
Correlation among Metabolic Changes in Tea Plant Camellia sinensis (L.) Shoots, Green Tea Quality and the Application of Cow Manure to Tea Plantation Soils

 , , and
, , and
Abstract
:
1. Introduction
2. Results
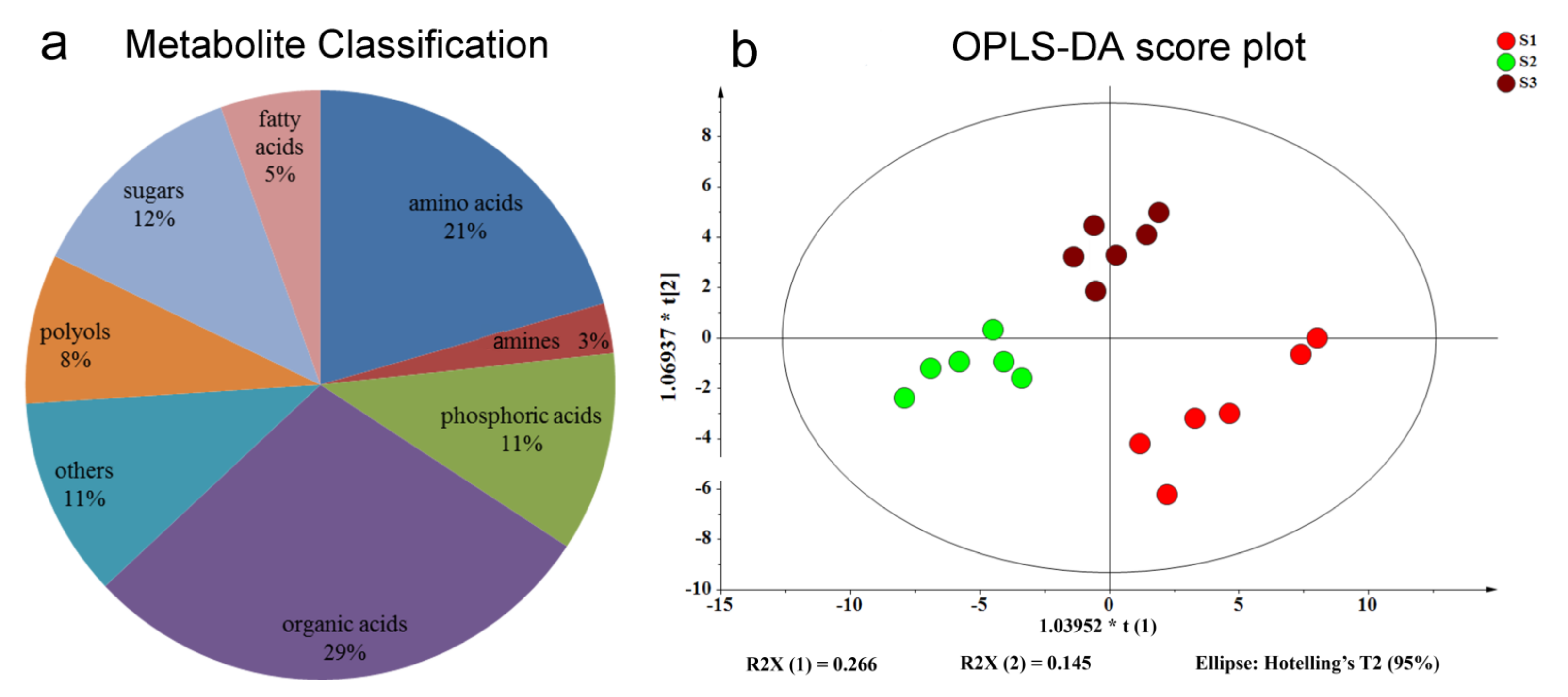
2.1. Analysis of Metabolic Profiling in Tea Shoots under Different Treatments
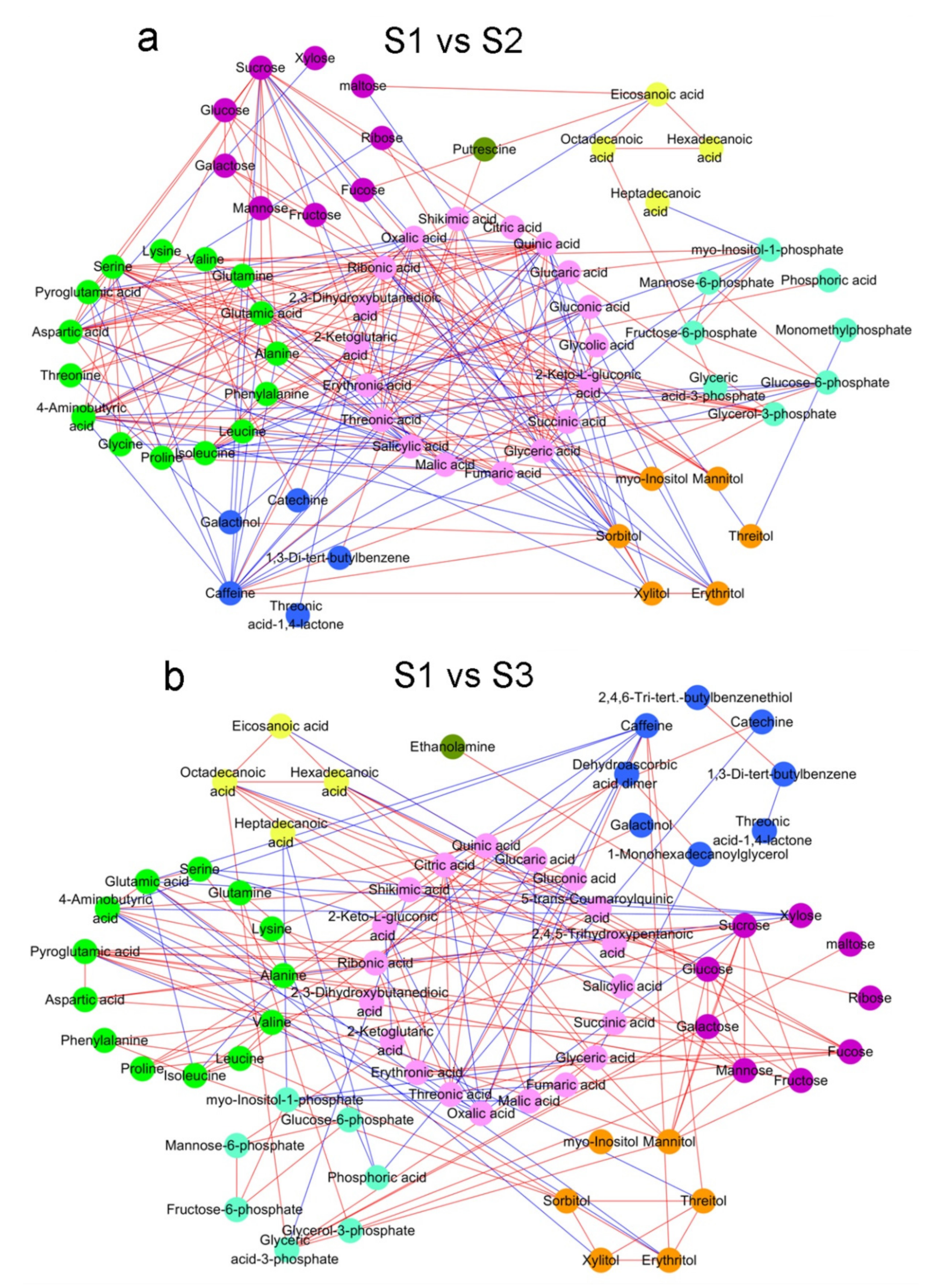
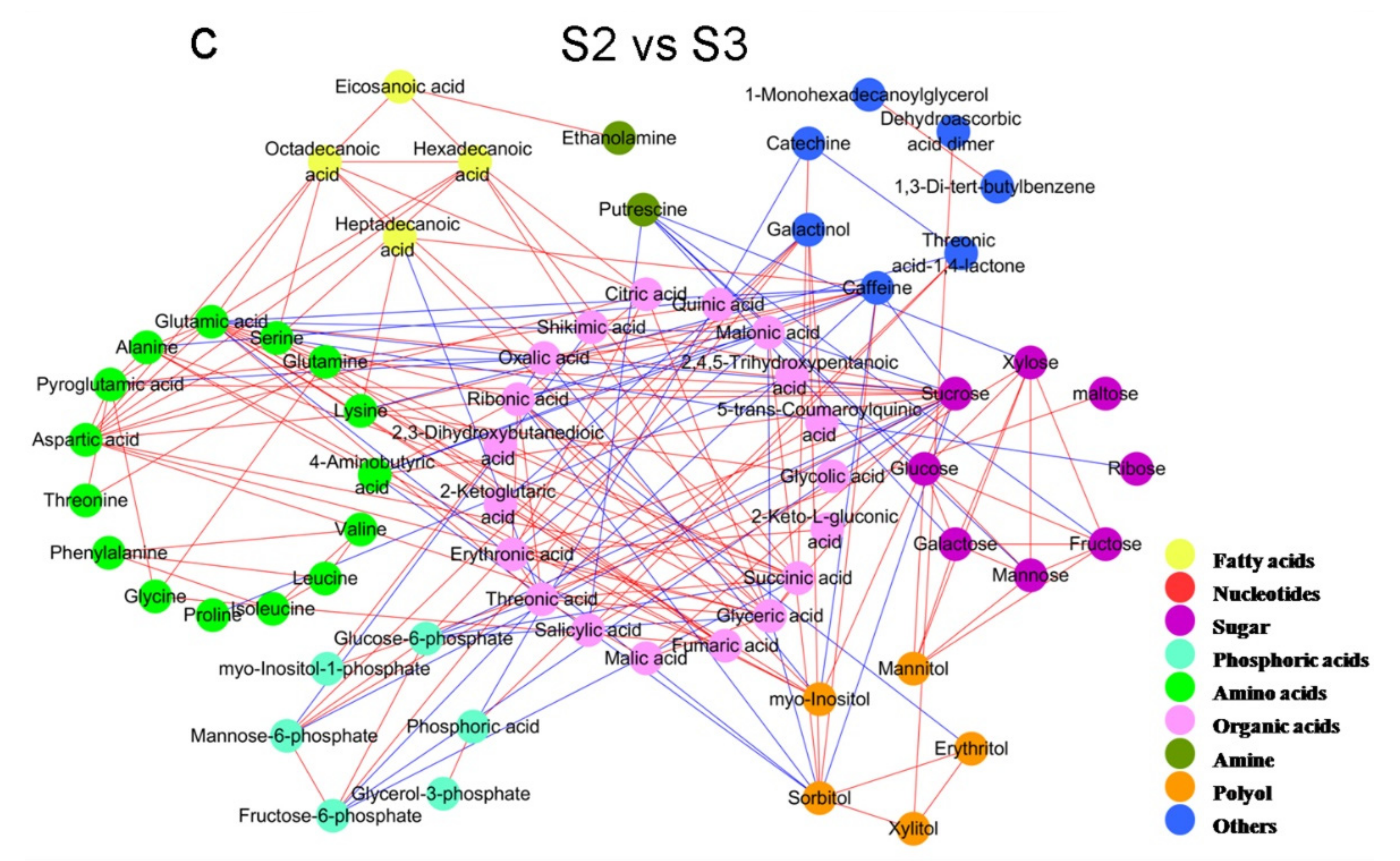
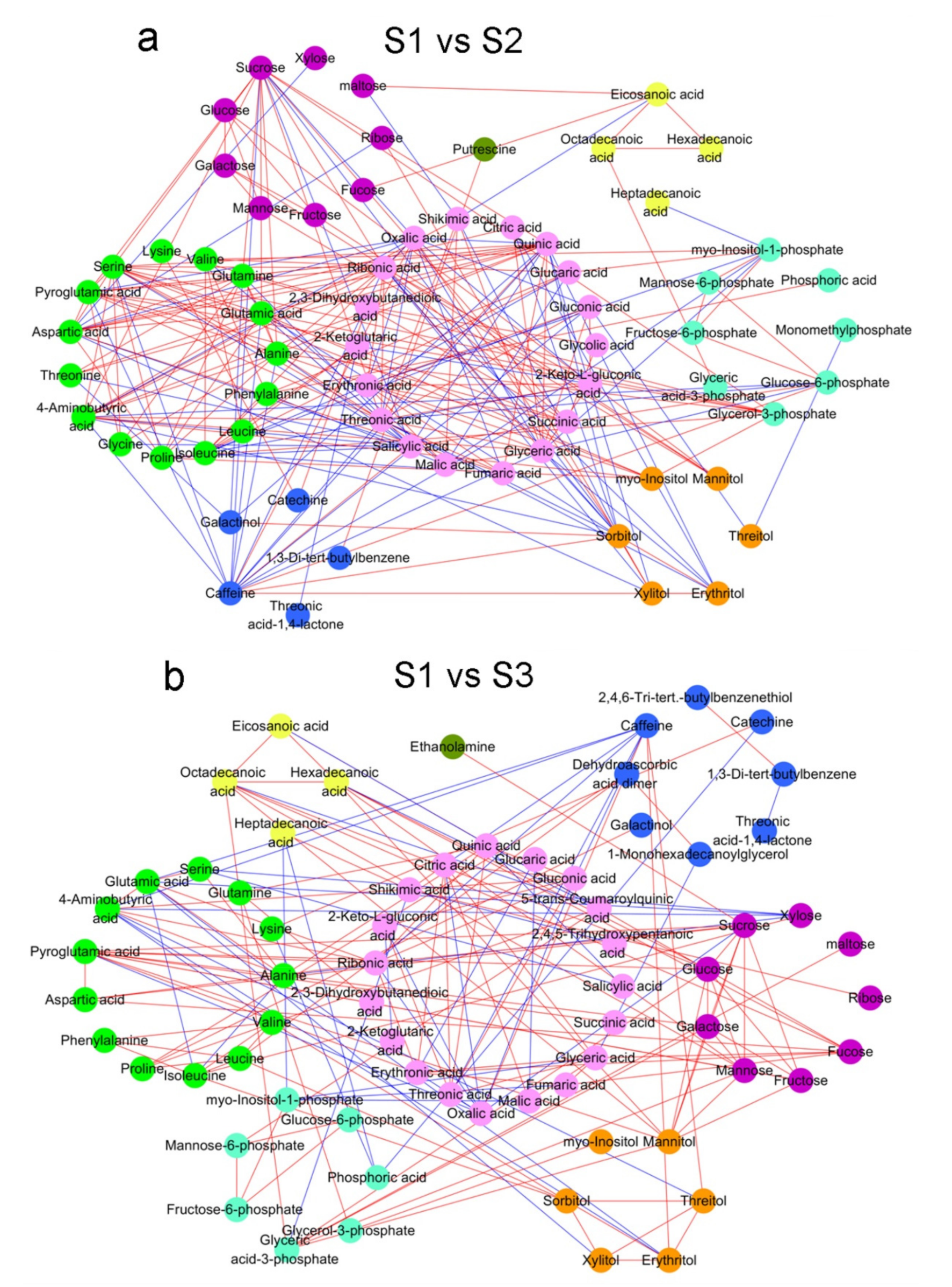
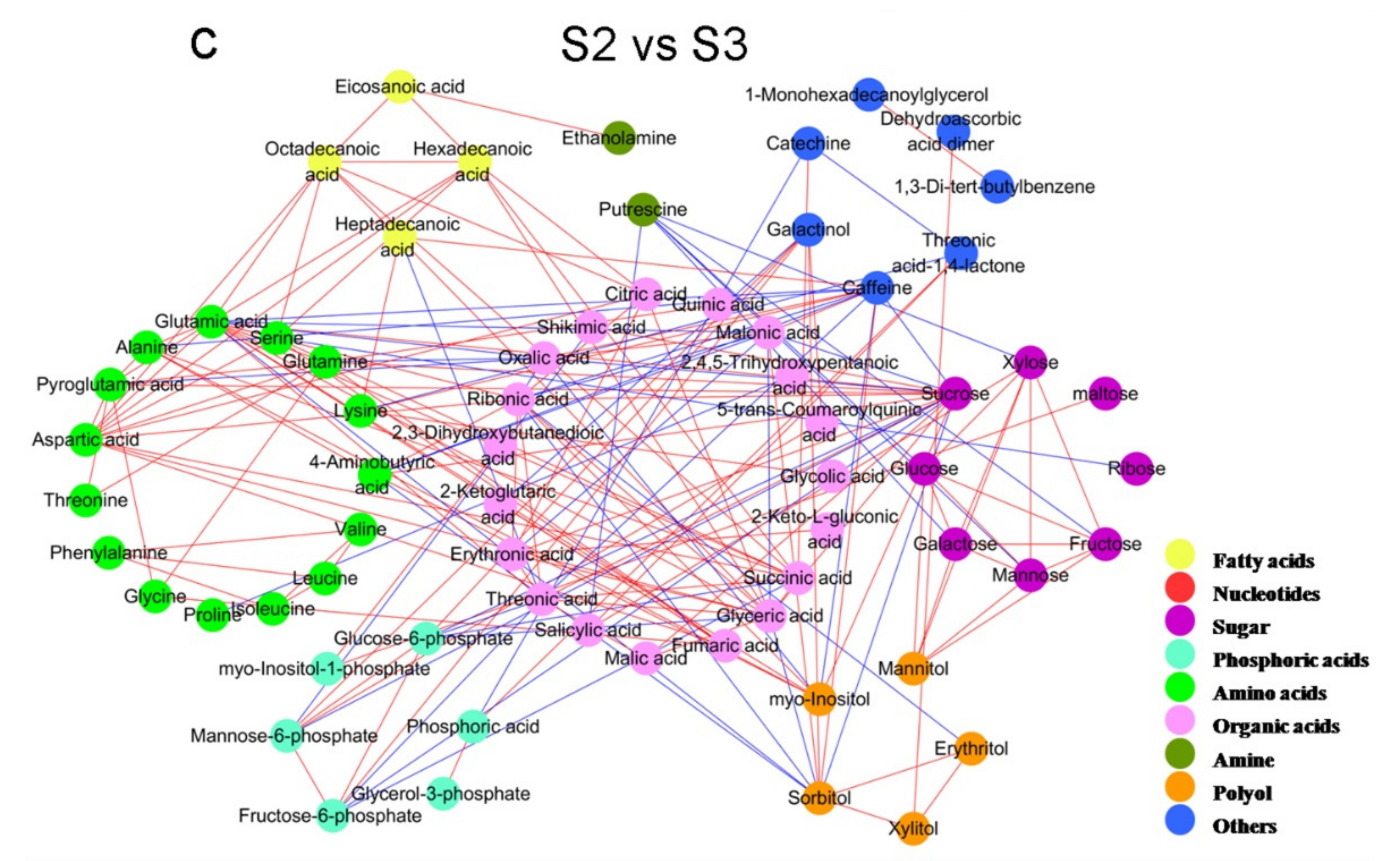
2.2. Analysis of Metabolite Correlations in Tea Shoots under Different Treatments
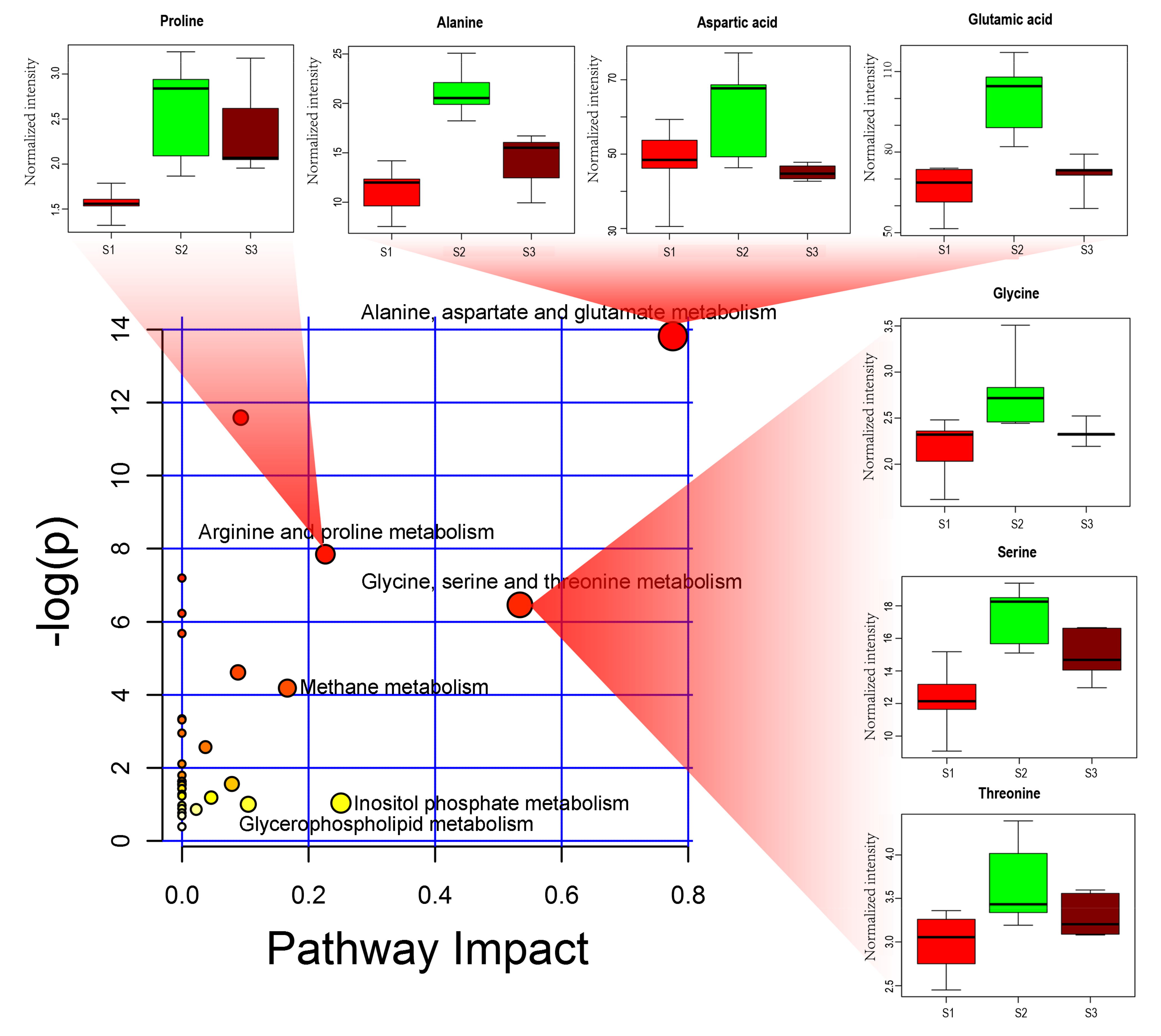
2.3. Analysis of Difference Metabolites in Tea Shoots under Different Treatments
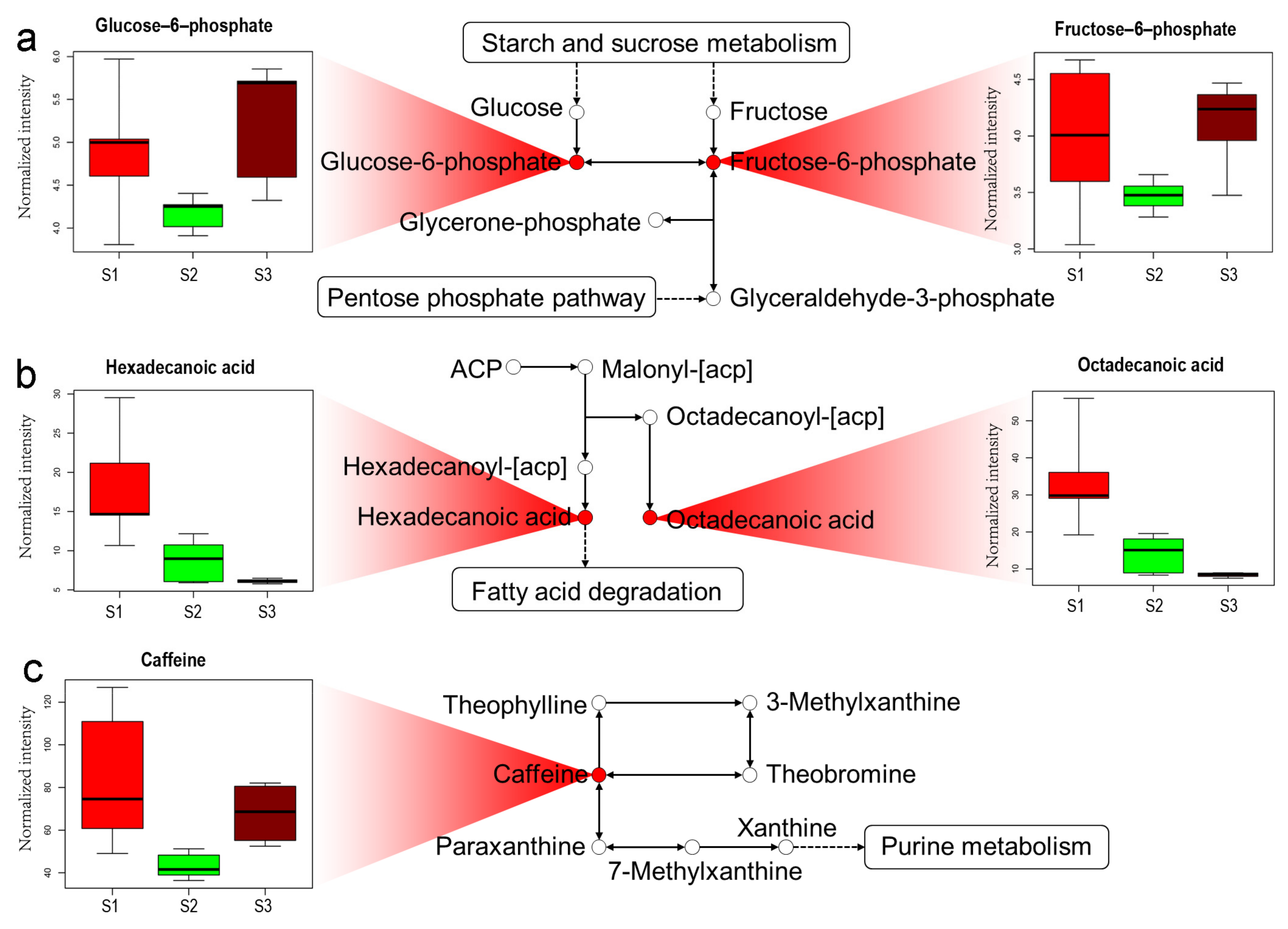
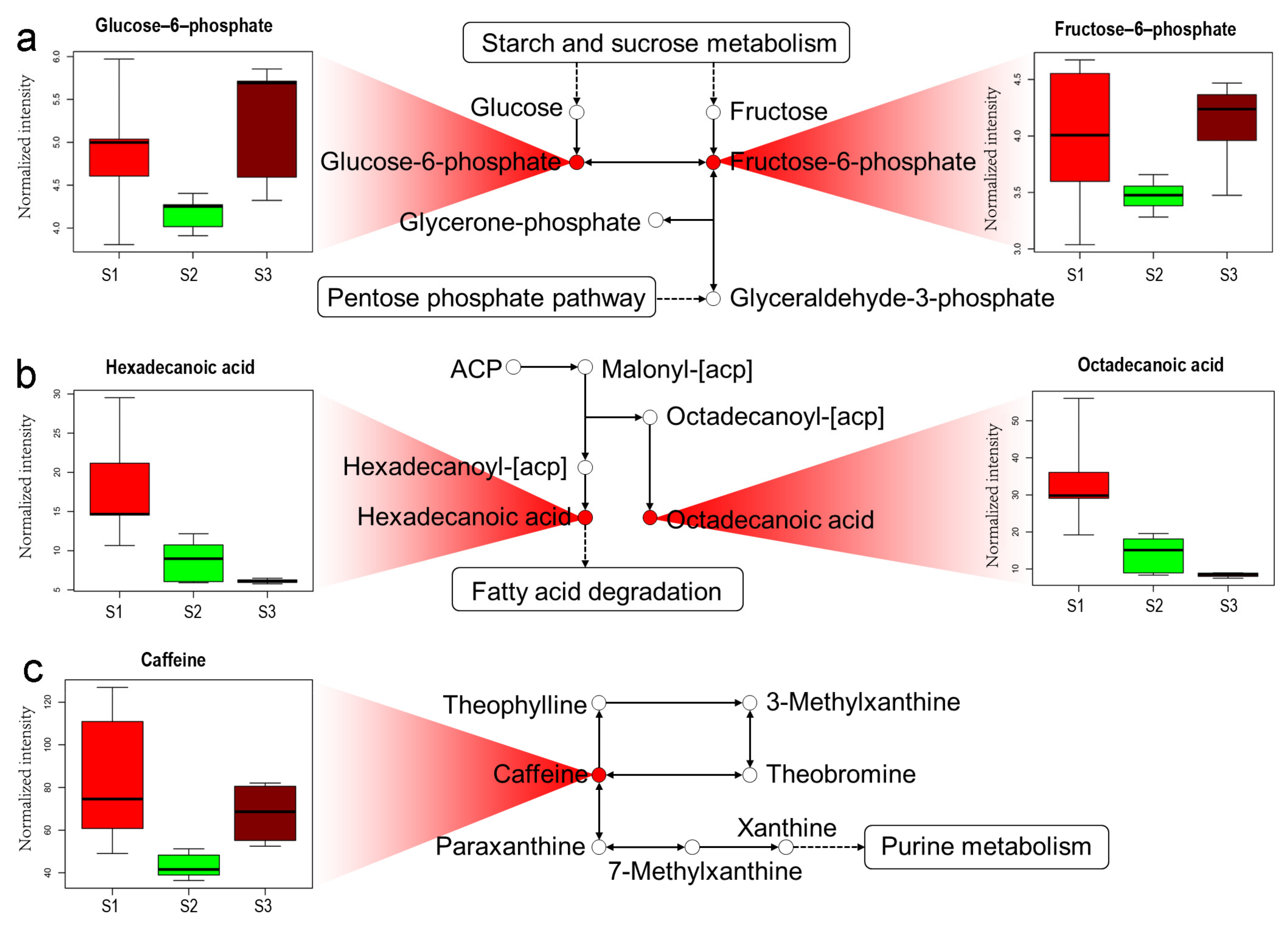
2.4. Analysis of Metabolic Pathway in Tea Shoots under Different Treatments
2.5. Sensory Evaluation of Green Teas under Different Treatments
3. Discussion
3.1. The Application of Cow Manure Affects the Amino Acid Metabolism in Tea Shoots
3.2. The Application of Cow Manure Affects the Sugar Metabolism in Tea Shoots
3.3. The Application of Cow Manure Affects the Fatty Acid Metabolism in Tea Shoots
4. Materials and Methods
4.1. Field Trial
4.2. Plant Sampling
4.3. GC–MS Analysis
4.4. Pathway Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- FAO. FAOSTAT Value of agricultural production-Gross production value of tea. In FAOSTAT, Value of Agricultural Production; FAO: Rome, Italy, 2019. [Google Scholar]
