
A Novel Approach to Unraveling the Apoptotic Potential of Rutin (Bioflavonoid) via Targeting Jab1 in Cervical Cancer Cells

 ,
,  ,
,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Requirements
2.2. MTT Assay
2.3. Extracellular Lactate Dehydrogenase (LDH) Activity Analysis in Rutin-Treated SiHa Cells
2.4. Investigation of Nuclear Morphology in Rutin-Treated Cells
2.5. Quantification of Apoptosis by Annexin V-FITC/PI Assay
2.6. Investigation of Caspases Activities in Rutin-Treated SiHa Cells
2.7. Investigation of the Effect of Caspase (Caspase-3 and Caspase-9) Inhibitors
2.8. Investigation of MMP (Mitochondrial Membrane Potential) in Rutin-Treated SiHa Cells
2.9. Measurement of Cytochrome C Level
2.10. Measurement of Intracellular ROS (Reactive Oxygen Species) Level
2.11. Investigation of ROS Inhibitor, NAC (N-Acetyl-L-Cysteine) Efficacy
2.12. Real-Time PCR Analysis
2.13. Statistical Analyses
3. Results
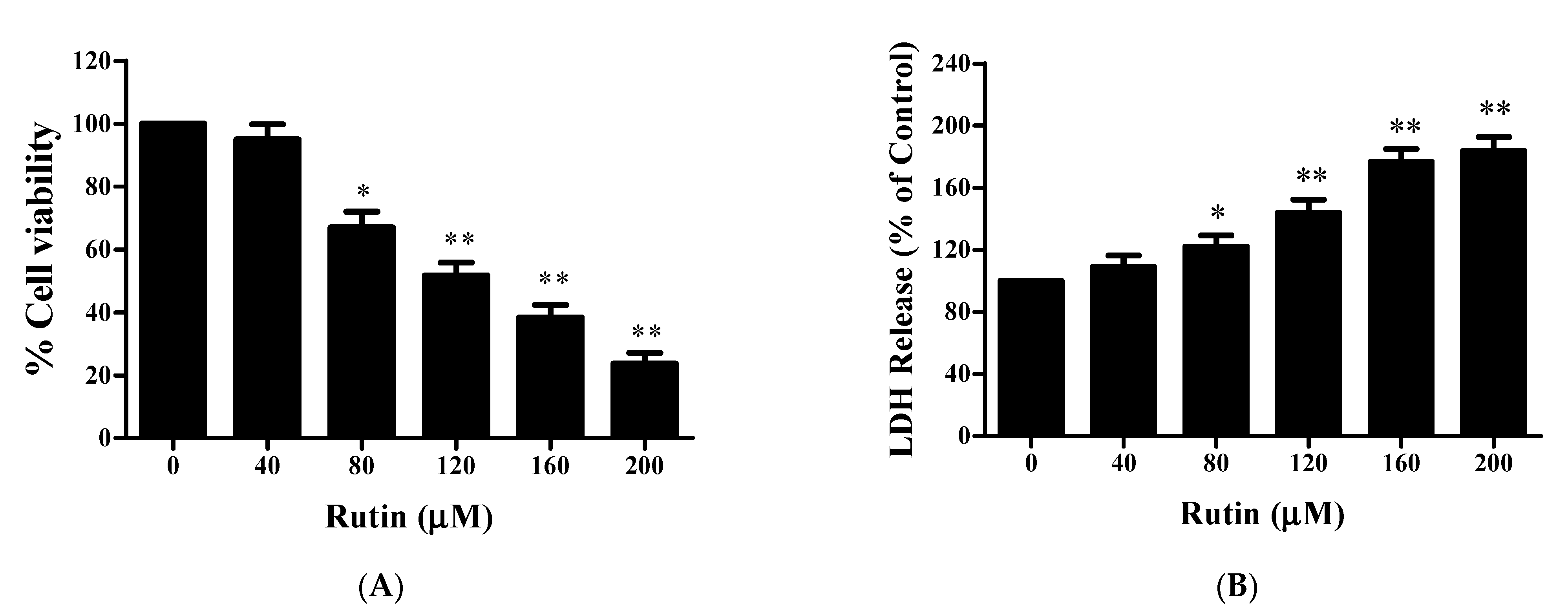
3.1. Rutin-Induced Antiproliferative Activity in SiHa Cervical Cancer Cells
3.2. Rutin Inhibits the Growth of SiHa Cells via Downregulation of Jab1 and Upregulation of p27
3.3. Rutin Induces Apoptosis in SiHa Cancer Cells through Upregulation of p53, Bax, and Downregulation of Bcl-2
3.4. Mitochondrial-Mediated Apoptosis Induction in Rutin-Treated SiHa Cancer Cells
3.5. Caspase-Mediated Apoptosis Induction in Rutin-Treated SiHa Cancer Cells
3.6. Effect of Rutin on ROS Generation in Apoptosis Induction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Moga, M.A.; Dimienescu, O.G.; Arvatescu, C.A.; Mironescu, A.; Dracea, L.; Ples, L. The Role of Natural Polyphenols in the Prevention and Treatment of Cervical Cancer-An Overview. Molecules 2016, 21, 1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of incidence and mortality of cervical cancer in 2018: A worldwide analysis. Lancet Glob. Health 2020, 8, e191–e203. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Khan, F. Jab1 Inhibition by Methanolic Extract of Moringa Oleifera Leaves in Cervical Cancer Cells: A Potent Targeted Therapeutic Approach. Nutr. Cancer 2020, 1–9. [Google Scholar] [CrossRef]
- Maurya, P.; Pandey, P.; Khan, F.; Mishra, R.; Chaudhary, R.; Singh, S.K. Study to Elucidate the Inhibitory Potential of Selected Flavonoids against Jab1 in Cervical Cancer. BioInterface Res. Appl. Chem. 2021, 12, 1290–1303. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Javanmardi, S.; Moradi-Ozarlou, M.; Mohammadinejad, R.; Farkhondeh, T.; Samarghandian, S.; Garg, M. Natural products and phytochemical nanoformulations targeting mitochondria in oncotherapy: An updated review on resveratrol. Biosci. Rep. 2020, 40, BSR20200257. [Google Scholar] [CrossRef] [Green Version]
