Effects of Hexane on Protein Profile, Solubility and Foaming Properties of Defatted Proteins Extracted from Tenebrio molitor Larvae



Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Materials
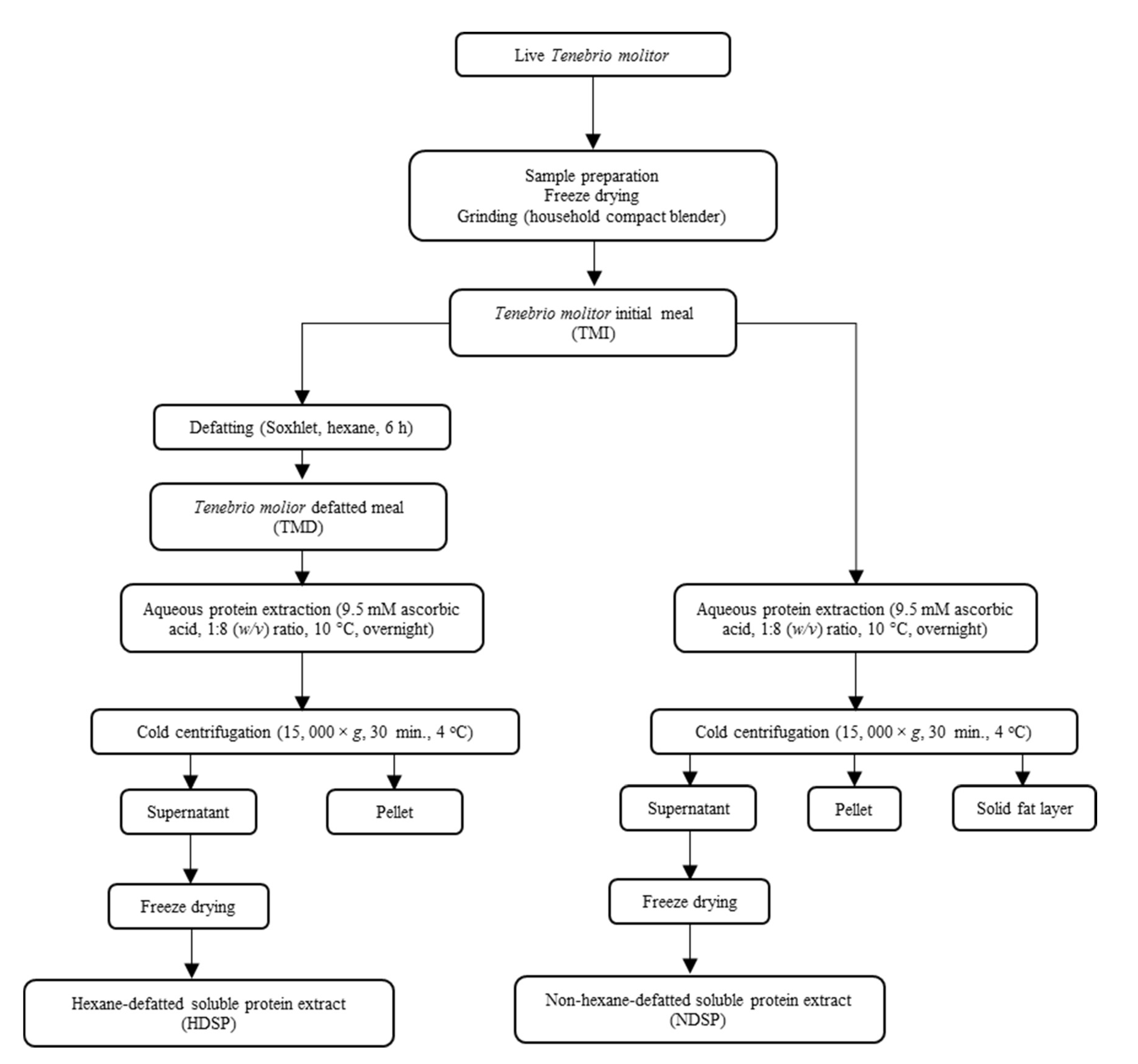
2.2. Methods
2.2.1. Lipid Extraction
2.2.2. Protein Extraction
2.3. Analysis
2.3.1. Proximate Composition
2.3.2. Protein Profile
2.3.3. Protein Identification by Mass Spectrometry
Mass Spectrometry
Database Searching
Criteria for Protein Identification
2.4. Protein Solubility
2.5. Foaming Properties
2.6. Statistical Analysis
3. Results
3.1. Proximate Composition of Meals and Protein Extracts
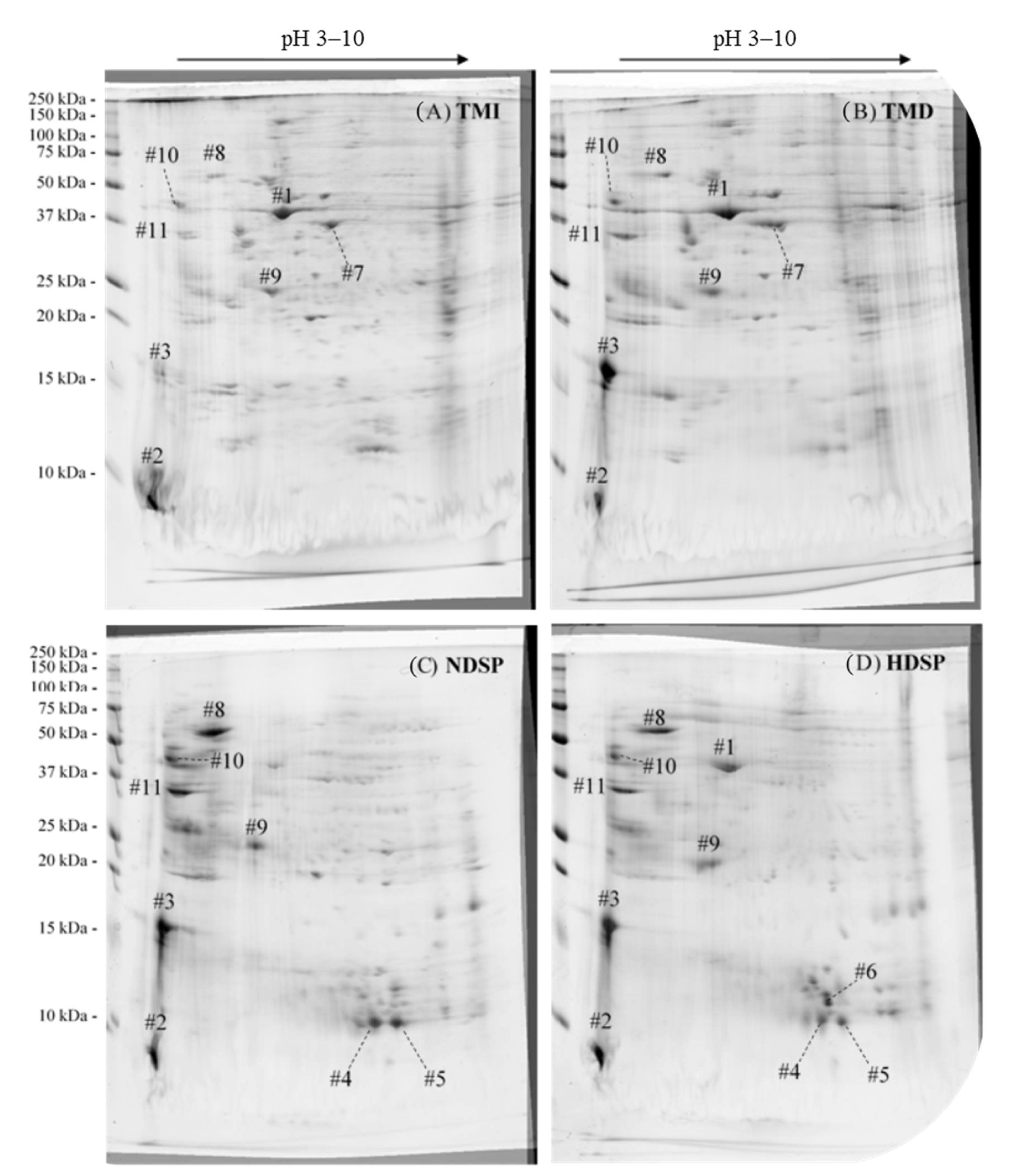
3.2. Profiles and Characterization of Proteins in Meals and Protein Extracts
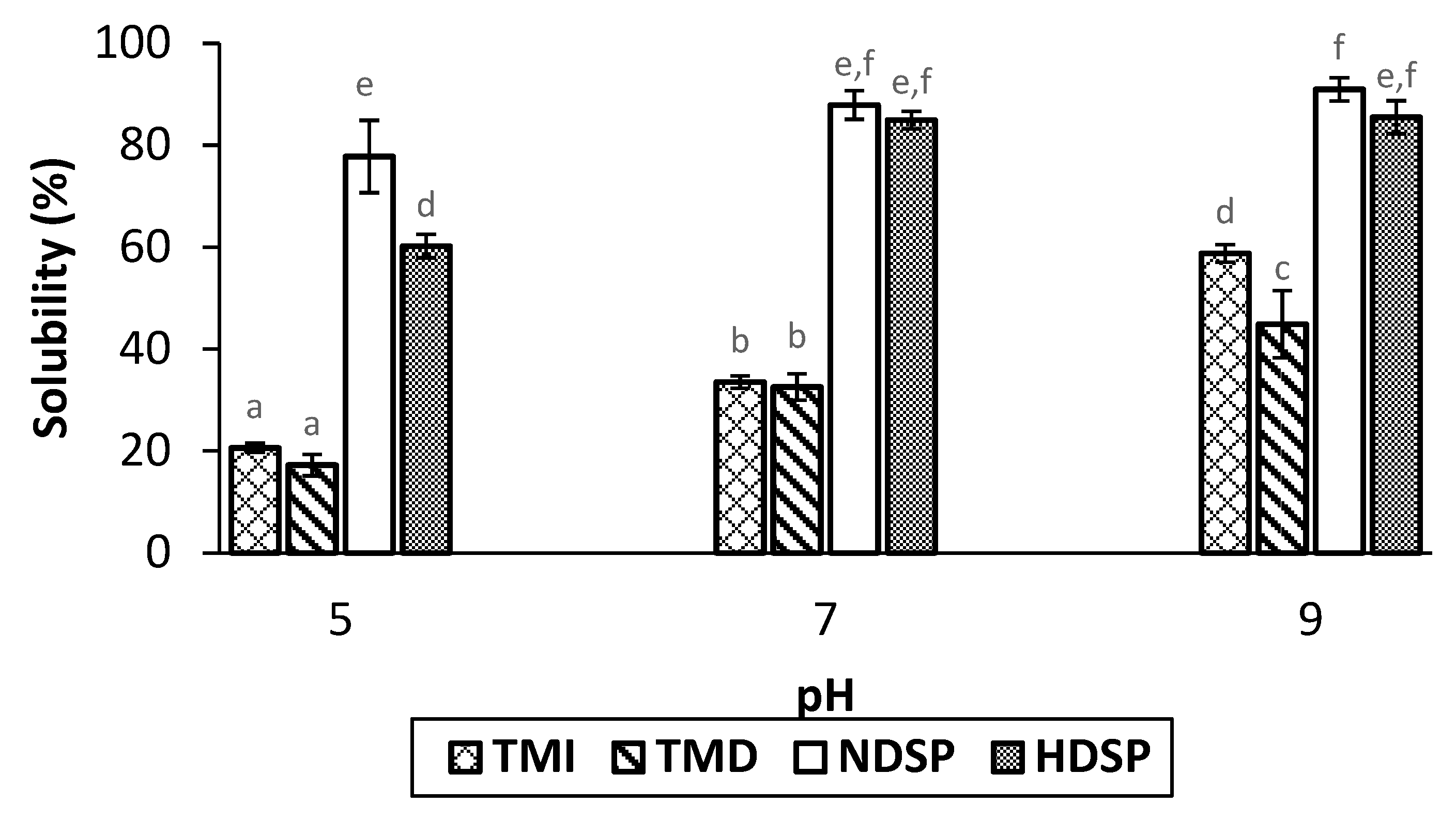
3.3. Protein Solubility
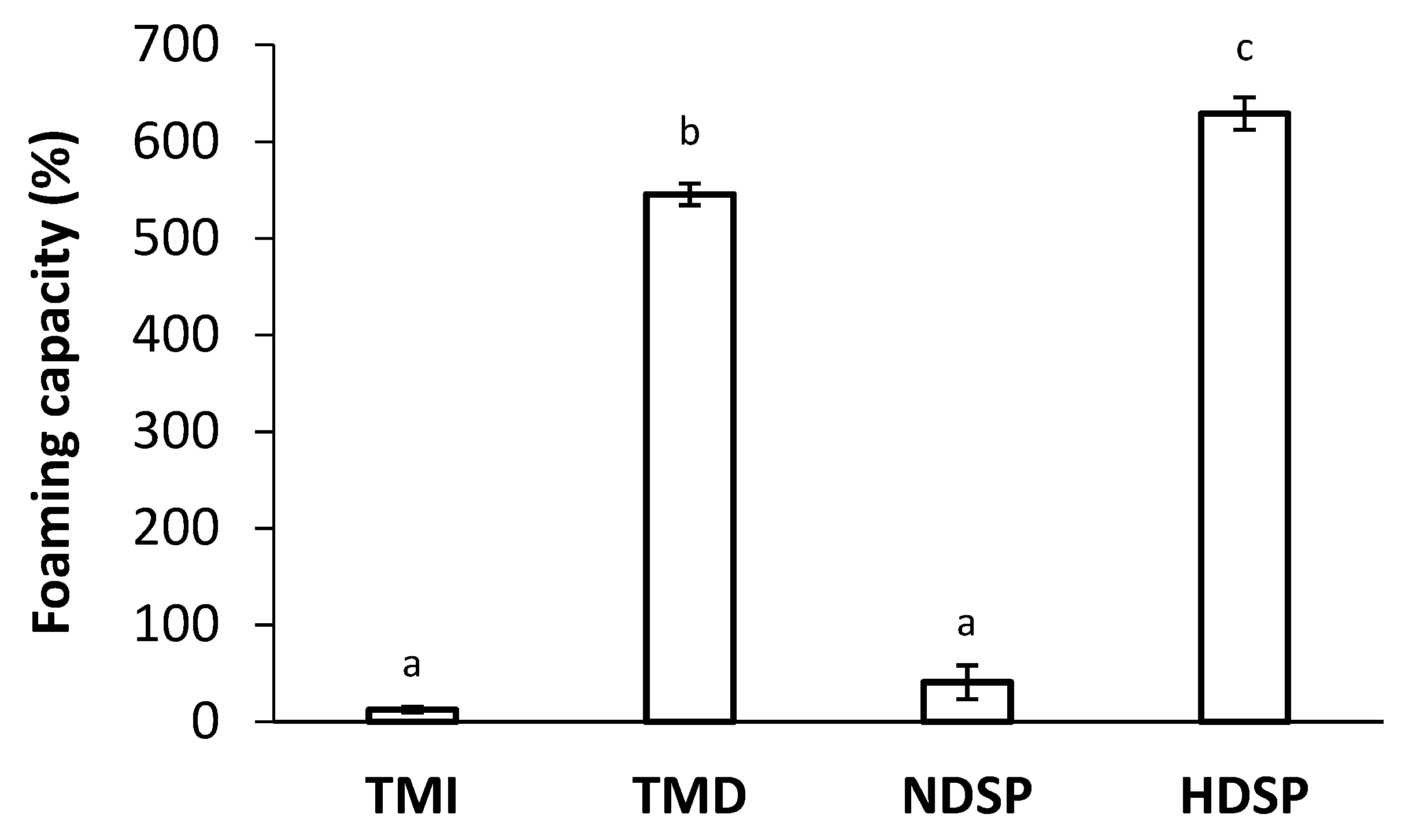
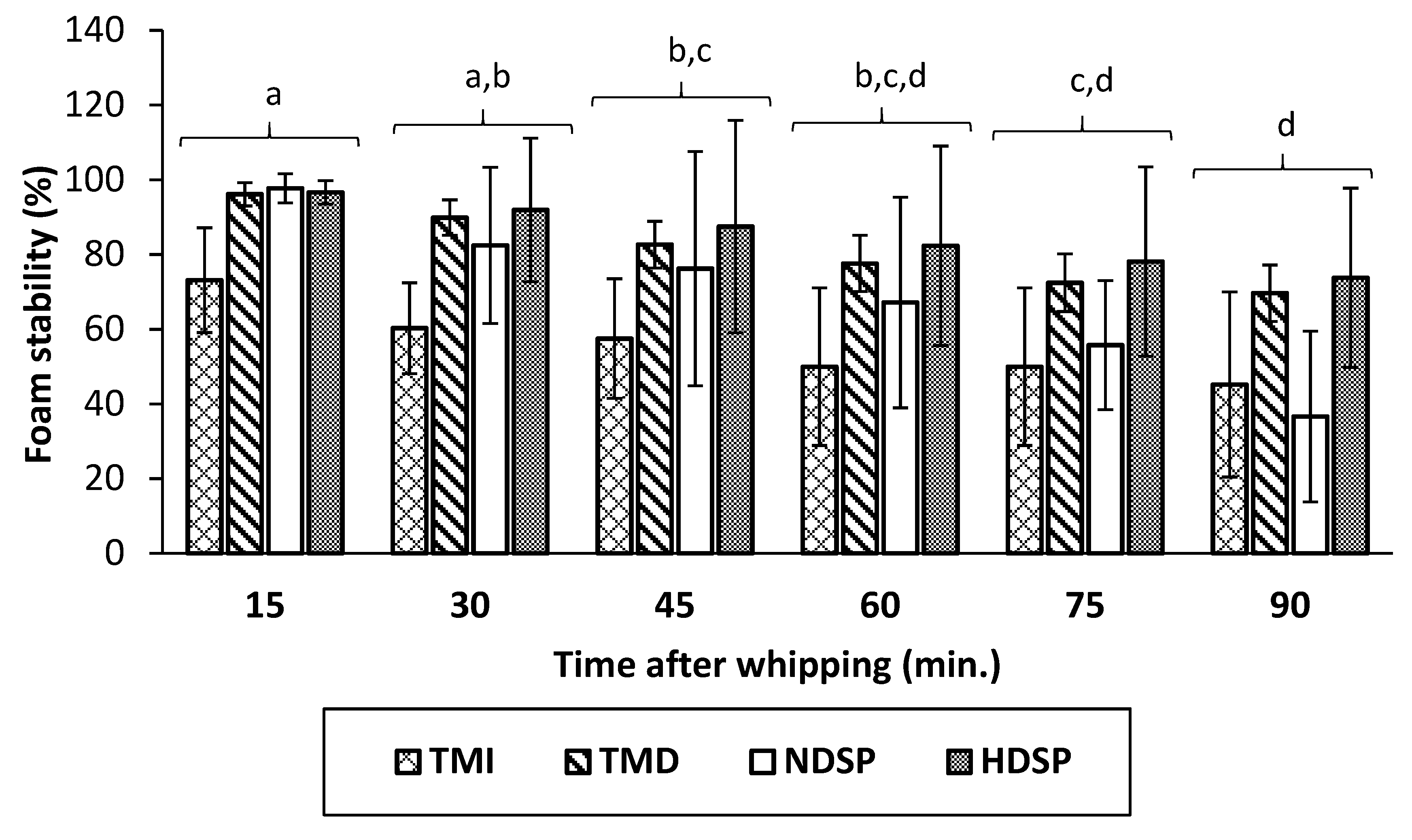
3.4. Foaming Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Oonincx, D.G.; van Itterbeeck, J.; Heetkamp, M.J.; van den Brand, H.; van Loon, J.J.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed]
- La Barbera, F.; Verneau, F.; Amato, M.; Grunert, K. Understanding Westerners’ disgust for the eating of insects: The role of food neophobia and implicit associations. Food Qual. Prefer. 2018, 64, 120–125. [Google Scholar] [CrossRef]
- Gere, A.; Székely, G.; Kovács, S.; Kókai, Z.; Sipos, L. Readiness to adopt insects in Hungary: A case study. Food Qual. Prefer. 2017, 59, 81–86. [Google Scholar] [CrossRef]
- Verbeke, W. Profiling consumers who are ready to adopt insects as a meat substitute in a Western society. Food Qual. Prefer. 2015, 39, 147–155. [Google Scholar] [CrossRef]