- Liu, J.; Zhang, Q.; Liu, M.; Ma, L.; Shi, Y.; Ruan, J. Metabolomic analyses reveal distinct change of metabolites and quality of green tea during the short duration of a single spring season. J. Agric. Food Chem. 2016, 64, 3302–3309. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Qi, D.; Yang, T.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Xie, D.; Tan, J. Nontargeted analysis using ultraperformance liquid chromatography–quadrupole time-of-flight mass spectrometry uncovers the effects of harvest season on the metabolites and taste quality of tea (Camellia sinensis L.). J. Agric. Food Chem. 2015, 63, 9869–9878. [Google Scholar] [CrossRef]
- Chen, G.-H.; Yang, C.-Y.; Lee, S.-J.; Wu, C.-C.; Tzen, J.T. Catechin content and the degree of its galloylation in oolong tea are inversely correlated with cultivation altitude. J. Food Drug Anal. 2014, 22, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Yan, J.; Cui, J.; Mao, S.; Li, M.; Liao, X.; Tong, H. Dynamic changes in amino acids, catechins, caffeine and gallic acid in green tea during withering. J. Food Compos. Anal. 2018, 66, 98–108. [Google Scholar] [CrossRef]
- Das, P.R.; Kim, Y.; Hong, S.-J.; Eun, J.-B. Profiling of volatile and non-phenolic metabolites—Amino acids, organic acids, and sugars of green tea extracts obtained by different extraction techniques. Food Chem. 2019, 296, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.