- Liontos, M.; Kyriazoglou, A.; Dimitriadis, I.; Dimopoulos, M.A.; Bamias, A. Systemic therapy in cervical cancer: 30 years in review. Crit. Rev. Oncol. Hematol. 2019, 137, 9–17. [Google Scholar] [CrossRef]
- Kuo, C.-Y.; Schelz, Z.; Tóth, B.; Vasas, A.; Ocsovszki, I.; Chang, F.-R.; Hohmann, J.; Zupkó, I.; Wang, H.-C. Investigation of natural phenanthrenes and the antiproliferative potential of juncusol in cervical cancer cell lines. Phytomedicine 2019, 58, 152770. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.R.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Banzato, T.P.; Gubiani, J.R.; Bernardi, D.I.; Nogueira, C.R.; Monteiro, A.F.; Juliano, F.F.; De Alencar, S.M.; Pilli, R.A.; de Lima, C.; Longato, G.; et al. Antiproliferative Flavanoid Dimers Isolated from Brazilian Red Propolis. J. Nat. Prod. 2020, 83, 1784–1793. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F.; Mazumder, A.; Rana, A.K.; Srivastava, Y. Inhibitory Potential of Dietary Phytocompounds of Nigella sativa against Key Targets of Novel Coronavirus (COVID-19). Indian J. Pharm. Educ. Res. 2021, 55. [Google Scholar] [CrossRef]
- Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; Gondal, T.A.; Mubarak, M.S. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G.; et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef]
- Uysal, S.; Aktumsek, A.; Picot-Allain, C.M.; Unuvar, H.; Mollica, A.; Georgiev, M.I.; Zengin, G.; Mahomoodally, M.F. Biological, chemical and in silico fingerprints of Dianthus calocephalus Boiss.: A novel source for rutin. Food Chem. Toxicol. 2018, 113, 179–186. [Google Scholar] [CrossRef]
- Gullón, B.; Lú-Chau, T.A.; Moreira, M.T.; Lema, J.M.; Eibes, G. Rutin: A review on extraction, identification and purification methods, biological activities and approaches to enhance its bioavailability. Trends Food Sci. Technol. 2017, 67, 220–235. [Google Scholar] [CrossRef]
- Farha, A.K.; Gan, R.-Y.; Li, H.-B.; Wu, D.-T.; Atanasov, A.G.; Gul, K.; Zhang, J.-R.; Yang, Q.-Q.; Corke, H. The anticancer potential of the dietary polyphenol rutin: Current status, challenges, and perspectives. Crit. Rev. Food Sci. Nutr. 2020, 1–28. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budzynska, B.; Faggio, C.; Kruk-Slomka, M.; Samec, D.; Nabavi, S.F.; Sureda, A.; Devi, K.P.; Nabavi, S.M. Rutin as Neuroprotective Agent: Rom Bench to Bedside. Curr. Med. Chem. 2019, 26, 5152–5164. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Rajput, V.S.; Nagpal, P.; Kukrety, H.; Grover, S.; Grover, A. Dual inhibition of SARS-CoV-2 spike and main protease through a repurposed drug, rutin. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef]
- Motamedshariaty, V.S.; Amel Farzad, S.; Nassiri-Asl, M.; Hosseinzadeh, H. Effects of rutin on acrylamide-induced neurotoxicity. Daru 2014, 22, 27. [Google Scholar] [CrossRef] [Green Version]
