
The Short-Term Exposure to SDHI Fungicides Boscalid and Bixafen Induces a Mitochondrial Dysfunction in Selective Human Cell Lines



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
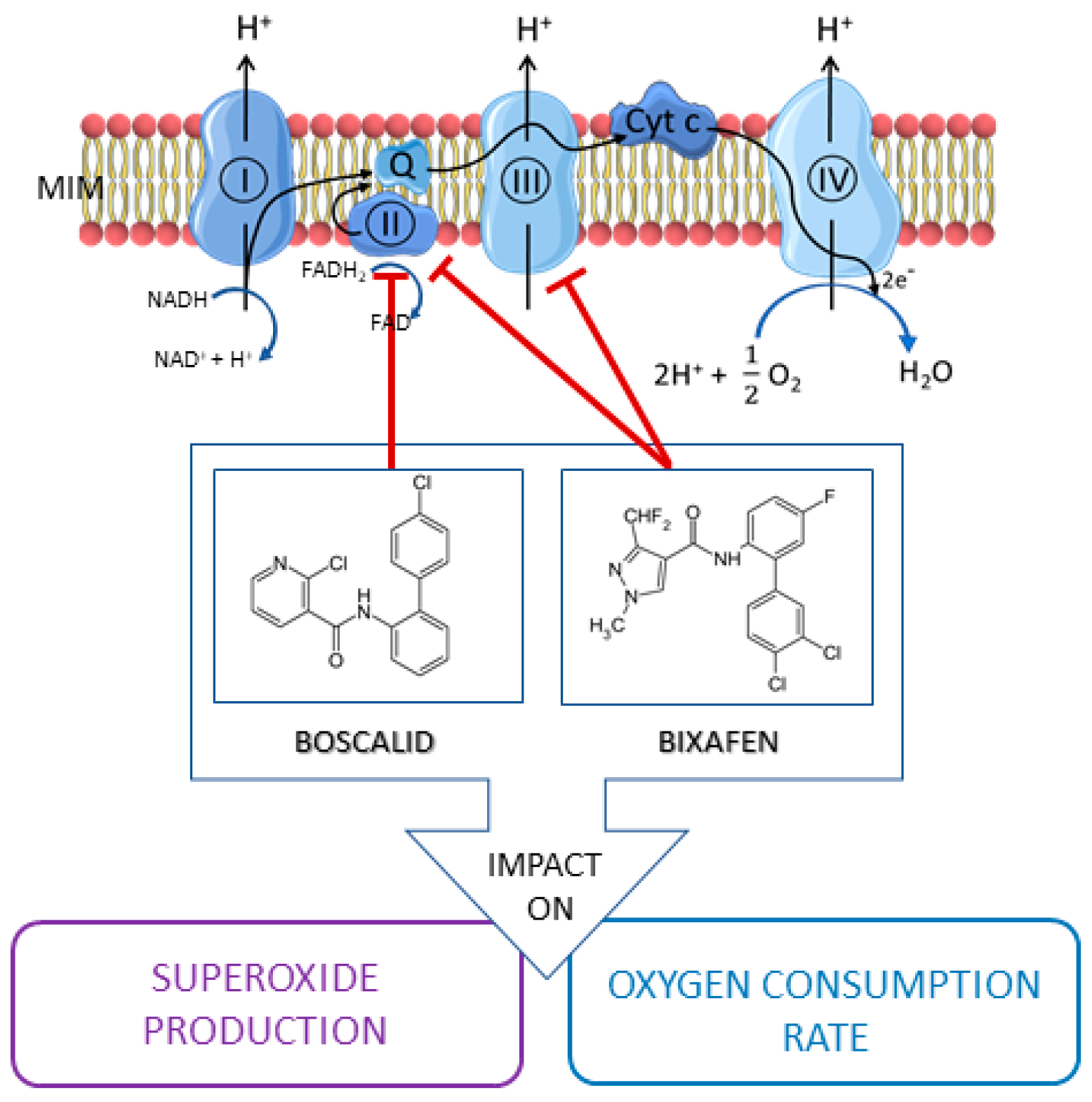
:1. Introduction
2. Results
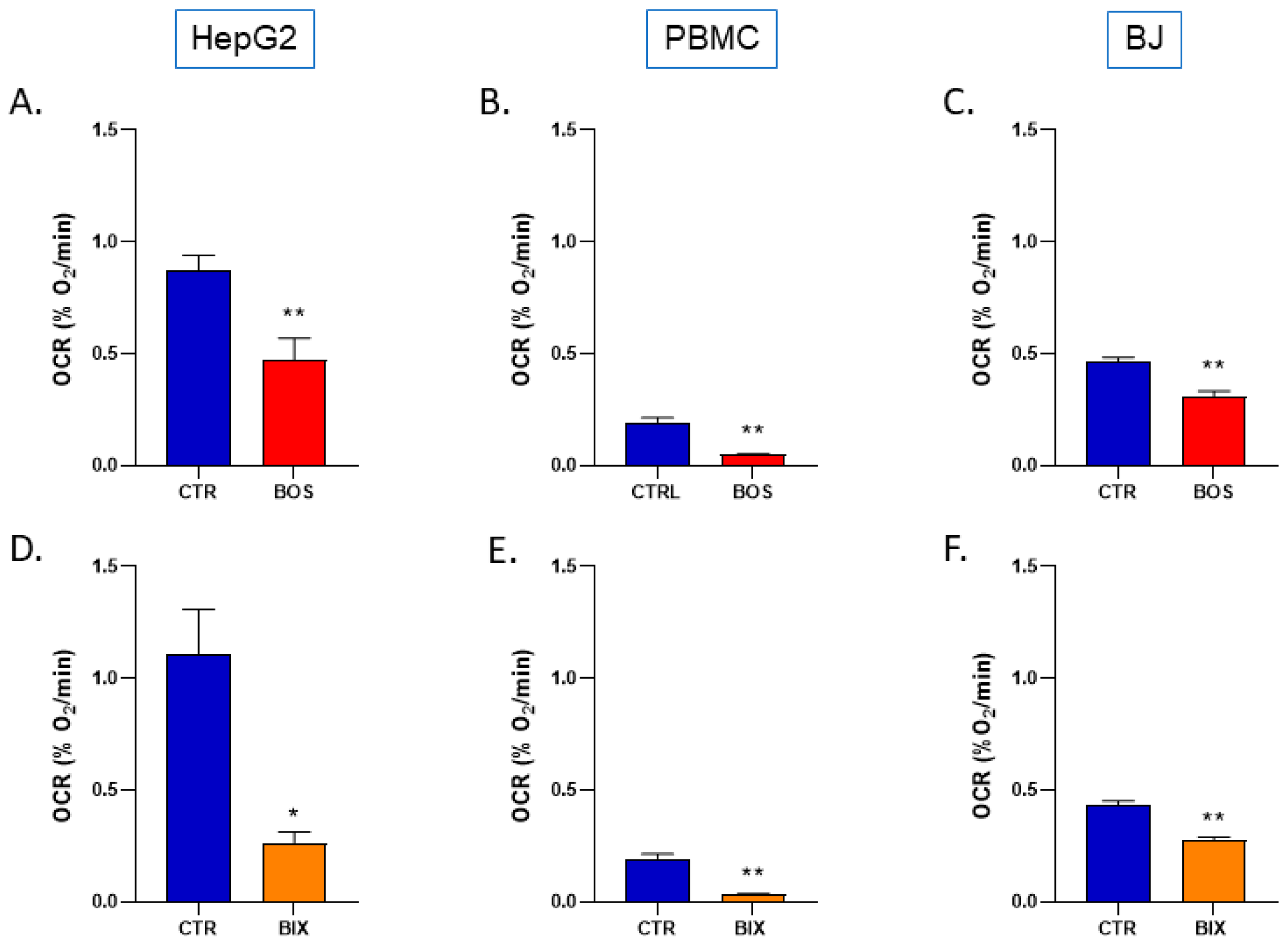
2.1. Boscalid and Bixafen Decreased OCR in the Three Human Cell Lines
2.2. Impact of SDHIs on Mitochondrial Superoxide Generation in Human Cell Lines
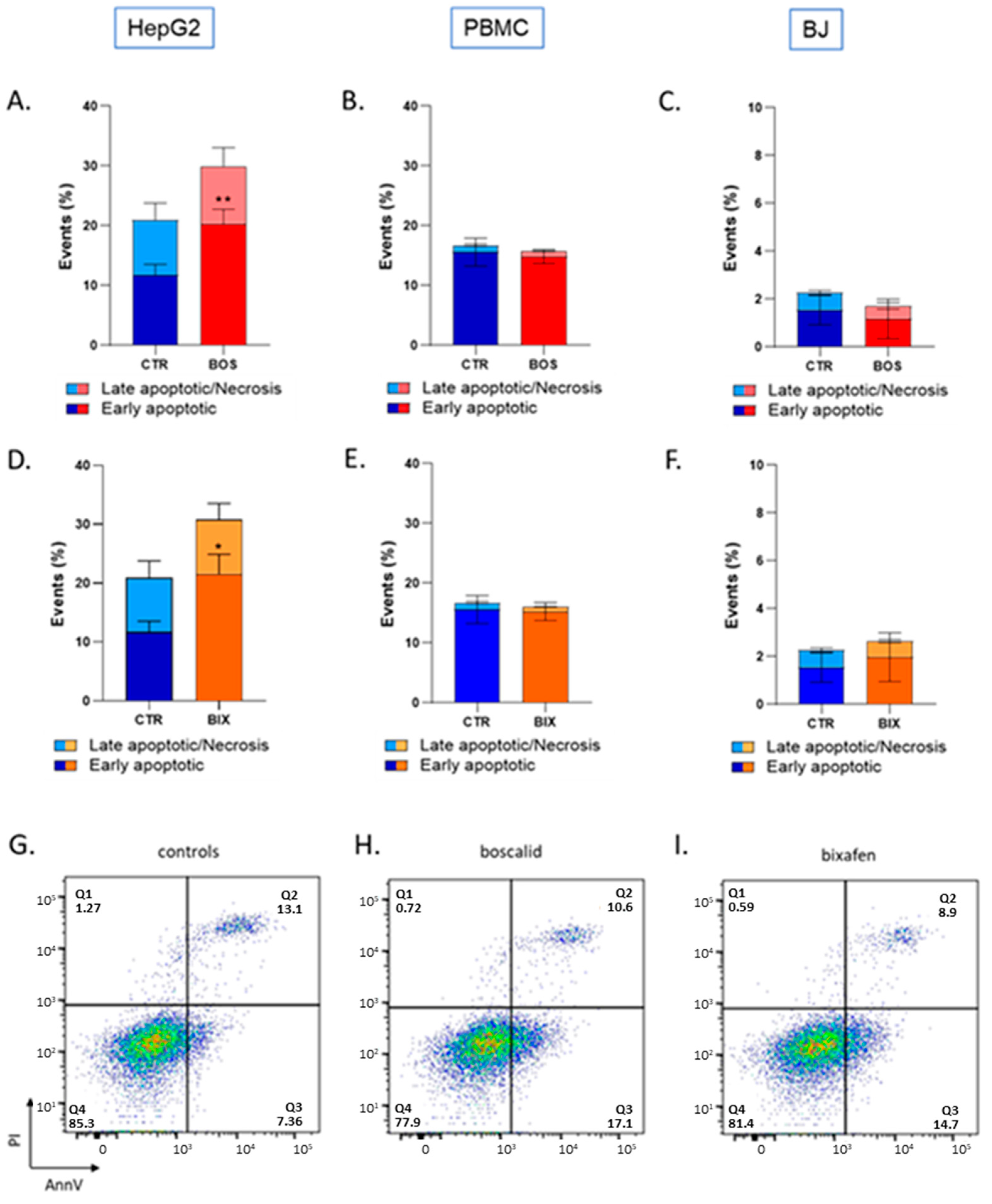
2.3. Bixafen and Boscalid Induced Apoptosis in HepG2 Cells, but Not in PBMC and BJ Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Culture
4.3. EPR Oximetry
4.4. EPR Superoxide Measurement
4.5. Cell Death Detection by Flow Cytometry
