
Cinnamomum cassia and Syzygium aromaticum Essential Oils Reduce the Colonization of Salmonella Typhimurium in an In Vivo Infection Model Using Caenorhabditis elegans


Abstract
:1. Introduction
2. Results and Discussion
2.1. EOs Composition
2.2. Assessment of Toxic Concentrations
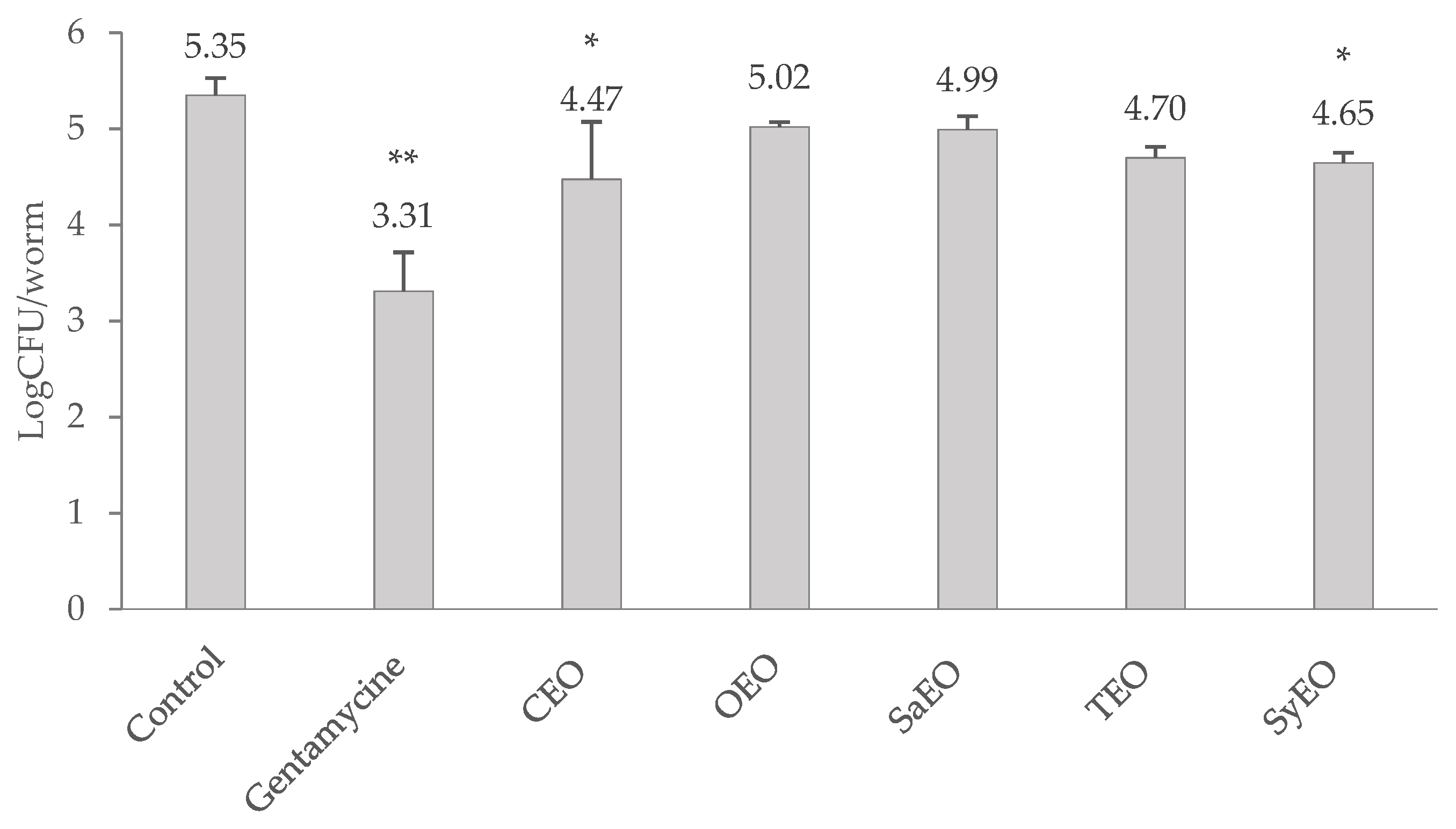
2.3. Effect of EOs on Bacterial Colonization
2.4. Antimicrobial Activities of EOs
2.5. Effects of EOs on Bacterial Biofilm Formation
2.6. Effects of EOs on Swimming Motility
3. Materials and Methods
3.1. Essential Oils
3.2. Bacterial Strains and Maintenance
3.3. Caenorhabditis Elegans Strain and Maintenance
3.4. Evaluation of Toxic Concentrations
3.5. Colonization Assay
3.6. MIC and MBC Determination
3.7. Inhibition of Biofilm Formation
3.8. Swimming Motility Assay
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bajpai, V.K.; Baek, K.H.; Kang, S.C. Control of Salmonella in foods by using essential oils: A review. Food Res. Int. 2012, 45, 722–734. [Google Scholar] [CrossRef]
- Merino, L.; Procura, F.; Trejo, F.M.; Bueno, D.J.; Golowczyc, M.A. Biofilm formation by Salmonella sp. in the poultry industry: Detection, control and eradication strategies. Food Res. Int. 2019, 119, 530–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piovezan, M.; Sayuri Uchida, N.; Fiori da Silva, A.; Grespan, R.; Regina Santos, P.; Leite Silva, E.; Kenji Nakamura Cuman, R.; Machinski Junior, M.; Martha Graton Mikcha, J. Effect of cinnamon essential oil and cinnamaldehyde on Salmonella Saintpaul biofilm on a stainless steel surface. J. Gen. Appl. Microbiol. 2014, 60, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Trampel, D.W.; Holder, T.G.; Gast, R.K. Integrated farm management to prevent Salmonella Enteritidis contamination of eggs. J. Appl. Poult. Res. 2014, 23, 353–365. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Shahid, M.; Ashraf, M.; Przybylski, R. Chemical composition, and antioxidant and antimicrobial activities of essential oil of spearmint (Mentha spicata L.) from pakistan. J. Essent. Oil Res. 2010, 22, 78–84. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cássia Da Silveira E Sá, R.; Andrade, L.N.; De Sousa, D.P. A review on anti-inflammatory activity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef] [PubMed]
