
The Peripheral Cannabinoid Receptor Type 1 (CB1) as a Molecular Target for Modulating Body Weight in Man

Abstract
1. Introduction
2. Peripheral CB1 Receptors

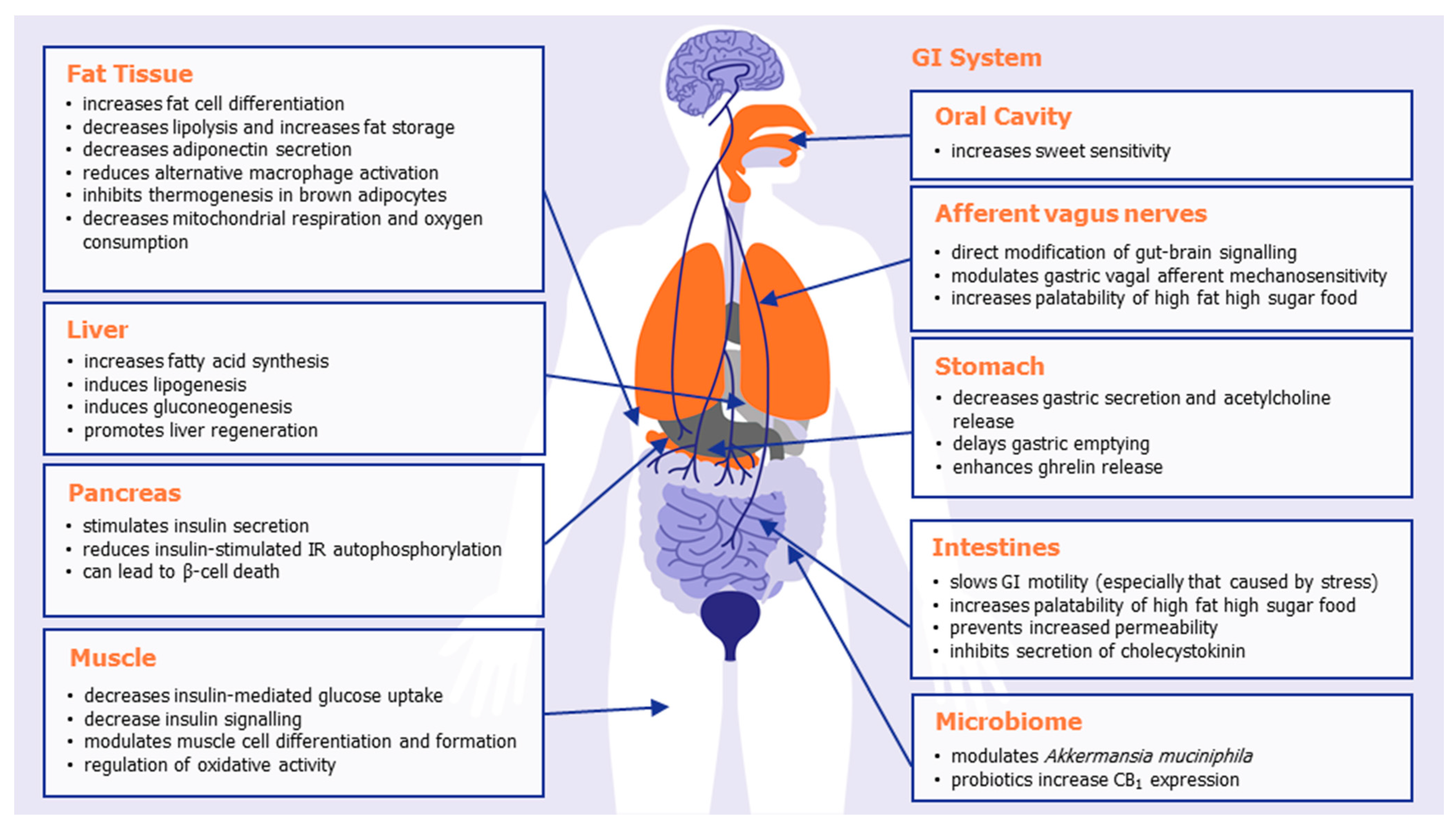{kind=link}
{kind=link}
| System/Organ | Tissue/Cell | Effect of CB1 Activation |
|---|---|---|
| GI system | Oral cavity | CB1 receptors are expressed in type II taste cells that also express the sweet-taste receptor, and their activation increases sweet sensitivity [35]. CB1 receptors on the tongue increase gustatory nerve responses [35]. |
| Stomach | CB1 is expressed on acid-secreting parietal cells [36]. CB1 activation decreases gastric secretion and acetylcholine release [37]. CB1 activation delays gastric emptying [38]. CB1 is expressed in ghrelin-positive gastric mucosal cells [39]. CB1 activation enhances ghrelin release from the stomach [40]. | |
| I cells of the small intestine | CB1 is expressed in enteroendocrine cells [41]. CB1 inhibits the secretion of the satiation hormone cholecystokinin [41]. | |
| Intestines | CB1 activation slows GI motility, particularly stress-induced motility [42,43]. CB1 activation prevents increased intestine permeability (leaky guts) [44]. Intestinal CB1 activation important for palatability of high fat high sugar foods [45]. CB1 deletion in intestinal epithelium reduces western diet preferences [24]. | |
| Afferent vagus nerves | CB1 receptors are expressed on vagal terminals [46,47]. Fasting increases CB1 expression on vagal afferent neurons [47]. The induction of feeding by peripherally CB1 activation is inhibited by vagal ablation [48]. CB1 activation modulates gastric vagal afferent mechanosensitivity to stretch/distension (leading to feeling of fullness) [39]. | |
| Microbiome | CB1 receptor antagonism [49] or THC [50] increases Akkermansia muciniphila. Probiotic treatment increases CB1 and/or CB2 expression [51,52]. | |
| Fat tissue | Adipocytes | CB1 is expressed on adipocytes [53]. CB1 deletion protects adult mice from diet-induced obesity [21]. CB1 increases adipocyte differentiation and adipogenesis [54]. CB1 activation increases PPARγ expression, a major regulator of adipose function [52]. CB1 enhances fat storage and reduces lipolysis [54,55]. CB1 decreases adiponectin production [54,56]. CB1 reduces alternative macrophage activation [21]. |
| White adipocyte mitochondria | CB1 activation decreases mitochondrial respiration and oxygen consumption [57,58]. | |
| Brown adipose tissue (BAT) | CB1 is upregulated during activation of BAT [59,60]. CB1 antagonism increases expression of uncoupling protein 1 (UCP-1) [61]. | |
| Liver | Hepatocytes | CB1 activation increases lipogenesis [62] CB1 activation increases fatty acid synthesis [62]. CB1 activation induces gluconeogenesis [63]. CB1 activation promotes liver regeneration by increasing mitotic progression [64]. CB1 knock-out mice are protected against diet-induced lipogenesis and steatosis [65]. |
| Pancreas | Pancreatic β-cells | CB1 activation stimulates basal and glucose-dependent insulin secretion [66,67]. CB1 activation impedes insulin-stimulated IR autophosphorylation [68]. CB1 receptors can lead to β-cell death [69]. |
| Muscle | Skeletal muscle cells | CB1 expression increases during skeletal muscle cell differentiation [31,33]. CB1 activation decreases insulin-mediated glucose uptake [31]. CB1 knockdown improves mitochondrial performance, increases whole-body muscle energy expenditure, and improves physical endurance [23]. CB1 receptor knockdown prevents diet-induced and age-induced insulin resistance [23]. |
| Myotubules | CB1 activation prevents myotubule formation [33]. CB1 activation inhibits sarcoplasmic Ca2+ release [70]. | |
| Skeletal muscle satellite cells | CB1 activation inhibits satellite cell differentiation [34]. | |
| Muscle Mitochondria | CB1 receptors regulates mitochondrial oxidative activity [20]. |
2.1. Effects of Peripheral CB1 Receptors on Appetite Hormones
2.2. Gut–Brain Axis
2.3. Microbiome
3. Therapeutic Utility of Peripheral CB1 Receptors as Molecular Targets
3.1. Peripherally Restricted CB1 Antagonists
| Peripherally Restricted CB1 Antagonists | Peripherally Restricted CB1 Agonists | Peripherally Restricted FAAH Inhibitors | |
|---|---|---|---|
