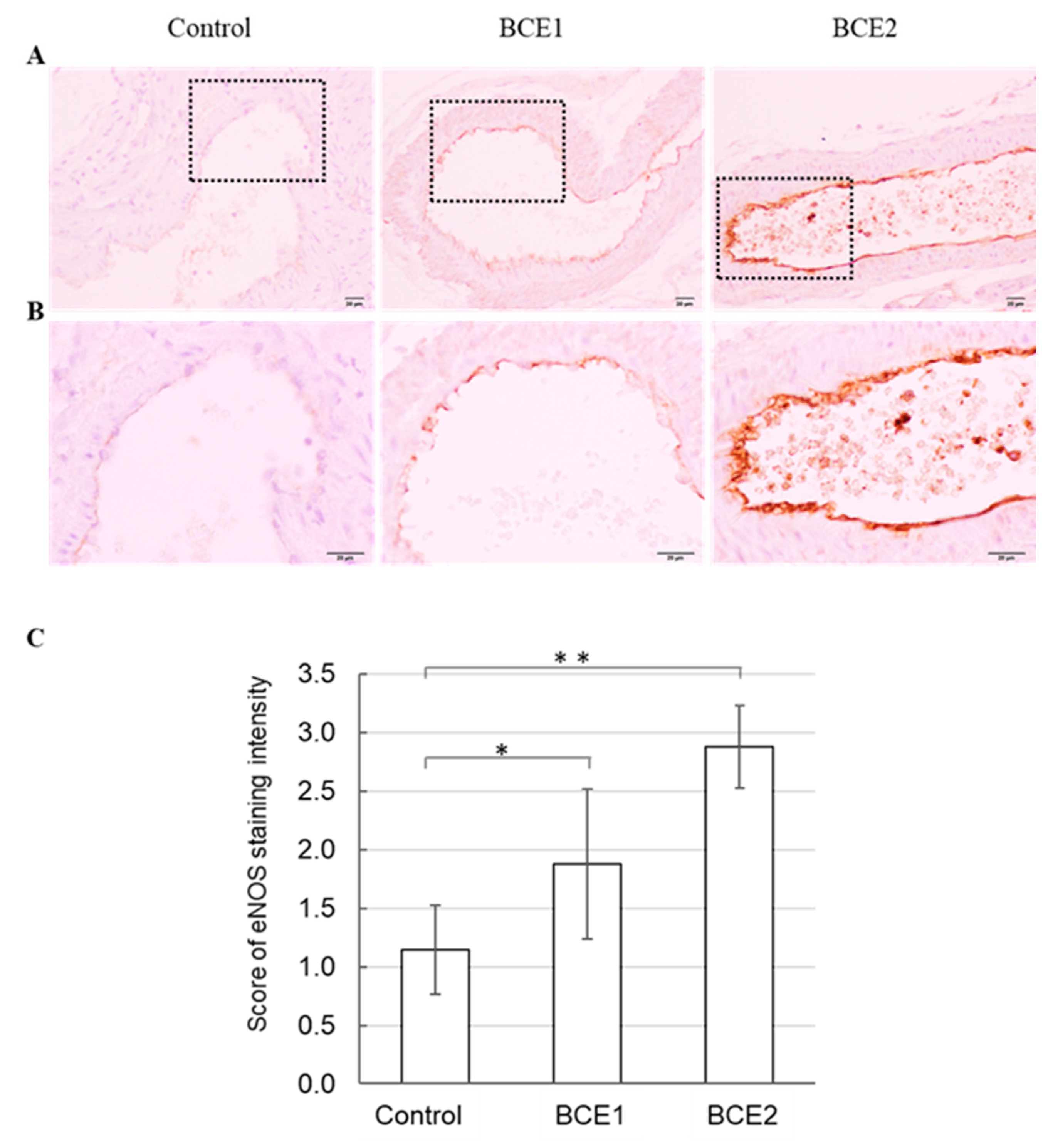
2.2. eNOS Protein Expression in BCE-Treated KK-Ay Mice
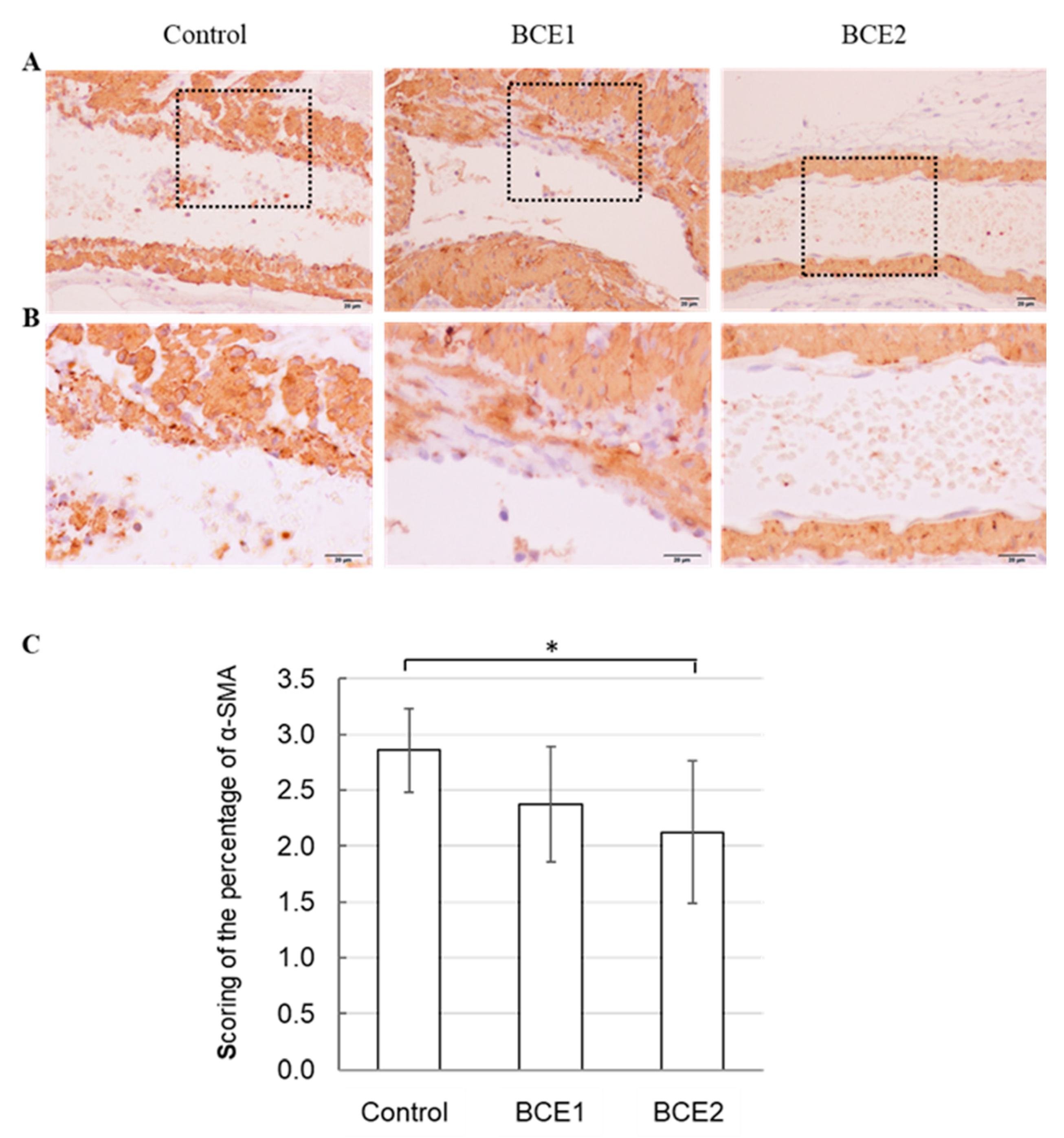
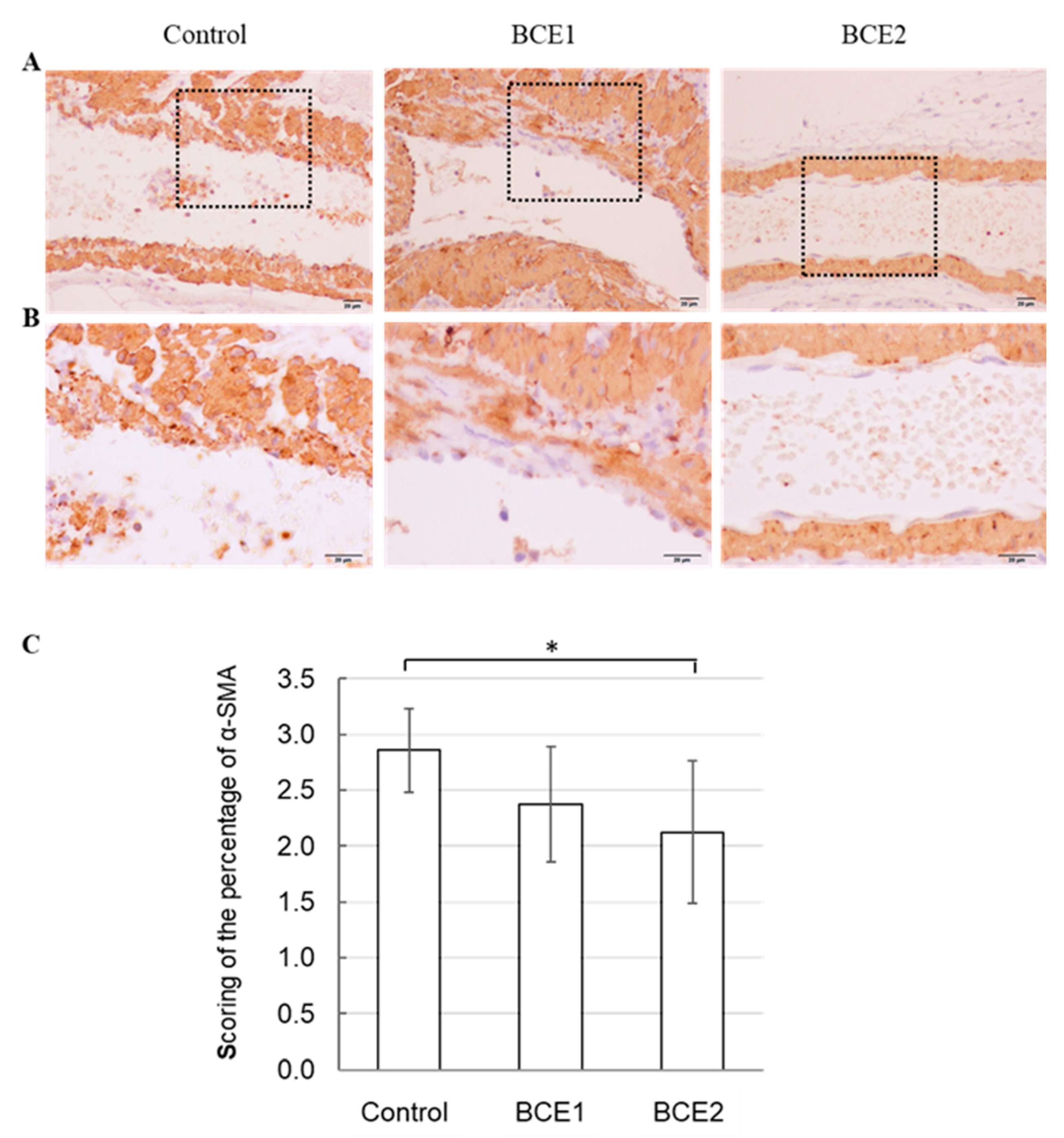
2.3. α-SMA Expression in BCE-Treated KK-Ay Mice
3. Materials and Methods
3.1. BCE and BCE-Containing Feed
3.2. Animals and Treatments
3.3. Histological Analyses
3.4. Immunohistochemical Staining of eNOS and α-SMA
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gajjar, S.; Patel, B.M. Common targets for a deadly duo of diabetes mellitus and colon cancer: Catching two fish with one worm. Eur. J. Pharmacol. 2021, 893, 173805. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Solini, A.; Zoppini, G.; Orsi, E.; Fondelli, C.; Trevisan, R.; Vedovato, M.; Cavalot, F.; Lamacchia, O.; Arosio, M.; Baroni, M.G.; et al. Resistant hypertension in patients with type 2 diabetes: Clinical correlates and association with complications. J. Hypertens. 2014, 32, 2401–2410. [Google Scholar] [CrossRef] [Green Version]
- Smith-Palmer, J.; Brändle, M.; Trevisan, R.; Federici, M.O.; Liabat, S.; Valentine, W. Assessment of the association between glycemic variability and diabetes-related complications in type 1 and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.A.; Creager, M.A.; Libby, P. Diabetes and atherosclerosis: Epidemiology, pathophysiology, and management. JAMA 2002, 287, 2570–2581. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sellers, S.; Stefanovic, N.; Leung, C.; Tan, S.M.; Huet, O.; Granville, D.J.; Cooper, M.E.; De Haan, J.B.; Bernatchez, P. Direct Endothelial Nitric Oxide Synthase Activation Provides Atheroprotection in Diabetes-Accelerated Atherosclerosis. Diabetes 2015, 64, 3937–3950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, A.V.; McDonnell, M.E. Pathogenesis of Cardiovascular Disease in Diabetes. Endocrinol. Metab. Clin. N. Am. 2018, 47, 51–63. [Google Scholar] [CrossRef]
