
A Systematic Review of the Biological Effects of Cordycepin

 and
and
Abstract
1. Introduction
2. Results
2.1. History of the Field
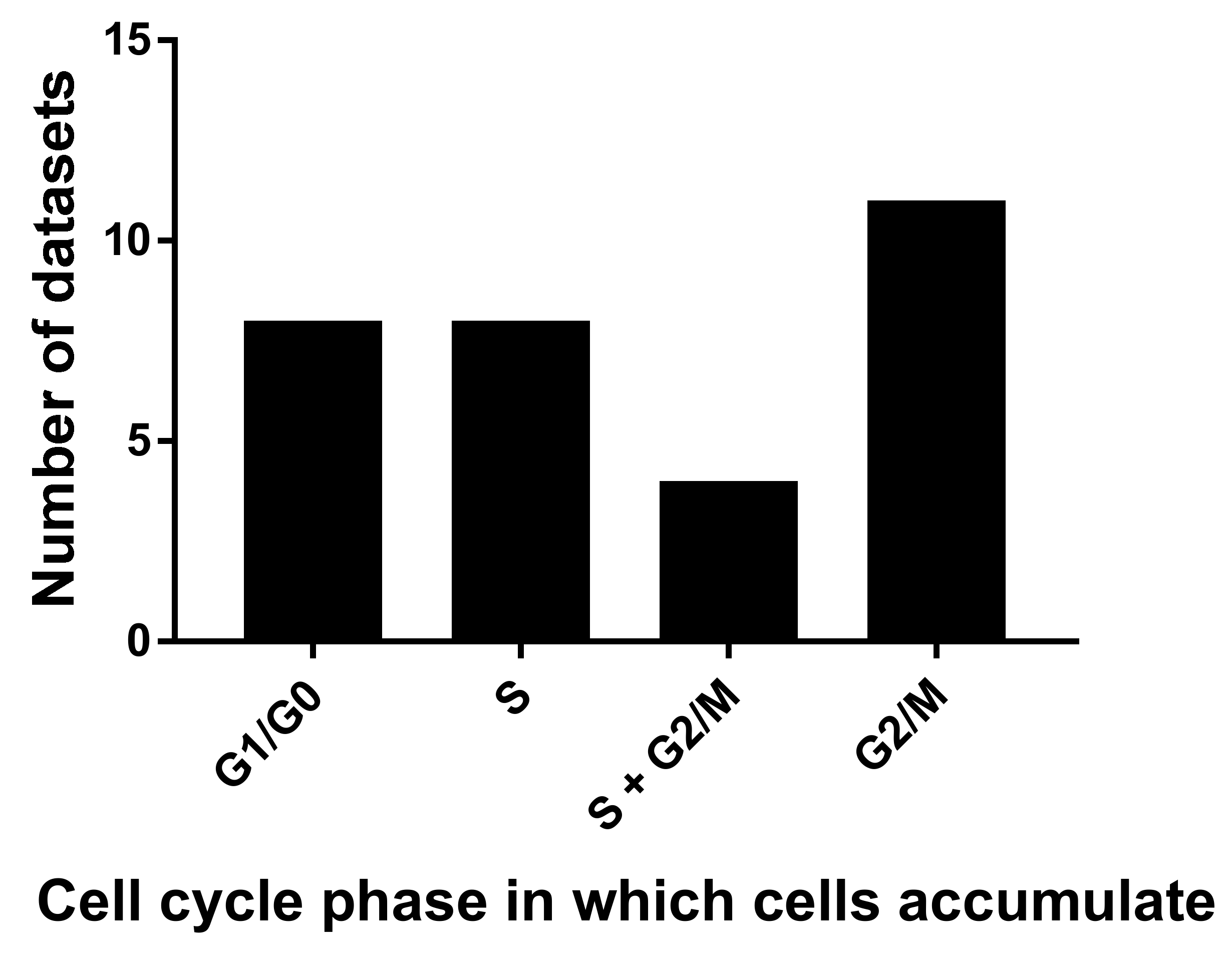
2.2. Cell Death, Survival and Division
2.3. Cell Migration
2.4. Effects on the Inflammatory Response
2.5. Effects on Signal Transduction Pathways
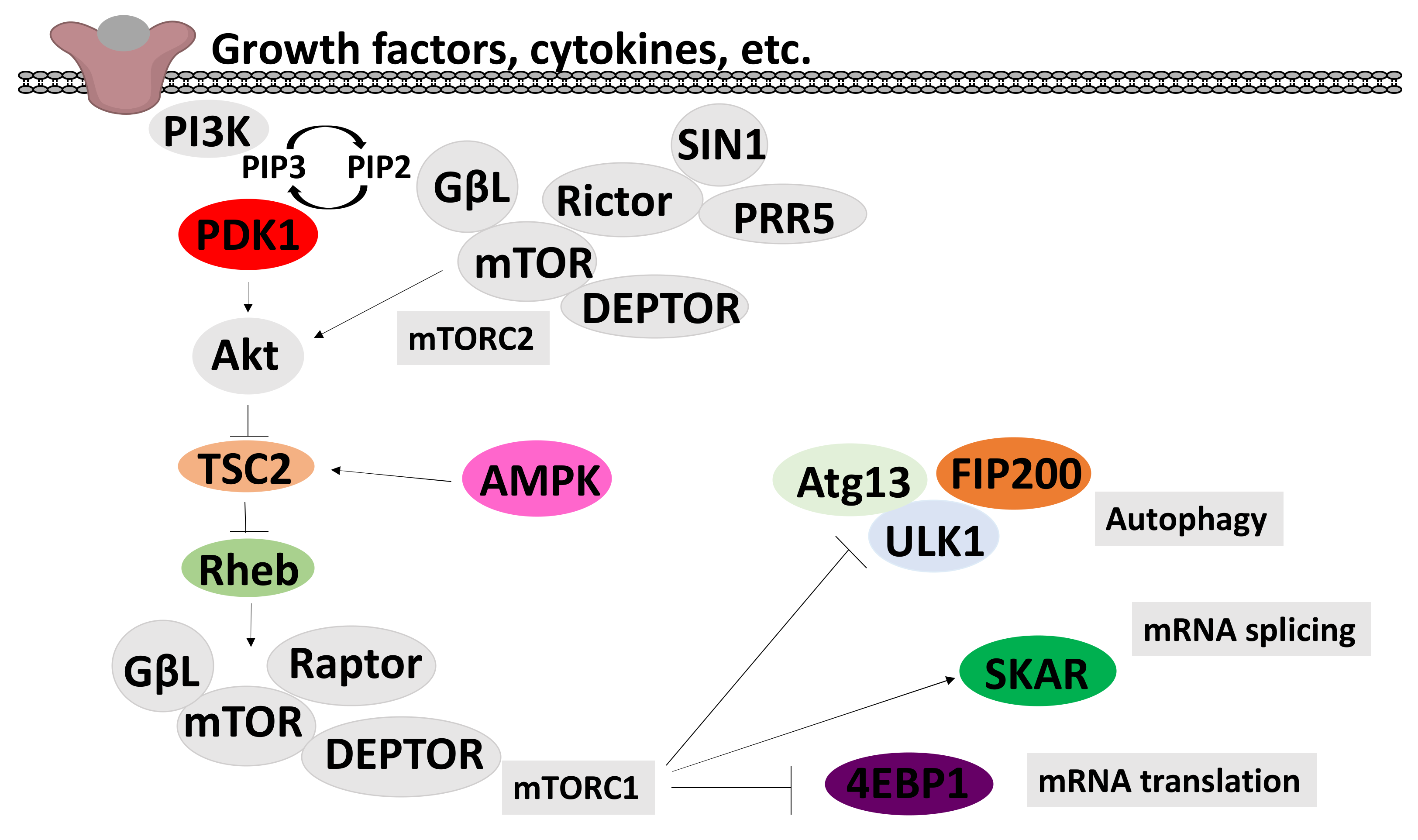
2.6. Effects on the PI3K/mTOR/Akt Pathway

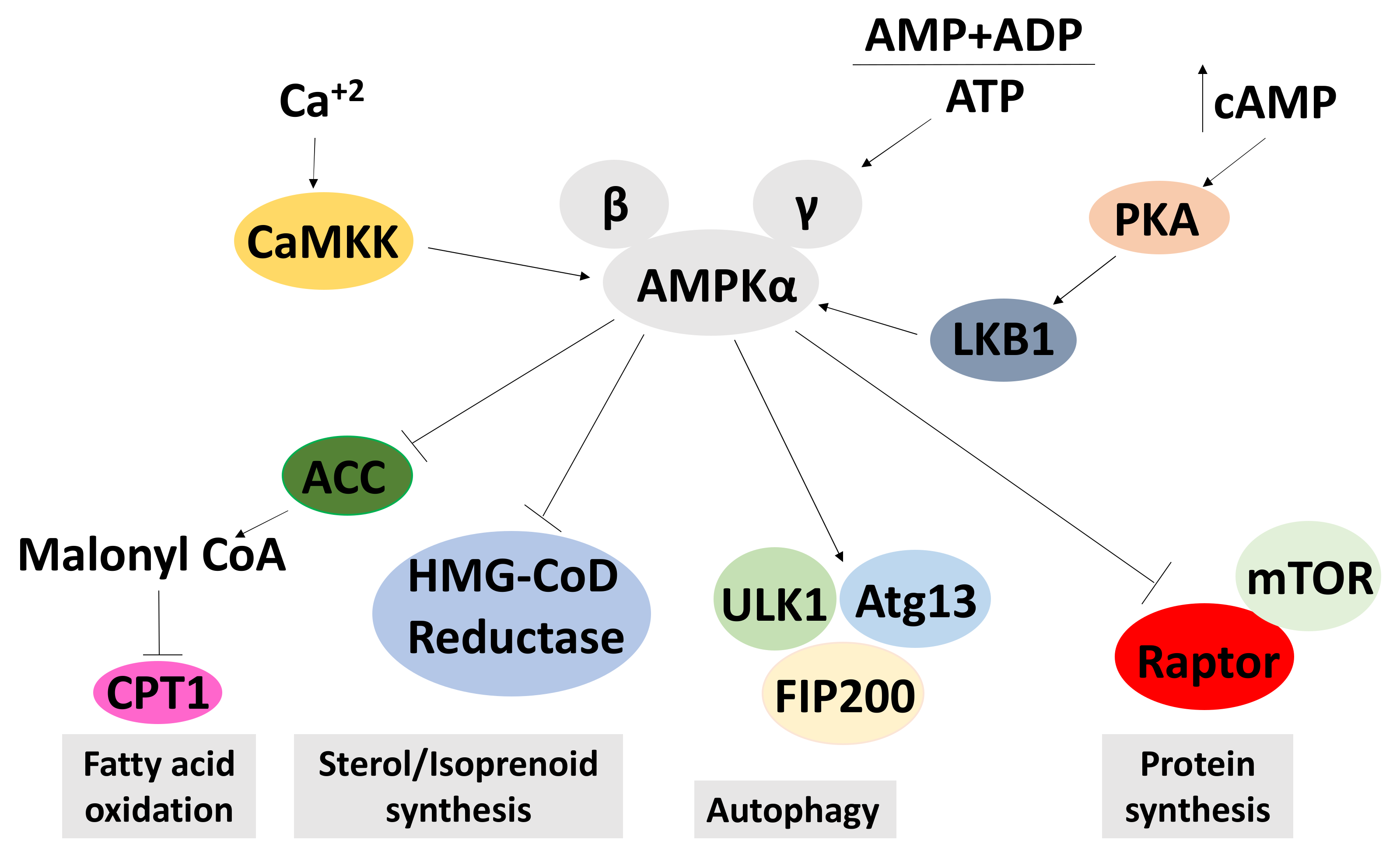
2.6.1. Effects on AMPK Signalling
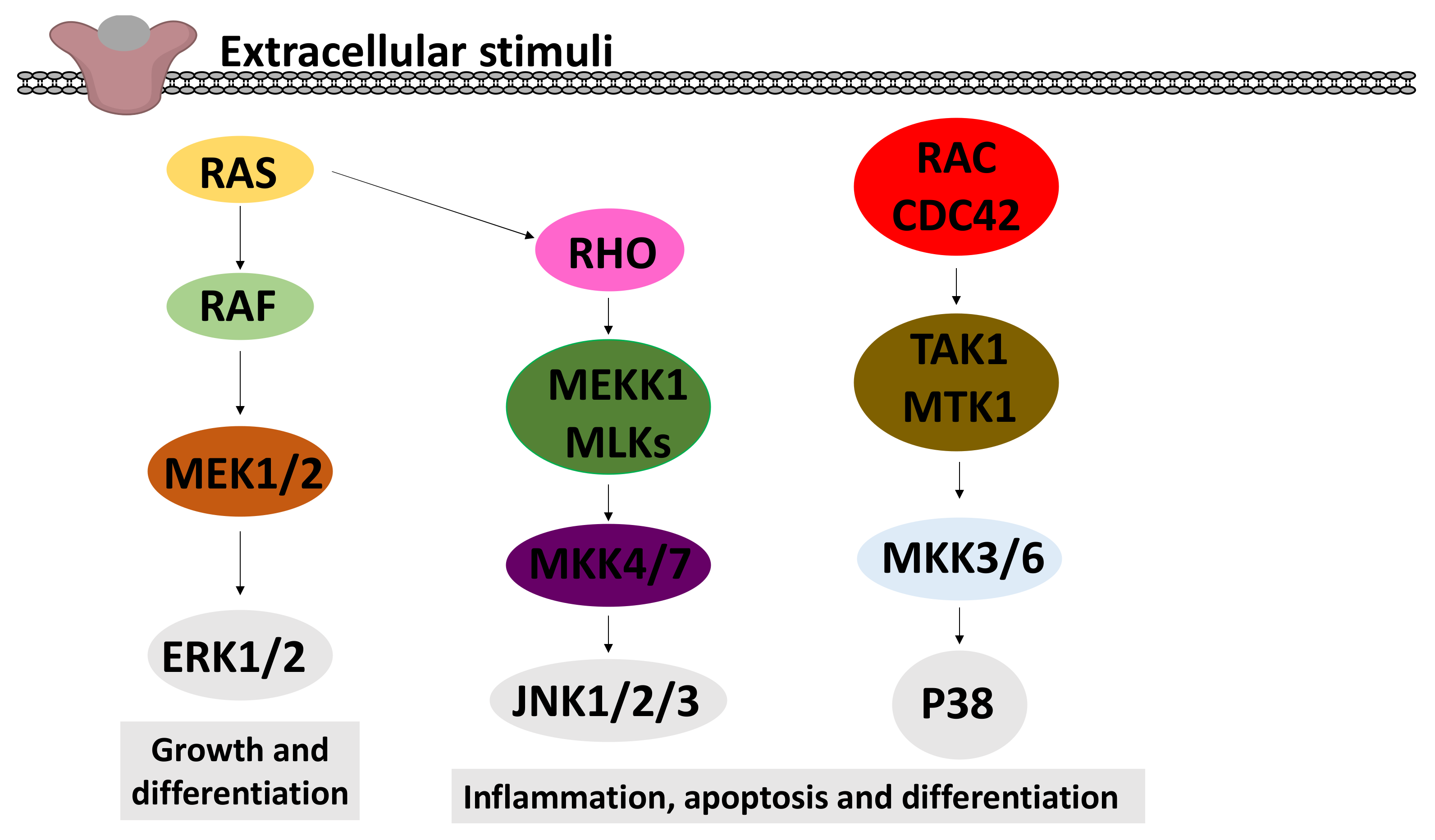
2.6.2. Effects on MAPK Signalling
2.6.3. Signal Transduction Effects in Animal Models
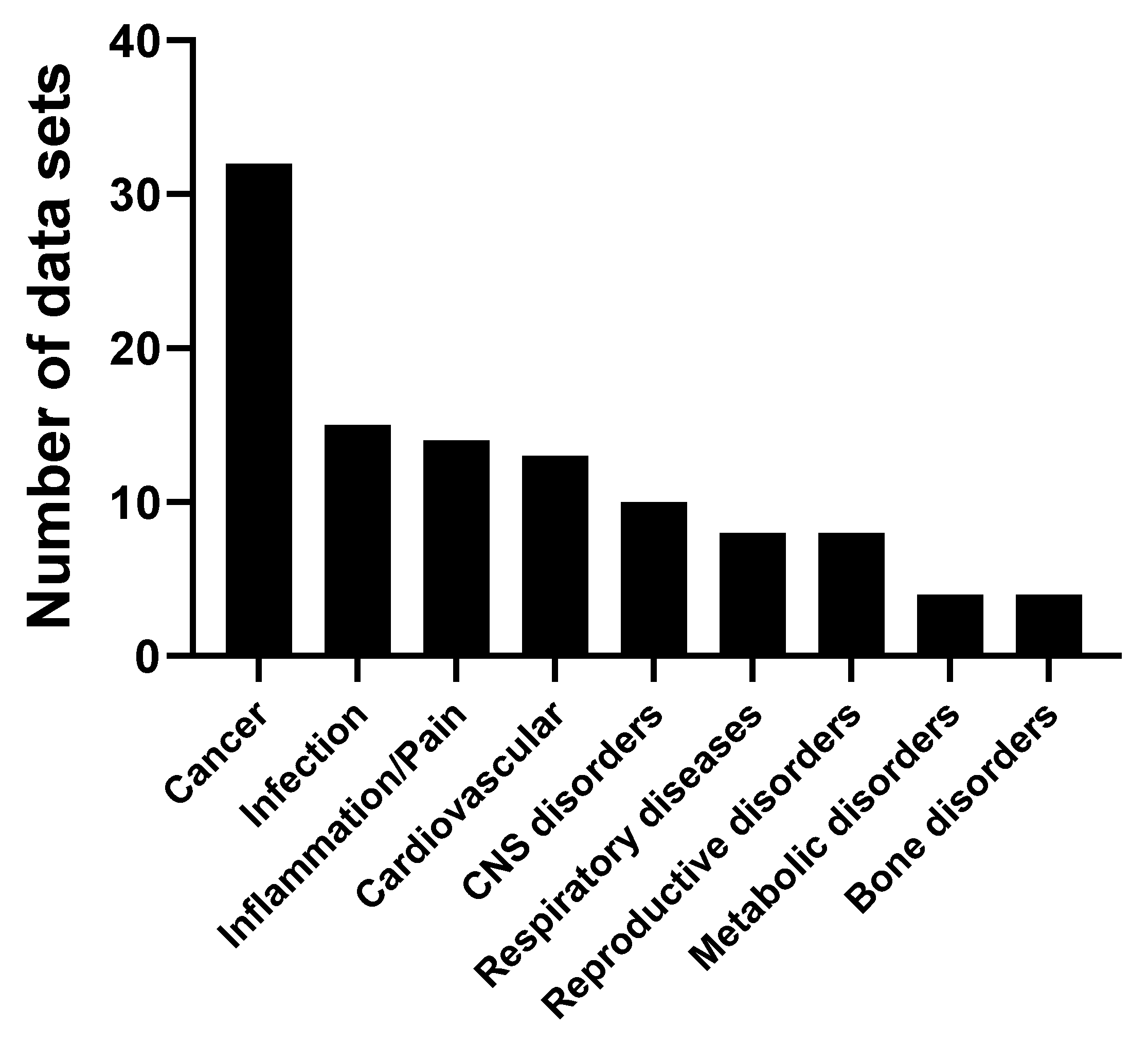
2.7. Cordycepin Activity in Animal Models
3. Discussion
4. Materials and Methods
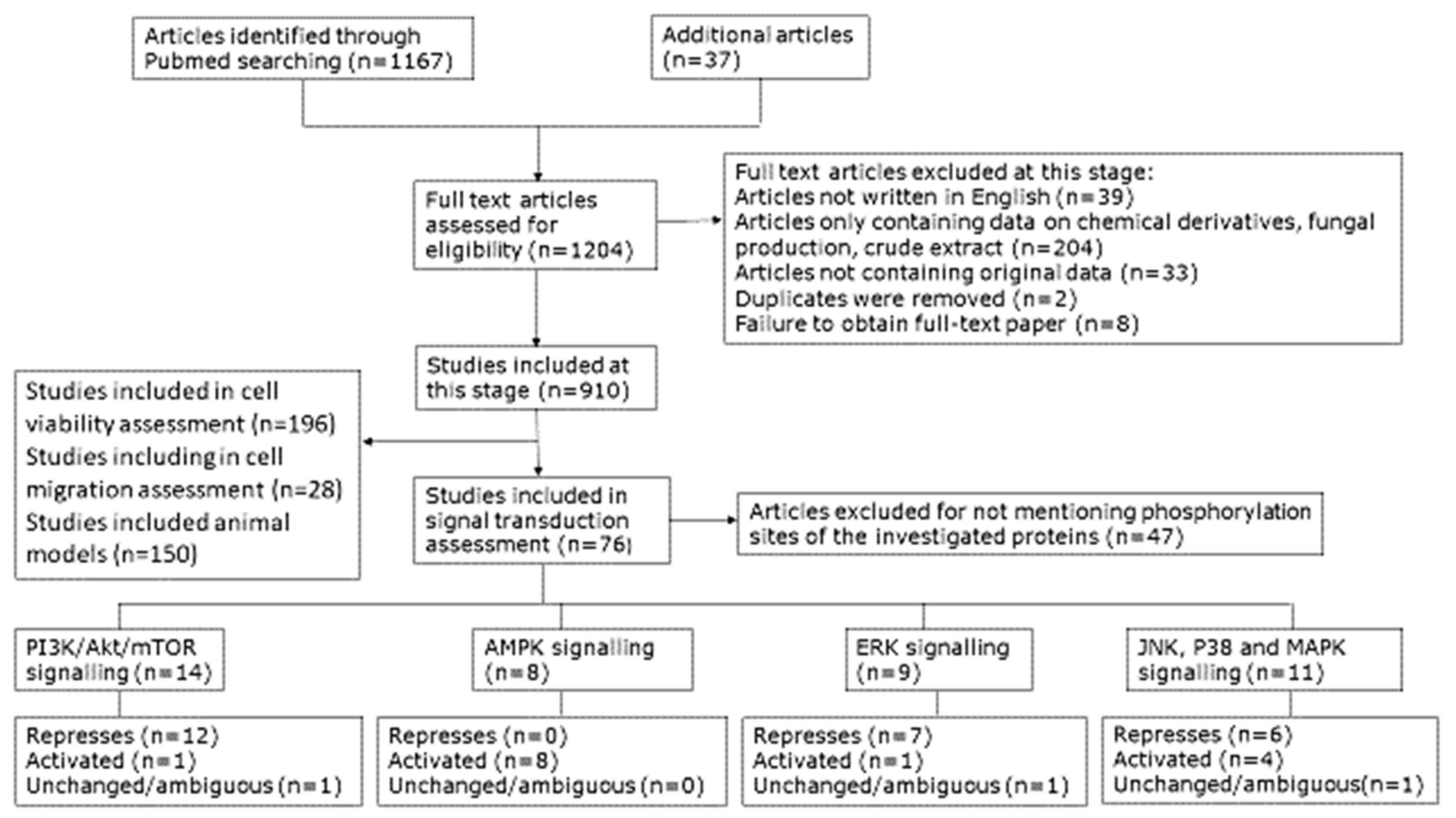
4.1. Scoping of the Review
4.2. Selecting Sources of Data
4.3. Quantifications
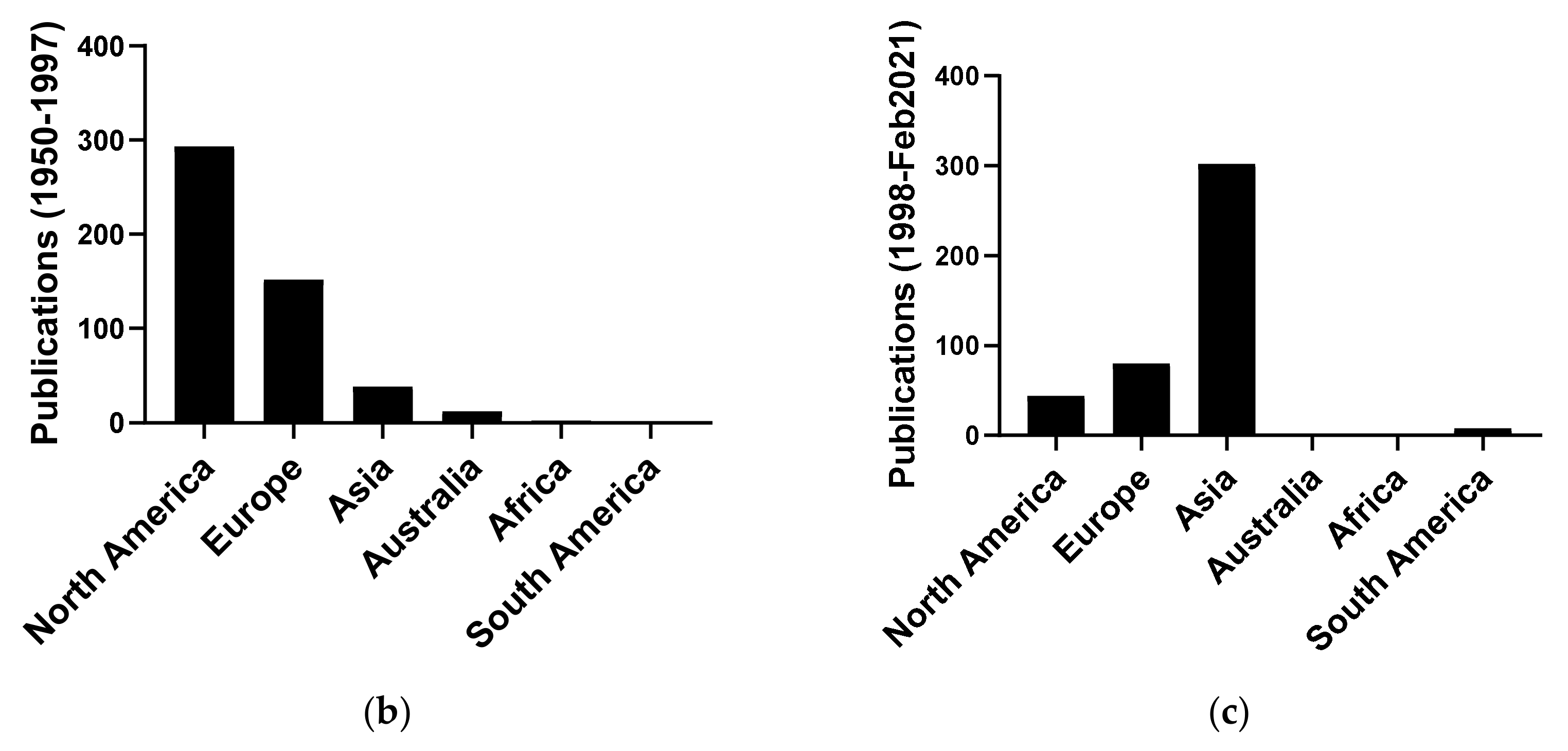
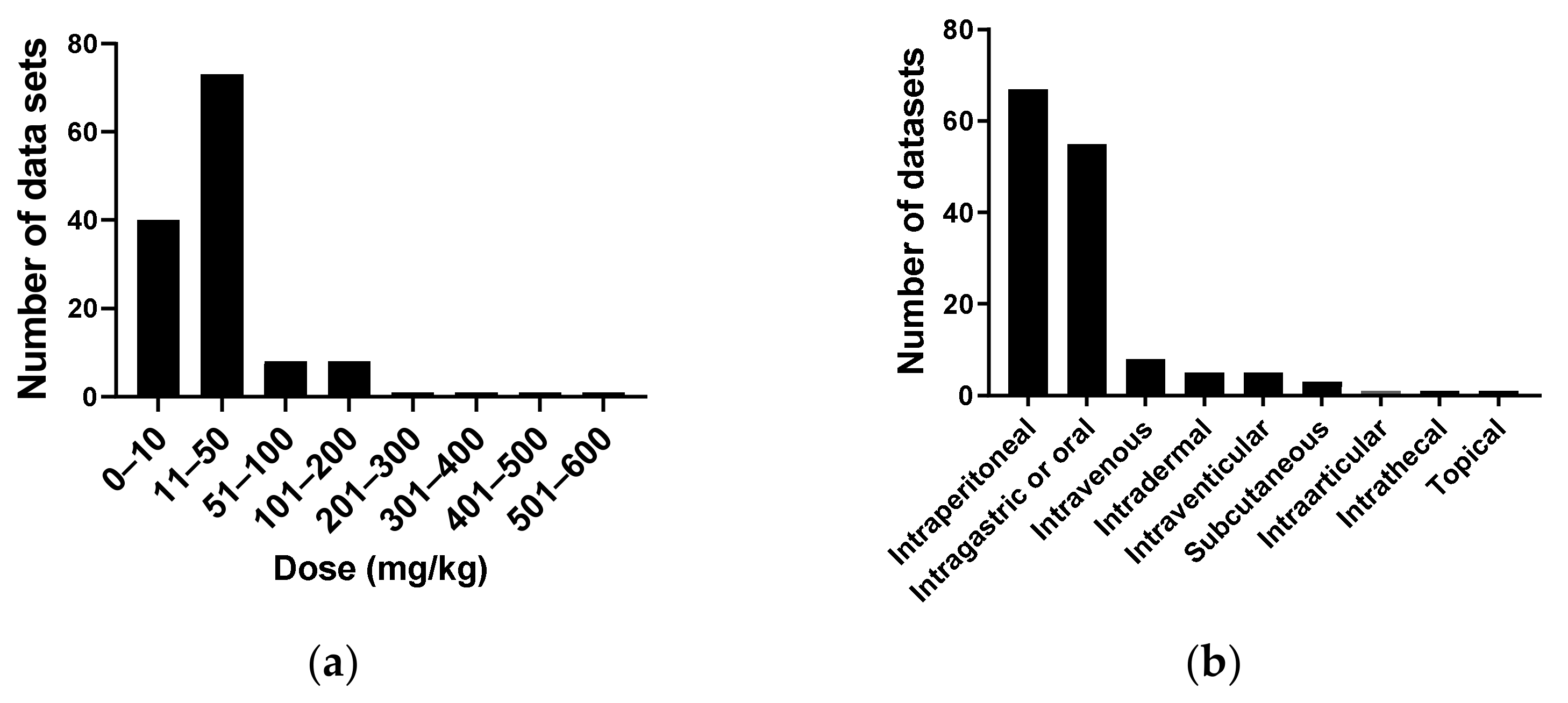
4.3.1. Publication Year
4.3.2. Geographic Origin
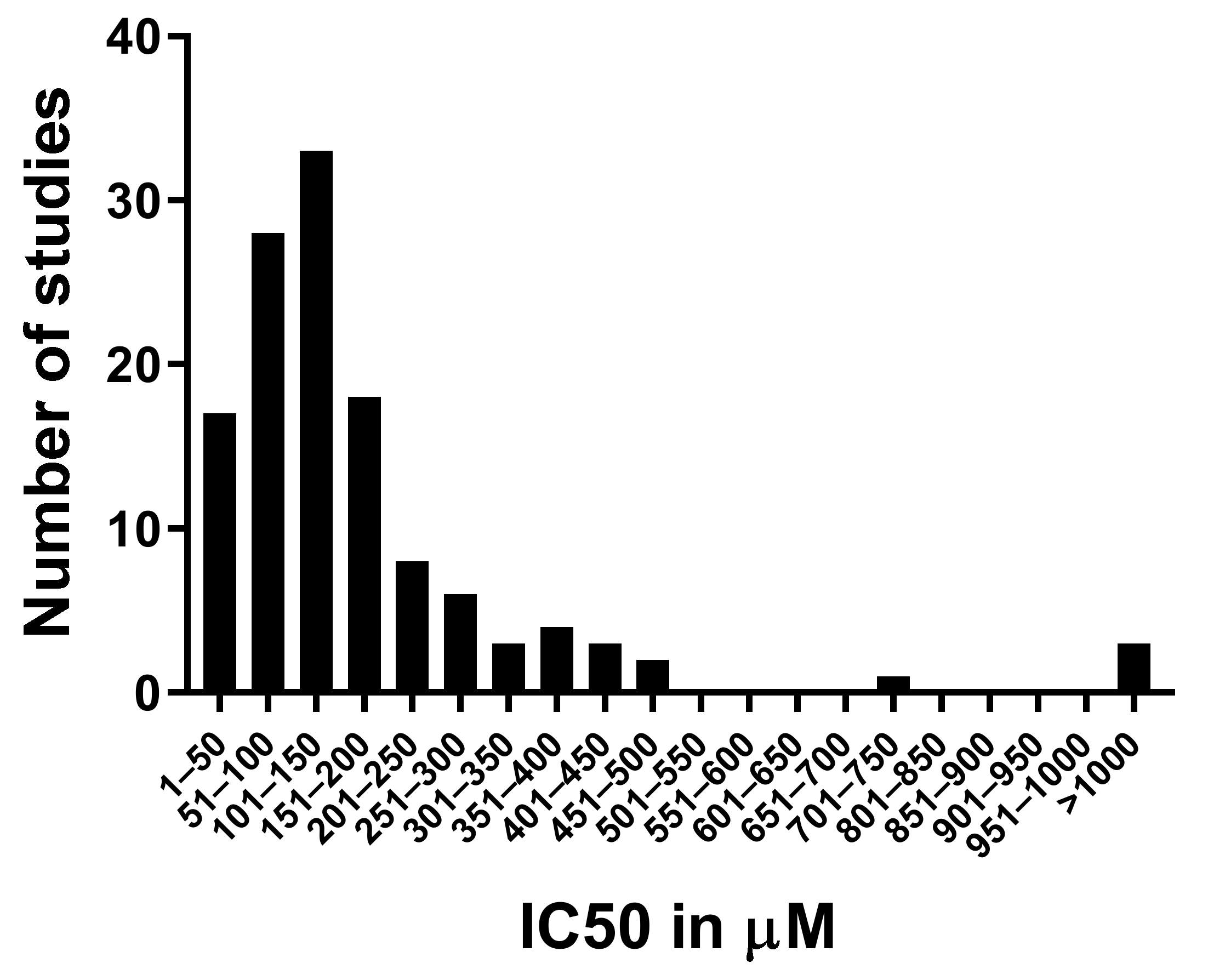
4.3.3. Cordycepin Concentrations and Cell Viability in Mammalian Tissue Culture
4.3.4. Cell Migration
4.3.5. Cellular Inflammation
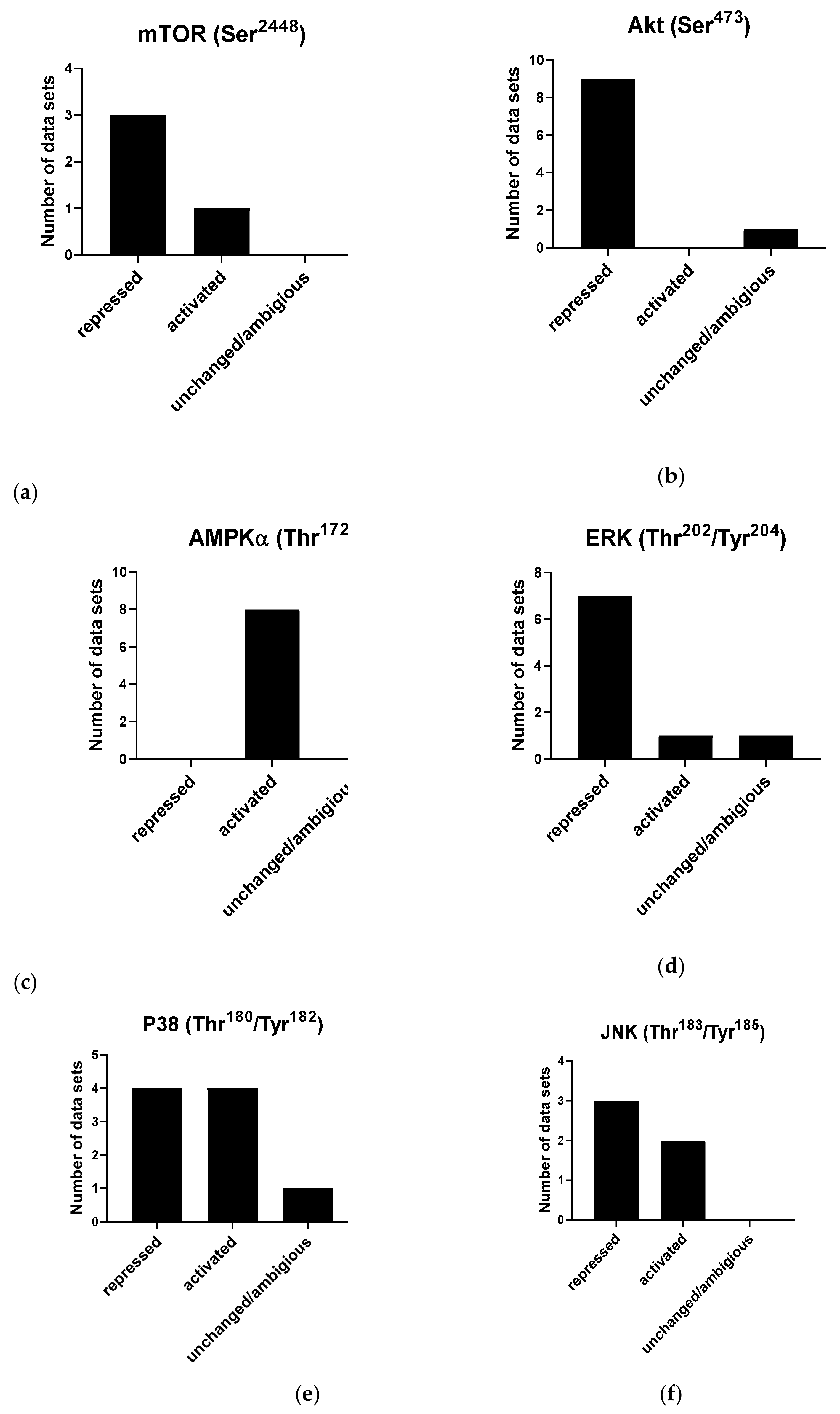
4.3.6. Signal Transduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | mTOR Ser2448 | Akt Ser473 | Akt Thr308 | Akt Total |
|---|---|---|---|---|
| Repressed | [11,44,142] | [11,101,103,142,144,149,150,151,152,153] | [154] | [152,155] |
| Activated | [38] | |||
| Unchanged/ambiguous | [145] | [145] | ||
| (b) | AMPKα Thr172 | AMPKβ Ser108 | ||
| Repressed | ||||
| Activated | [113,142,145,146,152,164,166,167] | [152] | ||
| Unchanged/ambiguous | ||||
| (c) | ERK Thr202/Tyr204 | |||
| Repressed | [55,103,143,151,174,177,178] | |||
| Activated | [11] | |||
| Unchanged/ambiguous | [176] | |||
| (d) | P38 Thr180/Tyr182 | JNK Thr183/Tyr185 | ||
| Repressed | [101,103,112,174] | [35,103,177] | ||
| Activated | [11,175,176,177] | [11,176] | ||
| Unchanged/ambiguous | [178] | |||
4.3.7. Effects in Animal Models
| mTOR Ser2448 | Akt Ser473 | AMPKα Thr172 | ERK Thr202/Tyr204 | JNK Thr183/Tyr185 | |
|---|---|---|---|---|---|
| Repressed | [142,184] | [142] | [184] | [35] | |
| Activated | [183] | [142,164,184] | |||
| Unchanged/Ambiguous |
| Animal Model | Publications |
|---|---|
| Cancer | [11,18,23,24,64,66,74,79,80,107,108,134,142,143,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203] |
| Cardiovascular | [37,76,120,122,164,183,184,204,206,207,208,209,211] |
| Infection | [220,221,222,223,224,225,226,227,228,229,230,231] |
| Central Nervous System | [28,38,116,119,232,233,234,235,236,237] |
| Respiratory Diseases | [94,117,118,123,124,128,129] |
| Reproductive Disorders | [244,245,246,247,248,249,250,251] |
| Metabolic Disorders | [166,238,239,240] |
| Bone | [59,126,334,342] |
| Inflammation/Pain | [42,97,110,121,131,133,212,213,214,215,216,217,218,219] |
4.3.8. Exclusion of Papers with Image Duplication
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cunningham, K.; Manson, W.; Spring, F.; Hutchinson, S. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) Link. Nature 1950, 166, 949. [Google Scholar] [CrossRef]
- Winkler, D. Cordyceps sinensis—a precious parasitic fungus infecting Tibet. Field Mycol. 2010, 11, 60. [Google Scholar] [CrossRef]
- Penman, S.; Rosbash, M.; Penman, M. Messenger and heterogeneous nuclear RNA in HeLa cells: Differential inhibition by cordycepin. Proc. Natl. Acad. Sci. USA 1970, 67, 1878–1885. [Google Scholar] [CrossRef]
- Philipson, L.; Wall, R.; Glickman, G.; Darnell, J. Addition of polyadenylate sequences to virus-specific RNA during adenovirus replication. Proc. Natl. Acad. Sci. USA 1971, 68, 2806–2809. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Ting, R.; Paran, M.; Gallo, R. Cordycepin inhibits induction of murine leukovirus production by 5-iodo-2′-deoxyuridine. Proc. Natl. Acad. Sci. USA 1972, 69, 3820–3824. [Google Scholar] [CrossRef] [PubMed]
- Brawerman, G.; Diez, J. Metabolism of the polyadenylate sequence of nuclear RNA and messenger RNA in mammalian cells. Cell 1975, 5, 271–280. [Google Scholar] [CrossRef]
- Diez, J.; Brawerman, G. Elongation of the polyadenylate segment of messenger RNA in the cytoplasm of mammalian cells. Proc. Natl. Acad. Sci. USA 1974, 71, 4091–4095. [Google Scholar] [CrossRef] [PubMed]
- Lotem, J.; Sachs, L. Induction of specific changes in the surface membrane of myeloid leukemic cells by steroid hormones. Int. J. Cancer 1975, 15, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Price, P.J.; Suk, W.A.; Peters, R.L.; Martin, C.E.; Bellew, T.M.; Huebner, R.J. Cordycepin inhibition of 3-methylcholanthrene-induced transformation in vitro. Proc. Soc. Exp. Biol. Med. 1975, 150, 650–653. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.; Zhu, S.; Zhou, R.; Wang, L.; Du, J.; Wang, Y.; Zhou, B.; Mai, L. Cordycepin induces apoptosis and autophagy in human neuroblastoma SK-N-SH and BE (2)-M17 cells. Oncol. Lett. 2015, 9, 2541–2547. [Google Scholar] [CrossRef]
- Pan, B.-S.; Wang, Y.-K.; Lai, M.-S.; Mu, Y.-F.; Huang, B.-M. Cordycepin induced MA-10 mouse Leydig tumor cell apoptosis by regulating p38 MAPKs and PI3K/AKT signaling pathways. Sci. Rep. 2015, 5, 13372. [Google Scholar] [CrossRef]
- Yu, X.; Ling, J.; Liu, X.; Guo, S.; Lin, Y.; Liu, X.; Su, L. Cordycepin induces autophagy-mediated c-FLIPL degradation and leads to apoptosis in human non-small cell lung cancer cells. Oncotarget 2017, 8, 6691. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Kim, S.O.; Kim, G.-Y.; Moon, S.-K.; Kim, W.-J.; Jeong, Y.K.; Yoo, Y.H.; Choi, Y.H. Involvement of autophagy in cordycepin-induced apoptosis in human prostate carcinoma LNCaP cells. Environ. Toxicol. Pharmacol. 2014, 38, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Lim, M.-H.; Kim, K.M.; Jeon, B.H.; Song, W.O.; Kim, T.W. Cordycepin-induced apoptosis and autophagy in breast cancer cells are independent of the estrogen receptor. Toxicol. Appl. Pharmacol. 2011, 257, 165–173. [Google Scholar] [CrossRef]
- Chaicharoenaudomrung, N.; Jaroonwitchawan, T.; Noisa, P. Cordycepin induces apoptotic cell death of human brain cancer through the modulation of autophagy. Toxicol. Vitr. 2018, 46, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-Y.; Wu, W.-S.; Lin, L.-C.; Wu, Y.-H.; Chiu, H.-W.; Yeh, Y.-L.; Huang, B.-M.; Wang, Y.-J. Cordycepin enhances radiosensitivity in oral squamous carcinoma cells by inducing autophagy and apoptosis through cell cycle arrest. Int. J. Mol. Sci. 2019, 20, 5366. [Google Scholar] [CrossRef]
- Jang, H.-J.; Yang, K.E.; Hwang, I.-H.; Huh, Y.H.; Kim, D.J.; Yoo, H.-S.; Park, S.J.; Jang, I.-S. Cordycepin inhibits human ovarian cancer by inducing autophagy and apoptosis through Dickkopf-related protein 1/β-catenin signaling. Am. J. Transl. Res. 2019, 11, 6890. [Google Scholar]
- Wang, D.; Zhang, Y.; Lu, J.; Wang, Y.; Wang, J.; Meng, Q.; Lee, R.J.; Wang, D.; Teng, L. Cordycepin, a natural antineoplastic agent, induces apoptosis of breast cancer cells via caspase-dependent pathways. Nat. Prod. Commun. 2016, 11, 1934578X1601100119. [Google Scholar] [CrossRef]
- Lee, D.; Lee, W.-Y.; Jung, K.; Kwon, Y.S.; Kim, D.; Hwang, G.S.; Kim, C.-E.; Lee, S.; Kang, K.S. The inhibitory effect of cordycepin on the proliferation of MCF-7 breast cancer cells, and its mechanism: An investigation using network pharmacology-based analysis. Biomolecules 2019, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhang, M.-f.; Ye, J.; Jiang, T.-t.; Fang, X.; Song, Y. Cordycepin induces apoptosis by enhancing JNK and p38 kinase activity and increasing the protein expression of Bcl-2 pro-apoptotic molecules. J. Zhejiang Univ. Sci. B 2010, 11, 654–660. [Google Scholar] [CrossRef]
- Lee, H.H.; Park, C.; Jeong, J.-W.; Kim, M.J.; Seo, M.J.; Kang, B.W.; Park, J.U.; Kim, G.-Y.; Choi, B.T.; Choi, Y.H. Apoptosis induction of human prostate carcinoma cells by cordycepin through reactive oxygen species-mediated mitochondrial death pathway. Int. J. Oncol. 2013, 42, 1036–1044. [Google Scholar] [CrossRef]
- Baik, J.-S.; Mun, S.-W.; Kim, K.-S.; Park, S.-J.; Yoon, H.-K.; Kim, D.-H.; Park, M.-K.; Kim, C.-H.; Lee, Y.-C. Apoptotic effects of cordycepin through the extrinsic pathway and p38 MAPK activation in human glioblastoma U87MG cells. J. Microbiol. Biotechnol. 2016, 26, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Su, N.-W.; Wu, S.-H.; Chi, C.-W.; Liu, C.-J.; Tsai, T.-H.; Chen, Y.-J. Metronomic cordycepin therapy prolongs survival of oral cancer-bearing mice and inhibits epithelial-mesenchymal transition. Molecules 2017, 22, 629. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-A.; Xiang, S.-S.; Li, H.-F.; Wu, X.-S.; Li, M.-L.; Shu, Y.-J.; Zhang, F.; Cao, Y.; Ye, Y.-Y.; Bao, R.-F. Cordycepin induces S phase arrest and apoptosis in human gallbladder cancer cells. Molecules 2014, 19, 11350–11365. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Jin, C.-Y.; Park, C.; Hong, S.H.; Kim, G.-Y.; Jeong, Y.K.; Lee, J.-D.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis by cordycepin via reactive oxygen species generation in human leukemia cells. Toxicol. Vitr. 2011, 25, 817–824. [Google Scholar] [CrossRef]
- Jang, K.-J.; Kwon, G.-S.; Jeong, J.-W.; Kim, C.-H.; Yoon, H.-M.; Kim, G.-Y.; Shim, J.-H.; Moon, S.-K.; Kim, W.-J.; Choi, Y.H. Cordyceptin induces apoptosis through repressing hTERT expression and inducing extranuclear export of hTERT. J. Biosci. Bioeng. 2015, 119, 351–357. [Google Scholar] [CrossRef]
- Jen, C.-Y.; Lin, C.-Y.; Huang, B.-M.; Leu, S.-F. Cordycepin induced MA-10 mouse Leydig tumor cell apoptosis through caspase-9 pathway. Evid. -Based Complementary Altern. Med. 2011, 2011, 984537. [Google Scholar] [CrossRef]
- Cheng, C.; Zhu, X. Cordycepin mitigates MPTP-induced Parkinson’s disease through inhibiting TLR/NF-κB signaling pathway. Life Sci. 2019, 223, 120–127. [Google Scholar] [CrossRef]
- Comella, J.X.; Sanz-Rodriguez, C.; Aldea, M.; Esquerda, J.E. Skeletal muscle-derived trophic factors prevent motoneurons from entering an active cell death program in vitro. J. Neurosci. 1994, 14, 2674–2686. [Google Scholar] [CrossRef]
- D’Arpa, P.; Beardmore, C.; Liu, L.F. Involvement of nucleic acid synthesis in cell killing mechanisms of topoisomerase poisons. Cancer Res. 1990, 50, 6919–6924. [Google Scholar] [PubMed]
- Gu, L.; Johno, H.; Nakajima, S.; Kato, H.; Takahashi, S.; Katoh, R.; Kitamura, M. Blockade of Smad signaling by 3′-deoxyadenosine: A mechanism for its anti-fibrotic potential. Lab. Investig. 2013, 93, 450–461. [Google Scholar] [CrossRef]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Oh, J.-I.; Lee, S.J.; Park, G. The neuroprotective effects of cordycepin inhibit glutamate-induced oxidative and ER stress-associated apoptosis in hippocampal HT22 cells. Neurotoxicology 2014, 41, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Kan, H.; Wang, Y.; Wang, D.; Sun, H.; Zhou, S.; Wang, H.; Guan, J.; Li, M. Cordycepin rescues lidocaine-induced neurotoxicity in dorsal root ganglion by interacting with inflammatory signaling pathway MMP3. Eur. J. Pharmacol. 2018, 827, 88–93. [Google Scholar] [CrossRef]
- Kaufmann, S.H. Antagonism between camptothecin and topoisomerase II-directed chemotherapeutic agents in a human leukemia cell line. Cancer Res. 1991, 51, 1129–1136. [Google Scholar]
- Kitamura, M.; Kato, H.; Saito, Y.; Nakajima, S.; Takahashi, S.; Johno, H.; Gu, L.; Katoh, R. Aberrant, differential and bidirectional regulation of the unfolded protein response towards cell survival by 3’-deoxyadenosine. Cell Death Differ 2011, 18, 1876–1888. [Google Scholar] [CrossRef]