-T.; Zheng, X.; Li, S. Tea aroma formation. Food Sci. Hum. Wellness 2015, 4, 9–27. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Fu, X.; Mei, X.; Zhou, Y.; Cheng, S.; Zeng, L.; Dong, F.; Yang, Z. Proteolysis of chloroplast proteins is responsible for accumulation of free amino acids in dark-treated tea (Camellia sinensis) leaves. J. Proteom. 2017, 157, 10–17. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Tai, Y.; Wang, X.; Ho, C.-T.; Wan, X. Gene discovery of characteristic metabolic pathways in the tea plant (Camellia sinensis) using ‘Omics’-based network approaches: A future perspective. Front. Plant Sci. 2018, 9, 480. [Google Scholar] [CrossRef]
- Maghanga, J.; Kituyi, J.; Kisinyo, P.; Ng’Etich, W. Impact of nitrogen fertilizer applications on surface water nitrate levels within a Kenyan tea plantation. J. Chem. 2013, 2013, 196516. [Google Scholar] [CrossRef] [Green Version]
- Ni, K.; Shi, Y.-Z.; Yi, X.-Y.; Zhang, Q.-F.; Fang, L.; Ma, L.-F.; Ruan, J. Effects of long-term nitrogen application on soil acidification and solution chemistry of a tea plantation in China. Agric. Ecosyst. Environ. 2018, 252, 74–82. [Google Scholar]
- Ji, L.; Wu, Z.; You, Z.; Yi, X.; Ni, K.; Guo, S.; Ruan, J. Effects of organic substitution for synthetic N fertilizer on soil bacterial diversity and community composition: A 10-year field trial in a tea plantation. Agric. Ecosyst. Environ. 2018, 268, 124–132. [Google Scholar] [CrossRef]