- Astani, A.; Reichling, J.; Schnitzler, P. Screening for antiviral activities of isolated compounds from essential oils. Evidence-based Complement. Altern. Med. 2011, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilau, M.R.; Alves, S.H.; Weiblen, R.; Arenhart, S.; Cueto, A.P.; Lovato, L.T. Antiviral activity of the Lippia graveolens (Mexican oregano) essential oil and its main compound carvacrol against human and animal viruses. Braz. J. Microbiol. 2011, 42, 1616–1624. [Google Scholar] [CrossRef] [Green Version]
- Du, E.; Gan, L.; Li, Z.; Wang, W.; Liu, D.; Guo, Y. In vitro antibacterial activity of thymol and carvacrol and their effects on broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2015, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Ballester-Costa, C.; Sendra, E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical composition and in vitro antibacterial properties of essential oils of four Thymus species from organic growth. Ind. Crop. Prod. 2013, 50, 304–311. [Google Scholar] [CrossRef]
- Zhou, F.; Ji, B.; Zhang, H.; Jiang, H.; Yang, Z.; Li, J.; Li, J.; Yan, W. The antibacterial effect of cinnamaldehyde, thymol, carvacrol and their combinations against the foodborne pathogen Salmonella typhimurium. J. Food Saf. 2007, 27, 124–133. [Google Scholar] [CrossRef]
- Soni, K.A.; Oladunjoye, A.; Nannapaneni, R.; Schilling, M.W.; Silva, J.L.; Mikel, B.; Bailey, R.H. Inhibition and inactivation of Salmonella typhimurium biofilms from polystyrene and stainless steel surfaces by essential oils and phenolic constituent carvacrol. J. Food Prot. 2013, 76, 205–212. [Google Scholar] [CrossRef]
- Ikeda, T.; Yasui, C.; Hoshino, K.; Arikawa, K.; Nishikawa, Y. Influence of lactic acid bacteria on longevity of Caenorhabditis elegans and host defense against Salmonella enterica serovar Enteritidis. Appl. Environ. Microbiol. 2007, 73, 6404–6409. [Google Scholar] [CrossRef] [Green Version]
- Moy, T.I.; Ball, A.R.; Anklesaria, Z.; Casadei, G.; Lewis, K.; Ausubel, F.M. Identification of novel antimicrobials using a live-animal infection model. Proc. Natl. Acad. Sci. USA 2006, 103, 10414–10419. [Google Scholar] [CrossRef] [Green Version]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The worm has turned-Microbial virulence modeled in Caenorhabditis elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar] [CrossRef]
- Powell, J.R.; Ausubel, F.M. Models of Caenorhabditis elegans infection by bacterial and fungal pathogens. Methods Mol. Biol. 2008, 415, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, P.S.; Rai, R.V. Inhibition of quorum-sensing-controlled virulence factors of Pseudomonas aeruginosa by Murraya koenigii essential oil: A study in a Caenorhabditis elegans infectious model. J. Med. Microbiol. 2016, 65, 1528–1535. [Google Scholar] [CrossRef]
