
Acanthaster planci Inhibits PCSK9 and Lowers Cholesterol Levels in Rats

Abstract
1. Introduction
2. Results
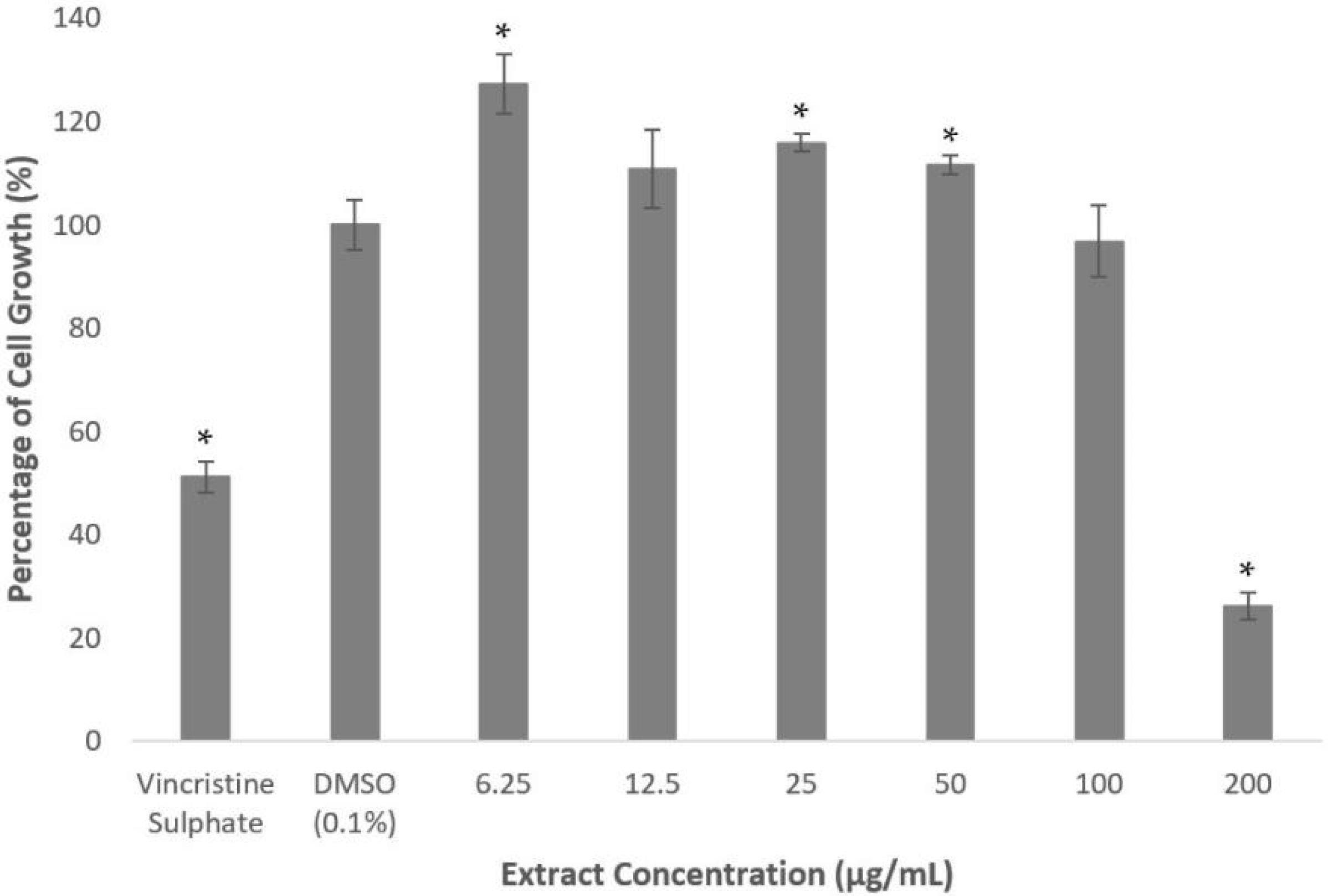