| Preclinical research | Obesity [81,82,83,86] Type 2 diabetes [85,105] Prader–Willi syndrome [88] Chronic kidney disease [101] Diabetic nephropathy [93] Alcoholic liver steatosis [94] Alcoholism [99,100] Non-alcoholic liver steatosis [96] Obesity-related liver steatosis [95] Liver fibrosis [102] Pulmonary fibrogenesis [97,98] Skin fibrosis [103] | Inflammatory pain [106,107] Neuropathic pain [107,108] Bone cancer pain [109] Chemotherapy-induced pain [110] Migraine and medication overuse headache [111] Spasticity in multiple sclerosis [112] Gastrointestinal motility in colitis [42,43] Anticipatory nausea [113] Cardiac disease [114] | Neuropathic pain [115] Chemotherapy-induced neuropathy [116] Inflammatory pain [115,117,118] Diabetic neuropathy [119] Visceral pain [115] Migraine [120,121] Anticipatory nausea [113] Cystitis [122] Bladder overactivity [123] Gastric lesions [118] |
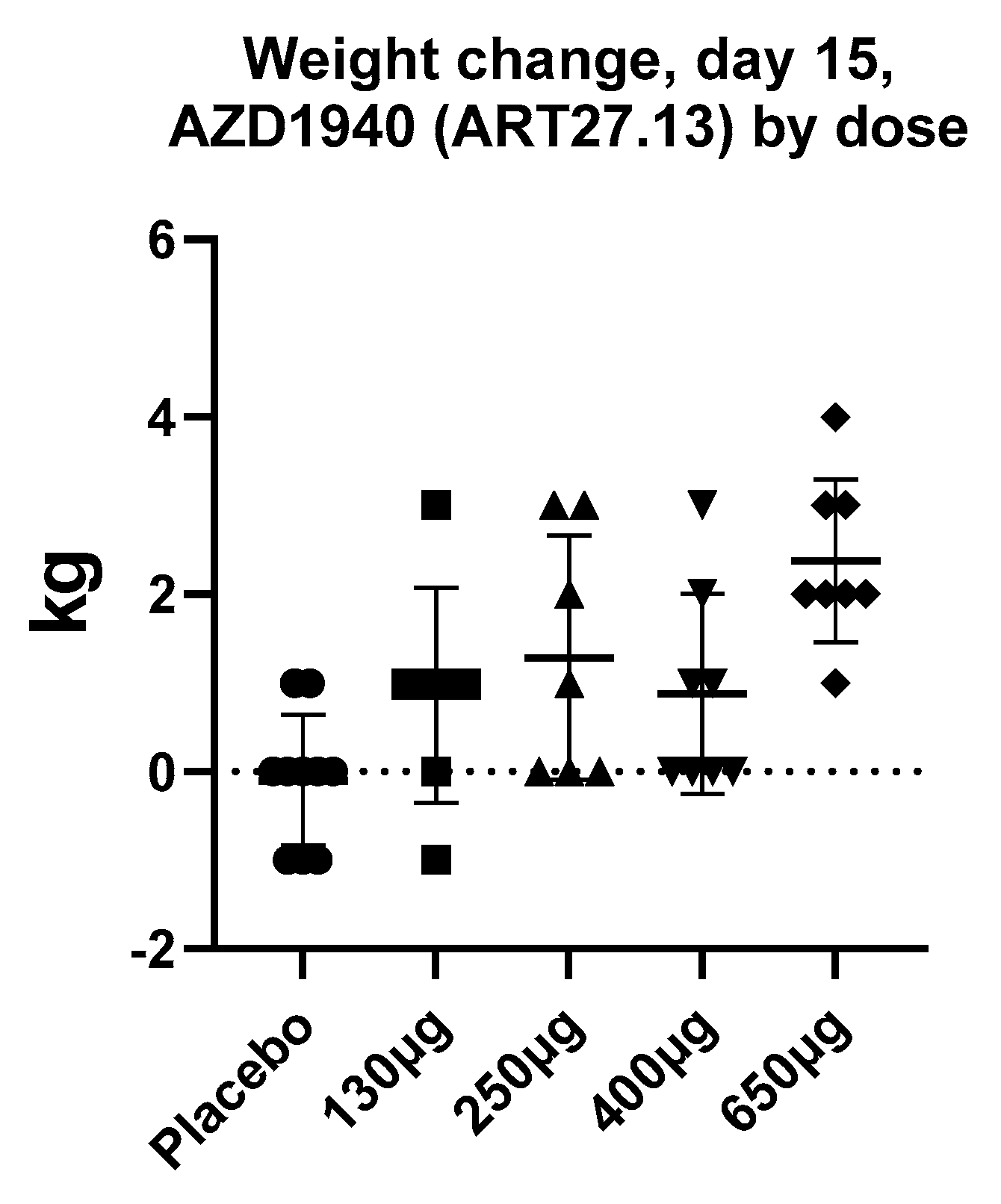
| Clinical research | INV-101 in Prader–Willi syndrome (PWS) and non-alcoholic steatohepatitis (NCT04531150) (Inversago Pharma) TM38837 in healthy subjects [104] (7TM Pharma) GFB-024 in diabetic nephropathy (Goldfinch Bio, NCT04880291) | AZD1940 in capsaicin-induced pain [124] and post-operative pain [125] ART27.13 (previously AZD1940) in Cancer anorexia (EudraCT NUMBER:2020-000464-27) (Artelo Biosciences) | URB937 is in the early stages of clinical development (Exxel Pharma) |
3.2. Peripherally Restricted CB1 Agonists
3.3. Peripherally Restricted Fatty Acid Amide Hydrolase (FAAH) Inhibitors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horn, H.; Böhme, B.; Dietrich, L.; Koch, M. Endocannabinoids in Body Weight Control. Pharmaceuticals 2018, 11, 55. [Google Scholar] [CrossRef]
- Sharkey, K.A.; Darmani, N.A.; Parker, L.A. Regulation of nausea and vomiting by cannabinoids and the endocannabinoid system. Eur. J. Pharmacol. 2014, 722, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.A.; Azariah, F.; Lavender, V.; Stoner, N.S.; Bettiol, S. Cannabinoids for nausea and vomiting in adults with cancer receiving chemotherapy. Cochrane Database Syst. Rev. 2015, 11, CD009464. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Tong, M.; Pan, H.; Li, D. Medical Cannabinoids for Cancer Cachexia: A Systematic Review and Meta-Analysis. BioMed Res. Int. 2019, 2019, 2864384. [Google Scholar] [CrossRef] [PubMed]
- Badowski, M.; Perez, S. Clinical utility of dronabinol in the treatment of weight loss associated with HIV and AIDS. HIV/AIDS-Res. Palliat. Care 2016, 8, 37–45. [Google Scholar] [CrossRef][Green Version]
- Struwe, M.; Kaempfer, S.H.; Geiger, C.J.; Pavia, A.T.; Plasse, T.F.; Shepard, K.V.; Ries, K.; Evans, T.G. Effect of Dronabinol on Nutritional Status in HIV Infection. Ann. Pharmacother. 1993, 27, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Andries, A.; Frystyk, J.; Flyvbjerg, A.; Stoving, R. Dronabinol in severe, enduring anorexia nervosa: A randomized controlled trial. Int. J. Eat. Disord. 2013, 47, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Andries, A.; Frystyk, J.; Flyvbjerg, A.; Støving, R.K. Changes in IGF-I, urinary free cortisol and adipokines during dronabinol therapy in anorexia nervosa: Results from a randomised, controlled trial. Growth Horm. IGF Res. 2015, 25, 247–252. [Google Scholar] [CrossRef]
- Volicer, L.; Stelly, M.; Morris, J.; McLaughlin, J.; Volicer, B.J. Effects of dronabinol on anorexia and disturbed behavior in patients with Alzheimer’s disease. Int. J. Geriatr. Psychiatry 1997, 12, 913–919. [Google Scholar] [CrossRef]
- Thomas, B.F.; Zhang, Y. Overcoming the Psychiatric Side Effects of the Cannabinoid CB1 Receptor Antagonists: Current Approaches for Therapeutics Development. Curr. Top. Med. Chem. 2019, 19, 1418–1435. [Google Scholar] [CrossRef]
- Christensen, R.; Kristensen, P.K.; Bartels, E.M.; Bliddal, H.; Astrup, A. Efficacy and safety of the weight-loss drug rimonabant: A meta-analysis of randomised trials. Lancet 2007, 370, 1706–1713. [Google Scholar] [CrossRef]
- Christopoulou, F.D.; Kiortsis, D.N. An overview of the metabolic effects of rimonabant in randomized controlled trials: Potential for other cannabinoid 1 receptor blockers in obesity. J. Clin. Pharm. Ther. 2011, 36, 10–18. [Google Scholar] [CrossRef]