- Ooi, S.K.; Lim, T.Y.; Lee, S.H.; Nathan, S. Burkholderia pseudomallei kills Caenorhabditis elegans through virulence mechanisms distinct from intestinal lumen colonization. Virulence 2012, 3, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.; Eng, S.-A.; Lim, M.-P.; Nathan, S. Beyond Traditional Antimicrobials: A Caenorhabditis elegans Model for Discovery of Novel Anti-infectives. Front. Microbiol. 2016, 7, 1956. [Google Scholar] [CrossRef]
- Peterson, N.D.; Pukkila-Worley, R. Caenorhabditis elegans in high-throughput screens for anti-infective compounds. Curr. Opin. Immunol. 2018, 54, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Anderson, Q.L.; Revtovich, A.V.; Kirienko, N.V. A High-throughput, High-content, Liquid-based C. elegans Pathosystem. J. Vis. Exp. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.W.; Ausubel, F.M. Caenorhabditis elegans: A model genetic host to study Pseudomonas aeruginosa pathogenesis. Curr. Opin. Microbiol. 2000, 3, 29–34. [Google Scholar] [CrossRef]
- Labrousse, A.; Chauvet, S.; Couillault, C.; Léopold Kurz, C.; Ewbank, J.J. Caenorhabditis elegans is a model host for Salmonella typhimurium. Curr. Biol. 2000, 10, 1543–1545. [Google Scholar] [CrossRef] [Green Version]
- Sem, X.H.; Rhen, M. Pathogenicity of Salmonella enterica in Caenorhabditis elegans Relies on Disseminated Oxidative Stress in the Infected Host. PLoS ONE 2012, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Desai, S.K.; Padmanabhan, A.; Harshe, S.; Zaidel-Bar, R.; Kenney, L.J. Salmonella biofilms program innate immunity for persistence in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2019, 116, 12462–12467. [Google Scholar] [CrossRef] [Green Version]
- Kulshreshtha, G.; Borza, T.; Rathgeber, B.; Stratton, G.S.; Thomas, N.A.; Critchley, A.; Hafting, J.; Prithiviraj, B. Red Seaweeds Sarcodiotheca gaudichaudii and Chondrus crispus down Regulate Virulence Factors of Salmonella Enteritidis and Induce Immune Responses in Caenorhabditis elegans. Front. Microbiol. 2016, 7, 421. [Google Scholar] [CrossRef]
- Horstmann, J.A.; Zschieschang, E.; Truschel, T.; de Diego, J.; Lunelli, M.; Rohde, M.; May, T.; Strowig, T.; Stradal, T.; Kolbe, M.; et al. Flagellin phase-dependent swimming on epithelial cell surfaces contributes to productive Salmonella gut colonisation. Cell. Microbiol. 2017, 19, e12739. [Google Scholar] [CrossRef] [Green Version]