- Li, L.; He, D.; Yang, J.; Wang, X. Cordycepin inhibits renal interstitial myofibroblast activation probably by inducing hepatocyte growth factor expression. J. Pharmacol. Sci. 2011, 1111300626. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Y.; Zhang, X.; Cao, X.; Wu, C.; Guo, P. Cordycepin stimulates autophagy in macrophages and prevents atherosclerotic plaque formation in ApoE-/-mice. Oncotarget 2017, 8, 94726. [Google Scholar] [CrossRef] [PubMed]
- Marcelo, A.; Brito, F.; Carmo-Silva, S.; Matos, C.A.; Alves-Cruzeiro, J.; Vasconcelos-Ferreira, A.; Koppenol, R.; Mendonça, L.; de Almeida, L.P.; Nóbrega, C. Cordycepin activates autophagy through AMPK phosphorylation to reduce abnormalities in Machado–Joseph disease models. Hum. Mol. Genet. 2019, 28, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Lawson, P.A.; Ralph, R.K. Inhibition of protein synthesis reduces the cytotoxicity of 4’-(9-acridinylamino) methane-sulfon-m-anisidide without affecting DNA breakage and DNA topoisomerase ii in a murine mastocytoma cell line. Biochem. Pharmacol. 1989, 38, 263–269. [Google Scholar] [CrossRef]
- Song, H.; Huang, L.-P.; Li, Y.; Liu, C.; Wang, S.; Meng, W.; Wei, S.; Liu, X.-P.; Gong, Y.; Yao, L.-H. Neuroprotective effects of cordycepin inhibit Aβ-induced apoptosis in hippocampal neurons. Neurotoxicology 2018, 68, 73–80. [Google Scholar] [CrossRef]
- Wotring, L.L.; Townsend, L.B. Identification of 6-azauridine triphosphate in l1210 cells and its possible relevance to cytotoxicity. Cancer Res. 1989, 49, 289–294. [Google Scholar] [PubMed]
- Xia, C.; Chen, P.; Mei, S.; Ning, L.; Lei, C.; Wang, J.; Zhang, J.; Ma, J.; Fan, S. Photo-crosslinked HAMA hydrogel with cordycepin encapsulated chitosan microspheres for osteoarthritis treatment. Oncotarget 2017, 8, 2835. [Google Scholar] [CrossRef]
- Yang, J.; Cao, Y.; Lv, Z.; Jiang, T.; Wang, L.; Li, Z. Cordycepin protected against the TNF-α-induced inhibition of osteogenic differentiation of human adipose-derived mesenchymal stem cells. Int. J. Immunopathol. Pharmacol. 2015, 28, 296–307. [Google Scholar] [CrossRef]
- Li, T.; Wen, L.; Cheng, B. Cordycepin alleviates hepatic lipid accumulation by inducing protective autophagy via PKA/mTOR pathway. Biochem. Biophys. Res. Commun. 2019, 516, 632–638. [Google Scholar] [CrossRef]
- Charlesworth, A.; Meijer, H.A.; de Moor, C.H. Specificity factors in cytoplasmic polyadenylation. Wiley Interdiscip. Rev. Rna 2013, 4, 437–461. [Google Scholar] [CrossRef] [PubMed]
- Kuge, H.; Inoue, A. Maturation of Xenopus laevis oocyte by progesterone requires poly (A) tail elongation of mRNA. Exp. Cell Res. 1992, 202, 52–58. [Google Scholar] [CrossRef]
- Nagahama, Y. 17α, 20β-Dihydroxy-4-pregnen-3-one, a maturation-inducing hormone in fish oocytes: Mechanisms of synthesis and action. Steroids 1997, 62, 190–196. [Google Scholar] [CrossRef]
- Barkoff, A.; Ballantyne, S.; Wickens, M. Meiotic maturation in Xenopus requires polyadenylation of multiple mRNAs. Embo J. 1998, 17, 3168–3175. [Google Scholar] [CrossRef] [PubMed]
- Katsu, Y.; Yamashita, M.; Nagahama, Y. Translational regulation of cyclin B mRNA by 17α, 20β-dihydroxy-4-pregnen-3-one (maturation-inducing hormone) during oocyte maturation in a teleost fish, the goldfish (Carassius auratus). Mol. Cell. Endocrinol. 1999, 158, 79–85. [Google Scholar] [CrossRef]
- Fuchimoto, D.-i.; Mizukoshi, A.; Schultz, R.M.; Sakai, S.; Aoki, F. Posttranscriptional regulation of cyclin A1 and cyclin A2 during mouse oocyte meiotic maturation and preimplantation development. Biol. Reprod. 2001, 65, 986–993. [Google Scholar] [CrossRef]
- Faerge, I.; Terry, B.; Kalous, J.; Wahl, P.; Lessl, M.; Ottesen, J.; Hyttel, P.; Grøndahl, C. Resumption of meiosis induced by meiosis-activating sterol has a different signal transduction pathway than spontaneous resumption of meiosis in denuded mouse oocytes cultured in vitro. Biol. Reprod. 2001, 65, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Traverso, J.M.; Donnay, I.; Lequarre, A.S. Effects of polyadenylation inhibition on meiosis progression in relation to the polyadenylation status of cyclins A2 and B1 during in vitro maturation of bovine oocytes. Mol. Reprod. Dev. Inc. Gamete Res. 2005, 71, 107–114. [Google Scholar] [CrossRef][Green Version]
- Hara, K.T.; Oda, S.; Naito, K.; Nagata, M.; Schultz, R.M.; Aoki, F. Cyclin A2-CDK2 regulates embryonic gene activation in 1-cell mouse embryos. Dev. Biol. 2005, 286, 102–113. [Google Scholar] [CrossRef]
- Gershon, E.; Galiani, D.; Dekel, N. Cytoplasmic polyadenylation controls cdc25B mRNA translation in rat oocytes resuming meiosis. Reproduction 2006, 132, 21–31. [Google Scholar] [CrossRef]
- Zhang, D.X.; Cui, X.S.; Kim, N.H. Involvement of polyadenylation status on maternal gene expression during in vitro maturation of porcine oocytes. Mol. Reprod. Dev. 2009, 76, 881–889. [Google Scholar] [CrossRef]
- Dobbs, K.B.; Spollen, W.G.; Springer, G.; Prather, R.S. The role of cytoplasmic polyadenylation element sequence on mRNA abundance during porcine embryogenesis and parthenogenetic development. Mol. Reprod. Dev. 2010, 77, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gao, Y.; Zhai, B.; Jiang, H.; Ding, Y.; Zhang, L.; Li, C.; Deng, Q.; Yu, X.; Zhang, J. The Effects of polyadenylation status on MPFs during in vitro porcine oocyte maturation. Cell. Physiol. Biochem. 2016, 39, 1735–1745. [Google Scholar] [CrossRef]
- Nair, R.; Victor, A.C.; Paul, V.; Paul-Prasanth, B. Effects of N-Nitrosodiethylamine, a Potent Carcinogen, on Sexual Development, Gametogenesis, and Oocyte Maturation. Sex. Dev. 2017, 11, 161–167. [Google Scholar] [CrossRef]
- Krischek, C.; Meinecke, B. In vitro maturation of bovine oocytes requires polyadenylation of mRNAs coding proteins for chromatin condensation, spindle assembly, MPF and MAP kinase activation. Anim. Reprod. Sci. 2002, 73, 129–140. [Google Scholar] [CrossRef]
- Novoa, I.; Gallego, J.; Ferreira, P.G.; Mendez, R. Mitotic cell-cycle progression is regulated by CPEB1 and CPEB4-dependent translational control. Nat. Cell Biol. 2010, 12, 447–456. [Google Scholar] [CrossRef]
- Osborn, J.; Moor, R. Time-dependent effects of α-amanitin on nuclear maturation and protein synthesis in mammalian oocytes. J. Embryol. Exp. Morphol. 1983, 73, 317–338. [Google Scholar] [PubMed]
- Park, J.-E.; Yi, H.; Kim, Y.; Chang, H.; Kim, V.N. Regulation of poly (A) tail and translation during the somatic cell cycle. Mol. Cell 2016, 62, 462–471. [Google Scholar] [CrossRef]
- Čermák, V.; Dostál, V.; Jelínek, M.; Libusová, L.; Kovář, J.; Rösel, D.; Brábek, J. Microtubule-targeting agents and their impact on cancer treatment. Eur. J. Cell Biol. 2020, 99, 151075. [Google Scholar] [CrossRef] [PubMed]
- Su, N.-W.; Wu, S.-H.; Chi, C.-W.; Tsai, T.-H.; Chen, Y.-J. Cordycepin, isolated from medicinal fungus Cordyceps sinensis, enhances radiosensitivity of oral cancer associated with modulation of DNA damage repair. Food Chem. Toxicol. 2019, 124, 400–410. [Google Scholar] [CrossRef]
- Thomadaki, H.; Tsiapalis, C.M.; Scorilas, A. The effect of the polyadenylation inhibitor cordycepin on human Molt-4 and Daudi leukaemia and lymphoma cell lines. Cancer Chemother. Pharmacol. 2008, 61, 703–711. [Google Scholar] [CrossRef]
- Wei, C.; Yao, X.; Jiang, Z.; Wang, Y.; Zhang, D.; Chen, X.; Fan, X.; Xie, C.; Cheng, J.; Fu, J. Cordycepin inhibits drug-resistance non-small cell lung cancer progression by activating AMPK signaling pathway. Pharmacol. Res. 2019, 144, 79–89. [Google Scholar] [CrossRef]
- Tomasovic, S.P.; Dewey, W.C. Acceleration of CHO cells into mitosis and reduction of X-ray-induced G2 delay by cordycepin. Exp. Cell Res. 1978, 114, 277–284. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Jin, C.-Y.; Park, C.; Han, M.H.; Kim, G.-Y.; Moon, S.-K.; Kim, C.G.; Jeong, Y.K.; Kim, W.-J.; Lee, J.-D. Inhibition of migration and invasion of LNCaP human prostate carcinoma cells by cordycepin through inactivation of Akt. Int. J. Oncol. 2012, 40, 1697–1704. [Google Scholar]
- Jeong, J.-W.; Park, C.; Cha, H.-J.; Hong, S.H.; Park, S.-H.; Kim, G.-Y.; Kim, W.J.; Kim, C.H.; Song, K.S.; Choi, Y.H. Cordycepin inhibits lipopolysaccharide-induced cell migration and invasion in human colorectal carcinoma HCT-116 cells through down-regulation of prostaglandin E2 receptor EP4. Bmb Rep. 2018, 51, 532. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, W.J.; Moon, S.K. Cordycepin suppresses TNF-alpha-induced invasion, migration and matrix metalloproteinase-9 expression in human bladder cancer cells. Phytother. Res. 2010, 24, 1755–1761. [Google Scholar] [CrossRef]
- Li, Y.; Li, K.; Mao, L.; Han, X.; Zhang, K.; Zhao, C.; Zhao, J. Cordycepin inhibits LPS-induced inflammatory and matrix degradation in the intervertebral disc. PeerJ 2016, 4, e1992. [Google Scholar] [CrossRef]
- Noh, E.-M.; Jung, S.H.; Han, J.-H.; Chung, E.-Y.; Jung, J.-Y.; Kim, B.-S.; Lee, S.-H.; Lee, Y.-R.; Kim, J.-S. Cordycepin inhibits TPA-induced matrix metalloproteinase-9 expression by suppressing the MAPK/AP-1 pathway in MCF-7 human breast cancer cells. Int. J. Mol. Med. 2010, 25, 255–260. [Google Scholar]
- Wang, Y.; Lv, Y.; Liu, T.S.; Di Yan, W.; Chen, L.Y.; Li, Z.H.; Piao, Y.S.; An, R.B.; Lin, Z.H.; Ren, X.S. Cordycepin suppresses cell proliferation and migration by targeting CLEC2 in human gastric cancer cells via Akt signaling pathway. Life Sci. 2019, 223, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Huang, C.; Fu, C.; Tian, Y.; Hu, Y.; Wang, B.; Strasner, A.; Song, Y.; Song, E. Cordycepin (3′-deoxyadenosine) suppressed HMGA2, Twist1 and ZEB1-dependent melanoma invasion and metastasis by targeting miR-33b. Oncotarget 2015, 6, 9834. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Stamenkovic, I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-β and promotes tumor invasion and angiogenesis. Genes Dev. 2000, 14, 163–176. [Google Scholar] [PubMed]
- Chang, W.; Lim, S.; Song, H.; Song, B.-W.; Kim, H.-J.; Cha, M.-J.; Sung, J.M.; Kim, T.W.; Hwang, K.-C. Cordycepin inhibits vascular smooth muscle cell proliferation. Eur. J. Pharmacol. 2008, 597, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Ning, Y.; Zhao, X.; Pan, T. The effects of cordycepin on the cell proliferation, migration and apoptosis in human lung cancer cell lines A549 and NCI-H460. J. Pharm. Pharmacol. 2016, 68, 901–911. [Google Scholar] [CrossRef]
- Cao, Z.; Dou, C.; Li, J.; Tang, X.; Xiang, J.; Zhao, C.; Zhu, L.; Bai, Y.; Xiang, Q.; Dong, S. Cordycepin inhibits chondrocyte hypertrophy of mesenchymal stem cells through PI3K/Bapx1 and Notch signaling pathway. Bmb Rep. 2016, 49, 548. [Google Scholar] [CrossRef]
- Hueng, D.-Y.; Hsieh, C.-H.; Cheng, Y.-C.; Tsai, W.-C.; Chen, Y. Cordycepin inhibits migration of human glioblastoma cells by affecting lysosomal degradation and protein phosphatase activation. J. Nutr. Biochem. 2017, 41, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-T.; Liang, S.-M.; Wu, Y.-J.; Wu, Y.-J.; Lu, Y.-J.; Jan, Y.-J.; Ko, B.-S.; Chuang, Y.-J.; Shyue, S.-K.; Kuo, C.-C. Cordycepin suppresses endothelial cell proliferation, migration, angiogenesis, and tumor growth by regulating focal adhesion kinase and p53. Cancers 2019, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.-L.; Ko, B.-S.; Liu, T.-A.; Liang, S.-M.; Liu, C.-C.; Lu, Y.-J.; Tzean, S.-S.; Shen, T.-L.; Liou, J.-Y. Cordycepin suppresses integrin/FAK signaling and epithelial-mesenchymal transition in hepatocellular carcinoma. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. -Anti-Cancer Agents) 2014, 14, 29–34. [Google Scholar] [CrossRef]