- Panagis, G.; Mackey, B.; Vlachou, S. Cannabinoid Regulation of Brain Reward Processing with an Emphasis on the Role of CB1 Receptors: A Step Back into the Future. Front. Psychiatry 2014, 5, 92. [Google Scholar] [CrossRef]
- Johansson, K.; Neovius, M.; DeSantis, S.M.; Rössner, S. Discontinuation due to adverse events in randomized trials of orlistat, sibutramine and rimonabant: A meta-analysis. Obes. Rev. 2009, 10, 564–575. [Google Scholar] [CrossRef]
- Engeli, S. Central and Peripheral Cannabinoid Receptors as Therapeutic Targets in the Control of Food Intake and Body Weight. Cytochrome P450 2012, 209, 357–381. [Google Scholar] [CrossRef]
- De Azua, I.R.; Lutz, B. Multiple endocannabinoid-mediated mechanisms in the regulation of energy homeostasis in brain and peripheral tissues. Cell. Mol. Life Sci. 2019, 76, 1341–1363. [Google Scholar] [CrossRef] [PubMed]
- Cinar, R.; Iyer, M.R.; Kunos, G. The therapeutic potential of second and third generation CB1R antagonists. Pharmacol. Ther. 2020, 208, 107477. [Google Scholar] [CrossRef] [PubMed]
- Quarta, C.; Cota, D. Anti-obesity therapy with peripheral CB1 blockers: From promise to safe(?) practice. Int. J. Obes. 2020, 44, 2179–2193. [Google Scholar] [CrossRef]
- Koch, M. Cannabinoid Receptor Signaling in Central Regulation of Feeding Behavior: A Mini-Review. Front. Neurosci. 2017, 11, 293. [Google Scholar] [CrossRef]
- Mendizabal-Zubiaga, J.; Melser, S.; Bénard, G.; Ramos-Uriarte, A.; Reguero, L.; Arrabal, S.; Elezgarai, I.; Gerrikagoitia, I.; Suárez, J.; De Fonseca, F.R.; et al. Cannabinoid CB1 Receptors Are Localized in Striated Muscle Mitochondria and Regulate Mitochondrial Respiration. Front. Physiol. 2016, 7, 476. [Google Scholar] [CrossRef] [PubMed]
- De Azua, I.R.; Mancini, G.; Srivastava, R.K.; Rey, A.A.; Cardinal, P.; Tedesco, L.; Zingaretti, C.M.; Sassmann, A.; Quarta, C.; Schwitter, C.; et al. Adipocyte cannabinoid receptor CB1 regulates energy homeostasis and alternatively activated macrophages. J. Clin. Investig. 2017, 127, 4148–4162. [Google Scholar] [CrossRef] [PubMed]
- Osei-Hyiaman, D.; Harvey-White, J.; Bátkai, S.; Kunos, G. The role of the endocannabinoid system in the control of energy homeostasis. Int. J. Obes. 2006, 30, S33–S38. [Google Scholar] [CrossRef] [PubMed]
- González-Mariscal, I.; Montoro, R.A.; O’Connel, J.F.; Kim, Y.; Gonzalez-Freire, M.; Liu, Q.; Alfaras, I.; Carlson, O.D.; Lehrmann, E.; Zhang, Y.; et al. Muscle cannabinoid 1 receptor regulates Il-6 and myostatin expression, governing physical performance and whole-body metabolism. FASEB J. 2019, 33, 5850–5863. [Google Scholar] [CrossRef] [PubMed]
- Avalos, B.; Argueta, D.; Perez, P.A.; Wiley, M.; Wood, C.; DiPatrizio, N.V. Cannabinoid CB1 Receptors in the Intestinal Epithelium Are Required for Acute Western-Diet Preferences in Mice. Nutrients 2020, 12, 2874. [Google Scholar] [CrossRef]
- Cavalcante, M.L.D.S.; Silva, M.S.; Cavalcante, A.K.M.; Santos, R.D.O.; Nunes, D.D.T.; Busquets, S.; Argiles, J.M.; Seelaender, M.; Neto, E.M.D.M.; Dos Santos, A.A.; et al. Win 55,212-2, atenolol and subdiaphragmatic vagotomy prevent acceleration of gastric emptying induced by cachexia via Yoshida-AH-130 cells in rats. Eur. J. Pharmacol. 2020, 877, 173087. [Google Scholar] [CrossRef] [PubMed]
- Engeli, S. Dysregulation of the Endocannabinoid System in Obesity. J. Neuroendocr. 2008, 20, 110–115. [Google Scholar] [CrossRef]
- Côté, M.; Matias, I.; Lemieux, I.; Petrosino, S.; Alméras, N.; Després, J.-P.; Di Marzo, V. Circulating endocannabinoid levels, abdominal adiposity and related cardiometabolic risk factors in obese men. Int. J. Obes. 2007, 31, 692–699. [Google Scholar] [CrossRef]
- Abdulnour, J.; Yasari, S.; Rabasa-Lhoret, R.; Faraj, M.; Petrosino, S.; Piscitelli, F.; Homme, D.P.; Di Marzo, V. Circulating endocannabinoids in insulin sensitive vs. Insulin resistant obese postmenopausal women. A MONET group study. Obesity 2014, 22, 211–216. [Google Scholar] [CrossRef]
- Silvestri, C.; Di Marzo, V. Second generation CB1 receptor blockers and other inhibitors of peripheral endocannabinoid overactivity and the rationale of their use against metabolic disorders. Expert Opin. Investig. Drugs 2012, 21, 1309–1322. [Google Scholar] [CrossRef]
- Nabilone. LiverTox: Clinical and Research Information on Drug-Induced Liver Injury. Available online: https://pubmed.ncbi.nlm.nih.gov/31643176/ (accessed on 10 September 2021).