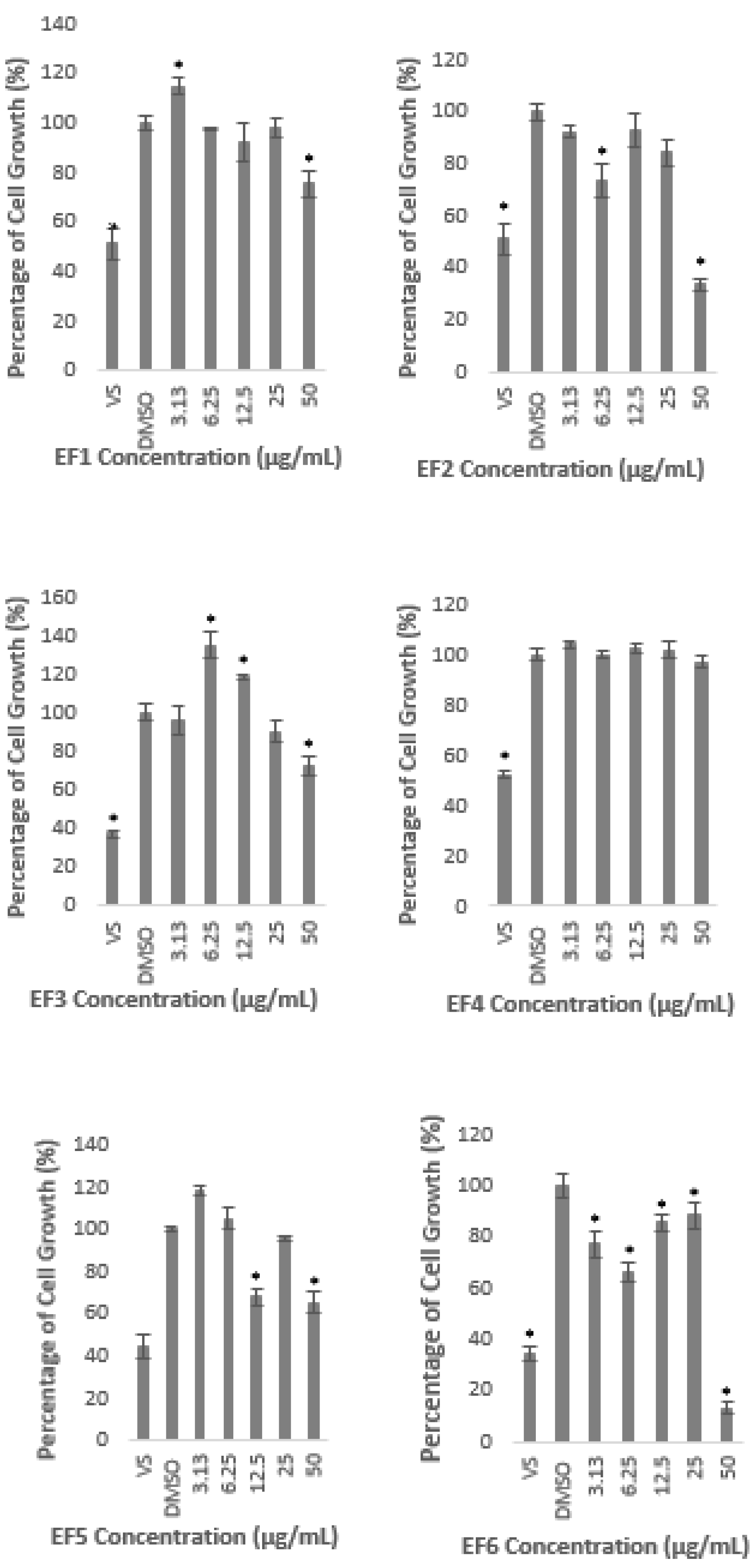