- Mitchell, J.P.; Carmody, R.J. NF-κB and the transcriptional control of inflammation. Int. Rev. Cell Mol. Biol. 2018, 335, 41–84. [Google Scholar]
- Khandia, R.; Munjal, A. Interplay between inflammation and cancer. Adv. Protein Chem. Struct. Biol. 2020, 119, 199–245. [Google Scholar]
- Stewart, A.G.; Thomas, B.; Koff, J. TGF-β: Master regulator of inflammation and fibrosis. Respirology 2018, 23, 1096–1097. [Google Scholar] [CrossRef]
- Ji, R.-R.; Chamessian, A.; Zhang, Y.-Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: The role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 2015, 16, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T. Ageing, neurodegeneration and brain rejuvenation. Nature 2016, 539, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Park, S.J.; Ko, W.G.; Kang, S.-M.; Lee, D.B.; Bang, J.; Park, B.-J.; Wee, C.-B.; Kim, D.J.; Jang, I.-S. Cordycepin induces human lung cancer cell apoptosis by inhibiting nitric oxide mediated ERK/Slug signaling pathway. Am. J. Cancer Res. 2017, 7, 417. [Google Scholar]
- Zhou, X.; Luo, L.; Dressel, W.; Shadier, G.; Krumbiegel, D.; Schmidtke, P.; Zepp, F.; Meyer, C.U. Cordycepin is an immunoregulatory active ingredient of Cordyceps sinensis. Am. J. Chin. Med. 2008, 36, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, M.K.; Otero-Vinas, M.; Falanga, V. Transforming growth factor beta (TGF-β) isoforms in wound healing and fibrosis. Wound Repair Regen. 2016, 24, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Finnson, K.W.; Chi, Y.; Bou-Gharios, G.; Leask, A.; Philip, A. TGF-β signaling in cartilage homeostasis and osteoarthritis. Front Biosci 2012, 4, 251–268. [Google Scholar] [CrossRef]
- Meng, X.-m.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.-G.; Yuan, Y.-P.; Wu, H.-M.; Zhang, X.; Tang, Q.-Z. Cardiac fibrosis: New insights into the pathogenesis. Int. J. Biol. Sci. 2018, 14, 1645. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cheung, F.W.; Chan, M.H.; Hui, P.K.; Ip, S.-P.; Ling, Y.H.; Che, C.-T.; Liu, W.K. Protective roles of Cordyceps on lung fibrosis in cellular and rat models. J. Ethnopharmacol. 2012, 143, 448–454. [Google Scholar] [CrossRef]
- Gu, L.; Johno, H.; Nakajima, S.; Yoshitomi, T.; Takahashi, S.; Kitamura, M. Intervention in Genotoxic Stress–Induced Senescence by Cordycepin Through Activation of eIF2α and Suppression of Sp1. Toxicol. Sci. 2013, 134, 345–354. [Google Scholar] [CrossRef][Green Version]
- Wang, C.-W.; Lee, B.-H.; Tai, C.-J. The inhibition of cordycepin on cancer stemness in TGF-beta induced chemo-resistant ovarian cancer cell. Oncotarget 2017, 8, 111912. [Google Scholar] [CrossRef]
- Ashraf, S.; Radhi, M.; Gowler, P.; Burston, J.J.; Gandhi, R.D.; Thorn, G.J.; Piccinini, A.M.; Walsh, D.A.; Chapman, V.; De Moor, C.H. The polyadenylation inhibitor cordycepin reduces pain, inflammation and joint pathology in rodent models of osteoarthritis. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Kim, G.-Y.; Lee, H.H. Anti-inflammatory effects of cordycepin in lipopolysaccharide-stimulated RAW 264.7 macrophages through Toll-like receptor 4-mediated suppression of mitogen-activated protein kinases and NF-κB signaling pathways. Drug Des. Dev. Ther. 2014, 8, 1941. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.Y.; Park, S.J.; Jo, E.; Hwang, I.-H.; Lee, K.-B.; Kim, S.-W.; Kim, D.J.; Joo, J.C.; Hong, S.H.; Lee, M.-G. Cordycepin induces apoptosis of human ovarian cancer cells by inhibiting CCL5-mediated Akt/NF-κB signaling pathway. Cell Death Discov. 2018, 4, 1–11. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Jin, C.-Y.; Kim, G.-Y.; Lee, J.-D.; Park, C.; Kim, G.-D.; Kim, W.-J.; Jung, W.-K.; Seo, S.K.; Choi, I.-W. Anti-inflammatory effects of cordycepin via suppression of inflammatory mediators in BV2 microglial cells. Int. Immunopharmacol. 2010, 10, 1580–1586. [Google Scholar] [CrossRef]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.-J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H.I. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef]
- Peng, J.; Wang, P.; Ge, H.; Qu, X.; Jin, X. Effects of cordycepin on the microglia-overactivation-induced impairments of growth and development of hippocampal cultured neurons. PLoS ONE 2015, 10, e0125902. [Google Scholar] [CrossRef]
- Yan, L.J.; Yang, H.T.; Duan, H.Y.; Wu, J.T.; Qian, P.; Fan, X.W.; Wang, S. Cordycepin inhibits vascular adhesion molecule expression in TNF-α-stimulated vascular muscle cells. Exp. Ther. Med. 2017, 14, 2335–2340. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ying, X.; Peng, L.; Chen, H.; Shen, Y.; Yu, K.; Cheng, S. Cordycepin prevented IL-β-induced expression of inflammatory mediators in human osteoarthritis chondrocytes. Int. Orthop. 2014, 38, 1519–1526. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, W.; Dai, G.; Huang, Y. Cordycepin suppresses the migration and invasion of human liver cancer cells by downregulating the expression of CXCR4. Int. J. Mol. Med. 2020, 45, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, A.; Meijer, H.A.; Barthet-Barateig, A.; Parker, H.N.; Khurshid, A.; Tessier, S.; Sicard, M.; Knox, A.J.; Pang, L.; De Moor, C.H. Inhibition of polyadenylation reduces inflammatory gene induction. Rna 2012, 18, 2236–2250. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.-H.; Oh, S.Y.; Jang, H.-J.; Jo, E.; Joo, J.C.; Lee, K.-B.; Yoo, H.-S.; Lee, M.Y.; Park, S.J.; Jang, I.-S. Cordycepin promotes apoptosis in renal carcinoma cells by activating the MKK7-JNK signaling pathway through inhibition of c-FLIPL expression. PLoS ONE 2017, 12, e0186489. [Google Scholar] [CrossRef]
- Liang, S.-M.; Lu, Y.-J.; Ko, B.-S.; Jan, Y.-J.; Shyue, S.-K.; Yet, S.-F.; Liou, J.-Y. Cordycepin disrupts leukemia association with mesenchymal stromal cells and eliminates leukemia stem cell activity. Sci. Rep. 2017, 7, 43930. [Google Scholar] [CrossRef]
- Sun, T.; Dong, W.; Jiang, G.; Yang, J.; Liu, J.; Zhao, L.; Ma, P. Cordyceps militaris improves chronic kidney disease by affecting TLR4/NF-κB redox signaling pathway. Oxidative Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, Y.; Shi, J. Cordycepin protects against acute pancreatitis by modulating NF-κB and NLRP3 inflammasome activation via AMPK. Life Sci. 2020, 117645. [Google Scholar] [CrossRef]
- Sun, J.; Jin, M.; Zhou, W.; Diao, S.; Zhou, Y.; Li, S.; Wang, X.; Pan, S.; Jin, X.; Li, G. A new ribonucleotide from Cordyceps militaris. Nat. Prod. Res. 2017, 31, 2537–2543. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.; Kang, K.S.; Chun, K.-H.; Hwang, G.S. Cordyceps militaris mushroom and cordycepin inhibit RANKL-induced osteoclast differentiation. J. Med. Food 2015, 18, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Xu, Y.; Shen, J. Cordycepin inhibits lipopolysaccharide (LPS)-induced tumor necrosis factor (TNF)-alpha production via activating amp-activated protein kinase (AMPK) signaling. Int. J. Mol. Sci. 2014, 15, 12119–12134. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-S.; Kim, K.-S.; Moon, H.-I.; An, H.-K.; Park, S.-J.; Kim, C.-H.; Lee, Y.-C. Cordycepin-mediated transcriptional regulation of human GD3 synthase (hST8Sia I) in human neuroblastoma SK-N-BE (2)-C cells. Acta Biochim Biophys Sin 2014, 46, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Han, M.W.; Ryu, I.S.; Lee, J.C.; Kim, S.H.; Chang, H.W.; Lee, Y.S.; Lee, S.; Kim, S.W.; Kim, S.Y. Phosphorylation of PI3K regulatory subunit p85 contributes to resistance against PI3K inhibitors in radioresistant head and neck cancer. Oral Oncol. 2018, 78, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, H.; Bao, H.; Zhang, D.; Feng, L.; Xiao, Y.; Zhu, K.; Hou, Y.; Luo, S.; Zhang, Y. Cordycepin (3′-deoxyadenosine) promotes remyelination via suppression of neuroinflammation in a cuprizone-induced mouse model of demyelination. Int. Immunopharmacol. 2019, 75, 105777. [Google Scholar] [CrossRef]
- Kim, H.; Naura, A.S.; Errami, Y.; Ju, J.; Boulares, A.H. Cordycepin blocks lung injury-associated inflammation and promotes BRCA1-deficient breast cancer cell killing by effectively inhibiting PARP. Mol. Med. 2011, 17, 893–900. [Google Scholar] [CrossRef]
- Qing, R.; Huang, Z.; Tang, Y.; Xiang, Q.; Yang, F. Cordycepin alleviates lipopolysaccharide-induced acute lung injury via Nrf2/HO-1 pathway. Int. Immunopharmacol. 2018, 60, 18–25. [Google Scholar] [CrossRef]
- Tianzhu, Z.; Shihai, Y.; Juan, D. Antidepressant-like effects of cordycepin in a mice model of chronic unpredictable mild stress. Evid. -Based Complementary Altern. Med. 2014, 2014, 438506. [Google Scholar] [CrossRef]
- Wang, M.-j.; Peng, X.-y.; Lian, Z.-q.; Zhu, H.-b. The cordycepin derivative IMM-H007 improves endothelial dysfunction by suppressing vascular inflammation and promoting AMPK-dependent eNOS activation in high-fat diet-fed ApoE knockout mice. Eur. J. Pharmacol. 2019, 852, 167–178. [Google Scholar] [CrossRef]
- Won, S.-Y.; Park, E.-H. Anti-inflammatory and related pharmacological activities of cultured mycelia and fruiting bodies of Cordyceps militaris. J. Ethnopharmacol. 2005, 96, 555–561. [Google Scholar] [CrossRef]
- Okur, M.H.; Arslan, S.; Aydogdu, B.; Zeytun, H.; Basuguy, E.; Arslan, M.S.; Ibiloglu, I.; Kaplan, I. Protective effect of cordycepin on experimental testicular ischemia/reperfusion injury in rats. J. Investig. Surg. 2018, 31, 1–8. [Google Scholar] [CrossRef]
- Tianzhu, Z.; Shihai, Y.; Juan, D. The effects of cordycepin on ovalbumin-induced allergic inflammation by strengthening Treg response and suppressing Th17 responses in ovalbumin-sensitized mice. Inflammation 2015, 38, 1036–1043. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; He, Y.; Li, T.; Wang, W.; Zhang, J.; Wei, J.; Deng, Y.; Lin, R. Cordycepin alleviates airway hyperreactivity in a murine model of asthma by attenuating the inflammatory process. Int. Immunopharmacol. 2015, 26, 401–408. [Google Scholar] [CrossRef]
- Hung, Y.-P.; Lee, C.-L. Higher anti-liver fibrosis effect of cordyceps militaris-fermented product cultured with deep ocean water via inhibiting proinflammatory factors and fibrosis-related factors expressions. Mar. Drugs 2017, 15, 168. [Google Scholar] [CrossRef]
- Zhang, D.-w.; Wang, Z.-l.; Qi, W.; Lei, W.; Zhao, G.-y. Cordycepin (3′-deoxyadenosine) down-regulates the proinflammatory cytokines in inflammation-induced osteoporosis model. Inflammation 2014, 37, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhong, L.; Zhu, H.; Wang, F. The protective effect of cordycepin on D-galactosamine/lipopolysaccharide-induced acute liver injury. Mediat. Inflamm. 2017, 2017, 3946706. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Zhang, X.; Zhang, G.-q.; Bao, W.-p.; Zhang, Y.-y.; Zhang, M.; Zhou, X. Cordycepin inhibits airway remodeling in a rat model of chronic asthma. Biomed. Pharmacother. 2017, 88, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Wei, Y.; Song, P.; Li, Y.; Zhang, T.; Feng, Q.; Xu, G. Cordycepin inhibits LPS-induced acute lung injury by inhibiting inflammation and oxidative stress. Eur. J. Pharmacol. 2018, 818, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Rottenberg, M.E.; Masocha, W.; Ferella, M.; Petitto-Assis, F.; Goto, H.; Kristensson, K.; McCaffrey, R.; Wigzell, H. Treatment of African trypanosomiasis with cordycepin and adenosine deaminase inhibitors in a mouse model. J. Infect. Dis. 2005, 192, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xi, D.; Mo, J.; Wang, K.; Luo, Y.; Xia, E.; Huang, R.; Luo, S.; Wei, J.; Ren, Z. Cordycepin exhibits a suppressive effect on T cells through inhibiting TCR signaling cascade in CFA-induced inflammation mice model. Immunopharmacol. Immunotoxicol. 2020, 42, 119–127. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, S.; Xu, L.; Song, B.; Huang, G.; Lu, J.; Guan, S. Suppression of T-cell activation in vitro and in vivo by cordycepin from Cordyceps militaris. J. Surg. Res. 2013, 185, 912–922. [Google Scholar] [CrossRef]