- Ahmed, S.; Stepp, J.R.; Orians, C.; Griffin, T.; Matyas, C.; Robbat, A.; Cash, S.; Xue, D.; Long, C.; Unachukwu, U. Effects of extreme climate events on tea (Camellia sinensis) functional quality validate indigenous farmer knowledge and sensory preferences in tropical China. PLoS ONE 2014, 9, e109126. [Google Scholar] [CrossRef] [Green Version]
- Bulluck Iii, L.; Brosius, M.; Evanylo, G.; Ristaino, J. Organic and synthetic fertility amendments influence soil microbial, physical and chemical properties on organic and conventional farms. Appl. Soil Ecol. 2002, 19, 147–160. [Google Scholar] [CrossRef]
- Xie, S.; Feng, H.; Yang, F.; Zhao, Z.; Hu, X.; Wei, C.; Liang, T.; Li, H.; Geng, Y. Does dual reduction in chemical fertilizer and pesticides improve nutrient loss and tea yield and quality? A pilot study in a green tea garden in Shaoxing, Zhejiang Province, China. Environ. Sci. Pollut. Res. 2019, 26, 2464–2476. [Google Scholar] [CrossRef]
- Liu, M.-Y.; Burgos, A.; Ma, L.; Zhang, Q.; Tang, D.; Ruan, J. Lipidomics analysis unravels the effect of nitrogen fertilization on lipid metabolism in tea plant (Camellia sinensis L.). BMC Plant Biol. 2017, 17, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Song, Q.; Li, D.; Wan, X. Discrimination of the production season of Chinese green tea by chemical analysis in combination with supervised pattern recognition. J. Agric. Food Chem. 2012, 60, 7064–7070. [Google Scholar] [CrossRef]