- Eckardt, K.; Sell, H.; Taube, A.; Koenen, M.; Platzbecker, B.; Cramer, A.; Horrighs, A.; Lehtonen, M.; Tennagels, N.; Eckel, J. Cannabinoid type 1 receptors in human skeletal muscle cells participate in the negative crosstalk between fat and muscle. Diabetology 2009, 52, 664–674. [Google Scholar] [CrossRef]
- Cavuoto, P.; McAinch, A.; Hatzinikolas, G.; Cameron-Smith, D.; Wittert, G. Effects of cannabinoid receptors on skeletal muscle oxidative pathways. Mol. Cell. Endocrinol. 2007, 267, 63–69. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Silvestri, C.; Mazzarella, E.; Martella, A.; Calvigioni, D.; Piscitelli, F.; Ambrosino, P.; Petrosino, S.; Czifra, G.; Biro, T.; et al. The endocannabinoid 2-AG controls skeletal muscle cell differentiation via CB1 receptor-dependent inhibition of Kv7 channels. Proc. Natl. Acad. Sci. 2014, 111, E2472–E2481. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Pagano, E.; Guardiola, O.; Adinolfi, S.; Saccone, V.; Consalvi, S.; Piscitelli, F.; Gazzerro, E.; Busetto, G.; Carrella, D.; et al. Genetic and pharmacological regulation of the endocannabinoid CB1 receptor in Duchenne muscular dystrophy. Nat. Commun. 2018, 9, 3950. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, R.; Niki, M.; Jyotaki, M.; Sanematsu, K.; Shigemura, N.; Ninomiya, Y. Modulation of sweet responses of taste receptor cells. Semin. Cell Dev. Biol. 2013, 24, 226–231. [Google Scholar] [CrossRef]
- Pazos, M.R.; Tolón, R.M.; Benito, C.; Rodríguez, C.F.; Gorgojo, J.J.; Nevado, M.; Álvarez, M.; Arias, F.; Almodóvar, F.; Fernández, M.T.P.; et al. Cannabinoid CB1Receptors Are Expressed by Parietal Cells of the Human Gastric Mucosa. J. Histochem. Cytochem. 2008, 56, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Adami, M.; Frati, P.; Bertini, S.; Kulkarni-Narla, A.; Brown, D.; De Caro, G.; Coruzzi, G.; Soldani, G. Gastric antisecretory role and immunohistochemical localization of cannabinoid receptors in the rat stomach. Br. J. Pharmacol. 2002, 135, 1598–1606. [Google Scholar] [CrossRef]
- Abalo, R.; Cabezos, P.A.; Vera, G.; Lopez-Miranda, V.; Herradón, E.; Martín-Fontelles, M.I. Cannabinoid-induced delayed gastric emptying is selectively increased upon intermittent administration in the rat: Role of CB1 receptors. Neurogastroenterol. Motil. 2011, 23, 457-e177. [Google Scholar] [CrossRef] [PubMed]
- Christie, S.; O’Rielly, R.; Li, H.; Wittert, G.; Page, A.J. Biphasic effects of methanandamide on murine gastric vagal afferent mechanosensitivity. J. Physiol. 2019, 598, 139–150. [Google Scholar] [CrossRef]
- Senin, L.L.; Al-Massadi, O.; Folgueira, C.; Castelao, C.; Pardo, M.; Barja-Fernandez, S.; Roca-Rivada, A.; Amil, M.; Crujeiras, A.B.; García-Caballero, T.; et al. The Gastric CB1 Receptor Modulates Ghrelin Production through the mTOR Pathway to Regulate Food Intake. PLoS ONE 2013, 8, e80339. [Google Scholar] [CrossRef] [PubMed]
- Argueta, D.; Perez, P.A.; Makriyannis, A.; DiPatrizio, N.V. Cannabinoid CB1 Receptors Inhibit Gut-Brain Satiation Signaling in Diet-Induced Obesity. Front. Physiol. 2019, 10, 704. [Google Scholar] [CrossRef]
- Cluny, N.L.; Keenan, C.M.; Duncan, M.; Fox, A.; Lutz, B.; Sharkey, K.A. Naphthalen-1-yl-(4-pentyloxynaphthalen-1-yl)methanone (SAB378), a Peripherally Restricted Cannabinoid CB1/CB2 Receptor Agonist, Inhibits Gastrointestinal Motility but Has No Effect on Experimental Colitis in Mice. J. Pharmacol. Exp. Ther. 2010, 334, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Keenan, C.M.; Storr, M.A.; Thakur, G.A.; Wood, J.T.; Wager-Miller, J.; Straiker, A.; Eno, M.R.; Nikas, S.P.; Bashashati, M.; Hu, H.; et al. AM841, a covalent cannabinoid ligand, powerfully slows gastrointestinal motility in normal and stressed mice in a peripherally restricted manner. Br. J. Pharmacol. 2015, 172, 2406–2418. [Google Scholar] [CrossRef] [PubMed]
- Karwad, M.; Couch, D.; Wright, K.; Tufarelli, C.; Larvin, M.; Lund, J.; O’Sullivan, S. Endocannabinoids and endocannabinoid-like compounds modulate hypoxia-induced permeability in CaCo-2 cells via CB1, TRPV1, and PPARα. Biochem. Pharmacol. 2019, 168, 465–472. [Google Scholar] [CrossRef]
- Argueta, D.; DiPatrizio, N.V. Peripheral endocannabinoid signaling controls hyperphagia in western diet-induced obesity. Physiol. Behav. 2017, 171, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Burdyga, G.; Lal, S.; Varro, A.; Dimaline, R.; Thompson, D.G.; Dockray, G.J. Expression of Cannabinoid CB1 Receptors by Vagal Afferent Neurons Is Inhibited by Cholecystokinin. J. Neurosci. 2004, 24, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Burdyga, G.; Varró, A.; Dimaline, R.; Thompson, D.G.; Dockray, G.J. Expression of cannabinoid CB1 receptors by vagal afferent neurons: Kinetics and role in influencing neurochemical phenotype. Am. J. Physiol. Liver Physiol. 2010, 299, G63–G69. [Google Scholar] [CrossRef]
- Gómez, R.; Navarro, M.; Ferrer, B.; Trigo, J.M.; Bilbao, A.; Del Arco, I.; Cippitelli, A.; Nava, F.A.; Piomelli, D.; De Fonseca, F.R. A Peripheral Mechanism for CB1 Cannabinoid Receptor-Dependent Modulation of Feeding. J. Neurosci. 2002, 22, 9612–9617. [Google Scholar] [CrossRef]
- Mehrpouya-Bahrami, P.; Chitrala, K.N.; Ganewatta, M.S.; Tang, C.; Murphy, E.A.; Enos, R.T.; Velazquez, K.T.; McCellan, J.; Nagarkatti, M.; Nagarkatti, P. Blockade of CB1 cannabinoid receptor alters gut microbiota and attenuates inflammation and diet-induced obesity. Sci. Rep. 2017, 7, 15645. [Google Scholar] [CrossRef] [PubMed]
- Cluny, N.L.; Keenan, C.M.; Reimer, R.A.; Le Foll, B.; Sharkey, K.A. Prevention of Diet-Induced Obesity Effects on Body Weight and Gut Microbiota in Mice Treated Chronically with Δ9-Tetrahydrocannabinol. PLoS ONE 2015, 10, e0144270. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Gioacchini, G.; Pengo, G.; Suchodolski, J.S.; Jergens, A.E.; Allenspach, K.; Gavazza, A.; Scarpona, S.; Berardi, S.; Galosi, L.; et al. Enterocolic increase of cannabinoid receptor type 1 and type 2 and clinical improvement after probiotic administration in dogs with chronic signs of colonic dysmotility without mucosal inflammatory changes. Neurogastroenterol. Motil. 2019, 32, e13717. [Google Scholar] [CrossRef] [PubMed]