- Yang, R.; Wang, X.; Xi, D.; Mo, J.; Wang, K.; Luo, S.; Wei, J.; Ren, Z.; Pang, H.; Luo, Y. Cordycepin Attenuates IFN-γ-Induced Macrophage IP-10 and Mig Expressions by Inhibiting STAT1 Activity in CFA-Induced Inflammation Mice Model. Inflammation 2020, 43, 752–764. [Google Scholar] [CrossRef]
- Zeng, Y.; Lian, S.; Li, D.; Lin, X.; Chen, B.; Wei, H.; Yang, T. Anti-hepatocarcinoma effect of cordycepin against NDEA-induced hepatocellular carcinomas via the PI3K/Akt/mTOR and Nrf2/HO-1/NF-κB pathway in mice. Biomed. Pharmacother. 2017, 95, 1868–1875. [Google Scholar] [CrossRef]
- Gong, X.; Li, T.; Wan, R.; Sha, L. Cordycepin attenuates high-fat diet-induced non-alcoholic fatty liver disease via down-regulation of lipid metabolism and inflammatory responses. Int. Immunopharmacol. 2021, 91, 107173. [Google Scholar] [CrossRef]
- Chamcheu, J.C.; Roy, T.; Uddin, M.B.; Banang-Mbeumi, S.; Chamcheu, R.-C.N.; Walker, A.L.; Liu, Y.-Y.; Huang, S. Role and therapeutic targeting of the PI3K/Akt/mTOR signaling pathway in skin cancer: A review of current status and future trends on natural and synthetic agents therapy. Cells 2019, 8, 803. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhang, L.; Zhang, Q.; Li, X.; Xia, X.-J.; Liu, Y.-Y.; Yang, Q.-S. Lentiviral vector-mediated SHC3 silencing exacerbates oxidative stress injury in nigral dopamine neurons by regulating the PI3K-AKT-FoxO signaling pathway in rats with Parkinson’s disease. Cell. Physiol. Biochem. 2018, 49, 971–984. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483. [Google Scholar] [CrossRef]
- Riquelme, I.; Tapia, O.; Espinoza, J.A.; Leal, P.; Buchegger, K.; Sandoval, A.; Bizama, C.; Araya, J.C.; Peek, R.M.; Roa, J.C. The gene expression status of the PI3K/AKT/mTOR pathway in gastric cancer tissues and cell lines. Pathol. Oncol. Res. 2016, 22, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.J.; Sharma, A.; Mair, W.B. Metabolic Communication and Healthy Aging: Where Should We Focus Our Energy? Dev. Cell 2020, 54, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Chiang, G.G.; Abraham, R.T. Phosphorylation of mammalian target of rapamycin (mTOR) at Ser-2448 is mediated by p70S6 kinase. J Biol Chem 2005, 280, 25485–25490. [Google Scholar] [CrossRef]
- Bi, Y.; Li, H.; Yi, D.; Sun, Y.; Bai, Y.; Zhong, S.; Song, Y.; Zhao, G.; Chen, Y. Cordycepin augments the chemosensitivity of human glioma cells to temozolomide by activating AMPK and inhibiting the AKT signaling pathway. Mol. Pharm. 2018, 15, 4912–4925. [Google Scholar] [CrossRef]
- Hsu, P.-Y.; Lin, Y.-H.; Yeh, E.-L.; Lo, H.-C.; Hsu, T.-H.; Su, C.-C. Cordycepin and a preparation from Cordyceps militaris inhibit malignant transformation and proliferation by decreasing EGFR and IL-17RA signaling in a murine oral cancer model. Oncotarget 2017, 8, 93712. [Google Scholar] [CrossRef]
- Takahashi, S.; Tamai, M.; Nakajima, S.; Kato, H.; Johno, H.; Nakamura, T.; Kitamura, M. Blockade of adipocyte differentiation by cordycepin. Br. J. Pharmacol. 2012, 167, 561–575. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Jiang, Z.; Luo, P.; Liu, L.; Huang, Y.; Wang, H.; Wang, Y.; Long, L.; Tan, X. Cordycepin prevents radiation ulcer by inhibiting cell senescence via NRF2 and AMPK in rodents. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Wu, W.-D.; Hu, Z.-M.; Shang, M.-J.; Zhao, D.-J.; Zhang, C.-W.; Hong, D.-F.; Huang, D.-S. Cordycepin down-regulates multiple drug resistant (MDR)/HIF-1α through regulating AMPK/mTORC1 signaling in GBC-SD gallbladder cancer cells. Int. J. Mol. Sci. 2014, 15, 12778–12790. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.C.; Madison, V. AKT crystal structure and AKT-specific inhibitors. Oncogene 2005, 24, 7493–7501. [Google Scholar] [CrossRef]
- Wei, Y.; Zhou, J.; Yu, H.; Jin, X. AKT phosphorylation sites of Ser473 and Thr308 regulate AKT degradation. Biosci Biotechnol Biochem 2019, 83, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.S.; Lu, Y.J.; Yao, W.L.; Liu, T.A.; Tzean, S.S.; Shen, T.L.; Liou, J.Y. Cordycepin regulates GSK-3beta/beta-catenin signaling in human leukemia cells. PLoS ONE 2013, 8, e76320. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mo, H.; Gu, J.; Chen, K.; Han, Z.; Liu, Y. Cordycepin induces apoptosis of human acute monocytic leukemia cells via downregulation of the ERK/Akt signaling pathway. Exp. Ther. Med. 2017, 14, 3067–3073. [Google Scholar] [CrossRef][Green Version]
- Wang, Z.; Wu, X.; Liang, Y.-N.; Wang, L.; Song, Z.-X.; Liu, J.-L.; Tang, Z.-S. Cordycepin induces apoptosis and inhibits proliferation of human lung cancer cell line H1975 via inhibiting the phosphorylation of EGFR. Molecules 2016, 21, 1267. [Google Scholar] [CrossRef]
- Wong, Y.Y.; Moon, A.; Duffin, R.; Barthet-Barateig, A.; Meijer, H.A.; Clemens, M.J.; de Moor, C.H. Cordycepin inhibits protein synthesis and cell adhesion through effects on signal transduction. J. Biol. Chem. 2010, 285, 2610–2621. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhao, L.; Yuan, W.; Wen, J. Cordycepin induces apoptotic cell death and inhibits cell migration in renal cell carcinoma via regulation of microRNA-21 and PTEN phosphatase. Biomed. Res. 2017, 38, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-S.; Lee, K.W.; Madhi, H.; Jeong, J.-W.; Park, S.; Kim, M.; Lee, Y.; Han, H.-T.; Hwangbo, C.; Yoo, J. Cordycepin Resensitizes T24R2 Cisplatin-Resistant Human Bladder Cancer Cells to Cisplatin by Inactivating Ets-1 Dependent MDR1 Transcription. Int. J. Mol. Sci. 2020, 21, 1710. [Google Scholar] [CrossRef]
- Cao, H.-L.; Liu, Z.-J.; Chang, Z. Cordycepin induces apoptosis in human bladder cancer cells via activation of A3 adenosine receptors. Tumor Biol. 2017, 39, 1010428317706915. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.-M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef]
- Wong, K.A.; Lodish, H.F. A revised model for AMP-activated protein kinase structure: The alpha-subunit binds to both the beta- and gamma-subunits although there is no direct binding between the beta- and gamma-subunits. J Biol Chem 2006, 281, 36434–36442. [Google Scholar] [CrossRef]
- Hardie, D.G. AMP-activated protein kinase—An energy sensor that regulates all aspects of cell function. Genes Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, B.; Chhipa, R.R. Evolving lessons on the complex role of AMPK in normal physiology and cancer. Trends Pharmacol. Sci. 2016, 37, 192–206. [Google Scholar] [CrossRef]
- Gauthier, M.-S.; O’Brien, E.L.; Bigornia, S.; Mott, M.; Cacicedo, J.M.; Xu, X.J.; Gokce, N.; Apovian, C.; Ruderman, N. Decreased AMP-activated protein kinase activity is associated with increased inflammation in visceral adipose tissue and with whole-body insulin resistance in morbidly obese humans. Biochem. Biophys. Res. Commun. 2011, 404, 382–387. [Google Scholar] [CrossRef]
- Valentine, R.J.; Coughlan, K.A.; Ruderman, N.B.; Saha, A.K. Insulin inhibits AMPK activity and phosphorylates AMPK Ser485/491 through Akt in hepatocytes, myotubes and incubated rat skeletal muscle. Arch. Biochem. Biophys. 2014, 562, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Willows, R.; Sanders, M.J.; Xiao, B.; Patel, B.R.; Martin, S.R.; Read, J.; Wilson, J.R.; Hubbard, J.; Gamblin, S.J.; Carling, D. Phosphorylation of AMPK by upstream kinases is required for activity in mammalian cells. Biochem J 2017, 474, 3059–3073. [Google Scholar] [CrossRef] [PubMed]
- Dite, T.A.; Ling, N.X.; Scott, J.W.; Hoque, A.; Galic, S.; Parker, B.L.; Ngoei, K.R.; Langendorf, C.G.; O’Brien, M.T.; Kundu, M. The autophagy initiator ULK1 sensitizes AMPK to allosteric drugs. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Kai, Q.; Gao, J.; Lian, Z.Q.; Wu, C.M.; Wu, C.A.; Zhu, H.B. Cordycepin prevents hyperlipidemia in hamsters fed a high-fat diet via activation of AMP-activated protein kinase. J Pharm. Sci 2010, 113, 395–403. [Google Scholar] [CrossRef]
- Hawley, S.A.; Ross, F.A.; Russell, F.M.; Atrih, A.; Lamont, D.J.; Hardie, D.G. Mechanism of Activation of AMPK by Cordycepin. Cell Chem. Biol. 2020, 27, 214–222.e214. [Google Scholar] [CrossRef]
- Qi, G.; Zhou, Y.; Zhang, X.; Yu, J.; Li, X.; Cao, X.; Wu, C.; Guo, P. Cordycepin promotes browning of white adipose tissue through an AMP-activated protein kinase (AMPK)-dependent pathway. Acta Pharm. Sin. B 2019, 9, 135–143. [Google Scholar] [CrossRef]
- Wu, C.; Guo, Y.; Su, Y.; Zhang, X.; Luan, H.; Zhang, X.; Zhu, H.; He, H.; Wang, X.; Sun, G. Cordycepin activates AMP-activated protein kinase (AMPK) via interaction with the γ1 subunit. J. Cell. Mol. Med. 2014, 18, 293–304. [Google Scholar] [CrossRef]
- Lee, C.; Kim, Y.; Jeon, J.H. JNK and p38 mitogen-activated protein kinase pathways contribute to porcine epidemic diarrhea virus infection. Virus Res. 2016, 222, 1–12. [Google Scholar] [CrossRef]
- Ko, H.M.; Joo, S.H.; Kim, P.; Park, J.H.; Kim, H.J.; Bahn, G.H.; Kim, H.Y.; Lee, J.; Han, S.-H.; Shin, C.Y. Effects of Korean Red Ginseng extract on tissue plasminogen activator and plasminogen activator inhibitor-1 expression in cultured rat primary astrocytes. J. Ginseng Res. 2013, 37, 401. [Google Scholar] [CrossRef]
- Koul, H.K.; Pal, M.; Koul, S. Role of p38 MAP kinase signal transduction in solid tumors. Genes Cancer 2013, 4, 342–359. [Google Scholar] [CrossRef]
- Lim, A.K.; Tesch, G.H. Inflammation in diabetic nephropathy. Mediat. Inflamm. 2012, 2012. [Google Scholar] [CrossRef]
- Ren, F.; Zhang, H.; Piao, Z.; Zheng, S.; Chen, Y.; Chen, D.; Duan, Z. Inhibition of glycogen synthase kinase 3b activity regulates Toll-like receptor 4-mediated liver inflammation. Zhonghua Gan Zang Bing Za Zhi = Zhonghua Ganzangbing Zazhi = Chin. J. Hepatol. 2012, 20, 693–697. [Google Scholar]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Et Biophys. Acta (Bba)-Mol. Cell Res. 2007, 1773, 1358–1375. [Google Scholar] [CrossRef]
- Imesch, P.; Hornung, R.; Fink, D.; Fedier, A. Cordycepin (3′-deoxyadenosine), an inhibitor of mRNA polyadenylation, suppresses proliferation and activates apoptosis in human epithelial endometriotic cells in vitro. Gynecol. Obstet. Investig. 2011, 72, 43–49. [Google Scholar] [CrossRef]
- Du, Y.; Yu, J.; Du, L.; Tang, J.; Feng, W.-H. Cordycepin enhances Epstein–Barr virus lytic infection and Epstein–Barr virus-positive tumor treatment efficacy by doxorubicin. Cancer Lett. 2016, 376, 240–248. [Google Scholar] [CrossRef]
- Lee, S.-J.; Kim, S.-K.; Choi, W.-S.; Kim, W.-J.; Moon, S.-K. Cordycepin causes p21WAF1-mediated G2/M cell-cycle arrest by regulating c-Jun N-terminal kinase activation in human bladder cancer cells. Arch. Biochem. Biophys. 2009, 490, 103–109. [Google Scholar] [CrossRef]