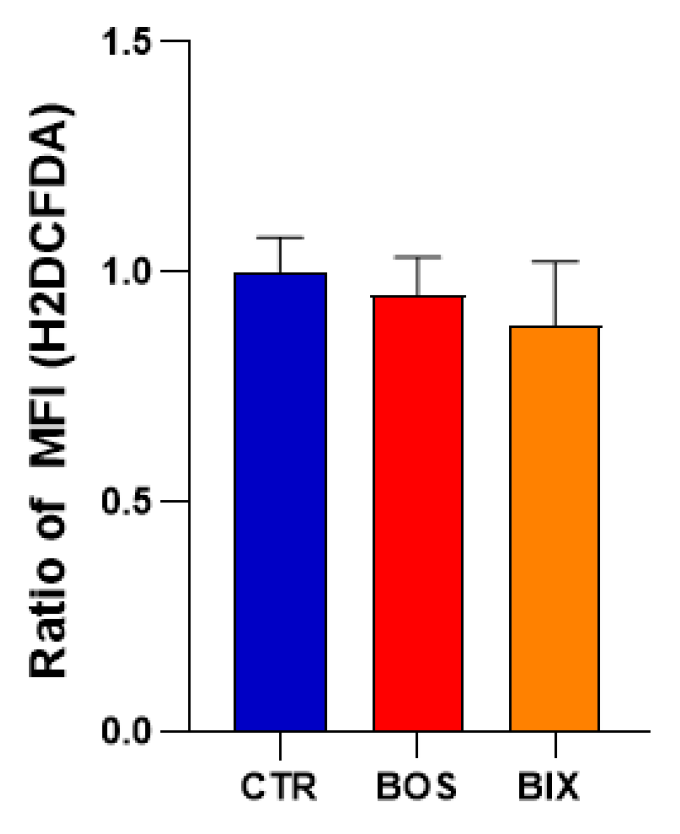
4.6. Global ROS Production Using H2DCFDA
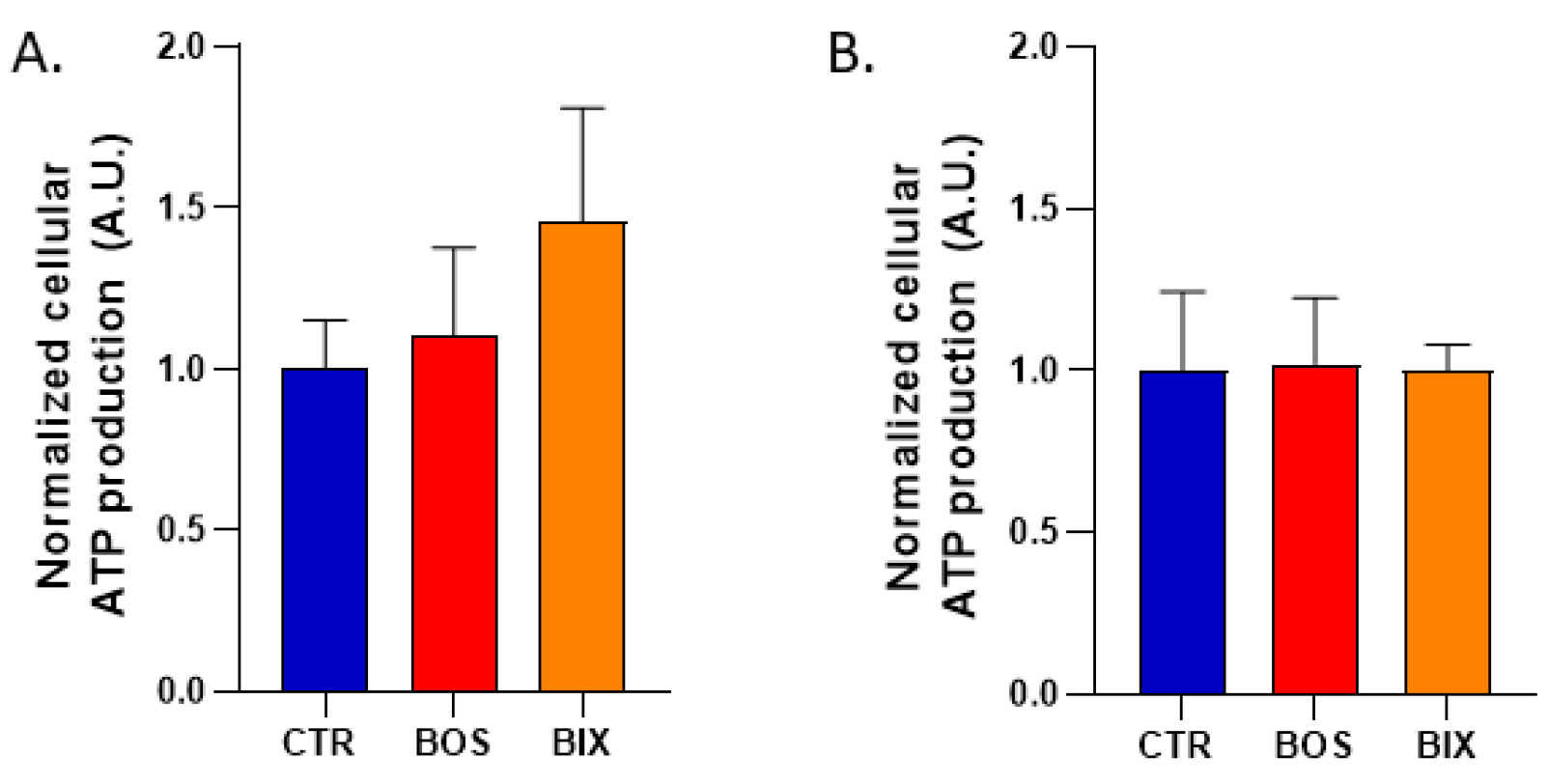
4.7. Cellular ATP Production
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Umetsu, N.; Shirai, Y. Development of novel pesticides in the 21th century. J. Pestic. Sci. 2020, 45, 54–74. [Google Scholar] [CrossRef] [Green Version]
- ANSES Report 2018-SA-0113. Available online: https://www.anses.fr/fr/system/files/PHYTO2018SA0113Ra.pdf (accessed on 26 July 2021).
- Kamp, H.; Wahrheit, J.; Stinchcombe, S.; Walk, T.; Stauber, F.; Ravenzwaay, B. Succinate dehydrogenase inhibitors: In silico flux analysis and in vivo metabolomics investigations show no severe metabolic consequences for rats and humans. Food Chem. Toxicol. 2021, 150, 112085. [Google Scholar] [CrossRef]