- Muccioli, G.G.; Naslain, D.; Bäckhed, F.; Reigstad, C.S.; Lambert, D.M.; Delzenne, N.; Cani, P.D. The endocannabinoid system links gut microbiota to adipogenesis. Mol. Syst. Biol. 2010, 6, 392. [Google Scholar] [CrossRef]
- Bensaid, M.; Gary-Bobo, M.; Esclangon, A.; Maffrand, J.P.; Le Fur, G.; Oury-Donat, F.; Soubrié, P. The Cannabinoid CB1Receptor Antagonist SR141716 Increases Acrp30 mRNA Expression in Adipose Tissue of Obese fa/fa Rats and in Cultured Adipocyte Cells. Mol. Pharmacol. 2003, 63, 908–914. [Google Scholar] [CrossRef]
- Matias, I.; Gonthier, M.-P.; Orlando, P.; Martiadis, V.; De Petrocellis, L.; Cervino, C.; Petrosino, S.; Hoareau, L.; Festy, F.; Pasquali, R.; et al. Regulation, Function, and Dysregulation of Endocannabinoids in Models of Adipose and β-Pancreatic Cells and in Obesity and Hyperglycemia. J. Clin. Endocrinol. Metab. 2006, 91, 3171–3180. [Google Scholar] [CrossRef]
- Cota, D.; Marsicano, G.; Tschöp, M.; Grübler, Y.; Flachskamm, C.; Schubert, M.; Auer, D.; Yassouridis, A.; Thöne-Reineke, C.; Ortmann, S.; et al. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J. Clin. Investig. 2003, 112, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Maury, E.; Rycken, L.; Gérard, J.; Noël, L.; Detry, R.; Navez, B.; Brichard, S.M. Endocannabinoids regulate adipokine production and the immune balance of omental adipose tissue in human obesity. Int. J. Obes. 2012, 37, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, L.; Valerio, A.; Dossena, M.; Cardile, A.; Ragni, M.; Pagano, C.; Pagotto, U.; Carruba, M.O.; Vettor, R.; Nisoli, E. Cannabinoid Receptor Stimulation Impairs Mitochondrial Biogenesis in Mouse White Adipose Tissue, Muscle, and Liver: The Role of eNOS, p38 MAPK, and AMPK Pathways. Diabetes 2010, 59, 2826–2836. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, L.; Valerio, A.; Cervino, C.; Cardile, A.; Pagano, C.; Vettor, R.; Pasquali, R.; Carruba, M.O.; Marsicano, G.; Lutz, B.; et al. Cannabinoid Type 1 Receptor Blockade Promotes Mitochondrial Biogenesis Through Endothelial Nitric Oxide Synthase Expression in White Adipocytes. Diabetes 2008, 57, 2028–2036. [Google Scholar] [CrossRef]
- Lahesmaa, M.; Eriksson, O.; Gnad, T.; Oikonen, V.; Bucci, M.; Hirvonen, J.; Koskensalo, K.; Teuho, J.; Niemi, T.; Taittonen, M.; et al. Cannabinoid Type 1 Receptors Are Upregulated During Acute Activation of Brown Adipose Tissue. Diabetes 2018, 67, 1226–1236. [Google Scholar] [CrossRef]
- Krott, L.M.; Piscitelli, F.; Heine, M.; Borrino, S.; Scheja, L.; Silvestri, C.; Heeren, J.; Di Marzo, V. Endocannabinoid regulation in white and brown adipose tissue following thermogenic activation. J. Lipid Res. 2016, 57, 464–473. [Google Scholar] [CrossRef]
- Bajzer, M.; Olivieri, M.; Haas, M.K.; Pfluger, P.T.; Magrisso, I.J.; Foster, M.T.; Tschöp, M.H.; Krawczewski-Carhuatanta, K.A.; Cota, D.; Obici, S. Cannabinoid receptor 1 (CB1) antagonism enhances glucose utilisation and activates brown adipose tissue in diet-induced obese mice. Diabetology 2011, 54, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Osei-Hyiaman, D.; DePetrillo, M.; Pacher, P.; Liu, J.; Radaeva, S.; Bátkai, S.; Harvey-White, J.; Mackie, K.; Offertáler, L.; Wang, L.; et al. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J. Clin. Investig. 2005, 115, 1298–1305. [Google Scholar] [CrossRef]
- Chanda, D.; Kim, D.-K.; Li, T.; Kim, Y.-H.; Koo, S.-H.; Lee, C.-H.; Chiang, J.; Choi, H.-S. Cannabinoid Receptor Type 1 (CB1R) Signaling Regulates Hepatic Gluconeogenesis via Induction of Endoplasmic Reticulum-bound Transcription Factor cAMP-responsive Element-binding Protein H (CREBH) in Primary Hepatocytes. J. Biol. Chem. 2011, 286, 27971–27979. [Google Scholar] [CrossRef]
- Mukhopadhyay, B.; Cinar, R.; Yin, S.; Liu, J.; Tam, J.; Godlewski, G.; Harvey-White, J.; Mordi, I.; Cravatt, B.F.; Lotersztajn, S.; et al. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via cannabinoid type 1 (CB1) receptors in the regenerating liver. Proc. Natl. Acad. Sci. USA 2011, 108, 6323–6328. [Google Scholar] [CrossRef]
- Osei-Hyiaman, U.; Liu, J.; Zhou, L.; Godlewski, G.; Harvey-White, J.; Jeong, W.-I.; Bátkai, S.; Marsicano, G.; Lutz, B.; Buettner, C.; et al. Hepatic CB1 receptor is required for development of diet-induced steatosis, dyslipidemia, and insulin and leptin resistance in mice. J. Clin. Investig. 2008, 118, 3160–3169. [Google Scholar] [CrossRef]
- Li, C.; Jones, P.; Persaud, S. Cannabinoid Receptors are Coupled to Stimulation of Insulin Secretion from Mouse MIN6 β-cells. Cell. Physiol. Biochem. 2010, 26, 187–196. [Google Scholar] [CrossRef]
- Malenczyk, K.; Jazurek, M.; Keimpema, E.; Silvestri, C.; Janikiewicz, J.; Mackie, K.; Di Marzo, V.; Redowicz, M.J.; Harkany, T.; Dobrzyn, A. CB1 Cannabinoid Receptors Couple to Focal Adhesion Kinase to Control Insulin Release. J. Biol. Chem. 2013, 288, 32685–32699. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Doyle, M.E.; Liu, Z.; Lao, Q.; Shin, Y.-K.; Carlson, O.D.; Kim, H.S.; Thomas, S.; Napora, J.K.; Lee, E.K.; et al. Cannabinoids Inhibit Insulin Receptor Signaling in Pancreatic -Cells. Diabetes 2011, 60, 1198–1209. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Lao, Q.; Shin, Y.-K.; Carlson, O.D.; Lee, E.K.; Gorospe, M.; Kulkarni, R.N.; Egan, J.M. Cannabinoids Induce Pancreatic—Cell Death by Directly Inhibiting Insulin Receptor Activation. Sci. Signal. 2012, 5, ra23. [Google Scholar] [CrossRef]
- Oláh, T.; Bodnár, D.; Tóth, A.; Vincze, J.; Fodor, J.; Reischl, B.; Kovács, A.; Ruzsnavszky, O.; Dienes, B.; Szentesi, P.; et al. Cannabinoid signalling inhibits sarcoplasmic Ca2+release and regulates excitation-contraction coupling in mammalian skeletal muscle. J. Physiol. 2016, 594, 7381–7398. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Szanda, G.; Drori, A.; Liu, Z.; Cinar, R.; Kashiwaya, Y.; Reitman, M.L.; Kunos, G. Peripheral cannabinoid-1 receptor blockade restores hypothalamic leptin signaling. Mol. Metab. 2017, 6, 1113–1125. [Google Scholar] [CrossRef]