- Pao, H.-Y.; Pan, B.-S.; Leu, S.-F.; Huang, B.-M. Cordycepin stimulated steroidogenesis in MA-10 mouse Leydig tumor cells through the protein kinase C Pathway. J. Agric. Food Chem. 2012, 60, 4905–4913. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Ling, J.; Zhang, G.; Liu, F.; Tao, S.; Han, Z.; Chen, S.; Chen, Z.; Le, H. Cordycepin induces cell cycle arrest and apoptosis by inducing DNA damage and up-regulation of p53 in Leukemia cells. Cell Cycle 2015, 14, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Yarza, R.; Vela, S.; Solas, M.; Ramirez, M.J. c-Jun N-terminal kinase (JNK) signaling as a therapeutic target for Alzheimer’s disease. Front. Pharmacol. 2016, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Zeke, A.; Misheva, M.; Reményi, A.; Bogoyevitch, M.A. JNK signaling: Regulation and functions based on complex protein-protein partnerships. Microbiol. Mol. Biol. Rev. 2016, 80, 793–835. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Kraus, I.; Besong Agbo, D.; Otto, M.; Wiltfang, J.; Klafki, H. Detection and Differentiation of Threonine- and Tyrosine-Monophosphorylated Forms of ERK1/2 by Capillary Isoelectric Focusing-Immunoassay. Sci. Rep. 2015, 5, 12767. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-S.; Kang, D.-H.; Yang, M.-K.; Kang, J.C.; Jang, Y.C.; Park, J.S.; Kim, S.-K.; Shin, H.-S. Cordycepin, 3′-deoxyadenosine, prevents rat hearts from ischemia/reperfusion injury via activation of Akt/GSK-3β/p70S6K signaling pathway and HO-1 expression. Cardiovasc. Toxicol. 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Duan, M.X.; Xu, M.; Huang, S.H.; Yang, J.; Yang, J.; Liu, L.B.; Huang, R.; Wan, C.X.; Ma, Z.G.; et al. Cordycepin ameliorates cardiac hypertrophy via activating the AMPKalpha pathway. J. Cell Mol. Med. 2019, 23, 5715–5727. [Google Scholar] [CrossRef]
- Jagger, D.V.; Kredich, N.M.; Guarino, A.J. Inhibition of Ehrlich mouse ascites tumor growth by cordycepin. Cancer Res. 1961, 21, 216–220. [Google Scholar]
- Bi, Y.; Li, H.; Yi, D.; Bai, Y.; Zhong, S.; Liu, Q.; Chen, Y.; Zhao, G. β-catenin contributes to cordycepin-induced MGMT inhibition and reduction of temozolomide resistance in glioma cells by increasing intracellular reactive oxygen species. Cancer Lett. 2018, 435, 66–79. [Google Scholar] [CrossRef]
- Dong, J.; Li, Y.; Xiao, H.; Luo, D.; Zhang, S.; Zhu, C.; Jiang, M.; Cui, M.; Lu, L.; Fan, S. Cordycepin sensitizes breast cancer cells toward irradiation through elevating ROS production involving Nrf2. Toxicol. Appl. Pharmacol. 2019, 364, 12–21. [Google Scholar] [CrossRef]
- Horsman, M.R.; Brown, D.M.; Hirst, D.G.; Brown, J.M. The effects of purine nucleoside analogs on the response of the RIF-1 tumor to melphalan in vivo. Int. J. Radiat. Oncol. Biol. Phys. 1986, 12, 801–806. [Google Scholar] [CrossRef]
- Wang, C.; Mao, Z.; Wang, L.; Zhang, F.; Wu, G.; Wang, D.; Shi, J. Cordycepin inhibits cell growth and induces apoptosis in human cholangiocarcinoma. Neoplasma 2017, 64, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Zhang, Q.X.; Leung, P.H. Inhibitory effects of ethyl acetate extract of Cordyceps sinensis mycelium on various cancer cells in culture and B16 melanoma in C57BL/6 mice. Phytomedicine 2007, 14, 43–49. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Nakamura, K.; Yamaguchi, Y.; Kagota, S.; Shinozuka, K.; Kunitomo, M. Antitumour activity of cordycepin in mice. Clin. Exp. Pharmacol. Physiol. 2004, 31, S51–S53. [Google Scholar] [CrossRef]
- Zhou, Y.; Guo, Z.; Meng, Q.; Lu, J.; Wang, N.; Liu, H.; Liang, Q.; Quan, Y.; Wang, D.; Xie, J. Cordycepin affects multiple apoptotic pathways to mediate hepatocellular carcinoma cell death. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. -Anti-Cancer Agents) 2017, 17, 143–149. [Google Scholar] [CrossRef]
- Aramwit, P.; Porasuphatana, S.; Srichana, T.; Nakpheng, T. Toxicity evaluation of cordycepin and its delivery system for sustained in vitro anti-lung cancer activity. Nanoscale Res. Lett. 2015, 10, 152. [Google Scholar] [CrossRef]
- Foss, F.M. Combination therapy with purine nucleoside analogs. Oncol. (Williston ParkNy) 2000, 14, 31–35. [Google Scholar]
- Johns, D.G.; Adamson, R.H. Enhancement of the biological activity of cordycepin (3’-deoxyadenosine) by the adenosine deaminase inhibitor 2’-deoxycoformycin. Biochem. Pharmacol. 1976, 25, 1441–1444. [Google Scholar] [CrossRef]
- Joo, J.C.; Hwang, J.H.; Jo, E.; Kim, Y.-R.; Kim, D.J.; Lee, K.-B.; Park, S.J.; Jang, I.-S. Cordycepin induces apoptosis by caveolin-1-mediated JNK regulation of Foxo3a in human lung adenocarcinoma. Oncotarget 2017, 8, 12211. [Google Scholar] [CrossRef]
- Sato, A.; Yoshikawa, N.; Kubo, E.; Kakuda, M.; Nishiuchi, A.; Kimoto, Y.; Takahashi, Y.; Kagota, S.; Shinozuka, K.; Nakamura, K. Inhibitory effect of cordycepin on experimental hepatic metastasis of B16-F0 mouse melanoma cells. Vivo 2013, 27, 729–732. [Google Scholar]
- Wu, P.-K.; Tao, Z.; Ouyang, Z.; Cao, J.-Y.; Geng, D.; Liu, J.; Wang, C.-M. The anti-tumor effects of cordycepin-loaded liposomes on the growth of hepatoma 22 tumors in mice and human hepatoma BEL-7402 cells in culture. Drug Dev. Ind. Pharm. 2016, 42, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, N.; Kunitomo, M.; Kagota, S.; Shinozuka, K.; Nakamura, K. Inhibitory effect of cordycepin on hematogenic metastasis of B16-F1 mouse melanoma cells accelerated by adenosine-5′-diphosphate. Anticancer Res. 2009, 29, 3857–3860. [Google Scholar] [PubMed]
- Liu, T.; Zhu, G.; Yan, W.; Lv, Y.; Wang, X.; Jin, G.; Cui, M.; Lin, Z.; Ren, X. Cordycepin inhibits cancer cell proliferation and angiogenesis through a DEK interaction via ERK signaling in cholangiocarcinoma. J. Pharmacol. Exp. Ther. 2020, 373, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Xue-Ying, L.; Homng, T.; Can, J.; Zhen-Yun, D.; Wen-Feng, L.; Qing-Jiu, T.; Kan, D. Cordycepin inhibits pancreatic cancer cell growth in vitro and in vivo via targeting FGFR2 and blocking ERK signaling. Chin. J. Nat. Med. 2020, 18, 345–355. [Google Scholar]
- Zhang, Y.; Zhang, X.X.; Yuan, R.Y.; Ren, T.; Shao, Z.Y.; Wang, H.F.; Cai, W.L.; Chen, L.T.; Wang, X.A.; Wang, P. Cordycepin induces apoptosis in human pancreatic cancer cells via the mitochondrial-mediated intrinsic pathway and suppresses tumor growth in vivo. Oncotargets Ther. 2018, 11, 4479. [Google Scholar] [CrossRef]
- Zheng, Q.; Sun, J.; Li, W.; Li, S.; Zhang, K. Cordycepin induces apoptosis in human tongue cancer cells in vitro and has antitumor effects in vivo. Arch. Oral Biol. 2020, 104846. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.-P.; Shang, Y.-J.; Liu, Q.-Y.; Wu, B.-Y.; Liu, W.-X.; Li, C.-H. Neuroprotection of cordycepin in NMDA-induced excitotoxicity by modulating adenosine A1 receptors. Eur. J. Pharmacol. 2019, 853, 325–335. [Google Scholar]
- Cheng, Z.; He, W.; Zhou, X.; Lv, Q.; Xu, X.; Yang, S.; Zhao, C.; Guo, L. Cordycepin protects against cerebral ischemia/reperfusion injury in vivo and in vitro. Eur. J. Pharmacol. 2011, 664, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.K.; Lim, S.S.; Yoo, K.-Y.; Lee, Y.S.; Kim, H.G.; Kang, I.-J.; Kwon, H.J.; Park, J.; Choi, S.Y.; Won, M.-H. A phytochemically characterized extract of Cordyceps militaris and cordycepin protect hippocampal neurons from ischemic injury in gerbils. Planta Med. 2008, 74, 114–119. [Google Scholar] [CrossRef]
- Gao, J.; Lian, Z.; Zhu, P.; Zhu, H. Lipid-lowering effect of cordycepin (3’-deoxyadenosine) from Cordyceps militaris on hyperlipidemic hamsters and rats. Yao Xue Xue Bao= Acta Pharm. Sin. 2011, 46, 669. [Google Scholar]
- Sun, Y.; Wang, Y.-H.; Qu, K.; Zhu, H.-B. Beneficial effects of cordycepin on metabolic profiles of liver and plasma from hyperlipidemic hamsters. J. Asian Nat. Prod. Res. 2011, 13, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wei, Y.; Yang, W.; Song, Y.; Shang, H.; Cai, Y.; Wu, Z.; Zhao, W. Cordycepin confers neuroprotection in mice models of intracerebral hemorrhage via suppressing NLRP3 inflammasome activation. Metab. Brain Dis. 2017, 32, 1133–1145. [Google Scholar] [CrossRef]
- Won, K.-J.; Lee, S.-C.; Lee, C.-K.; Lee, H.M.; Lee, S.H.; Fang, Z.; Choi, O.B.; Jin, M.; Kim, J.; Park, T. Cordycepin attenuates neointimal formation by inhibiting reactive oxygen species–mediated responses in vascular smooth muscle cells in rats. J. Pharmacol. Sci. 2009, 109, 403–412. [Google Scholar] [CrossRef]
- Xingqiang, L.; Fen, N.; Zhongpeng, Y.; Tiantian, W.; Lei, Z.; Jiali, F.; Junjie, M.; Guanghui, L.; Lu, X.; Yuhe, G. Ethylene carbodiimide-fixed donor splenocytes combined with cordycepin induce long-term protection to mice cardiac allografts. Transpl. Immunol. 2019, 56, 101196. [Google Scholar] [CrossRef]
- Araldi, D.; Ferrari, L.F.; Levine, J.D. Gi-protein coupled 5-HT1B/D receptor agonist sumatriptan induces type I hyperalgesic priming. Pain 2016, 157, 1773. [Google Scholar] [CrossRef] [PubMed]
- Araldi, D.; Ferrari, L.F.; Levine, J.D. Mu-opioid receptor (MOR) biased agonists induce biphasic dose-dependent hyperalgesia and analgesia, and hyperalgesic priming in the rat. Neuroscience 2018, 394, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, L.F.; Araldi, D.; Bogen, O.; Green, P.G.; Levine, J.D. Systemic morphine produces dose-dependent nociceptor-mediated biphasic changes in nociceptive threshold and neuroplasticity. Neuroscience 2019, 398, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, L.F.; Araldi, D.; Levine, J.D. Distinct terminal and cell body mechanisms in the nociceptor mediate hyperalgesic priming. J. Neurosci. 2015, 35, 6107–6116. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, L.F.; Bogen, O.; Chu, C.; Levine, J.D. Peripheral administration of translation inhibitors reverses increased hyperalgesia in a model of chronic pain in the rat. J. Pain 2013, 14, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Khomula, E.V.; Araldi, D.; Levine, J.D. In vitro nociceptor neuroplasticity associated with in vivo opioid-induced hyperalgesia. J. Neurosci. 2019, 39, 7061–7073. [Google Scholar] [CrossRef]
- Song, R.; Zheng, J.; Liu, Y.; Tan, Y.; Yang, Z.; Song, X.; Yang, S.; Fan, R.; Zhang, Y.; Wang, Y. A natural cordycepin/chitosan complex hydrogel with outstanding self-healable and wound healing properties. Int. J. Biol. Macromol. 2019, 134, 91–99. [Google Scholar] [CrossRef]
- Yang, F.; Sun, W.; Luo, W.-J.; Yang, Y.; Yang, F.; Wang, X.-L.; Chen, J. SDF1-CXCR4 signaling contributes to the transition from acute to chronic pain state. Mol. Neurobiol. 2017, 54, 2763–2775. [Google Scholar] [CrossRef]
- Aiyedun, B.; Williamson, J.; Amodu, A. The effect of cordy-cepin on tsetse-borne Trypanosoma vivax infections. Acta Trop. 1973, 30, 276–278. [Google Scholar]
- Da Silva, A.S.; Wolkmer, P.; Nunes, J.T.; Duck, M.R.; Oliveira, C.B.; Gressler, L.T.; Costa, M.M.; Zanette, R.A.; Mazzanti, C.M.; Lopes, S.T. Susceptibility of Trypanosoma evansi to cordycepin. Biomed. Pharmacother. 2011, 65, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Dalla Rosa, L.; Da Silva, A.S.; Oliveira, C.B.; Gressler, L.T.; Arnold, C.B.; Baldissera, M.D.; Sagrillo, M.; Sangoi, M.; Moresco, R.; Mendes, R.E. Dose finding of 3′ deoxyadenosine and deoxycoformycin for the treatment of Trypanosoma evansi infection: An effective and nontoxic dose. Microb. Pathog. 2015, 85, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Dalla Rosa, L.; Da Silva, A.S.; Ruchel, J.B.; Gressler, L.T.; Oliveira, C.B.; França, R.T.; Lopes, S.T.; Leal, D.B.; Monteiro, S.G. Influence of treatment with 3′-deoxyadenosine associated deoxycoformycin on hematological parameters and activity of adenosine deaminase in infected mice with Trypanosoma evansi. Exp. Parasitol. 2013, 135, 357–362. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dalla Rosa, L.; Da Silva, A.S.; Gressler, L.T.; Oliveira, C.B.; Dambros, M.G.; Miletti, L.C.; T. FRANÇA, R.; Lopes, S.T.; Samara, Y.N.; Da Veiga, M.L. Cordycepin (3’-deoxyadenosine) pentostatin (deoxycoformycin) combination treatment of mice experimentally infected with Trypanosoma evansi. Parasitology 2013, 140, 663. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, G.M.; de Sá, M.F.; Grando, T.H.; Gressler, L.T.; Baldissera, M.D.; Monteiro, S.G.; Henker, L.C.; Mendes, R.E.; Stefani, L.M.; Da Silva, A.S. Cordycepin (3′-deoxyadenosine) and pentostatin (deoxycoformycin) against Trypanosoma cruzi. Exp. Parasitol. 2019, 199, 47–51. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, G.M.; Doleski, P.H.; de Sá, M.F.; Grando, T.H.; Azevedo, M.I.; Manzoni, A.G.; Leal, D.B.; Gressler, L.T.; Henker, L.C.; Mendes, R.E. Treatment with 3′-deoxyadenosine and deoxycoformycin in mice infected by Trypanosoma cruzi and its side effect on purinergic enzymes. Microb. Pathog. 2017, 113, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Vodnala, S.K.; Ferella, M.; Lundén-Miguel, H.; Betha, E.; Van Reet, N.; Amin, D.N.; Öberg, B.; Andersson, B.; Kristensson, K.; Wigzell, H. Preclinical assessment of the treatment of second-stage African trypanosomiasis with cordycepin and deoxycoformycin. PLoS Negl. Trop. Dis. 2009, 3, e495. [Google Scholar] [CrossRef]