- Schosler, H.; de Boer, J.; Boersema, J.J. Can we cut out the meat of the dish? Constructing consumer-oriented pathways towards meat substitution. Appetite 2012, 58, 39–47. [Google Scholar] [CrossRef]
- Williams, J.P.; Williams, J.R.; Kirabo, A.; Chester, D.; Peterson, M. Chapter 3—Nutrient Content and Health Benefits of Insects. In Insects as Sustainable Food Ingredients; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 61–84. [Google Scholar]
- Zielińska, E.; Karaś, M.; Baraniak, B. Comparison of functional properties of edible insects and protein preparations thereof. LWT 2018, 91, 168–174. [Google Scholar] [CrossRef]
- Stone, A.K.; Tanaka, T.; Nickerson, M.T. Protein quality and physicochemical properties of commercial cricket and mealworm powders. J. Food Sci. Technol. 2019, 56, 3355–3363. [Google Scholar] [CrossRef]
- Mishyna, M.; Martinez, J.I.; Chen, J.; Benjamin, O. Extraction, characterization and functional properties of soluble proteins from edible grasshopper (Schistocerca gregaria) and honey bee (Apis mellifera). Food Res. Int. 2019, 116, 697–706. [Google Scholar] [CrossRef]
- Gravel, A.; Doyen, A. The use of edible insect proteins in food: Challenges and issues related to their functional properties. Innov. Food Sci. Emerg. Technol. 2020, 59. [Google Scholar] [CrossRef]
- Cappelli, A.; Cini, E.; Lorini, C.; Oliva, N.; Bonaccorsi, G. Insects as food: A review on risks assessments of Tenebrionidae and Gryllidae in relation to a first machines and plants development. Food Control. 2020, 108, 106877. [Google Scholar] [CrossRef]
- L’hocine, L.; Boye, J.I.; Arcand, Y. Composition and Functional Properties of Soy Protein Isolates Prepared Using Alternative Defatting and Extraction Procedures. J. Food Sci. 2006, 71, C137–C145. [Google Scholar] [CrossRef]