- Thabet, N.M.; Moustafa, E.M. Protective effect of rutin against brain injury induced by acrylamide or gamma radiation: Role of PI3K/AKT/GSK-3β/NRF-2 signalling pathway. Arch. Physiol. Biochem. 2018, 124, 185–193. [Google Scholar] [CrossRef]
- Pandey, P.; Sayyed, U.; Tiwari, R.K.; Siddiqui, M.H.; Pathak, N.; Bajpai, P. Hesperidin Induces ROS-Mediated Apoptosis along with Cell Cycle Arrest at G2/M Phase in Human Gall Bladder Carcinoma. Nutr. Cancer 2019, 71, 676–687. [Google Scholar] [CrossRef]
- Khan, F.; Pandey, P.; Jha, N.K.; Jafri, A.; Khan, I. Antiproliferative effect of Moringa oleifera methanolic leaf extract by down-regulation of Notch signaling in DU145 prostate cancer cells. Gene Rep. 2020, 19, 100619. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F.; Maurya, P. Targeting Jab1 using hesperidin (dietary phytocompound) for inducing apoptosis in HeLa cervical cancer cells. J. Food Biochem. 2021, 45, e13800. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Claret, F.X.; Shen, Q. The novel Jab1 inhibitor CSN5i-3 suppresses cell proliferation and induces apoptosis in human breast cancer cells. Neoplasma 2019, 66, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, Y.; Zhao, Y.; Shu, Y.; Liu, Z.; Zhou, H.; Wang, H.; Zhang, W. The pivotal oncogenic role of Jab1/CSN5 and its therapeutic implications in human cancer. Gene 2019, 687, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Pan, Y.; Liu, G.; Yang, T.; Jin, Y.; Zhou, F.; Wei, Y. MRPS30-DT Knockdown Inhibits Breast Cancer Progression by Targeting Jab1/Cops5. Front. Oncol. 2019, 9, 1170. [Google Scholar] [CrossRef]
- Pistollato, F.; Giampieri, F.; Battino, M. The use of plant-derived bioactive compounds to target cancer stem cells and modulate tumor microenvironment. Food Chem. Toxicol. 2015, 75, 58–70. [Google Scholar] [CrossRef]
- Pistollato, F.; Bremer-Hoffmann, S.; Basso, G.; Cano, S.S.; Elio, I.; Vergara, M.M.; Giampieri, F.; Battino, M. Targeting Glioblastoma with the Use of Phytocompounds and Nanoparticles. Target. Oncol. 2015, 11, 1–16. [Google Scholar] [CrossRef]
- Anwanwan, D.; Singh, S.K.; Singh, S.; Saikam, V.; Singh, R. Challenges in liver cancer and possible treatment approaches. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188314. [Google Scholar] [CrossRef]
- Pistollato, F.; Iglesias, R.C.; Ruiz, R.; Aparicio, S.; Crespo, J.; Lopez, L.A.D.; Giampieri, F.; Battino, M. The use of natural compounds for the targeting and chemoprevention of ovarian cancer. Cancer Lett. 2017, 411, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, B.A.; Iqbal, J.; Ahmad, R.; Bibi, S.; Mahmood, T.; Kanwal, S.; Bashir, S.; Gul, F.; Hameed, S. Potential phytochemicals in the prevention and treatment of esophagus cancer: A green therapeutic approach. Pharmacol. Rep. 2019, 71, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Deepika, M.S.; Thangam, R.; Sheena, T.S.; Sasirekha, R.; Sivasubramanian, S.; Babu, M.D.; Jeganathan, K.; Thirumurugan, R. A novel rutin-fucoidan complex based phytotherapy for cervical cancer through achieving enhanced bioavailability and cancer cell apoptosis. Biomed. Pharmacother. 