- Vodnala, S.K.; Lundbäck, T.; Yeheskieli, E.; Sjöberg, B.; Gustavsson, A.-L.; Svensson, R.; Olivera, G.C.; Eze, A.A.; de Koning, H.P.; Hammarström, L.G. Structure–activity relationships of synthetic cordycepin analogues as experimental therapeutics for African trypanosomiasis. J. Med. Chem. 2013, 56, 9861–9873. [Google Scholar] [CrossRef]
- Williamson, J. Further experiments with the nucleoside trypanocide, cordycepin. Trans. R. Soc. Trop. Med. Hyg. 1972, 66, 354. [Google Scholar] [CrossRef]
- Williamson, J.; Macadam, R. Drug effects on the fine structure of Trypanosoma rhodesiense: Puromycin and its aminonucleoside, Cordycepin and Nucleocidin. Trans. R. Soc. Trop. Med. Hyg. 1976, 70, 130–137. [Google Scholar] [CrossRef]
- Williamson, J.; Scott-Finnigan, T. Trypanocidal activity of antitumor antibiotics and other metabolic inhibitors. Antimicrob. Agents Chemother. 1978, 13, 735–744. [Google Scholar] [CrossRef]
- Li, B.; Hou, Y.; Zhu, M.; Bao, H.; Nie, J.; Zhang, G.Y.; Shan, L.; Yao, Y.; Du, K.; Yang, H. 3’-deoxyadenosine (cordycepin) produces a rapid and robust antidepressant effect via enhancing prefrontal AMPA receptor signaling pathway. Int. J. Neuropsychopharmacol. 2016, 19, pyv112. [Google Scholar] [CrossRef]
- Cai, Z.-L.; Wang, C.-Y.; Jiang, Z.-J.; Li, H.-H.; Liu, W.-X.; Gong, L.-W.; Xiao, P.; Li, C.-H. Effects of cordycepin on Y-maze learning task in mice. Eur. J. Pharmacol. 2013, 714, 249–253. [Google Scholar] [CrossRef]
- Cao, Z.-P.; Dai, D.; Wei, P.-J.; Han, Y.-Y.; Guan, Y.-Q.; Li, H.-H.; Liu, W.-X.; Xiao, P.; Li, C.-H. Effects of cordycepin on spontaneous alternation behavior and adenosine receptors expression in hippocampus. Physiol. Behav. 2018, 184, 135–142. [Google Scholar] [CrossRef]
- Han, Y.Y.; Chen, Z.H.; Shang, Y.J.; Yan, W.W.; Wu, B.Y.; Li, C.H. Cordycepin improves behavioral-LTP and dendritic structure in hippocampal CA1 area of rats. J. Neurochem. 2019, 151, 79–90. [Google Scholar] [CrossRef]
- Zearfoss, N.R.; Alarcon, J.M.; Trifilieff, P.; Kandel, E.; Richter, J.D. A molecular circuit composed of CPEB-1 and c-Jun controls growth hormone-mediated synaptic plasticity in the mouse hippocampus. J. Neurosci. 2008, 28, 8502–8509. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Lee, C.-I.; Shah, V.K.; Oh, E.-H.; Han, J.-Y.; Bae, J.-R.; Lee, K.; Chong, M.-S.; Hong, J.T.; Oh, K.-W. Cordycepin increases nonrapid eye movement sleep via adenosine receptors in rats. Evid. -Based Complementary Altern. Med. 2013, 2013, 840134. [Google Scholar] [CrossRef]
- An, Y.; Li, Y.; Wang, X.; Chen, Z.; Xu, H.; Wu, L.; Li, S.; Wang, C.; Luan, W.; Wang, X. Cordycepin reduces weight through regulating gut microbiota in high-fat diet-induced obese rats. Lipids Health Dis. 2018, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Wang, X.; Xu, H.; Wang, C.; An, Y.; Luan, W.; Wang, X.; Li, S.; Ma, F. Cordycepin modulates body weight by reducing prolactin via an adenosine A1 receptor. Curr. Pharm. Des. 2018, 24, 3240–3249. [Google Scholar] [CrossRef]
- Xu, H.; Wu, B.; Wang, X.; Ma, F.; Li, Y.; An, Y.; Wang, C.; Wang, X.; Luan, W.; Li, S. Cordycepin regulates body weight by inhibiting lipid droplet formation, promoting lipolysis and recruiting beige adipocytes. J. Pharm. Pharmacol. 2019, 71, 1429–1439. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Xu, R.; Xu, Y.; Liu, Y.; Qi, D.; Wan, Q. The protective effect of Cordycepin on diabetic nephropathy through autophagy induction in vivo and in vitro. Int. Urol. Nephrol. 2019, 51, 1883–1892. [Google Scholar] [CrossRef]
- Cimbala, M.; Lamers, W.; Nelson, K.; Monahan, J.; Yoo-Warren, H.; Hanson, R. Rapid changes in the concentration of phosphoenolpyruvate carboxykinase mRNA in rat liver and kidney. Effects of insulin and cyclic AMP. J. Biol. Chem. 1982, 257, 7629–7636. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, S.; Du, M. Cordycepin from Cordyceps militaris prevents hyperglycemia in alloxan-induced diabetic mice. Nutr. Res. 2015, 35, 431–439. [Google Scholar] [CrossRef]
- Borgan, J.-L.; Bonnafous, J.-C.; Mousseron-Canet, M.; Mani, J.-C.; Cazaubon, C. Cordycepin and early effects of estradiol on the immature rat uterus. Biochimie 1976, 58, 443–454. [Google Scholar] [CrossRef]
- Fernandez-Noval, A.; Leroy, F. 3′-DEOXYADENOSINE AND IMPLANTATION OF DELAYED BLASTOCYSTS IN MICE. J. Endocrinol. 1979, 81, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Leu, S.-F.; Poon, S.L.; Pao, H.-Y.; Huang, B.-M. The in vivo and in vitro stimulatory effects of cordycepin on mouse leydig cell steroidogenesis. Biosci. Biotechnol. Biochem. 2011, 75, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-H.; Tsai, M.-T.; Chen, Y.-S.; Hou, R.C.-W.; Hung, H.-F.; Li, C.-H.; Wang, H.-K.; Lai, M.-N.; Jeng, K.-C.G. Improvement of sperm production in subfertile boars by Cordyceps militaris supplement. Am. J. Chin. Med. 2007, 35, 631–641. [Google Scholar] [CrossRef]
- Sohn, S.-H.; Lee, S.-C.; Hwang, S.-Y.; Kim, S.-W.; Kim, I.-W.; Michael, B.Y.; Kim, S.-K. Effect of long-term administration of cordycepin from Cordyceps militaris on testicular function in middle-aged rats. Planta Med. 2012, 78, 1620–1625. [Google Scholar] [CrossRef]
- Ulibarri, C.; Yahr, P. Poly-A+ mRNA and defeminization of sexual behavior and gonadotropin secretion in rats. Physiol. Behav. 1987, 39, 767–774. [Google Scholar] [CrossRef]
- Yahr, P.; Ulibarri, C. Estrogen induction of sexual behavior in female rats and synthesis of polyadenylated messenger RNA in the ventromedial nucleus of the hypothalamus. Mol. Brain Res. 1986, 1, 153–165. [Google Scholar] [CrossRef]
- Yahr, P.; Ulibarri, C. Polyadenylated and nonadenylated messenger RNA and androgen control of sexual behavior and scent marking in male gerbils. Horm. Behav. 1987, 21, 53–64. [Google Scholar] [CrossRef]
- Dickson, B.M.; Roelofs, A.J.; Rochford, J.J.; Wilson, H.M.; De Bari, C. The burden of metabolic syndrome on osteoarthritic joints. Arthritis Res. Ther. 2019, 21, 1–10. [Google Scholar] [CrossRef]
- Kang, S.; Kumanogoh, A. The spectrum of macrophage activation by immunometabolism. Int. Immunol. 2020, 32, 467–473. [Google Scholar] [CrossRef]
- Noe, J.T.; Mitchell, R.A. Tricarboxylic acid cycle metabolites in the control of macrophage activation and effector phenotypes. J. Leukoc. Biol. 2019, 106, 359–367. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Pearce, E.J. Immunometabolism governs dendritic cell and macrophage function. J. Exp. Med. 2016, 213, 15–23. [Google Scholar] [CrossRef]
- Marslin, G.; Khandelwal, V.; Franklin, G. Cordycepin nanoencapsulated in poly (Lactic-Co-Glycolic acid) exhibits better cytotoxicity and lower hemotoxicity than free drug. Nanotechnol. Sci. Appl. 2020, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Adrower, C.; Qin, C.; Fischer, P.M.; de Moor, C.H.; Gershkovich, P. Development of cordycepin formulations for preclinical and clinical studies. Aaps Pharmscitech 2017, 18, 3219–3226. [Google Scholar] [CrossRef]
- Zhang, J.-Q.; Wu, D.; Jiang, K.-M.; Zhang, D.; Zheng, X.; Wan, C.-P.; Zhu, H.-Y.; Xie, X.-G.; Jin, Y.; Lin, J. Preparation, spectroscopy and molecular modelling studies of the inclusion complex of cordycepin with cyclodextrins. Carbohydr. Res. 2015, 406, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Zhou, Y.; Wang, M.; Li, L.; Lee, R.J.; Xie, J.; Teng, L. Targeted delivery of cordycepin to liver cancer cells using transferrin-conjugated liposomes. Anticancer Res. 2017, 37, 5207–5214. [Google Scholar] [PubMed]
- Cheek, M.A.; Dobrikov, M.I.; Wennefors, C.K.; Xu, Z.; Hashmi, S.N.; Shen, X.; Shaw, B.R. Synthesis and properties of (α-P-borano)-nucleoside 5′-triphosphate analogues as potential antiviral agents. Nucleic Acids Symp. Ser. 2008, 52, 81–82. [Google Scholar]
- Hulpia, F.; Mabille, D.; Campagnaro, G.D.; Schumann, G.; Maes, L.; Roditi, I.; Hofer, A.; De Koning, H.P.; Caljon, G.; Van Calenbergh, S. Combining tubercidin and cordycepin scaffolds results in highly active candidates to treat late-stage sleeping sickness. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.-X.; Yang, L.; Jiang, C.-Y.; Wang, Y.-H.; Zhu, H.-B. Evaluation of dose-related effects of 2’, 3’, 5’-tri-O-acetyl-N6-(3-hydroxylaniline) adenosine using NMR-based metabolomics. Yao Xue Xue Bao= Acta Pharm. Sin. 2014, 49, 679–685. [Google Scholar]
- Kubota, Y.; Ehara, M.; Haraguchi, K.; Tanaka, H. Phenylsulfanylation of 3′, 4′-Unsaturated Adenosine Employing Thiophenol-N-Iodosuccinimide Leads to 4′-Phenylsulfanylcordycepin: Synthesis of 4′-Substituted Cordycepins on the Basis of Substitution of the Phenylsulfanyl Leaving Group. J. Org. Chem. 2011, 76, 8710–8717. [Google Scholar] [CrossRef]
- Fong, P.; Ao, C.N.; Tou, K.I.; Huang, K.M.; Cheong, C.C.; Meng, L.R. Experimental and in silico analysis of cordycepin and its derivatives as endometrial cancer treatment. Oncol. Res. 2019, 27, 237. [Google Scholar] [CrossRef]
- Chaicharoenaudomrung, N.; Kunhorm, P.; Promjantuek, W.; Heebkaew, N.; Rujanapun, N.; Noisa, P. Fabrication of 3D calcium-alginate scaffolds for human glioblastoma modeling and anticancer drug response evaluation. J. Cell. Physiol. 2019, 234, 20085–20097. [Google Scholar] [CrossRef]
- El Khadem, H.S.; El Sayed, H. Synthesis of a c-nucleoside analog of the antibiotic cordycepin. Carbohydr. Res. 1974, 32, 339–348. [Google Scholar] [CrossRef]
- Kaokaen, P.; Jaiboonma, A.; Chaicharoenaudomrung, N.; Kunhorm, P.; Janebodin, K.; Noisa, P.; Jitprasertwong, P. Cordycepin-loaded Nanoparticles from Cassava Starch Promote the Proliferation of Submandibular Gland Cells and Inhibit the Growth of Oral Squamous Carcinoma Cells. Nutr. Cancer 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Holbein, S.; Wengi, A.; Decourty, L.; Freimoser, F.M.; Jacquier, A.; Dichtl, B. Cordycepin interferes with 3′ end formation in yeast independently of its potential to terminate RNA chain elongation. RNA 2009, 15, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Qu, K.; Zhu, P.; Guo, N.; Zhang, R.; Abliz, Z.; Yu, H.; Zhu, H. Binding of Cordycepin Monophosphate to AMP-Activated Protein Kinase and its Effect on AMP-Activated Protein Kinase Activation. Chem. Biol. Drug Des. 2010, 76, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.Y.; Kim, J.H.; Baek, K.-S.; Kim, G.S.; Lee, S.E.; Lee, D.Y.; Choi, J.H.; Kim, S.Y.; Park, H.B.; Sung, G.-H. A direct protein kinase B-targeted anti-inflammatory activity of cordycepin from artificially cultured fruit body of Cordyceps militaris. Pharmacogn. Mag. 2015, 11, 477. [Google Scholar] [PubMed]