- Ooi, L.S.M.; Li, Y.; Kam, S.-L.; Wang, H.; Wong, E.Y.L.; Ooi, V.E.C. Antimicrobial Activities of Cinnamon Oil and Cinnamaldehyde from the Chinese Medicinal Herb Cinnamomum cassia Blume. Am. J. Chin. Med. 2006, 34, 511–522. [Google Scholar] [CrossRef]
- Fratini, F.; Casella, S.; Leonardi, M.; Pisseri, F.; Ebani, V.V.; Pistelli, L.; Pistelli, L. Antibacterial activity of essential oils, their blends and mixtures of their main constituents against some strains supporting livestock mastitis. Fitoterapia 2014, 96, 1–7. [Google Scholar] [CrossRef]
- Alinkina, E.S.; Misharina, T.A.; Fatkullina, L.D. Antiradical properties of oregano, thyme, and savory essential oils. Appl. Biochem. Microbiol. 2012, 49, 73–78. [Google Scholar] [CrossRef]
- Murbach Teles Andrade, B.F.; Nunes Barbosa, L.; da Silva Probst, I.; Fernandes Júnior, A. Antimicrobial activity of essential oils. J. Essent. Oil Res. 2014, 26, 34–40. [Google Scholar] [CrossRef]
- Woods, K.E.; Chhetri, B.K.; Jones, C.D.; Goel, N.; Setzer, W.N. Bioactivities and compositions of Betula nigra essential oils. J. Med. Act. Plants 2013, 2, 1–9. [Google Scholar] [CrossRef]
- Satyal, P.; Paudel, P.; Poudel, A.; Dosoky, N.S.; Pokharel, K.K.; Setzer, W.N. Bioactivities and compositional analyses of Cinnamomum essential oils from Nepal: C. camphora, C. tamala, and C. glaucescens. Nat. Prod. Commun. 2013, 8, 1777–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosoky, N.; Satyal, P.; Gautam, T.; Setzer, W. Composition and Biological Activities of Murraya paniculata (L.) Jack Essential Oil from Nepal. Medicines 2016, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Husain, F.M.; Ahmad, I.; Asif, M.; Tahseen, Q. Influence of clove oil on certain quorum-sensing-regulated functions and biofilm of Pseudomonas aeruginosa and Aeromonas hydrophila. J. Biosci. 2013, 38, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ahmad, I.; Khan, M.S.; Ahmad, E.; Tahseen, Q.; Khan, M.S.; Alshabib, N.A. Sub-MICs of Mentha piperita essential oil and menthol inhibits AHL mediated quorum sensing and biofilm of Gram-negative bacteria. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Faleiro, M.L. The mode of antibacterial action of essential oils. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; Volume 3, pp. 1143–1156, ISBN-13: 978-84-939843-1-1. [Google Scholar]