- Katakami, N. Mechanism of Development of Atherosclerosis and Cardiovascular Disease in Diabetes Mellitus. J. Atheroscler. Thromb. 2018, 25, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Beckman, J.A.; Paneni, F.; Cosentino, F.; Creager, M.A. Diabetes and vascular disease: Pathophysiology, clinical consequences, and medical therapy: Part II. Eur. Heart J. 2013, 34, 2444–2452. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.V.; de Lemos, J.A.; Rosenson, R.S.; Ballantyne, C.M.; Liu, Y.; Mues, K.E.; Alam, S.; Elliott-Davey, M.; Bhatt, D.L.; Cannon, C.P.; et al. Use of Guideline-Recommended Risk Reduction Strategies Among Patients With Diabetes and Atherosclerotic Cardiovascular Disease. Circulation 2019, 140, 618–620. [Google Scholar] [CrossRef]
- Nasir, A.; Bullo, M.; Ahmed, Z.; Imtiaz, A.; Yaqoob, E.; Jadoon, M.; Ahmed, H.; Afreen, A.; Yaqoob, S. Nutrigenomics: Epigenetics and cancer prevention: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 60, 1375–1387. [Google Scholar] [CrossRef] [PubMed]
- Cena, H.; Calder, P.C. Defining a Healthy Diet: Evidence for the Role of Contemporary Dietary Patterns in Health and Disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behl, T.; Bungau, S.; Kumar, K.; Zengin, G.; Khan, F.; Kumar, A.; Kaur, R.; Venkatachalam, T.; Tit, D.M.; Vesa, C.M.; et al. Pleotropic Effects of Polyphenols in Cardiovascular System. Biomed. Pharmacother. 2020, 130, 110714. [Google Scholar] [CrossRef]
- Vitale, M.; Vaccaro, O.; Masulli, M.; Bonora, E.; Del Prato, S.; Giorda, C.; Nicolucci, A.; Squatrito, S.; Auciello, S.; Babini, A.C.; et al. Polyphenol intake and cardiovascular risk factors in a population with type 2 diabetes: The TOSCA.IT study. Clin. Nutr. 2017, 36, 1686–1692. [Google Scholar] [CrossRef]
- Intake of Total Polyphenols and Some Classes of Polyphenols Is Inversely Associated with Diabetes in Elderly People at High Cardiovascular Disease Risk. J. Nutr. 2015, 146, 767–777. [CrossRef] [PubMed] [Green Version]
- Tresserra-Rimbau, A.; Rimm, E.; Medina-Remón, A.; Martinez-Gonzalez, M.A.; de la Torre, R.; Corella, D.; Salas-Salvadó, J.; Gómez-Gracia, E.; Lapetra, J.; Arós, F.; et al. Inverse association between habitual polyphenol intake and incidence of cardiovascular events in the PREDIMED study. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 639–647. [Google Scholar] [CrossRef]