2019, 109, 1181–1195. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Pandey, P.; Upadhyay, T.; Jafri, A.; Jha, N.K.; Mishra, R.; Singh, V. Anti-Cancerous Effect of Rutin Against HPV-C33A Cervical Cancer Cells via G0/G1 Cell Cycle Arrest and Apoptotic Induction. Endocr. Metab. Immune Disord.-Drug Targets 2020, 20, 409–418. [Google Scholar] [CrossRef]
- Li, Q.; Ren, L.; Zhang, Y.; Gu, Z.; Tan, Q.; Zhang, T.; Qin, M.; Chen, S. P38 Signal Transduction Pathway Has More Cofactors on Apoptosis of SGC-7901 Gastric Cancer Cells Induced by Combination of Rutin and Oxaliplatin. BioMed Res. Int. 2019, 2019, 6407210. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhang, Q.; Tian, L.; Wang, X.; Fan, X.; Zhang, H.; Claret, F.X.; Yang, H. Jab1/CSN5 Negatively Regulates p27 and Plays a Role in the Pathogenesis of Nasopharyngeal Carcinoma. Cancer Res. 2012, 72, 1890–1900. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Bajpai, P.; Siddiqui, M.H.; Sayyed, U.; Tiwari, R.; Shekh, R.; Mishra, K.; Kapoor, V. Elucidation of the Chemopreventive Role of Stigmasterol Against Jab1 in Gall Bladder Carcinoma. Endocr. Metab. Immune Disord.—Drug Targets 2019, 19, 826–837. [Google Scholar] [CrossRef]
- Pandey, P.; Siddiqui, M.H.; Behari, A.; Kapoor, V.K.; Mishra, K.; Sayyed, U.; Tiwari, R.K.; Shekh, R.; Bajpai, P. Jab1-siRNA Induces Cell Growth Inhibition and Cell Cycle Arrest in Gall Bladder Cancer Cells via Targeting Jab1 Signalosome. Anti-Cancer Agents Med. Chem. 2020, 19, 2019–2033. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, D.; Liu, G.; Pan, Y. Jab1/Cops5: A promising target for cancer diagnosis and therapy. Int. J. Clin. Oncol. 2021, 26, 1159–1169. [Google Scholar] [CrossRef]
- Samsa, W.E.; Mamidi, M.K.; Bashur, L.A.; Elliott, R.; Miron, A.; Chen, Y.; Lee, B.; Greenfield, E.M.; Chan, R.; Danielpour, D.; et al. The crucial p53-dependent oncogenic role of JAB1 in osteosarcoma in vivo. Oncogene 2020, 39, 4581–4591. [Google Scholar] [CrossRef]
- Sang, M.M.; Du, W.Q.; Zhang, R.Y.; Zheng, J.N.; Pei, D.S. Suppression of CSN5 promotes the apoptosis of gastric cancer cells through regulating p53-related apoptotic pathways. Bioorg. Med. Chem. Lett. 2015, 25, 2897–2901. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Claret, F.X.; Zhou, F.; Pan, Y. Jab1/COPS5 as a Novel Biomarker for Diagnosis, Prognosis, Therapy Prediction and Therapeutic Tools for Human Cancer. Front Pharmacol. 2018, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zheng, J.; Nussinov, R.; Ma, B. Release of Cytochrome C from Bax Pores at the Mitochondrial Membrane. Sci. Rep. 2017, 7, 2635. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.U.; Soujanya, M.; Mishra, R.K. Deterioration of nuclear morphology and architecture: A hallmark of senescence and aging. Ageing Res. Rev. 2021, 67, 101264. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadeh, H.; Keikhaei, B.; Mottaghi, S. A review of molecular mechanisms involved in anticancer and antiangiogenic effects of natural polyphenolic compounds. Phytother. Res. 2019, 33, 2002–2014. [Google Scholar] [CrossRef]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Líšková, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Kesavardhana, S.; Malireddi, R.K.S.; Kanneganti, T.D. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [Green Version]
- Zamaraev, A.V.; Kopeina, G.S.; Prokhorova, E.A.; Zhivotovsky, B.; Lavrik, I.N. Post-translational Modification of Caspases: The Other Side of Apoptosis Regulation. Trends Cell Biol. 2017, 27, 322–339. [Google Scholar] [CrossRef]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, Q.; Halim, A.; Song, G. Targeting lipid metabolism of cancer cells: A promising therapeutic strategy for cancer. Cancer Lett. 2017, 401, 39–45. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Bax | AGGGTGGCTGGGAAGGC | TGAGCGAGGCGGTGAGG |
| Bcl-2 | ATCGCTCTGTGGATGACTGAGTAC | AGAGACAGCCAGGAGAAATCAAAC |
| Jab1 | GGCGCCTTTAGGACATACC | CATGAAACTCCCTCGTCCC |
| Caspase-3 | ACCAAAGATCATACATGGAAGCGA | CGAGATGTCATTCCAGTGCT |
| Caspase-9 | TGGTGATGTCGGTGCTCTTG | ACCATGAAATGCAGCGAGGA |
| p27 | TCTGGAACAGCGTGCCATTGATCT | ATTACTGAGGGCCACTTCCACCTT |
| p53 | ACTAAGCGAGCACTGCCCAA | ATGGCGGGAGGTAGACTGAC |
| GAPDH | AAGTTCAACGGCACAGTCAAGG | CATACTCAGCACCAGCATCACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, P.; Khan, F.; Alzahrani, F.A.; Qari, H.A.; Oves, M. A Novel Approach to Unraveling the Apoptotic Potential of Rutin (Bioflavonoid) via Targeting Jab1 in Cervical Cancer Cells. Molecules 2021, 26, 5529. https://doi.org/10.3390/molecules26185529
Pandey P, Khan F, Alzahrani FA, Qari HA, Oves M. A Novel Approach to Unraveling the Apoptotic Potential of Rutin (Bioflavonoid) via Targeting Jab1 in Cervical Cancer Cells. Molecules. 2021; 26(18):5529. https://doi.org/10.3390/molecules26185529
Chicago/Turabian StylePandey, Pratibha, Fahad Khan, Faisal Abdulrahman Alzahrani, Huda A. Qari, and Mohammad Oves. 2021. "A Novel Approach to Unraveling the Apoptotic Potential of Rutin (Bioflavonoid) via Targeting Jab1 in Cervical Cancer Cells" Molecules 26, no. 18: 5529. https://doi.org/10.3390/molecules26185529
APA StylePandey, P., Khan, F., Alzahrani, F. A., Qari, H. A., & Oves, M. (2021). A Novel Approach to Unraveling the Apoptotic Potential of Rutin (Bioflavonoid) via Targeting Jab1 in Cervical Cancer Cells. Molecules, 26(18), 5529. https://doi.org/10.3390/molecules26185529