- Kalai Chelvam, K.; Chai, L.C.; Thong, K.L. Variations in motility and biofilm formation of Salmonella enterica serovar Typhi. Gut Pathog. 2014, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 100, 296–305. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Saucier, L.; Lacroix, M. Inhibitory effects of selected plant essential oils on the growth of four pathogenic bacteria: E. coli O157:H7, Salmonella Typhimurium, Staphylococcus aureus and Listeria monocytogenes. Food Control 2007, 18, 414–420. [Google Scholar] [CrossRef]
- Niu, C.; Gilbert, E.S. Colorimetric Method for Identifying Plant Essential Oil Components That Affect Biofilm Formation and Structure. Appl. Environ. Microbiol. 2004, 70, 6951–6956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollanoor-Johny, A.; Mattson, T.; Baskaran, S.A.; Amalaradjou, M.A.; Babapoor, S.; March, B.; Valipe, S.; Darre, M.; Hoagland, T.; Schreiber, D.; et al. Reduction of Salmonella enterica serovar enteritidis colonization in 20-day-old broiler chickens by the plant-derived compounds trans-cinnamaldehyde and eugenol. Appl. Environ. Microbiol. 2012, 78, 2981–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta-de-la-Riva, M.; Fontrodona, L.; Villanueva, A.; Cerón, J. Basic Caenorhabditis elegans methods: Synchronization and observation. J. Vis. Exp. 2012, 64, e4019. [Google Scholar] [CrossRef] [Green Version]
- Conery, A.L.; Larkins-Ford, J.; Ausubel, F.M.; Kirienko, N.V. High-Throughput Screening for Novel Anti-Infectives Using a C. elegans Pathogenesis Model. Curr. Protoc. Chem. Biol. 2014, 6, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Konga, C.; Rahmanb, N.A.; Nathana, S. Identification of potential anti-infectives against staphylococcus aureus using a Caenorhabditis elegans infection model. AIP Conf. Proc. 2014, 1614, 513–516. [Google Scholar] [CrossRef]
- Lang, M.; Ferron, P.-J.; Bursztyka, J.; Montjarret, A.; Duteil, E.; Bazire, A.; Bedoux, G. Evaluation of immunomodulatory activities of essential oils by high content analysis. J. Biotechnol. 2019, 303, 65–71. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
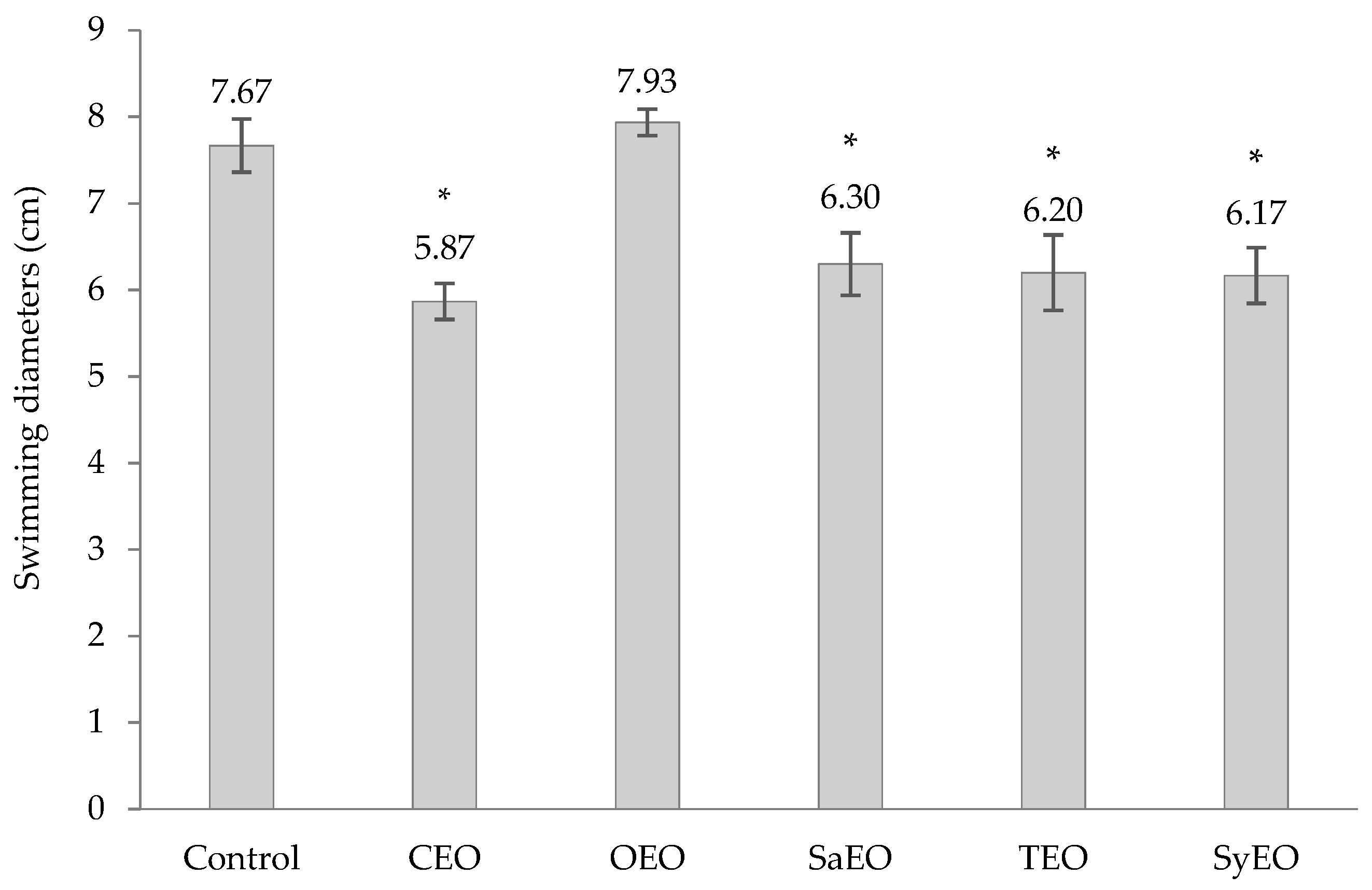{kind=link}
| Cinnamomum cassia | Origanum vulgare | Satureja hortensis | Thymus vulgaris | Syzygium aromaticum | |
|---|---|---|---|---|---|
| (CEO) | (OEO) | (SaEO) | (TEO) | (SyEO) | |
| Eugenol | 0.02 | 80.67 | |||
| Cinnamaldehyde (cis + trans) | 79.98 | ||||
| Carvacrol | 70.72 | 47.37 | 2.13 | ||
| Thymol | 2.12 | 1.61 | 40.20 | ||
| p-Cymene | 3.70 | 11.59 | 22.04 | ||
| γ-Terpinene | 4.97 | 27.36 | 11.46 | ||
| β-Caryophyllene | 3.58 | 1.58 | 3.55 | 9.6 | |
| o-Methoxycinnamaldehyde | 8.00 | ||||