- Gopalan, A.; Reuben, S.C.; Ahmed, S.; Darvesh, A.S.; Hohmann, J.; Bishayee, A. The health benefits of blackcurrants. Food Funct. 2012, 3, 795–809. [Google Scholar] [CrossRef]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black Currant Anthocyanins Attenuate Weight Gain and Improve Glucose Metabolism in Diet-Induced Obese Mice with Intact, but Not Disrupted, Gut Microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef]
- Iizuka, Y.; Ozeki, A.; Tani, T.; Tsuda, T. Blackcurrant Extract Ameliorates Hyperglycemia in Type 2 Diabetic Mice in Association with Increased Basal Secretion of Glucagon-Like Peptide-1 and Activation of AMP-Activated Protein Kinase. J. Nutr. Sci. Vitaminol. 2018, 64, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Tani, T.; Nishikawa, S.; Kato, M.; Tsuda, T. Delphinidin 3-rutinoside-rich blackcurrant extract ameliorates glucose tolerance by increasing the release of glucagon-like peptide-1 secretion. Food Sci. Nutr. 2017, 5, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Lappi, J.; Raninen, K.; Väkeväinen, K.; Kårlund, A.; Törrönen, R.; Kolehmainen, M. Blackcurrant (Ribes nigrum) lowers sugar-induced postprandial glycaemia independently and in a product with fermented quinoa: A randomised crossover trial. Br. J. Nutr. 2020, 126, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Jurgoński, A.; Juskiewicz, J.; Zdunczyk, Z.; Matusevicius, P.; Kołodziejczyk, K. Polyphenol-rich extract from blackcurrant pomace attenuates the intestinal tract and serum lipid changes induced by a high-fat diet in rabbits. Eur. J. Nutr. 2014, 53, 1603–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benn, T.; Kim, B.; Park, Y.-K.; Yang, Y.; Pham, T.X.; Ku, C.S.; Farruggia, C.; Harness, E.; Smyth, J.A.; Lee, J.-Y. Polyphenol-rich blackcurrant extract exerts hypocholesterolaemic and hypoglycaemic effects in mice fed a diet containing high fat and cholesterol. Br. J. Nutr. 2015, 113, 1697–1703. [Google Scholar] [CrossRef] [Green Version]
- Willems, M.; Myers, S.D.; Gault, M.L.; Cook, M. Beneficial Physiological Effects with Blackcurrant Intake in Endurance Athletes. Int. J. Sport Nutr. Exerc. Metab. 2015, 25, 367–374. [Google Scholar] [CrossRef]
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J.; McCarthy, D.; Burton-Freeman, B.M. Effect of Black Currant Anthocyanins on the Activation of Endothelial Nitric Oxide Synthase (eNOS) in Vitro in Human Endothelial Cells. J. Agric. Food Chem. 2011, 59, 8616–8624. [Google Scholar] [CrossRef]