- Bard, J.; Zhelkovsky, A.M.; Helmling, S.; Earnest, T.N.; Moore, C.L.; Bohm, A. Structure of yeast poly (A) polymerase alone and in complex with 3’-dATP. Science 2000, 289, 1346–1349. [Google Scholar] [CrossRef] [PubMed]
- Ryner, L.C.; Manley, J.L. Requirements for accurate and efficient mRNA 3’end cleavage and polyadenylation of a simian virus 40 early pre-RNA in vitro. Mol. Cell. Biol. 1987, 7, 495–503. [Google Scholar] [CrossRef]
- Ju, D.; Zhang, W.; Yan, J.; Zhao, H.; Li, W.; Wang, J.; Liao, M.; Xu, Z.; Wang, Z.; Zhou, G. Chemical perturbations reveal that RUVBL2 regulates the circadian phase in mammals. Sci. Transl. Med. 2020, 12, eaba0769. [Google Scholar] [CrossRef] [PubMed]
- Kadomatsu, M.; Nakajima, S.; Kato, H.; Gu, L.; Chi, Y.; Yao, J.; Kitamura, M. Cordycepin as a sensitizer to tumour necrosis factor (TNF)-α-induced apoptosis through eukaryotic translation initiation factor 2α (eIF2α)-and mammalian target of rapamycin complex 1 (mTORC1)-mediated inhibition of nuclear factor (NF)-κB. Clin. Exp. Immunol. 2012, 168, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, N.; Yamada, S.; Takeuchi, C.; Kagota, S.; Shinozuka, K.; Kunitomo, M.; Nakamura, K. Cordycepin (3′-deoxyadenosine) inhibits the growth of B16-BL6 mouse melanoma cells through the stimulation of adenosine A 3 receptor followed by glycogen synthase kinase-3β activation and cyclin D 1 suppression. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 377, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, S.-H.; Hueng, D.-Y.; Syu, J.-P.; Liao, C.-C.; Wu, Y.-C. Cordycepin induces apoptosis of C6 glioma cells through the adenosine 2A receptor-p53-caspase-7-PARP pathway. Chem. -Biol. Interact. 2014, 216, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.-S.; Lin, C.-Y.; Huang, B.-M. The effect of cordycepin on steroidogenesis and apoptosis in MA-10 mouse Leydig tumor cells. Evid. -Based Complementary Altern. Med. 2011, 2011, 750468. [Google Scholar] [CrossRef]
- Zarkower, D.; Wickens, M. Formation of mRNA 3′ termini: Stability and dissociation of a complex involving the AAUAAA sequence. Embo J. 1987, 6, 177–186. [Google Scholar] [CrossRef]
- Kim, S.O.; Cha, H.-J.; Park, C.; Lee, H.; Hong, S.H.; Jeong, S.-J.; Park, S.-H.; Kim, G.-Y.; Leem, S.-H.; Jin, C.-Y. Cordycepin induces apoptosis in human bladder cancer T24 cells through ROS-dependent inhibition of the PI3K/Akt signaling pathway. Biosci. Trends 2019, 13, 324–333. [Google Scholar] [CrossRef]
- Nasser, M.I.; Masood, M.; Wei, W.; Li, X.; Zhou, Y.; Liu, B.; Li, J.; Li, X. Cordycepin induces apoptosis in SGC-7901 cells through mitochondrial extrinsic phosphorylation of PI3K/Akt by generating ROS. Int. J. Oncol. 2017, 50, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Wellham, P.A.; Kim, D.-H.; Brock, M.; de Moor, C.H. Coupled biosynthesis of cordycepin and pentostatin in Cordyceps militaris: Implications for fungal biology and medicinal natural products. Ann. Transl. Med. 2019, 7, S85. [Google Scholar] [CrossRef]
- Hiraoka, W.; Tanabe, K.; Kuwabara, M.; Sato, F. Metabolic Effects of 3′-Deoxyadenosine (Cordycepin) and 2-Halo-3′-Deoxyadenosine on Repair of X-Ray-Induced Potentially Lethal Damage in Chinese Hamster V79 Cells. Radiat. Res. 1988, 114, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Kefford, R.F.; Taylor, I.W.; Fox, R.M. Flow cytometric analysis of adenosine analogue lymphocytotoxicity. Cancer Res. 1983, 43, 5112–5119. [Google Scholar] [PubMed]
- Zieve, G.W.; Feeney, R.J.; Roemer, E.J. Cordycepin disrupts the microtubule networks and arrests nil 8 hamster fibroblasts at the onset of mitosis. Cell Motil. Cytoskelet. 1987, 7, 337–346. [Google Scholar] [CrossRef]
- Overgaard-Hansen, K. The inhibition of 5-phosphoribosyl-1-pyrophosphate formation by cordycepin triphosphate in extracts of Ehrlich ascites tumor cells. Biochim. Et Biophys. Acta (Bba)-Spec. Sect. Nucleic Acids Relat. Subj. 1964, 80, 504–507. [Google Scholar] [CrossRef]
- Lee, J.B.; Radhi, M.; Cipolla, E.; Gandhi, R.D.; Sarmad, S.; Zgair, A.; Kim, T.H.; Feng, W.; Qin, C.; Adrower, C. A novel nucleoside rescue metabolic pathway may be responsible for therapeutic effect of orally administered cordycepin. Sci. Rep. 2019, 9, 15760. [Google Scholar] [CrossRef]
- Tsai, Y.-J.; Lin, L.-C.; Tsai, T.-H. Pharmacokinetics of adenosine and cordycepin, a bioactive constituent of Cordyceps sinensis in rat. J. Agric. Food Chem. 2010, 58, 4638–4643. [Google Scholar] [CrossRef]
- Li, G.; Nakagome, I.; Hirono, S.; Itoh, T.; Fujiwara, R. Inhibition of adenosine deaminase (ADA)-mediated metabolism of cordycepin by natural substances. Pharmacol. Res. Perspect. 2015, 3, e00121. [Google Scholar] [CrossRef]
- Kodama, E.N.; McCaffrey, R.P.; Yusa, K.; Mitsuya, H. Antileukemic activity and mechanism of action of cordycepin against terminal deoxynucleotidyl transferase-positive (TdT+) leukemic cells. Biochem. Pharmacol. 2000, 59, 273–281. [Google Scholar] [CrossRef]
- Hirschhorn, R. Adenosine deaminase deficiency. Immunodefic. Rev. 1990, 2, 175–198. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.-J.; Tao, R.-Y.; Liu, Q.; Tian, J.-Y.; Ye, F.; Zhu, P.; Zhu, H.-B. Improvement on lipid metabolic disorder by 3′-deoxyadenosine in high-fat-diet-induced fatty mice. Am. J. Chin. Med. 2010, 38, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Rodman, L.E.; Farnell, D.R.; Coyne, J.M.; Allan, P.W.; Hill, D.L.; Duncan, K.L.; Tomaszewski, J.E.; Smith, A.C.; Page, J.G. Toxicity of cordycepin in combination with the adenosine deaminase inhibitor 2′-deoxycoformycin in beagle dogs. Toxicol. Appl. Pharmacol. 1997, 147, 39–45. [Google Scholar] [CrossRef]
- Trigo, P.; Gutteridge, W.; Williamson, J. The effects of cordycepin on malaria parasites. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 514–520. [Google Scholar] [CrossRef]
- Inoue, T.; Murakami, K.; Fujii, T. Mutagenic potential of cordycepin (3′-deoxyadenosine) in Salmonella and soybean tester strains. Mutat. Res. Lett. 1986, 174, 179–182. [Google Scholar] [CrossRef]
- Ramesh, T.; Yoo, S.-K.; Kim, S.-W.; Hwang, S.-Y.; Sohn, S.-H.; Kim, I.-W.; Kim, S.-K. Cordycepin (3′-deoxyadenosine) attenuates age-related oxidative stress and ameliorates antioxidant capacity in rats. Exp. Gerontol. 2012, 47, 979–987. [Google Scholar] [CrossRef]
- Mannick, J.B.; Morris, M.; Hockey, H.-U.P.; Roma, G.; Beibel, M.; Kulmatycki, K.; Watkins, M.; Shavlakadze, T.; Zhou, W.; Quinn, D. TORC1 inhibition enhances immune function and reduces infections in the elderly. Sci. Transl. Med. 2018, 10, eaaq1564. [Google Scholar] [CrossRef]
- Sano, N.; Rajjou, L.; North, H.M. Lost in translation: Physiological roles of stored mrnas in seed germination. Plants 2020, 9, 347. [Google Scholar] [CrossRef]
- Jung, S.-M.; Park, S.-S.; Kim, W.-J.; Moon, S.-K. Ras/ERK1 pathway regulation of p27KIP1-mediated G1-phase cell-cycle arrest in cordycepin-induced inhibition of the proliferation of vascular smooth muscle cells. Eur. J. Pharmacol. 2012, 681, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Imesch, P.; Goerens, A.; Fink, D.; Fedier, A. MLH1-deficient HCT116 colon tumor cells exhibit resistance to the cytostatic and cytotoxic effect of the poly (A) polymerase inhibitor cordycepin (3’-deoxyadenosine) in vitro. Oncol. Lett. 2012, 3, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Wang, J.-Y.; Pan, B.-S.; Mu, Y.-F.; Lai, M.-S.; So, E.C.; Wong, T.-S.; Huang, B.-M. Cordycepin enhances cisplatin apoptotic effect through caspase/MAPK pathways in human head and neck tumor cells. Oncotargets Ther. 2013, 6, 983. [Google Scholar]
- Wang, F.; Yin, P.; Lu, Y.; Zhou, Z.; Jiang, C.; Liu, Y.; Yu, X. Cordycepin prevents oxidative stress-induced inhibition of osteogenesis. Oncotarget 2015, 6, 35496. [Google Scholar] [CrossRef]
- Shao, L.W.; Huang, L.H.; Yan, S.; Jin, J.D.; Ren, S.Y. Cordycepin induces apoptosis in human liver cancer HepG2 cells through extrinsic and intrinsic signaling pathways. Oncol. Lett. 2016, 12, 995–1000. [Google Scholar] [CrossRef]
- Hwang, J.-H.; Joo, J.C.; Kim, D.J.; Jo, E.; Yoo, H.-S.; Lee, K.-B.; Park, S.J.; Jang, I.-S. Cordycepin promotes apoptosis by modulating the ERK-JNK signaling pathway via DUSP5 in renal cancer cells. Am. J. Cancer Res. 2016, 6, 1758. [Google Scholar]
- Li, S.Z.; Ren, J.W.; Fei, J.; Zhang, X.D.; Du, R.L. Cordycepin induces Bax-dependent apoptosis in colorectal cancer cells. Mol. Med. Rep. 2019, 19, 901–908. [Google Scholar] [CrossRef]
- Wu, W.-C.; Hsiao, J.-R.; Lian, Y.-Y.; Lin, C.-Y.; Huang, B.-M. The apoptotic effect of cordycepin on human OEC-M1 oral cancer cell line. Cancer Chemother. Pharmacol. 2007, 60, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Shichiri, H.; Uda, A.; Yamashita, K.; Nishioka, T.; Kume, M.; Makimoto, H.; Nakagawa, T.; Hirano, T.; Hirai, M. Apoptotic effects of the extracts of cordyceps militaris via Erk phosphorylation in a renal cell carcinoma cell line. Phytother. Res. 2015, 29, 707–713. [Google Scholar] [CrossRef]
- Tania, M.; Shawon, J.; Saif, K.; Kiefer, R.; Khorram, M.S.; Halim, M.A.; Khan, M. Cordycepin downregulates Cdk-2 to interfere with cell cycle and increases apoptosis by generating ROS in cervical cancer cells: In vitro and in silico study. Curr. Cancer Drug Targets 2019, 19, 152–159. [Google Scholar] [CrossRef]
- Jiapeng, T.; Yiting, L.; Li, Z. Optimization of fermentation conditions and purification of cordycepin from Cordyceps militaris. Prep. Biochem. Biotechnol. 2014, 44, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Debnath, T.; Kim, S.-K.; Lim, B.O. Anti-cancer effect and apoptosis induction of cordycepin through DR3 pathway in the human colonic cancer cell HT-29. Food Chem. Toxicol. 2013, 60, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-S.; Kwon, H.-Y.; Kim, K.-S. Cordycepin induces apoptosis in human neuroblastoma SK-N-BE (2)-C and melanoma SK-MEL-2 cells. Indian J. Biochem. Biophys. 2012, 49, 86–91. [Google Scholar] [PubMed]
- Shi, P.; Huang, Z.; Tan, X.; Chen, G. Proteomic detection of changes in protein expression induced by cordycepin in human hepatocellular carcinoma BEL-7402 cells. Methods Find. Exp. Clin. Pharmacol. 2008, 30, 347. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.; Seibert, G.; Beyer, R.; Breter, H.J.; Maidhof, A.; Zahn, R.K. Effect of cordycepin on nucleic acid metabolism in L5178Y cells and on nucleic acid-synthesizing enzyme systems. Cancer Res. 1977, 37, 3824–3833. [Google Scholar]
- Rosowsky, A.; Lazarus, H.; Yamashita, A. Nucleosides. 1. 9-(3’-Alkyl-3’-deoxy-. beta.-D-ribofuranosyl) adenines as lipophilic analogs of cordycepin. Synthesis and preliminary biological studies. J. Med. Chem. 1976, 19, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-C.; Zhou, X.-P.; Wang, X.-A.; Xu, M.-D.; Chen, T.; Chen, T.-Y.; Zhou, P.-H.; Zhang, Y.-Q. Cordycepin induces apoptosis and G2/M phase arrest through the ERK pathways in esophageal cancer cells. J. Cancer 2019, 10, 2415. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Qi, M.; Li, L.; Yuan, Y.; Wu, X.; Fu, J. Natural cordycepin induces apoptosis and suppresses metastasis in breast cancer cells by inhibiting the Hedgehog pathway. Food Funct. 2020, 11, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Tao, L.; Guo, W.; Wu, Z.-X.; Du, H.; Wang, J.; Zhang, J.; Chen, H.; Chen, Z.-S.; Lin, L. Combination of Cordycepin and Apatinib synergistically inhibits NSCLC cells by down-regulating VEGF/PI3K/Akt signaling pathway. Front. Oncol. 2020, 10, 1732. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Y.; Tsai, S.-W.; Chien, H.-H.; Chen, T.-Y.; Sheu, S.-Y.; So, E.C.; Huang, B.-M. Cordycepin Inhibits Human Gestational Choriocarcinoma Cell Growth by Disrupting Centrosome Homeostasis. Drug Des. Dev. Ther. 2020, 14, 2987. [Google Scholar] [CrossRef] [PubMed]