- Capellini, M.C.; Novais, J.S.; Monteiro, R.F.; Veiga, B.Q.; Osiro, D.; Rodrigues, C.E.C. Thermal, structural and functional properties of rice bran defatted with alcoholic solvents. J. Cereal Sci. 2020, 95, 103067. [Google Scholar] [CrossRef]
- Teh, S.-S.; Bekhit, A.E.-D.; Carne, A.; Birch, J. Effect of the defatting process, acid and alkali extraction on the physicochemical and functional properties of hemp, flax and canola seed cake protein isolates. J. Food Meas. Charact. 2013, 8, 92–104. [Google Scholar] [CrossRef]
- Yi, L.; Van Boekel, M.A.J.S.; Boeren, S.; Lakemond, C.M.M. Protein identification and in vitro digestion of fractions from Tenebrio molitor. Eur. Food Res. Technol. 2016, 242, 1285–1297. [Google Scholar] [CrossRef]
- Choi, B.D.; Wong, N.A.K.; Auh, J.H. Defatting and Sonication Enhances Protein Extraction from Edible Insects. Korean J. Food Sci. Anim. Resour. 2017, 37, 955–961. [Google Scholar] [PubMed]
- Son, Y.J.; Choi, S.Y.; Hwang, I.K.; Nho, C.W.; Kim, S.H. Could Defatted Mealworm (Tenebrio molitor) and Mealworm Oil Be Used as Food Ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Russin, T.A.; Boye, J.I.; Arcand, Y.; Rajamohamed, S.H. Alternative Techniques for Defatting Soy: A Practical Review. Food Bioprocess. Technol. 2010, 4, 200–223. [Google Scholar] [CrossRef]
- Laroche, M.; Perreault, V.; Marciniak, A.; Gravel, A.; Chamberland, J.; Doyen, A. Comparison of Conventional and Sustainable Lipid Extraction Methods for the Production of Oil and Protein Isolate from Edible Insect Meal. Foods 2019, 8, 572. [Google Scholar] [CrossRef]
- Kim, T.K.; Yong, H.I.; Kim, Y.B.; Jung, S.; Kim, H.W.; Choi, Y.S. Effects of organic solvent on functional properties of defatted proteins extracted from Protaetia brevitarsis larvae. Food Chem 2020, 336, 127679. [Google Scholar] [CrossRef]