- Willems, M.E.T.; Parktin, N.; Widjaja, W.; Ajjimaporn, A. Effect of New Zealand Blackcurrant Extract on Physiological Responses at Rest and during Brisk Walking in Southeast Asian Men: A Randomized, Double-Blind, Placebo-Controlled, Crossover Study. Nutrients 2018, 10, 1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, K.; Nanashima, N.; Maeda, H.; Tomisawa, T.; Oey, I. Blackcurrant (Ribes nigrum L.) Extract Exerts Potential Vasculoprotective Effects in Ovariectomized Rats, Including Prevention of Elastin Degradation and Pathological Vascular Remodeling. Nutrients 2021, 13, 560. [Google Scholar] [CrossRef]
- Horie, K.; Nanashima, N.; Maeda, H. Phytoestrogenic Effects of Blackcurrant Anthocyanins Increased Endothelial Nitric Oxide Synthase (eNOS) Expression in Human Endothelial Cells and Ovariectomized Rats. Molecules 2019, 24, 1259. [Google Scholar] [CrossRef] [Green Version]
- Nanashima, N.; Horie, K.; Yamanouchi, K.; Tomisawa, T.; Kitajima, M.; Oey, I.; Maeda, H. Blackcurrant (Ribes nigrum) Extract Prevents Dyslipidemia and Hepatic Steatosis in Ovariectomized Rats. Nutrients 2020, 12, 1541. [Google Scholar] [CrossRef]
- Nanashima, N.; Horie, K.; Kitajima, M.; Takamagi, S.; Mikami, K.; In, N.; Tomisawa, T. Hypocholesterolemic Effect of Blackcurrant (Ribes nigrum) Extract in Healthy Female Subjects: A Pilot Study. Molecules 2021, 26, 4085. [Google Scholar] [CrossRef]
- Tomisawa, T.; Nanashima, N.; Kitajima, M.; Mikami, K.; Takamagi, S.; Maeda, H.; Horie, K.; Lai, F.-C.; Osanai, T. Effects of Blackcurrant Anthocyanin on Endothelial Function and Peripheral Temperature in Young Smokers. Molecules 2019, 24, 4295. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.K.; Hutchinson, J.R.; Sachidanandam, K.; Johnson, M.H.; Dorrance, A.M.; Stepp, D.W.; Fagan, S.C.; Ergul, A. Type 2 Diabetes Causes Remodeling of Cerebrovasculature via Differential Regulation of Matrix Metalloproteinases and Collagen Synthesis: Role of Endothelin-1. Diabetes 2005, 54, 2638–2644. [Google Scholar] [CrossRef] [Green Version]