- Bénit, P.; Kahn, A.; Chretien, D.; Bortoli, S.; Huc, L.; Schiff, M.; Gimenez-Roqueplo, A.-P.; Favier, J.; Gressens, P.; Rak, M.; et al. Evolutionarily conserved susceptibility of the mitochondrial respiratory chain to SDHI pesticides and its consequence on the impact of SDHIs on human cultured cells. PLoS ONE 2019, 14, e0224132. [Google Scholar] [CrossRef] [Green Version]
- Brenet, A.; Hassan-Abdi, R.; Soussi-Yanicostas, N. Bixafen, a succinate dehydrogenase inhibitor fungicide, causes microcephaly and motor neuron axon defects during development. Chemosphere 2020, 265, 128781. [Google Scholar] [CrossRef]
- Wang, H.; Meng, Z.; Liu, F.; Zhou, L.; Su, M.; Meng, Y.; Zhang, S.; Liao, X.; Cao, Z.; Lu, H. Characterization of boscalid-induced oxidative stress and neurodevelopmental toxicity in zebrafish embryos. Chemosphere 2019, 238, 124753. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Li, W.; Xiao, P. Bixafen causes cardiac toxicity in zebrafish (Danio rerio) embryos. Environ. Sci. Pollut. Res. 2021, 28, 1–11. [Google Scholar] [CrossRef]
- Qian, L.; Zhang, J.; Chen, X.; Qi, S.; Wu, P.; Wang, C.; Wang, C. Toxic effects of boscalid in adult zebrafish (Danio rerio) on carbohydrate and lipid metabolism. Environ. Pollut. 2019, 247, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Cayyir, A.; Coskun, M.; Coskun, M. Micronuclei, nucleoplasmic bridges, and nuclear buds induced in human lymphocytes by the fungicide signum and its active ingredients (boscalid and pyraclostrobin). Environ. Toxicol. 2014, 29, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Graillot, V.; Tomasetig, F.; Cravedi, J.-P.; Audebert, M. Evidence of the in vitro genotoxicity of methyl-pyrazole pesticides in human cells. Mutat. Res. Toxicol. Environ. Mutagen. 2012, 748, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Evolutionarily Conserved Susceptibility of the Mitochondrial Respiratory Chain to SDHI Pesticides and Its Consequence on the Impact of SDHIs on Human Cultured Cells. Available online: https://journals.plos.org/plosone/article/comment?id=10.1371/annotation/400d52f8-f61f-4132-a94c-bee53565f35c (accessed on 26 July 2021).