| Eugenyl acetate | 6.0 | ||||
| Linalool | 3.00 | 1.04 | 3.66 | ||
| Cinnamyl acetate | 2.52 | ||||
| Myrcene | 2.00 | 2.14 | |||
| α-Terpinene | 0.58 | 2.09 | 1.80 | ||
| Coumarin | 1.56 | ||||
| D-Limonene | 0.04 | 0.72 | 0.48 | ||
| Eucalyptol | 1.41 | ||||
| (D,L)-Borneol | 1.20 | 1.34 | |||
| α-Pinene | 1.18 | ||||
| β-Pinene | 1.15 | ||||
| 4-Terpinenol | 1.08 | ||||
| Benzaldehyde | 0.83 | ||||
| Camphor | 0.83 | ||||
| α-Terpineol | 0.76 | 0.17 | |||
| 4-Carvomenthenol | 0.75 | ||||
| Camphene | 0.70 | ||||
| Phenylethyl alcohol | 0.70 | ||||
| trans-Sabinene hydrate | 0.50 | ||||
| Styrene | 0.21 | ||||
| Salicylaldehyde | 0.20 | ||||
| Terpinolene | 0.15 | ||||
| Cinnamic acid | 0.11 | ||||
| Cinnamyl alcohol | 0.11 | ||||
| Benzyl Benzoate | 0.06 | ||||
| Furfural | 0.06 |
| CEO | OEO | SaEO | TEO | SyEO | ||
|---|---|---|---|---|---|---|
| C. elegans | LD10 | 43.7 ± 5.4 a | 52.8 ± 4.2 ab | 71.6 ± 18.4 bc | 91.8 ± 1.1 c | 156.8 ± 16.0 d |
| LD50 | 74.5 ± 9.6 a | 113.9 ± 8.8 b | 136.9 ± 20.6 bc | 186.6 ± 33.2 c | 271.6 ± 14.9 d | |
| LD90 | 127.3 ± 18.2 a | 263.4 ± 27.7 b | 265.7 ± 33.0 b | 305.7 ± 65.8 b | 471.2 ± 3.3 c | |
| S. Typhimurium | MIC MBC | - 312.5 ± 0.0 | - 312.5 ± 0.0 | - 312.5 ± 0.0 | - 312.5 ± 0.0 | - 625.0 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, M.; Montjarret, A.; Duteil, E.; Bedoux, G. Cinnamomum cassia and Syzygium aromaticum Essential Oils Reduce the Colonization of Salmonella Typhimurium in an In Vivo Infection Model Using Caenorhabditis elegans. Molecules 2021, 26, 5598. https://doi.org/10.3390/molecules26185598
Lang M, Montjarret A, Duteil E, Bedoux G. Cinnamomum cassia and Syzygium aromaticum Essential Oils Reduce the Colonization of Salmonella Typhimurium in an In Vivo Infection Model Using Caenorhabditis elegans. Molecules. 2021; 26(18):5598. https://doi.org/10.3390/molecules26185598
Chicago/Turabian StyleLang, Marie, Aude Montjarret, Emmanuel Duteil, and Gilles Bedoux. 2021. "Cinnamomum cassia and Syzygium aromaticum Essential Oils Reduce the Colonization of Salmonella Typhimurium in an In Vivo Infection Model Using Caenorhabditis elegans" Molecules 26, no. 18: 5598. https://doi.org/10.3390/molecules26185598
APA StyleLang, M., Montjarret, A., Duteil, E., & Bedoux, G. (2021). Cinnamomum cassia and Syzygium aromaticum Essential Oils Reduce the Colonization of Salmonella Typhimurium in an In Vivo Infection Model Using Caenorhabditis elegans. Molecules, 26(18), 5598. https://doi.org/10.3390/molecules26185598