- Spinetti, G.; Kraenkel, N.; Emanueli, C.; Madeddu, P. Diabetes and vessel wall remodelling: From mechanistic insights to regenerative therapies. Cardiovasc. Res. 2008, 78, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Stary, H.C.; Chandler, A.B.; Glagov, S.; Guyton, J.R.; Insull, W.; Rosenfeld, M.E.; Schaffer, S.A.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A definition of initial, fatty streak, and intermediate lesions of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 1994, 89, 2462–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Ilyas, I.; Little, P.J.; Li, H.; Kamato, D.; Zheng, X.; Luo, S.; Li, Z.; Liu, P.; Han, J.; et al. Endothelial Dysfunction in Atherosclerotic Cardiovascular Diseases and Beyond: From Mechanism to Pharmacotherapies. Pharmacol. Rev. 2021, 73, 924–967. [Google Scholar] [CrossRef]
- Daiber, A.; Xia, N.; Steven, S.; Oelze, M.; Hanf, A.; Kröller-Schön, S.; Münzel, T.; Li, H. New Therapeutic Implications of Endothelial Nitric Oxide Synthase (eNOS) Function/Dysfunction in Cardiovascular Disease. Int. J. Mol. Sci. 2019, 20, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudau, M.; Genis, A.; Lochner, A.; Strijdom, H. Endothelial dysfunction: The early predictor of atherosclerosis. Cardiovasc. J. Afr. 2012, 23, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Endemann, D.H.; Schiffrin, E.L. Endothelial dysfunction. J. Am. Soc. Nephrol. JASN 2004, 15, 1983–1992. [Google Scholar] [CrossRef]
- Tan, K.C.; Chow, W.-S.; Ai, V.H.; Metz, C.; Bucala, R.; Lam, K.S. Advanced glycation end products and endothelial dysfunction in type 2 diabetes. Diabetes Care 2002, 25, 1055–1059. [Google Scholar] [CrossRef] [Green Version]
- Nakagami, H.; Kaneda, Y.; Ogihara, T.; Morishita, R. Endothelial dysfunction in hyperglycemia as a trigger of atherosclerosis. Curr. Diabetes Rev. 2005, 1, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Felaco, M.; Grilli, A.; De Lutiis, M.A.; Patruno, A.; Libertini, N.; Taccardi, A.A.; Di Napoli, P.; Di Giulio, C.; Barbacane, R.; Conti, P. Endothelial nitric oxide synthase (eNOS) expression and localization in healthy and diabetic rat hearts. Ann. Clin. Lab. Sci. 2001, 31, 179–186. [Google Scholar]
- Hu, D.; Yin, C.; Luo, S.; Habenicht, A.; Mohanta, S.K. Vascular Smooth Muscle Cells Contribute to Atherosclerosis Immunity. Front. Immunol. 2019, 10, 1101. [Google Scholar] [CrossRef]