- Ding, Z.; Jia, S.; Wang, Y.; Xiao, J.; Zhang, Y. Phosphate stresses affect ionome and metabolome in tea plants. Plant Physiol. Biochem. 2017, 120, 30–39. [Google Scholar] [CrossRef]
- Keenan, E.K.; Finnie, M.D.; Jones, P.S.; Rogers, P.J.; Priestley, C.M. How much theanine in a cup of tea? Effects of tea type and method of preparation. Food Chem. 2011, 125, 588–594. [Google Scholar] [CrossRef]
- Li, Y.; Chen, C.; Li, Y.; Ding, Z.; Shen, J.; Wang, Y.; Zhao, L.; Xu, M. The identification and evaluation of two different color variations of tea. J. Agric. Food Chem. 2016, 96, 4951–4961. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Wang, Y.; Hu, J.; Ding, Z.; Liang, Q.; Zhang, Y.; Wang, H. Mineral and metabolic profiles in tea leaves and flowers during flower development. Plant Physiol. Biochem. 2016, 106, 316–326. [Google Scholar] [CrossRef]
- Mao, A.; Su, H.; Fang, S.; Chen, X.; Ning, J.; Ho, C.; Wan, X. Effects of roasting treatment on non-volatile compounds and taste of green tea. Int. J. Food Sci. Technol. 2018, 53, 2586–2594. [Google Scholar] [CrossRef]
- Wang, L.-F.; Kim, D.-M.; Lee, C.Y. Effects of heat processing and storage on flavanols and sensory qualities of green tea beverage. J. Agric. Food Chem. 2000, 48, 4227–4232. [Google Scholar] [CrossRef]