- Kengkittipat, W.; Kaewmalun, S.; Khongkow, M.; Iempridee, T.; Jantimaporn, A.; Bunwatcharaphansakun, P.; Yostawonkul, J.; Yata, T.; Phoolcharoen, W.; Namdee, K. Improvement of the multi-performance biocharacteristics of cordycepin using BiloNiosome-core/chitosan-shell hybrid nanocarriers. Colloids Surf. B: Biointerfaces 2021, 197, 111369. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, D.-L.; Zhou, M.; He, M.-F.; Huang, S.; Liao, X.-Z.; Zhang, J.-X. Cordycepin enhances the chemosensitivity of esophageal cancer cells to cisplatin by inducing the activation of AMPK and suppressing the AKT signaling pathway. Cell Death Dis. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Chang, M.-M.; Hong, S.-Y.; Yang, S.-H.; Wu, C.-C.; Wang, C.-Y.; Huang, B.-M. Anti-Cancer Effect of Cordycepin on FGF9-Induced Testicular Tumorigenesis. Int. J. Mol. Sci. 2020, 21, 8336. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.-C.; Lin, Y.-T.; Huang, S.-H.; Wang, S.-M. Cordycepin Induces Apoptosis of CGTH W-2 Thyroid Carcinoma Cells through the Calcium− Calpain− Caspase 7-PARP Pathway. J. Agric. Food Chem. 2010, 58, 11645–11652. [Google Scholar] [CrossRef]
- Lu, H.; Li, X.; Zhang, J.; Shi, H.; Zhu, X.; He, X. Effects of cordycepin on HepG2 and EA. hy926 cells: Potential antiproliferative, antimetastatic and anti-angiogenic effects on hepatocellular carcinoma. Oncol. Lett. 2014, 7, 1556–1562. [Google Scholar] [CrossRef]
- Cho, S.H.; Kang, I.-C. The inhibitory effect of Cordycepin on the proliferation of cisplatin-resistant A549 lung cancer cells. Biochem. Biophys. Res. Commun. 2018, 498, 431–436. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Liu, J.; Xu, P.; Zhang, X.M.; Tian, Y.Y.; Xue, Y.M.; Gao, X.Y.; Liu, Y.; Wang, J.H. Synergistic effect of HMGB1 knockdown and cordycepin in the K562 human chronic myeloid leukemia cell line. Mol. Med. Rep. 2015, 12, 4462–4468. [Google Scholar] [CrossRef]
- Chang, M.M.; Pan, B.S.; Wang, C.Y.; Huang, B.M. Cordycepin-induced unfolded protein response-dependent cell death, and AKT/MAPK-mediated drug resistance in mouse testicular tumor cells. Cancer Med. 2019, 8, 3949–3964. [Google Scholar] [CrossRef]
- Hu, P.; Chen, W.; Bao, J.; Jiang, L.; Wu, L. Cordycepin modulates inflammatory and catabolic gene expression in interleukin-1beta-induced human chondrocytes from advanced-stage osteoarthritis: An in vitro study. Int. J. Clin. Exp. Pathol. 2014, 7, 6575. [Google Scholar] [PubMed]
- Ryu, E.; Son, M.; Lee, M.; Lee, K.; Cho, J.Y.; Cho, S.; Lee, S.K.; Lee, Y.M.; Cho, H.; Sung, G.-H. Cordycepin is a novel chemical suppressor of Epstein-Barr virus replication. Oncoscience 2014, 1, 866. [Google Scholar] [CrossRef]
- Dou, C.; Cao, Z.; Ding, N.; Hou, T.; Luo, F.; Kang, F.; Yang, X.; Jiang, H.; Xie, Z.; Hu, M. Cordycepin prevents bone loss through inhibiting osteoclastogenesis by scavenging ROS generation. Nutrients 2016, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Ouyang, Z.; Su, Z. Cordycepin protects PC12 cells against 6-hydroxydopamine induced neurotoxicity via its antioxidant properties. Biomed. Pharmacother. 2016, 81, 7–14. [Google Scholar] [CrossRef]
- Yu, S.-B.; Kim, H.-J.; Kang, H.-M.; Park, B.-S.; Lee, J.-H.; Kim, I.-R. Cordycepin accelerates osteoblast mineralization and attenuates osteoclast differentiation in vitro. Evid. -Based Complementary Altern. Med. 2018, 2018, 5892957. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-X.; Chen, Y.; Yang, Y.; Chen, X.-X.; Zhang, D.-D. Screening five Qi-Tonifying herbs on M2 phenotype macrophages. Evid. -Based Complementary Altern. Med. 2019, 2019. [Google Scholar] [CrossRef]
- Zhou, X.; Meyer, C.U.; Schmidtke, P.; Zepp, F. Effect of cordycepin on interleukin-10 production of human peripheral blood mononuclear cells. Eur. J. Pharmacol. 2002, 453, 309–317. [Google Scholar] [CrossRef]
- Potten, C.; Darzynkiewicz, Z.; Sasaki, K.; Reshkin, S.; Tedone, T.; Correale, M.; Mangia, A.; Casavola, V.; Paradiso, A. Association of pS2 (TFF1) release with breast tumour proliferative rate: In vitro and in vivo studies. Cell Prolif. 1999, 32, 107–118. [Google Scholar] [CrossRef]
- Maytin, E.V. Differential effects of heat shock and UVB light upon stress protein expression in epidermal keratinocytes. J. Biol. Chem. 1992, 267, 23189–23196. [Google Scholar] [CrossRef]
- Jaiboonma, A.; Kaokaen, P.; Chaicharoenaudomrung, N.; Kunhorm, P.; Janebodin, K.; Noisa, P.; Jitprasertwong, P. Cordycepin attenuates salivary hypofunction through the prevention of oxidative stress in human submandibular gland cells. Int. J. Med. Sci. 2020, 17, 1733. [Google Scholar] [CrossRef]
- Wang, C.-H.; Chang, C.-H.; Lin, T.-L.; Fu, R.-H.; Huang, Y.-C.; Chen, S.-Y.; Shyu, W.-C.; Liu, S.-P. The novel application of cordycepin in maintaining stem cell pluripotency and increasing iPS cell generation efficiency. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Seong, D.B.; Hong, S.; Muthusami, S.; Kim, W.-D.; Yu, J.-R.; Park, W.-Y. Cordycepin increases radiosensitivity in cervical cancer cells by overriding or prolonging radiation-induced G2/M arrest. Eur. J. Pharmacol. 2016, 771, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Moon, G.-S.; Jung, K.-H.; Kim, W.-J.; Moon, S.-K. c-Jun N-terminal kinase 1 is required for cordycepin-mediated induction of G2/M cell-cycle arrest via p21WAF1 expression in human colon cancer cells. Food Chem. Toxicol. 2010, 48, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-W.; Hsu, W.-H.; Tai, C.-J. Antimetastatic effects of cordycepin mediated by the inhibition of mitochondrial activity and estrogen-related receptor α in human ovarian carcinoma cells. Oncotarget 2017, 8, 3049. [Google Scholar] [CrossRef] [PubMed]
- Yoou, M.-s.; Yoon, K.W.; Choi, Y.; Kim, H.-M.; Jeong, H.-J. Cordycepin diminishes thymic stromal lymphopoietin-induced interleukin-13 production. Eur. J. Pharmacol. 2017, 802, 1–6. [Google Scholar] [CrossRef]
- Yoou, M.-s.; Jin, M.H.; Lee, S.Y.; Lee, S.H.; Kim, B.; Roh, S.S.; Choi, I.H.; Lee, M.S.; Kim, H.-M.; Jeong, H.-J. Cordycepin suppresses thymic stromal lymphopoietin expression via blocking caspase-1 and receptor-interacting protein 2 signaling pathways in mast cells. Biol. Pharm. Bull. 2016, 39, 90–96. [Google Scholar] [CrossRef][Green Version]
- Shin, S.; Lee, S.; Kwon, J.; Moon, S.; Lee, S.; Lee, C.-K.; Cho, K.; Ha, N.-J.; Kim, K. Cordycepin suppresses expression of diabetes regulating genes by inhibition of lipopolysaccharide-induced inflammation in macrophages. Immune Netw. 2009, 9, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Moon, S.; Park, Y.; Kwon, J.; Lee, S.; Lee, C.-K.; Cho, K.; Ha, N.-J.; Kim, K. Role of cordycepin and adenosine on the phenotypic switch of macrophages via induced anti-inflammatory cytokines. Immune Netw. 2009, 9, 255–264. [Google Scholar] [CrossRef]
- Noh, E.-M.; Kim, J.-S.; Hur, H.; Park, B.-H.; Song, E.-K.; Han, M.-K.; Kwon, K.-B.; Yoo, W.-H.; Shim, I.-K.; Lee, S. Cordycepin inhibits IL-1β-induced MMP-1 and MMP-3 expression in rheumatoid arthritis synovial fibroblasts. Rheumatology 2009, 48, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Imamura, K.; Asai, M.; Sugamoto, K.; Matsumoto, T.; Yamasaki, Y.; Kamei, I.; Hattori, T.; Kishimoto, M.; Niisaka, S.; Kubo, M. Suppressing effect of cordycepin on the lipopolysaccharide-induced nitric oxide production in RAW 264.7 cells. Biosci. Biotechnol. Biochem. 2015, 79, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Qing, R.; Huang, Z.; Tang, Y.; Xiang, Q.; Yang, F. Cordycepin negatively modulates lipopolysaccharide-induced cytokine production by up-regulation of heme oxygenase-1. Int. Immunopharmacol. 2017, 47, 20–27. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.-z.; Hylemon, P.B.; Zhang, L.-y.; Zhou, H.-p. Cordycepin inhibits LPS-induced inflammatory responses by modulating NOD-Like Receptor Protein 3 inflammasome activation. Biomed. Pharmacother. 2017, 95, 1777–1788. [Google Scholar] [CrossRef]
- Lu, M.-Y.; Chen, C.-C.; Lee, L.-Y.; Lin, T.-W.; Kuo, C.-F. N 6-(2-Hydroxyethyl) adenosine in the medicinal mushroom Cordyceps cicadae attenuates lipopolysaccharide-stimulated pro-inflammatory responses by suppressing TLR4-mediated NF-κB signaling pathways. J. Nat. Prod. 2015, 78, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.J.; Kim, M.J.; Lee, H.H.; Park, J.U.; Kang, B.W.; Kim, G.-Y.; Rhu, E.J.; Kim, J.-I.; Kim, K.H.; Jeong, Y.K. Effect of cordycepin on the expression of the inflammatory cytokines TNF-alpha, IL-6, and IL-17A in C57BL/6 mice. J. Microbiol. Biotechnol. 2013, 23, 156–160. [Google Scholar] [CrossRef]
- Ren, Z.; Cui, J.; Huo, Z.; Xue, J.; Cui, H.; Luo, B.; Jiang, L.; Yang, R. Cordycepin suppresses TNF-α-induced NF-κB activation by reducing p65 transcriptional activity, inhibiting IκBα phosphorylation, and blocking IKKγ ubiquitination. Int. Immunopharmacol. 2012, 14, 698–703. [Google Scholar] [CrossRef]
- Jeong, M.-H.; Seo, M.J.; Park, J.U.; Kang, B.W.; Kim, K.-S.; Lee, J.Y.; Kim, G.-Y.; Kim, J.-I.; Choi, Y.H.; Kim, K.H. Effect of cordycepin purified from Cordyceps militaris on Th1 and Th2 cytokines in mouse splenocytes. J. Microbiol. Biotechnol. 2012, 22, 1161–1164. [Google Scholar] [CrossRef]
- Aksamit, R.; Backlund, P.; Cantoni, G. Chemotaxis and the synthesis of specific proteins are inhibited by 3-deazaadenosine and other adenosine analogs in a mouse macrophage cell line. J. Biol. Chem. 1983, 258, 20–23. [Google Scholar] [CrossRef]
- Sugar, A.M.; McCaffrey, R.P. Antifungal activity of 3′-deoxyadenosine (cordycepin). Antimicrob. Agents Chemother. 1998, 42, 1424–1427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-w.; Deng, H.; Qi, W.; Zhao, G.-y.; Cao, X.-r. Osteoprotective effect of cordycepin on estrogen deficiency-induced osteoporosis in vitro and in vivo. Biomed Res. Int. 2015, 2015, 423869. [Google Scholar] [PubMed]
- Chen, Y.-X.; Zhu, D.-Y.; Xu, Z.-L.; Yin, J.-H.; Yu, X.-W.; Mei, J.; Gao, Y.-S.; Zhang, C.-Q. The protective effect of cordycepin on alcohol-induced osteonecrosis of the femoral head. Cell. Physiol. Biochem. 2017, 42, 2391–2403. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radhi, M.; Ashraf, S.; Lawrence, S.; Tranholm, A.A.; Wellham, P.A.D.; Hafeez, A.; Khamis, A.S.; Thomas, R.; McWilliams, D.; de Moor, C.H. A Systematic Review of the Biological Effects of Cordycepin. Molecules 2021, 26, 5886. https://doi.org/10.3390/molecules26195886
Radhi M, Ashraf S, Lawrence S, Tranholm AA, Wellham PAD, Hafeez A, Khamis AS, Thomas R, McWilliams D, de Moor CH. A Systematic Review of the Biological Effects of Cordycepin. Molecules. 2021; 26(19):5886. https://doi.org/10.3390/molecules26195886
Chicago/Turabian StyleRadhi, Masar, Sadaf Ashraf, Steven Lawrence, Asta Arendt Tranholm, Peter Arthur David Wellham, Abdul Hafeez, Ammar Sabah Khamis, Robert Thomas, Daniel McWilliams, and Cornelia Huiberdina de Moor. 2021. "A Systematic Review of the Biological Effects of Cordycepin" Molecules 26, no. 19: 5886. https://doi.org/10.3390/molecules26195886
APA StyleRadhi, M., Ashraf, S., Lawrence, S., Tranholm, A. A., Wellham, P. A. D., Hafeez, A., Khamis, A. S., Thomas, R., McWilliams, D., & de Moor, C. H. (2021). A Systematic Review of the Biological Effects of Cordycepin. Molecules, 26(19), 5886. https://doi.org/10.3390/molecules26195886