- Nishimura, M. Breeding of Mice Strains for Diabetes Mellitus. Exp. Anim. 1969, 18, 147–157. [Google Scholar] [CrossRef]
- Mita, T.; Goto, H.; Azuma, K.; Jin, W.L.; Nomiyama, T.; Fujitani, Y.; Hirose, T.; Kawamori, R.; Watada, H. Impact of insulin resistance on enhanced monocyte adhesion to endothelial cells and atherosclerogenesis independent of LDL cholesterol level. Biochem. Biophys. Res. Commun. 2010, 395, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Mauldin, J.P.; Srinivasan, S.; Mulya, A.; Gebre, A.; Parks, J.S.; Daugherty, A.; Hedrick, C.C. Reduction in ABCG1 in Type 2 Diabetic Mice Increases Macrophage Foam Cell Formation. J. Biol. Chem. 2006, 281, 21216–21224. [Google Scholar] [CrossRef] [Green Version]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Gaspari, A.; Di Minno, G.; Ritieni, A. Red Wine Consumption and Cardiovascular Health. Molecules 2019, 24, 3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsos, A.; Riccadonna, S.; Ulaszewska, M.M.; Franceschi, P.; Trošt, K.; Galvin, A.; Braune, T.; Fava, F.; Perenzoni, D.; Mattivi, F.; et al. Two apples a day lower serum cholesterol and improve cardiometabolic biomarkers in mildly hypercholesterolemic adults: A randomized, controlled, crossover trial. Am. J. Clin. Nutr. 2020, 111, 307–318. [Google Scholar] [CrossRef]
- Nagasako-Akazome, Y.; Kanda, T.; Ohtake, Y.; Shimasaki, H.; Kobayashi, T. Apple Polyphenols Influence Cholesterol Metabolism in Healthy Subjects with Relatively High Body Mass Index. J. Oleo Sci. 2007, 56, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, D.; Ji, Y.; Liu, Y.; Xu, L.; Guo, Y. Dietary Supplementation of Black Rice Anthocyanin Extract Regulates Cholesterol Metabolism and Improves Gut Microbiota Dysbiosis in C57BL/6J Mice Fed a High-Fat and Cholesterol Diet. Mol. Nutr. Food Res. 2020, 64, e1900876. [Google Scholar] [CrossRef]
- Yao, S.-L.; Xu, Y.; Zhang, Y.-Y.; Lu, Y.-H. Black rice and anthocyanins induce inhibition of cholesterol absorption in vitro. Food Funct. 2013, 4, 1602–1608. [Google Scholar] [CrossRef]
- Chamnansilpa, N.; Aksornchu, P.; Adisakwattana, S.; Thilavech, T.; Mäkynen, K.; Dahlan, W.; Ngamukote, S. Anthocyanin-rich fraction from Thai berries interferes with the key steps of lipid digestion and cholesterol absorption. Heliyon 2020, 6, 05408. [Google Scholar] [CrossRef]