- Wang, C.; Lv, S.; Wu, Y.; Gao, X.; Li, J.; Zhang, W.; Meng, Q. Oolong tea made from tea plants from different locations in Yunnan and Fujian, China showed similar aroma but different taste characteristics. SpringerPlus 2016, 5, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fhatuwani, M.N.; Nokwanda, M.P. Effect of Seasonal Variations and Growth Conditions on Carbohydrate Partitioning in Different Organs and the Quality of Bush Tea. HortScience 2018, 53, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.-Q.; Li, Q.-S.; Xiang, L.-P.; Liang, Y.-R. Recent advances in volatiles of teas. Molecules 2016, 21, 338. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Li, W.-L.; Chen, L.-B.; Wang, Y.-R.; Chen, X. Identification of the different aroma compounds between conventional and freeze dried wuyi rock tea (dangui) using headspace solid phase microextraction. J. Food Sci. Technol. 2013, 19, 805–811. [Google Scholar]
- Lee, J.; Chambers, D.H.; Chambers, E.; Adhikari, K.; Yoon, Y. Volatile aroma compounds in various brewed green teas. Molecules 2013, 18, 10024–10041. [Google Scholar] [CrossRef]
- Wright, A.J.; Fishwick, M.J. Lipid degradation during manufacture of black tea. Phytochemistry 1979, 18, 1511–1513. [Google Scholar] [CrossRef]
- Mahanta, P.; Harazika, M.; Takeo, T. Flavour volatiles and lipids in various components of tea shoots Camellia sinensis, (L.), O. kuntze. J. Sci. Food Agric. 1985, 36, 1130–1132. [Google Scholar]
- Bondarovich, H.; Giammarino, A.S.; Renner, J.; Shephard, F.; Shingler, A.; Gianturco, M.A. Volatiles in tea some aspects of the chemistry of tea. a contribution to the knowledge of the volatile constituents. J. Agric. Food Chem. 1967, 15, 36–47. [Google Scholar] [CrossRef]