- Pesticide Fact Sheet. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/fs_PC-128008_01-Jul-03.pdf (accessed on 26 July 2021).
- BIXAFEN (262). Available online: http://www.fao.org/fileadmin/templates/agphome/documents/Pests_Pesticides/JMPR/Evaluation13/Bixafen.pdf (accessed on 26 July 2021).
- European Food Safety Authority (EFSA). Conclusion on Pesticide Peer Review. EFSA J. 2012, 10, 2917. [Google Scholar]
- Pozza, E.D.; Dando, I.; Pacchiana, R.; Liboi, E.; Scupoli, M.T.; Donadelli, M.; Palmieri, M. Regulation of succinate dehydrogenase and role of succinate in cancer. Semin. Cell Dev. Biol. 2019, 98, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Lu, M.; Andera, L.; Lahm, H.; Gellert, N.; Fariss, M.W.; Korinek, V.; Sattler, W.; Ucker, D.S.; Terman, A.; et al. Vitamin E succinate is a potent novel antineoplastic agent with high selectivity and cooperativity with tumor necrosis factor-related apoptosis-inducing ligand (Apo2 ligand) in vivo. Clin. Cancer Res. 2002, 8, 863–869. [Google Scholar] [PubMed]
- Dong, L.-F.; Jameson, V.J.; Tilly, D.; Prochazka, L.; Rohlena, J.; Valis, K.; Truksa, J.; Zobalova, R.; Mahdavian, E.; Kluckova, K.; et al. Mitochondrial targeting of α-tocopheryl succinate enhances its pro-apoptotic efficacy: A new paradigm for effective cancer therapy. Free. Radic. Biol. Med. 2011, 50, 1546–1555. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, M.G.; Galindo, M.F.; De Mera, R.M.M.-F.; Fernandez-Gomez, F.J.; Concannon, C.G.; Segura, M.F.; Comella, J.X.; Prehn, J.H.M.; Jordan, J.; De Mera, R.M.M.F. Reactive Oxygen Species and p38 Mitogen-Activated Protein Kinase Activate Bax to Induce Mitochondrial Cytochrome c Release and Apoptosis in Response to Malonate. Mol. Pharmacol. 2006, 71, 736–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.-S.; Sun, G.; Cobessi, D.; Wang, A.C.; Shen, J.T.; Tung, E.Y.; Anderson, V.E.; Berry, E.A. 3-Nitropropionic Acid Is a Suicide Inhibitor of Mitochondrial Respiration That, upon Oxidation by Complex II, Forms a Covalent Adduct with a Catalytic Base Arginine in the Active Site of the Enzyme. J. Biol. Chem. 2006, 281, 5965–5972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, G.; Zhang, Q.; Pan, J.; Lee, Y.; Ouari, O.; Hardy, M.; Zielonka, M.; Myers, C.R.; Zielonka, J.; Weh, K.; et al. Targeting lonidamine to mitochondria mitigates lung tumorigenesis and brain metastasis. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, B.F.; Grégoire, V.; Demeure, R.J.; Sonveaux, P.; Feron, O.; O’Hara, J.; VanHulle, V.P.; Delzenne, N.; Gallez, B. Insulin increases the sensitivity of tumors to irradiation: Involvement of an increase in tumor oxygenation mediated by a nitric oxide-dependent decrease of the tumor cells oxygen consumption. Cancer Res. 2002, 62, 3555–3561. [Google Scholar]
- Diepart, C.; Verrax, J.; Calderon, P.B.; Feron, O.; Jordan, B.F.; Gallez, B. Comparison of methods for measuring oxygen consumption in tumor cells in vitro. Anal. Biochem. 2009, 396, 250–256. [Google Scholar] [CrossRef]