- Purschke, B.; Stegmann, T.; Schreiner, M.; Jäger, H. Pilot-scale supercritical CO2 extraction of edible insect oil from Tenebrio molitor L. larvae-Influence of extraction conditions on kinetics, defatting performance and compositional properties. Eur. J. Lipid Sci. Technol. 2017, 119. [Google Scholar] [CrossRef]
- Borremans, A.; Bussler, S.; Sagu, S.T.; Rawel, H.; Schluter, O.K.; Leen, V.C. Effect of Blanching Plus Fermentation on Selected Functional Properties of Mealworm (Tenebrio molitor) Powders. Foods 2020, 9, 917. [Google Scholar] [CrossRef]
- Tzompa-Sosa, D.A.; Yi, L.; van Valenberg, H.J.F.; van Boekel, M.A.J.S.; Lakemond, C.M.M. Insect lipid profile: Aqueous versus organic solvent-based extraction methods. Food Res. Int. 2014, 62, 1087–1094. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.; Sagis, L.M.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A. Extraction and characterisation of protein fractions from five insect species. Food Chem 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Simonne, A.H.; Simonne, E.H.; Eitenmiller, R.R.; Mills, H.A.; Cresman, C. Could the Dumas method replace the Kjeldahl digestion for nitrogen and crude protein determinations in foods? J. Sci. Food Agric. 1997, 73, 39–45. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.P.; van den Broek, L.A.; Fogliano, V.; Lakemond, C.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2017. [Google Scholar]
- Spinelli, J.; Lehman, L.; Wieg, D. Composition, Processing, and Utilization of Red Crab (Pleuroncodes planipes) as an Aquacultural Feed Ingredient. J. l’Office Rech. Pêcheries Can. 1974, 31, 1025–1029. [Google Scholar] [CrossRef]