- Mahanta, P.K.; Tamuli, P.; Bhuyan, L.P. Changes of fatty acid contents, lipoxygenase activities, and volatiles during black tea manufacture. J. Agric. Food Chem. 1993, 41, 1677–1683. [Google Scholar] [CrossRef]
- Owuor, P.; Okal, A.; Kamau, D.; Msomba, S.; Uwimana, M.; Kamunya, S.; Mang’uro, L. Influence of nitrogen fertilizer rates and harvesting intervals on clonal tea green leaf fatty acid levels in the Lake Victoria basin of Kenya. J. Food Agric. Environ. 2013, 11, 667–674. [Google Scholar]
- Huppe, H.; Turpin, D. Integration of carbon and nitrogen metabolism in plant and algal cells. Annu. Rev. Plant Physiol. 1994, 45, 577–607. [Google Scholar] [CrossRef]
- Ferrario-Méry, S.; Thibaud, M.-C.; Betsche, T.; Valadier, M.-H.; Foyer, C.H. Modulation of carbon and nitrogen metabolism, and of nitrate reductase, in untransformed and transformed Nicotiana plumbaginifolia during CO2 enrichment of plants grown in pots and in hydroponic culture. Planta 1997, 202, 510–521. [Google Scholar] [CrossRef]
- Brupbacher, R.H. Analytical methods and procedures used in the soil testing laboratory. In LSU Agricultural Experiment Station Reports; Louisiana State University and Agricultural and Mechanical College, Agricultural Experiment Station: Baton Rouge, Louisiana, 1968; p. 454. Available online: http://digitalcommons.lsu.edu/agexp/454 (accessed on 30 August 2020).