- Stary, H.C.; Chandler, A.B.; Dinsmore, R.E.; Fuster, V.; Glagov, S.; Insull, W.; Rosenfeld, M.E.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A Definition of Advanced Types of Atherosclerotic Lesions and a Histological Classification of Atherosclerosis. Circulation 1995, 92, 1355–1374. [Google Scholar] [CrossRef]
- Heiss, C.; Rodriguez-Mateos, A.; Kelm, M. Central Role of eNOS in the Maintenance of Endothelial Homeostasis. Antioxidants Redox Signal. 2015, 22, 1230–1242. [Google Scholar] [CrossRef] [Green Version]
- Raij, L. Nitric Oxide in the Pathogenesis of Cardiac Disease. J. Clin. Hypertens. 2006, 8, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Förstermann, U. Nitric oxide in the pathogenesis of vascular disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Taguchi, K.; Tano, I.; Kaneko, N.; Matsumoto, T.; Kobayashi, T. Plant polyphenols Morin and Quercetin rescue nitric oxide production in diabetic mouse aorta through distinct pathways. Biomed. Pharmacother. 2020, 129, 110463. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Jin, C.-Y.; Bi, X.-K.; Zhao, Y.-B.; Xu, S.-J.; Wang, M.-H.; Yu, L.; Sun, Y.-X.; Hu, D. Green Tea Polyphenol Epigallocatechin-3-Gallate Promotes Reendothelialization in Carotid Artery of Diabetic Rabbits by Reactivating Akt/eNOS Pathway. Front. Pharmacol. 2018, 9, 1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuuchi, R.; Shimizu, I.; Yoshida, Y.; Hayashi, Y.; Ikegami, R.; Suda, M.; Katsuumi, G.; Wakasugi, T.; Nakao, M.; Minamino, T. Boysenberry polyphenol inhibits endothelial dysfunction and improves vascular health. PLoS ONE 2018, 13, e0202051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doran, A.C.; Meller, N.; McNamara, C.A. Role of Smooth Muscle Cells in the Initiation and Early Progression of Atherosclerosis. Arter. Thromb. Vasc. Biol. 2008, 28, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, Y.; Imaizumi, T.; Matsuura, H.; Ohara, S.; Takano, K.; Suyama, K.; Hashimoto, K.; Nozawa, R.; Suzuki, H.; Hosoya, M. Renal expression of alpha-smooth muscle actin and c-Met in children with Henoch–Schönlein purpura nephritis. Pediatr. Nephrol. 2008, 23, 913–919. [Google Scholar] [CrossRef]