- Kumar, M.; Singh, R.; Meena, A.; Patidar, B.S.; Prasad, R.; Chhabra, S.K.; Bansal, S.K. An Improved 2-Dimensional Gel Electrophoresis Method for Resolving Human Erythrocyte Membrane Proteins. Proteom. Insights 2017, 8, 1178641817700880. [Google Scholar] [CrossRef]
- Natale, M.; Maresca, B.; Abrescia, P.; Bucci, E.M. Image Analysis Workflow for 2-D Electrophoresis Gels Based on Image. J. Proteom. Insights 2011, 4. [Google Scholar] [CrossRef]
- Lundgren, D.H.; Hwang, S.-I.; Wu, L.; Han, D.K. Role of spectral counting in quantitative proteomics. Expert Rev. Proteom. 2010, 7, 39–53. [Google Scholar] [CrossRef]
- Boukil, A.; Perreault, V.; Chamberland, J.; Mezdour, S.; Pouliot, Y.; Doyen, A. High Hydrostatic Pressure-Assisted Enzymatic Hydrolysis Affect Mealworm Allergenic Proteins. Molecules 2020, 25, 2685. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Xiong, Y.L.; Qin, F.; Jian, H.; Huang, X.; Chen, J. Surface properties of heat-induced soluble soy protein aggregates of different molecular masses. J. Food Sci. 2015, 80, C279–C287. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Purschke, B.; Mendez Sanchez, Y.D.; Jäger, H. Centrifugal fractionation of mealworm larvae (Tenebrio molitor L.) for protein recovery and concentration. LWT 2018, 89, 224–228. [Google Scholar] [CrossRef]
- Buβler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schluter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, e00218. [Google Scholar]
- Lam, A.C.Y.; Can Karaca, A.; Tyler, R.T.; Nickerson, M.T. Pea protein isolates: Structure, extraction, and functionality. Food Rev. Int. 2018, 34, 126–147. [Google Scholar] [CrossRef]
- Barre, A.; Pichereaux, C.; Velazquez, E.; Maudouit, A.; Simplicien, M.; Garnier, L.; Bienvenu, F.; Bienvenu, J.; Burlet-Schiltz, O.; Auriol, C.; et al. Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor). Foods 2019, 8, 515. [Google Scholar] [CrossRef]