- D’Hose, D.; Danhier, P.; Northshield, H.; Isenborghs, P.; Jordan, B.F.; Gallez, B. A versatile EPR toolbox for the simultaneous measurement of oxygen consumption and superoxide production. Redox Biol. 2020, 40, 101852. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Kirilyuk, I.; Voynov, M.; Grigor’Ev, I. EPR detection of cellular and mitochondrial superoxide using cyclic hydroxylamines. Free. Radic. Res. 2010, 45, 417–430. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for Detection of Mitochondrial and Cellular Reactive Oxygen Species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Scheinok, S.; Leveque, P.; Sonveaux, P.; Driesschaert, B.; Gallez, B. Comparison of different methods for measuring the superoxide radical by EPR spectroscopy in buffer, cell lysates and cells. Free Radic. Res. 2018, 52, 1182–1196. [Google Scholar] [CrossRef]
- Scheinok, S.; Driesschaert, B.; D’Hose, D.; Sonveaux, P.; Robiette, R.; Gallez, B. Synthesis and characterization of a 5-membered ring cyclic hydroxylamine coupled to triphenylphosphonium to detect mitochondrial superoxide by EPR spectrometry. Free Radic. Res. 2019, 53, 1135–1143. [Google Scholar] [CrossRef]
- Scheinok, S.; Capeloa, T.; Porporato, P.E.; Sonveaux, P.; Gallez, B. An EPR Study Using Cyclic Hydroxylamines to Assess the Level of Mitochondrial ROS in Superinvasive Cancer Cells. Cell Biophys. 2020, 78, 249–254. [Google Scholar] [CrossRef]
- d’Hose, D.; Gallez, B. Measurement of mitochondrial (dys)function in cellular systems using Electron Paramagnetic Resonance (EPR): Oxygen consumption rate and superoxide production. Methods Mol. Biol. 2021, in press. [Google Scholar]
- Lorenz, M.; Saretzki, G.; Sitte, N.; Metzkow, S.; von Zglinicki, T. BJ fibroblasts display high antioxidant capacity and slow telomere shortening independent of hTERT transfection. Free. Radic. Biol. Med. 2001, 31, 824–831. [Google Scholar] [CrossRef]
- Qian, L.; Qi, S.; Cao, F.; Zhang, J.; Zhao, F.; Li, C.; Wang, C. Toxic effects of boscalid on the growth, photosynthesis, antioxidant system and metabolism of Chlorella vulgaris. Environ. Pollut. 2018, 242, 171–181. [Google Scholar] [CrossRef]
- Hardy, M.; Zielonka, J.; Karoui, H.; Sikora, A.; Michalski, R.; Podsiadły, R.; Lopez, M.; Vasquez-Vivar, J.; Kalyanaraman, B.; Ouari, O. Detection and Characterization of Reactive Oxygen and Nitrogen Species in Biological Systems by Monitoring Species-Specific Products. Antioxid Redox Signal. 2018, 28, 1416–1432. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Hardy, M.; Podsiadly, R.; Cheng, G.; Zielonka, J. Recent developments in detection of superoxide radical anion and hydrogen peroxide: Opportunities, challenges, and implications in redox signaling. Arch. Biochem. Biophys. 2016, 617, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyanaraman, B.; Dranka, B.; Hardy, M.; Michalski, R.; Zielonka, J. HPLC-based monitoring of products formed from hydroethidine-based fluorogenic probes—The ultimate approach for intra- and extracellular superoxide detection. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1840, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knasmüller, S.; Schwab, C.E.; Land, S.J.; Wang, C.Y.; Sanyal, R.; Kundi, M.; Parzefall, W.; Darroudi, F. Genotoxic effects of heterocyclic aromatic amines in human derived hepatoma (HepG2) cells. Mutagenesis 1999, 14, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Schoonen, W.G.; de Roos, J.A.; Westerink, W.M.; Débiton, E. Cytotoxic effects of 110 reference compounds on HepG2 cells and for 60 compounds on HeLa, ECC-1 and CHO cells: II Mechanistic assays on NAD(P)H, ATP and DNA contents. Toxicol. Vitr. 2005, 19, 491–503. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.J.; Irwin, W.; Diaz, D.; Howard-Cofield, E.; Krejsa, C.M.; Slaughter, M.R.; Gao, B.; Kaludercic, N.; Angeline, A.; Bernardi, P.; et al. High concordance of drug-induced human hepatotoxicity with in vitro cytotoxicity measured in a novel cell-based model using high content screening. Arch. Toxicol. 2006, 80, 580–604. [Google Scholar] [CrossRef]
- Gerets, H.; Hanon, E.; Cornet, M.; Dhalluin, S.; Depelchin, O.; Canning, M.; Atienzar, F. Selection of cytotoxicity markers for the screening of new chemical entities in a pharmaceutical context: A preliminary study using a multiplexing approach. Toxicol. Vitr. 2009, 23, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Briere, J.-J.; Favier, J.; El Ghouzzi, V.; Djouadi, F.; Bénit, P.; Gimenez, A.-P.; Rustin, P. Succinate dehydrogenase deficiency in human. Experientia 2005, 62, 2317–2324. [Google Scholar] [CrossRef] [PubMed]
- Polledri, E.; Mercadante, R.; Nijssen, R.; Consonni, D.; Mol, H.; Fustinoni, S. Hair as a matrix to evaluate cumulative and aggregate exposure to pesticides in winegrowers. Sci. Total. Environ. 2019, 687, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Polledri, E.; Mercadante, R.; Consonni, D.; Fustinoni, S. Cumulative pesticides exposure of children and their parents living near vineyards by hair analysis. Int. J. Environ. Res. Public Health 2021, 18, 3723. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S.; Minor, R.L., Jr.; White, C.W.; Repine, J.E.; Rosen, G.M.; Freeman, B.A. Superoxide dismutase and catalase conjugated to polyethylene glycol increases endothelial enzyme activity and oxidant resistance. J. Biol. Chem. 1988, 263, 6884–6892. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
d’Hose, D.; Isenborghs, P.; Brusa, D.; Jordan, B.F.; Gallez, B. The Short-Term Exposure to SDHI Fungicides Boscalid and Bixafen Induces a Mitochondrial Dysfunction in Selective Human Cell Lines. Molecules 2021, 26, 5842. https://doi.org/10.3390/molecules26195842
d’Hose D, Isenborghs P, Brusa D, Jordan BF, Gallez B. The Short-Term Exposure to SDHI Fungicides Boscalid and Bixafen Induces a Mitochondrial Dysfunction in Selective Human Cell Lines. Molecules. 2021; 26(19):5842. https://doi.org/10.3390/molecules26195842
Chicago/Turabian Styled’Hose, Donatienne, Pauline Isenborghs, Davide Brusa, Bénédicte F. Jordan, and Bernard Gallez. 2021. "The Short-Term Exposure to SDHI Fungicides Boscalid and Bixafen Induces a Mitochondrial Dysfunction in Selective Human Cell Lines" Molecules 26, no. 19: 5842. https://doi.org/10.3390/molecules26195842
APA Styled’Hose, D., Isenborghs, P., Brusa, D., Jordan, B. F., & Gallez, B. (2021). The Short-Term Exposure to SDHI Fungicides Boscalid and Bixafen Induces a Mitochondrial Dysfunction in Selective Human Cell Lines. Molecules, 26(19), 5842. https://doi.org/10.3390/molecules26195842