- Alen, F.; Crespo, I.; Ramírez-López, M.T.; Jagerovic, N.; Goya, P.; De Fonseca, F.R.; De Heras, R.G.; Orio, L. Ghrelin-Induced Orexigenic Effect in Rats Depends on the Metabolic Status and Is Counteracted by Peripheral CB1 Receptor Antagonism. PLoS ONE 2013, 8, e60918. [Google Scholar] [CrossRef] [PubMed]
- Farokhnia, M.; McDiarmid, G.R.; Newmeyer, M.; Munjal, V.; Abulseoud, O.A.; Huestis, M.A.; Leggio, L. Effects of oral, smoked, and vaporized cannabis on endocrine pathways related to appetite and metabolism: A randomized, double-blind, placebo-controlled, human laboratory study. Transl. Psychiatry 2020, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Weltens, N.; Depoortere, I.; Tack, J.; Van Oudenhove, L. Effect of acute Δ9-tetrahydrocannabinol administration on subjective and metabolic hormone responses to food stimuli and food intake in healthy humans: A randomized, placebo-controlled study. Am. J. Clin. Nutr. 2019, 109, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Arai, H.; Inui, A. The regulatory approval of anamorelin for treatment of cachexia in patients with non-small cell lung cancer, gastric cancer, pancreatic cancer, and colorectal cancer in Japan: Facts and numbers. J. Cachex-Sarcopenia Muscle 2021, 12, 14–16. [Google Scholar] [CrossRef] [PubMed]
- DiPatrizio, N. Endocannabinoids and the Gut-Brain Control of Food Intake and Obesity. Nutrition 2021, 13, 1214. [Google Scholar] [CrossRef]
- Christie, S.; O’Rielly, R.; Li, H.; Nunez-Salces, M.; Wittert, G.A.; Page, A.J. Modulatory effect of methanandamide on gastric vagal afferent satiety signals depends on nutritional status. J. Physiol. 2020, 598, 2169–2182. [Google Scholar] [CrossRef]
- DiPatrizio, N.V.; Astarita, G.; Schwartz, G.; Li, X.; Piomelli, D. Endocannabinoid signal in the gut controls dietary fat intake. Proc. Natl. Acad. Sci. USA 2011, 108, 12904–12908. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Di Marzo, V. The gut microbiome, endocannabinoids and metabolic disorders. J. Endocrinol. 2021, 248, R83–R97. [Google Scholar] [CrossRef]
- Manca, C.; Boubertakh, B.; Leblanc, N.; Deschênes, T.; Lacroix, S.; Martin, C.; Houde, A.; Veilleux, A.; Flamand, N.; Muccioli, G.G.; et al. Germ-free mice exhibit profound gut microbiota-dependent alterations of intestinal endocannabinoidome signaling. J. Lipid Res. 2020, 61, 70–85. [Google Scholar] [CrossRef]
- LoVerme, J.; Duranti, A.; Tontini, A.; Spadoni, G.; Mor, M.; Rivara, S.; Stella, N.; Xu, C.; Tarzia, G.; Piomelli, D. Synthesis and characterization of a peripherally restricted CB1 cannabinoid antagonist, URB447, that reduces feeding and body-weight gain in mice. Bioorganic Med. Chem. Lett. 2009, 19, 639–643. [Google Scholar] [CrossRef]
- Cluny, N.; Vemuri, V.; Chambers, A.; Limebeer, C.; Bedard, H.; Wood, J.; Lutz, B.; Zimmer, A.; Parker, L.; Makriyannis, A.; et al. A novel peripherally restricted cannabinoid receptor antagonist, AM6545, reduces food intake and body weight, but does not cause malaise, in rodents. Br. J. Pharmacol. 2010, 161, 629–642. [Google Scholar] [CrossRef]
- Chen, W.; Shui, F.; Liu, C.; Zhou, X.; Li, W.; Zheng, Z.; Fu, W.; Wang, L. Novel Peripherally Restricted Cannabinoid 1 Receptor Selective Antagonist TXX-522 with Prominent Weight-Loss Efficacy in Diet Induced Obese Mice. Front. Pharmacol. 2017, 8, 707. [Google Scholar] [CrossRef]
- Alonso, M.; Serrano, A.; Vida, M.; Crespillo, A.; Hernandez-Folgado, L.; Jagerovic, N.; Goya, P.; Reyes-Cabello, C.; Perez-Valero, V.; Decara, J.; et al. Anti-obesity efficacy of LH-21, a cannabinoid CB1 receptor antagonist with poor brain penetration, in diet-induced obese rats. Br. J. Pharmacol. 2012, 165, 2274–2291. [Google Scholar] [CrossRef] [PubMed]
- Chorvat, R.J.; Berbaum, J.; Seriacki, K.; McElroy, J.F. JD-5006 and JD-5037: Peripherally restricted (PR) cannabinoid-1 receptor blockers related to SLV-319 (Ibipinabant) as metabolic disorder therapeutics devoid of CNS liabilities. Bioorg. Med. Chem. Lett. 2012, 22, 6173–6180. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Vemuri, V.K.; Liu, J.; Bátkai, S.; Mukhopadhyay, B.; Godlewski, G.; Osei-Hyiaman, D.; Ohnuma, S.; Ambudkar, S.V.; Pickel, J.; et al. Peripheral CB1 cannabinoid receptor blockade improves cardiometabolic risk in mouse models of obesity. J. Clin. Investig. 2010, 120, 2953–2966. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Cinar, R.; Liu, J.; Godlewski, G.; Wesley, D.; Jourdan, T.; Szanda, G.; Mukhopadhyay, B.; Chedester, L.; Liow, J.-S.; et al. Peripheral Cannabinoid-1 Receptor Inverse Agonism Reduces Obesity by Reversing Leptin Resistance. Cell Metab. 2012, 16, 167–179. [Google Scholar] [CrossRef]
- Knani, I.; Earley, B.J.; Udi, S.; Nemirovski, A.; Hadar, R.; Gammal, A.; Cinar, R.; Hirsch, H.J.; Pollak, Y.; Gross, I.; et al. Targeting the endocannabinoid/CB1 receptor system for treating obesity in Prader–Willi syndrome. Mol. Metab. 2016, 5, 1187–1199. [Google Scholar] [CrossRef]
- Ding, Y.; Qiu, Y.; Jing, L.; Thorn, D.A.; Zhang, Y.; Li, J.-X. Behavioral effects of the cannabinoid CB1receptor allosteric modulator ORG27569 in rats. Pharmacol. Res. Perspect. 2014, 2, e00069. [Google Scholar] [CrossRef]
- Ferrante, C.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di Nisio, C.; Martinotti, S.; Mollica, A.; Macedonio, G.; Stefanucci, A.; Dvorácskó, S.; et al. Anorexigenic effects induced by RVD-hemopressin(α) administration. Pharmacol. Rep. 2017, 69, 1402–1407. [Google Scholar] [CrossRef]
- Horswill, J.G.; Bali, U.; Shaaban, S.; Keily, J.F.; Jeevaratnam, P.; Babbs, A.J.; Reynet, C.; In, P.W.K. PSNCBAM-1, a novel allosteric antagonist at cannabinoid CB1 receptors with hypophagic effects in rats. Br. J. Pharmacol. 2007, 152, 805–814. [Google Scholar] [CrossRef]