- Verhoeckx, K.C.; van Broekhoven, S.; den Hartog-Jager, C.F.; Gaspari, M.; de Jong, G.A.; Wichers, H.J.; van Hoffen, E.; Houben, G.F.; Knulst, A.C. YiHouse dust mite (Der p 10) and crustacean allergic patients may react to food containing Yellow mealworm proteins. Food Chem. Toxicol. 2014, 65, 364–373. [Google Scholar] [CrossRef]
- Francis, F.; Doyen, V.; Debaugnies, F.; Mazzucchelli, G.; Caparros, R.; Alabi, T.; Blecker, C.; Haubruge, E.; Corazza, F. Limited cross reactivity among arginine kinase allergens from mealworm and cricket edible insects. Food Chem. 2019, 276, 714–718. [Google Scholar] [CrossRef]
- Salgado, V.L.; Sparks, T.C. 6.5—The Spinosyns: Chemistry, Biochemistry, Mode of Action, and Resistance. In Comprehensive Molecular Insect Science; Gilbert, L.I., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 137–173. [Google Scholar]
- Kanost, M.R. Chapter 117—Hemolymph. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 446–449. [Google Scholar]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Andersen, S.O. Characteristic properties of proteins from pre-ecdysial cuticle of larvae and pupae of the mealworm Tenebrio molitor. Insect Biochem. Mol. Biol. 2002, 32, 1077–1087. [Google Scholar] [CrossRef]
- Andersen, S.O.; Rafn, K.; Roepstorff, P. Sequence studies of proteins from larval and pupal cuticle of the yellow meal worm, Tenebrio molitor. Insect Biochem. Mol. Biol. 1997, 27, 121–131. [Google Scholar] [CrossRef]
- Azagoh, C.; Ducept, F.; Garcia, R.; Rakotozafy, L.; Cuvelier, M.E.; Keller, S.; Lewandowski, R.; Mezdour, S. Extraction and physicochemical characterization of Tenebrio molitor proteins. Food Res. Int. 2016, 88, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Galves, C.; Stone, A.K.; Szarko, J.; Liu, S.; Shafer, K.; Hargreaves, J.; Siarkowski, M.; Nickerson, M.T. Effect of pH and defatting on the functional attributes of safflower, sunflower, canola, and hemp protein concentrates. Cereal Chem. 2019, 96, 1036–1047. [Google Scholar] [CrossRef]