- Gai, Z.; Wang, Y.; Jiang, J.; Xie, H.; Ding, Z.; Ding, S.; Wang, H. The quality evaluation of tea (Camellia sinensis) varieties based on the metabolomics. HortScience 2019, 54, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.D.; Wiemann, S. KEGGgraph: A graph approach to KEGG PATHWAY in R and bioconductor. J. Bioinform. 2009, 25, 1470–1471. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the human adult urinary metabolome variations with age, body mass index, and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Groups | Identified Chemicals | log2fc_S2/S1 | log2fc_S3/S1 | log2fc_S3/S2 |
|---|---|---|---|---|
| Amino acids | alanine | 0.9265 | 0.3433 | −0.5832 |
| proline | 0.7331 | 0.6034 | −0.1297 | |
| salicylic acid | −0.6537 | −0.7162 | −0.0625 | |
| pyroglutamic acid | 0.7666 | 0.1668 | −0.5998 | |
| 4-aminobutyric acid | 0.6777 | 0.2170 | −0.4607 | |
| glycine | 0.3689 | 0.1127 | −0.2562 | |
| glutamic acid | 0.6056 | 0.1152 | −0.4905 | |
| glutamine | 2.5405 | 1.1647 | −1.3757 | |
| serine | 0.5055 | 0.2923 | −0.2132 | |
| threonine | 0.3040 | 0.1511 | −0.1528 | |
| aspartic acid | 0.3761 | −0.0801 | −0.4562 | |
| Organic acids | succinic acid | 0.4559 | −0.0315 | −0.4874 |
| quinic acid | 0.9348 | 0.7440 | −0.1908 | |
| malonic acid | 0.1325 | 0.3861 | 0.2536 | |
| threonic acid | 0.7460 | 0.2558 | −0.4902 | |
| glycolic acid | 0.2467 | 0.3599 | 0.1132 | |
| Fatty acids | hexadecanoic acid | −1.0449 | −1.5632 | −0.5183 |
| octadecanoic acid | −1.2808 | −2.0232 | −0.7425 | |
| eicosanoic acid | −0.4435 | −0.6362 | −0.1927 | |
| Sugars | Sucrose | 1.0156 | 0.0591 | −0.9565 |
| Polyols | myo-inositol | 0.5233 | 0.2252 | −0.2982 |
| Phosphoric acids | glycerol-3-phosphate | 0.3133 | 0.3073 | −0.0060 |
| Others | Caffeine | −0.9640 | −0.3172 | 0.6468 |
| Sensory Indexes (SIs) * | Spring (20 April) | Early Summer (6 June) | Late Summer (11 August) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| (SI%) | S1 | S2 | S3 | S1 | S2 | S3 | S1 | S2 | S3 |
| Appearance (25%) | 81.67 ± 0.58 b | 86.33 ± 0.58 a | 87.67 ± 0.58 a | 90.33 ± 0.58 a | 88.33 ± 0.58 b | 88.33 ± 0.58 b | 86.33 ± 0.58 b | 88.00 ± 1.00 b | 92.00 ± 0.00 a |
| Aroma (25%) | 88.00 ± 0.00 b | 85.33 ± 0.58 c | 90.33 ± 0.58 a | 90.33 ± 0.58 a | 87.33 ± 0.58 b | 81.67 ± 0.58 c | 83.00 ± 0.00 c | 88.67 ± 0.58 b | 90.33 ± 0.58 a |
| Infusion color (10%) | 85.33 ± 0.58 b | 88.33 ± 0.58 a | 88.33 ± 0.58 a | 90.67 ± 0.58 a | 87.67 ± 0.58 b | 80.00 ± 0.00 c | 87.33 ± 0.58 c | 88.67 ± 0.58 b | 90.00 ± 0.00 a |
| Infusion taste (30%) | 81.67 ± 0.58 c | 85.67 ± 0.58 b | 89.00 ± 0.00 a | 90.33 ± 0.58 a | 87.00 ± 0.00 b | 82.00 ± 0.00 c | 83.00 ± 1.00 c | 86.00 ± 0.00 b | 90.00 ± 0.00 a |
| Infused leaf (color/shape) (10%) | 82.67 ± 0.58 c | 86.00 ± 0.00 b | 88.00 ± 0.00 a | 90.00 ± 0.00 a | 87.67 ± 0.58 b | 88.33 ± 0.58 b | 77.67 ± 0.58 c | 83.00 ± 1.00 b | 88.33 ± 0.58 a |
| Overall | 83.72 | 86.05 | 88.83 | 90.33 | 87.55 | 83.93 | 83.73 | 87.13 | 90.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Fan, K.; Wang, L.; Ma, D.; Wang, Y.; Kong, X.; Li, H.; Ren, Y.; Ding, Z. Correlation among Metabolic Changes in Tea Plant Camellia sinensis (L.) Shoots, Green Tea Quality and the Application of Cow Manure to Tea Plantation Soils. Molecules 2021, 26, 6180. https://doi.org/10.3390/molecules26206180
Sun L, Fan K, Wang L, Ma D, Wang Y, Kong X, Li H, Ren Y, Ding Z. Correlation among Metabolic Changes in Tea Plant Camellia sinensis (L.) Shoots, Green Tea Quality and the Application of Cow Manure to Tea Plantation Soils. Molecules. 2021; 26(20):6180. https://doi.org/10.3390/molecules26206180
Chicago/Turabian StyleSun, Litao, Kai Fan, Linlin Wang, Dexin Ma, Yu Wang, Xiaojun Kong, Hongyan Li, Yonglin Ren, and Zhaotang Ding. 2021. "Correlation among Metabolic Changes in Tea Plant Camellia sinensis (L.) Shoots, Green Tea Quality and the Application of Cow Manure to Tea Plantation Soils" Molecules 26, no. 20: 6180. https://doi.org/10.3390/molecules26206180
APA StyleSun, L., Fan, K., Wang, L., Ma, D., Wang, Y., Kong, X., Li, H., Ren, Y., & Ding, Z. (2021). Correlation among Metabolic Changes in Tea Plant Camellia sinensis (L.) Shoots, Green Tea Quality and the Application of Cow Manure to Tea Plantation Soils. Molecules, 26(20), 6180. https://doi.org/10.3390/molecules26206180