- Barutta, F.; Grimaldi, S.; Gambino, R.; Vemuri, K.; Makriyannis, A.; Annaratone, L.; Di Marzo, V.; Bruno, G.; Gruden, G. Dual therapy targeting the endocannabinoid system prevents experimental diabetic nephropathy. Nephrol. Dial. Transplant. 2017, 32, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Barutta, F.; Bellini, S.; Mastrocola, R.; Gambino, R.; Piscitelli, F.; Di Marzo, V.; Corbetta, B.; Vemuri, V.; Makriyannis, A.; Annaratone, L.; et al. Reversal of albuminuria by combined AM6545 and perindopril therapy in experimental diabetic nephropathy. Br. J. Pharmacol. 2018, 175, 4371–4385. [Google Scholar] [CrossRef]
- Amato, G.; Manke, A.; Harris, D.L.; Wiethe, R.W.; Vasukuttan, V.; Snyder, R.W.; Lefever, T.W.; Cortes, R.; Zhang, Y.; Wang, S.; et al. Blocking Alcoholic Steatosis in Mice with a Peripherally Restricted Purine Antagonist of the Type 1 Cannabinoid Receptor. J. Med. Chem. 2018, 61, 4370–4385. [Google Scholar] [CrossRef]
- Azar, S.; Udi, S.; Drori, A.; Hadar, R.; Nemirovski, A.; Vemuri, K.V.; Miller, M.; Sherill-Rofe, D.; Arad, Y.; Gur-Wahnon, D.; et al. Reversal of diet-induced hepatic steatosis by peripheral CB1 receptor blockade in mice is p53/miRNA-22/SIRT1/PPARα dependent. Mol. Metab. 2020, 42, 101087. [Google Scholar] [CrossRef] [PubMed]
- Kale, V.P.; Gibbs, S.; Taylor, J.A.; Zmarowski, A.; Novak, J.; Patton, K.; Sparrow, B.; Gorospe, J.; Anand, S.; Cinar, R.; et al. Preclinical toxicity evaluation of JD5037, a peripherally restricted CB1 receptor inverse agonist, in rats and dogs for treatment of nonalcoholic steatohepatitis. Regul. Toxicol. Pharmacol. 2019, 109, 104483. [Google Scholar] [CrossRef]
- Bronova, I.; Smith, B.; Aydogan, B.; Weichselbaum, R.R.; Vemuri, K.; Erdélyi, K.; Makriyannis, A.; Pacher, P.; Berdyshev, E.V. Protection from Radiation-Induced Pulmonary Fibrosis by Peripheral Targeting of Cannabinoid Receptor-1. Am. J. Respir. Cell Mol. Biol. 2015, 53, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Cinar, R.; Gochuico, B.R.; Iyer, M.R.; Jourdan, T.; Yokoyama, T.; Park, J.K.; Coffey, N.; Pri-Chen, H.; Szanda, G.; Liu, Z.; et al. Cannabinoid CB1 receptor overactivity contributes to the pathogenesis of idiopathic pulmonary fibrosis. JCI Insight 2017, 2, e92281. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, G.; Cinar, R.; Coffey, N.; Liu, J.; Jourdan, T.; Mukhopadhyay, B.; Chedester, L.; Liu, Z.; Osei-Hyiaman, D.; Iyer, M.R.; et al. Targeting Peripheral CB1 Receptors Reduces Ethanol Intake via a Gut-Brain Axis. Cell Metab. 2019, 29, 1320–1333.e8. [Google Scholar] [CrossRef]
- Santos-Molina, L.; Herrerias, A.; Zawatsky, C.N.; Gunduz-Cinar, O.; Cinar, R.; Iyer, M.R.; Wood, C.M.; Lin, Y.; Gao, B.; Kunos, G.; et al. Effects of a Peripherally Restricted Hybrid Inhibitor of CB1 Receptors and iNOS on Alcohol Drinking Behavior and Alcohol-Induced Endotoxemia. Molecules 2021, 26, 5089. [Google Scholar] [CrossRef]
- Udi, S.; Hinden, L.; Ahmad, M.; Drori, A.; Iyer, M.R.; Cinar, R.; Herman-Edelstein, M.; Tam, J. Dual inhibition of cannabinoid CB 1 receptor and inducible NOS attenuates obesity-induced chronic kidney disease. Br. J. Pharmacol. 2020, 177, 110–127. [Google Scholar] [CrossRef]
- Cinar, R.; Iyer, M.R.; Liu, Z.; Cao, Z.; Jourdan, T.; Erdelyi, K.; Godlewski, G.; Szanda, G.; Liu, J.; Park, J.K.; et al. Hybrid inhibitor of peripheral cannabinoid-1 receptors and inducible nitric oxide synthase mitigates liver fibrosis. JCI Insight 2016, 1, e87336. [Google Scholar] [CrossRef] [PubMed]
- Zawatsky, C.N.; Park, J.K.; Abdalla, J.; Kunos, G.; Iyer, M.R.; Cinar, R. Peripheral Hybrid CB1R and iNOS Antagonist MRI-1867 Displays Anti-Fibrotic Efficacy in Bleomycin-Induced Skin Fibrosis. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Klumpers, L.E.; Fridberg, M.; De Kam, M.L.; Little, P.B.; Jensen, N.O.; Kleinloog, H.D.; Elling, C.E.; Van Gerven, J.M.A. Peripheral selectivity of the novel cannabinoid receptor antagonist TM38837 in healthy subjects. Br. J. Clin. Pharmacol. 2013, 76, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Cinar, R.; Godlewski, G.; Liu, J.; Tam, J.; Jourdan, T.; Mukhopadhyay, B.; Harvey-White, J.; Kunos, G. Hepatic cannabinoid-1 receptors mediate diet-induced insulin resistance by increasingde novosynthesis of long-chain ceramides. Hepatology 2014, 59, 143–153. [Google Scholar] [CrossRef]
- Cheng, Y.-X.; Pourashraf, M.; Luo, X.; Srivastava, S.; Walpole, C.; Salois, D.; St-Onge, S.; Payza, K.; Lessard, E.; Yu, X.H.; et al. γ-Carbolines: A novel class of cannabinoid agonists with high aqueous solubility and restricted CNS penetration. Bioorg. Med. Chem. Lett. 2012, 22, 1619–1624. [Google Scholar] [CrossRef]
- Yu, X.H.; Cao, C.Q.; Martino, G.; Puma, C.; Morinville, A.; St-Onge, S.; Lessard, É.; Perkins, M.N.; Laird, J.M. A peripherally restricted cannabinoid receptor agonist produces robust anti-nociceptive effects in rodent models of inflammatory and neuropathic pain. Pain 2010, 151, 337–344. [Google Scholar] [CrossRef]
- Dziadulewicz, E.K.; Bevan, S.J.; Brain, C.T.; Coote, P.R.; Culshaw, A.J.; Davis, A.J.; Edwards, L.; Fisher, A.J.; Fox, A.J.; Gentry, C.; et al. Naphthalen-1-yl-(4-pentyloxynaphthalen-1-yl)methanone: A Potent, Orally Bioavailable Human CB1/CB2 Dual Agonist with Antihyperalgesic Properties and Restricted Central Nervous System Penetration. J. Med. Chem. 2007, 50, 3851–3856. [Google Scholar] [CrossRef]
- Zhang, H.; Lund, D.M.; Ciccone, H.A.; Staatz, W.D.; Ibrahim, M.M.; Largent-Milnes, T.M.; Seltzman, H.H.; Spigelman, I.; Vanderah, T.W. Peripherally restricted cannabinoid 1 receptor agonist as a novel analgesic in cancer-induced bone pain. Pain 2018, 159, 1814–1823. [Google Scholar] [CrossRef]
- Mulpuri, Y.; Marty, V.N.; Munier, J.J.; Mackie, K.; Schmidt, B.L.; Seltzman, H.H.; Spigelman, I. Synthetic peripherally-restricted cannabinoid suppresses chemotherapy-induced peripheral neuropathy pain symptoms by CB1 receptor activation. Neuropharmacology 2018, 139, 85–97. [Google Scholar] [CrossRef]