- Zayas, J.F. Solubility of Proteins. In Functionality of Proteins in Food; Zayas, J.F., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 6–75. [Google Scholar]
- Akpossan, R.; Digbeu, Y.; Koffi, M.; Kouadio, J.; Dué, E.; Kouamé, P. Protein Fractions and Functional Properties of Dried Imbrasia oyemensis Larvae Full-Fat and Defatted Flours. Int. J. Biochem. Res. Rev. 2015, 5, 116–126. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Processing Stage | Protein | Lipid | Chitin | Ash | Moisture | Total Solids |
|---|---|---|---|---|---|---|
| % (w/w) | ||||||
| TMI | 38.6 ± 1.4 a | 28.5 ± 1.0 a | 5.0 ± 0.6 a | 3.4 ± 0.1 a | 9.7 ± 3.2 a | 90.3 ± 3.2 a |
| TMD | 59.1 ± 0.9 b,c | 0.4 ± 0.2 b | 7.8 ± 0.6 b | 4.8 ± 0.1 a | 4.8 ± 0.1 b | 95.2 ± 0.1 b |
| HDSP | 62.7 ± 3.0 c | 0.3 ± 0.1 b | 0.0 ± 0.0 c | 8.4 ± 1.1 b | 7.3 ± 0.8 a,b | 92.7 ± 0.8 a,b |
| NDSP | 54.0 ± 5.0 b | 0.5 ± 0.1 b | 0.0 ± 0.0 c | 10.3 ± 1.3 b | 9.3 ± 1.2 a,b | 90.7 ± 1.2 a,b |
| # | Identified Proteins | Molecular Weight MW (kDa) | UniProt ID | Number of Unique Peptides | Coverage (%) | Total Spectrum Count | Most Abundant in Fractions |
|---|---|---|---|---|---|---|---|
| 1 | Actin-87E-like protein (Tribolium castaneum) | 42 | D6WF19 | 39 | 77 | 736 | TMI, TMD, and HDSP |
| 2 | Cockroach allergen-like protein (Tenebrio molitor) | 65 | Q7YZB8 | 5 | 11 | 53 | TMI, TMD, NDSP, and HDSP |
| 3 | 9 | 15 | 158 | TMD, NDSP, and HDSP | |||
| 4 | 12 kDa hemolymph protein b (Tenebrio molitor) | 14 | Q7YWD7 | 2 | 81 | 297 | NDSP and HDSP |
| 5 | 2 | 81 | 271 | ||||