- Lin, H.-C.; Hsieh, M.-J.; Peng, C.-H.; Yang, S.-F.; Huang, C.-N. Pterostilbene Inhibits Vascular Smooth Muscle Cells Migration and Matrix Metalloproteinase-2 through Modulation of MAPK Pathway. J. Food Sci. 2015, 80, H2331–H2335. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.N.; Narkevich, I.A.; Akamova, A.V.; Nemyatykh, O.D.; Flisyuk, E.V.; Luzhanin, V.G.; Povydysh, M.N.; Mikhailova, I.V.; Pozharitskaya, O.N. Medical Species Used in Russia for the Management of Diabetes and Related Disorders. Front. Pharmacol. 2021, 12, 697411. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Wu, G.; Han, D.; Stipkovits, L.; Wu, X.; Tang, S.; Brennan, M.A.; Brennan, C.S. The effects of bioactive compounds from blueberry and blackcurrant powders on the inhibitory activities of oat bran pastes against α-amylase and α-glucosidase linked to type 2 diabetes. Food Res. Int. 2020, 138, 109756. [Google Scholar] [CrossRef] [PubMed]
- Nanashima, N.; Horie, K.; Tomisawa, T.; Chiba, M.; Nakano, M.; Fujita, T.; Maeda, H.; Kitajima, M.; Takamagi, S.; Uchiyama, D.; et al. Phytoestrogenic activity of blackcurrant (Ribes nigrum) anthocyanins is mediated through estrogen receptor alpha. Mol. Nutr. Food Res. 2015, 59, 2419–2431. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horie, K.; Maeda, H.; Nanashima, N.; Oey, I. Potential Vasculoprotective Effects of Blackcurrant (Ribes nigrum) Extract in Diabetic KK-Ay Mice. Molecules 2021, 26, 6459. https://doi.org/10.3390/molecules26216459
Horie K, Maeda H, Nanashima N, Oey I. Potential Vasculoprotective Effects of Blackcurrant (Ribes nigrum) Extract in Diabetic KK-Ay Mice. Molecules. 2021; 26(21):6459. https://doi.org/10.3390/molecules26216459
Chicago/Turabian StyleHorie, Kayo, Hayato Maeda, Naoki Nanashima, and Indrawati Oey. 2021. "Potential Vasculoprotective Effects of Blackcurrant (Ribes nigrum) Extract in Diabetic KK-Ay Mice" Molecules 26, no. 21: 6459. https://doi.org/10.3390/molecules26216459
APA StyleHorie, K., Maeda, H., Nanashima, N., & Oey, I. (2021). Potential Vasculoprotective Effects of Blackcurrant (Ribes nigrum) Extract in Diabetic KK-Ay Mice. Molecules, 26(21), 6459. https://doi.org/10.3390/molecules26216459