- Yamamoto, T.; Mulpuri, Y.; Izraylev, M.; Li, Q.; Simonian, M.; Kramme, C.; Schmidt, B.L.; Seltzman, H.H.; Spigelman, I. Selective targeting of peripheral cannabinoid receptors prevents behavioral symptoms and sensitization of trigeminal neurons in mouse models of migraine and medication overuse headache. Pain 2021, 162, 2246–2262. [Google Scholar] [CrossRef]
- Pryce, G.; Visintin, C.C.; Ramagopalan, S.V.S.; Al-Izki, S.S.; De Faveri, L.; Nuamah, R.R.; Mein, C.; Montpetit, A.A.; Hardcastle, A.; Kooij, G.G.; et al. Control of spasticity in a multiple sclerosis model using central nervous system-excluded CB 1 cannabinoid receptor agonists. FASEB J. 2014, 28, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Rock, E.M.; Moreno-Sanz, G.; Limebeer, C.L.; Petrie, G.N.; Angelini, R.; Piomelli, D.A. Suppression of acute and anticipatory nausea by peripherally restricted fatty acid amide hydrolase inhibitor in animal models: Role of PPARα and CB1 receptors. Br. J. Pharmacol. 2017, 174, 3837–3847. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Lee, D.; Chowdhury, S.R.; Lu, P.; Kamboj, A.; Anderson, C.M.; Fernyhough, P.; Anderson, H.D. Activation of Cannabinoid Receptors Attenuates Endothelin-1–Induced Mitochondrial Dysfunction in Rat Ventricular Myocytes. J. Cardiovasc. Pharmacol. 2020, 75, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Clapper, J.R.; Moreno-Sanz, G.; Russo, R.; Guijarro, A.; Vacondio, F.; Duranti, A.; Tontini, A.; Sanchini, S.; Sciolino, N.R.; Spradley, J.M.; et al. Anandamide suppresses pain initiation through a peripheral endocannabinoid mechanism. Nat. Neurosci. 2010, 13, 1265–1270. [Google Scholar] [CrossRef]
- Slivicki, R.A.; Xu, Z.; Mali, S.; Hohmann, A.G. Brain permeant and impermeant inhibitors of fatty-acid amide hydrolase suppress the development and maintenance of paclitaxel-induced neuropathic pain without producing tolerance or physical dependence in vivo and synergize with paclitaxel to reduce tumor cell line viability in vitro. Pharmacol. Res. 2019, 142, 267–282. [Google Scholar] [CrossRef]
- Moreno-Sanz, G.; Sasso, O.; Guijarro, A.; Oluyemi, O.; Bertorelli, R.; Reggiani, A.; Piomelli, D. Pharmacological characterization of the peripheral FAAH inhibitor URB937 in female rodents: Interaction with the Abcg2 transporter in the blood-placenta barrier. Br. J. Pharmacol. 2012, 167, 1620–1628. [Google Scholar] [CrossRef]
- Sasso, O.; Bertorelli, R.; Bandiera, T.; Scarpelli, R.; Colombano, G.; Armirotti, A.; Moreno-Sanz, G.; Reggiani, A.; Piomelli, D. Peripheral FAAH inhibition causes profound antinociception and protects against indomethacin-induced gastric lesions. Pharmacol. Res. 2012, 65, 553–563. [Google Scholar] [CrossRef]
- Sasso, O.; Wagner, K.; Morisseau, C.; Inceoglu, B.; Hammock, B.D.; Piomelli, D. Peripheral FAAH and soluble epoxide hydrolase inhibitors are synergistically antinociceptive. Pharmacol. Res. 2015, 97, 7–15. [Google Scholar] [CrossRef]
- Greco, R.; Demartini, C.; Zanaboni, A.; Casini, I.; De Icco, R.; Reggiani, A.; Misto, A.; Piomelli, D.; Tassorelli, C. Characterization of the peripheral FAAH inhibitor, URB937, in animal models of acute and chronic migraine. Neurobiol. Dis. 2021, 147, 105157. [Google Scholar] [CrossRef]
- Greco, R.; Demartini, C.; Zanaboni, A.M.; Tumelero, E.; Reggiani, A.; Misto, A.; Piomelli, D.; Tassorelli, C. FAAH inhibition as a preventive treatment for migraine: A pre-clinical study. Neurobiol. Dis. 2020, 134, 104624. [Google Scholar] [CrossRef]
- Charrua, A.; Matos, R.; Oliveira, R.; Marczylo, T.; Nagy, I.; Cruz, F. Fatty acid amide hydrolase inhibition normalises bladder function and reduces pain through normalising the anandamide/palmitoylethanolamine ratio in the inflamed bladder of rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 393, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, N.; Gandaglia, G.; Hedlund, P.; Fujimura, T.; Fukuhara, H.; Montorsi, F.; Homma, Y.; Igawa, Y. URB937, a peripherally restricted inhibitor for fatty acid amide hydrolase, reduces prostaglandin E2-induced bladder overactivity and hyperactivity of bladder mechano-afferent nerve fibres in rats. BJU Int. 2015, 117, 821–828. [Google Scholar] [CrossRef]
- Kalliomäki, J.; Annas, P.; Huizar, K.; Clarke, C.; Zettergren, A.; Karlsten, R.; Segerdahl, M. Evaluation of the analgesic efficacy and psychoactive effects of AZD1940, a novel peripherally acting cannabinoid agonist, in human capsaicin-induced pain and hyperalgesia. Clin. Exp. Pharmacol. Physiol. 2013, 40, 212–218. [Google Scholar] [CrossRef]
- Kalliomäki, J.; Segerdahl, M.; Webster, L.; Reimfelt, A.; Huizar, K.; Annas, P.; Karlsten, R.; Quiding, H. Evaluation of the analgesic efficacy of AZD1940, a novel cannabinoid agonist, on post-operative pain after lower third molar surgical removal. Scand. J. Pain 2013, 4, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Ando, H.; Unno, S.; Kitagawa, J. Targeting Peripherally Restricted Cannabinoid Receptor 1, Cannabinoid Receptor 2, and Endocannabinoid-Degrading Enzymes for the Treatment of Neuropathic Pain Including Neuropathic Orofacial Pain. Int. J. Mol. Sci. 2020, 21, 1423. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Sullivan, S.E.; Yates, A.S.; Porter, R.K. The Peripheral Cannabinoid Receptor Type 1 (CB1) as a Molecular Target for Modulating Body Weight in Man. Molecules 2021, 26, 6178. https://doi.org/10.3390/molecules26206178
O’Sullivan SE, Yates AS, Porter RK. The Peripheral Cannabinoid Receptor Type 1 (CB1) as a Molecular Target for Modulating Body Weight in Man. Molecules. 2021; 26(20):6178. https://doi.org/10.3390/molecules26206178
Chicago/Turabian StyleO’Sullivan, Saoirse Elizabeth, Andrew S. Yates, and Richard K. Porter. 2021. "The Peripheral Cannabinoid Receptor Type 1 (CB1) as a Molecular Target for Modulating Body Weight in Man" Molecules 26, no. 20: 6178. https://doi.org/10.3390/molecules26206178
APA StyleO’Sullivan, S. E., Yates, A. S., & Porter, R. K. (2021). The Peripheral Cannabinoid Receptor Type 1 (CB1) as a Molecular Target for Modulating Body Weight in Man. Molecules, 26(20), 6178. https://doi.org/10.3390/molecules26206178