| 6 | 86 kDa early-staged encapsulation inducing protein (Tenebrio molitor) | 91 | Q9Y1W5 | 21 | 27 | 93 | HDSP |
| Hexamerin 2 (Tenebrio molitor) | 85 | Q95PI7 | 12 | 20 | 81 | ||
| 7 | Arginine kinase (Fragment) (Tenebrio molitor) | 27 | A0A0U4ARJ8 | 8 | 85 | 414 | TMI and TMD |
| 8 | α-amylase (Tenebrio molitor) | 51 | P56634 | 30 | 80 | 731 | NDSP and HDSP |
| 9 | 28 kDa desiccation stress protein (Tenebrio molitor) | 25 | Q27013 | 21 | 49 | 202 | TMI, TMD, NDSP, and HDSP |
| 10 | Melanin-inhibiting protein (Tenebrio molitor) | 40 | Q4LE89 | 25 | 64 | 225 | NDSP and HDSP |
| 11 | Chitinase (Tenebrio molitor) | 40 | Q7YZB9 | 11 | 38 | 156 | NDSP and HDSP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gravel, A.; Marciniak, A.; Couture, M.; Doyen, A. Effects of Hexane on Protein Profile, Solubility and Foaming Properties of Defatted Proteins Extracted from Tenebrio molitor Larvae. Molecules 2021, 26, 351. https://doi.org/10.3390/molecules26020351
Gravel A, Marciniak A, Couture M, Doyen A. Effects of Hexane on Protein Profile, Solubility and Foaming Properties of Defatted Proteins Extracted from Tenebrio molitor Larvae. Molecules. 2021; 26(2):351. https://doi.org/10.3390/molecules26020351
Chicago/Turabian StyleGravel, Alexia, Alice Marciniak, Manon Couture, and Alain Doyen. 2021. "Effects of Hexane on Protein Profile, Solubility and Foaming Properties of Defatted Proteins Extracted from Tenebrio molitor Larvae" Molecules 26, no. 2: 351. https://doi.org/10.3390/molecules26020351
APA StyleGravel, A., Marciniak, A., Couture, M., & Doyen, A. (2021). Effects of Hexane on Protein Profile, Solubility and Foaming Properties of Defatted Proteins Extracted from Tenebrio molitor Larvae. Molecules, 26(2), 351. https://doi.org/10.3390/molecules26020351