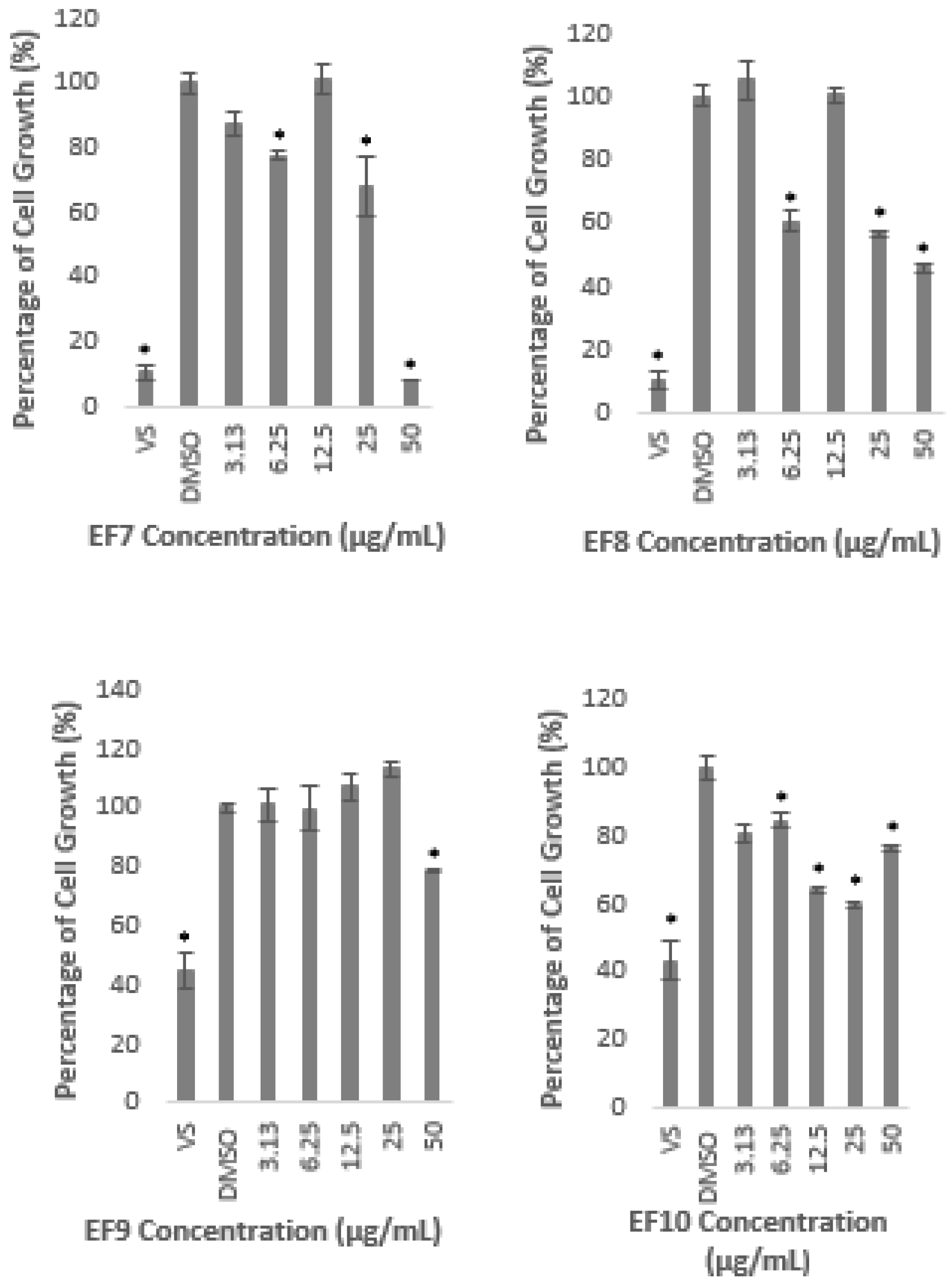
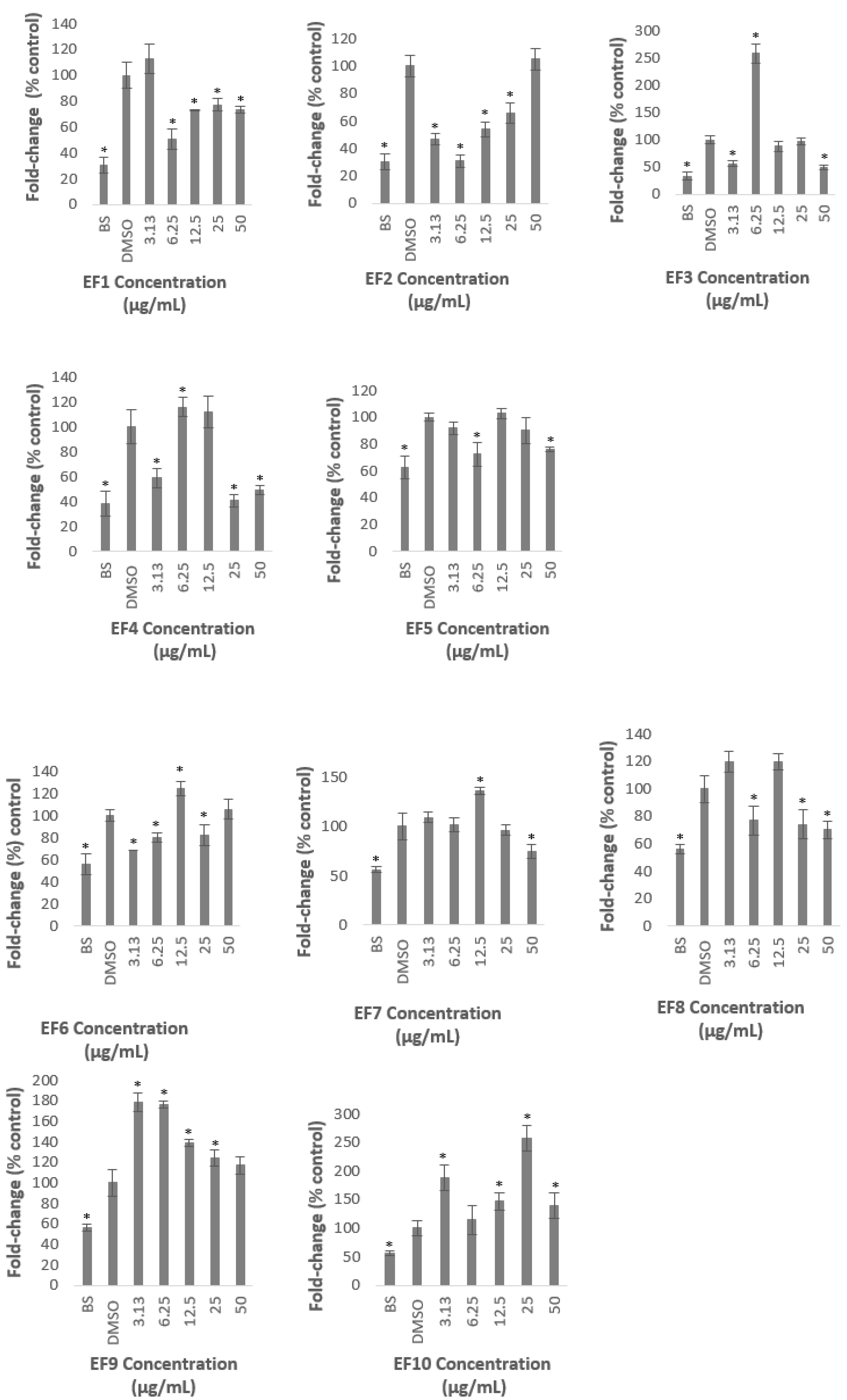
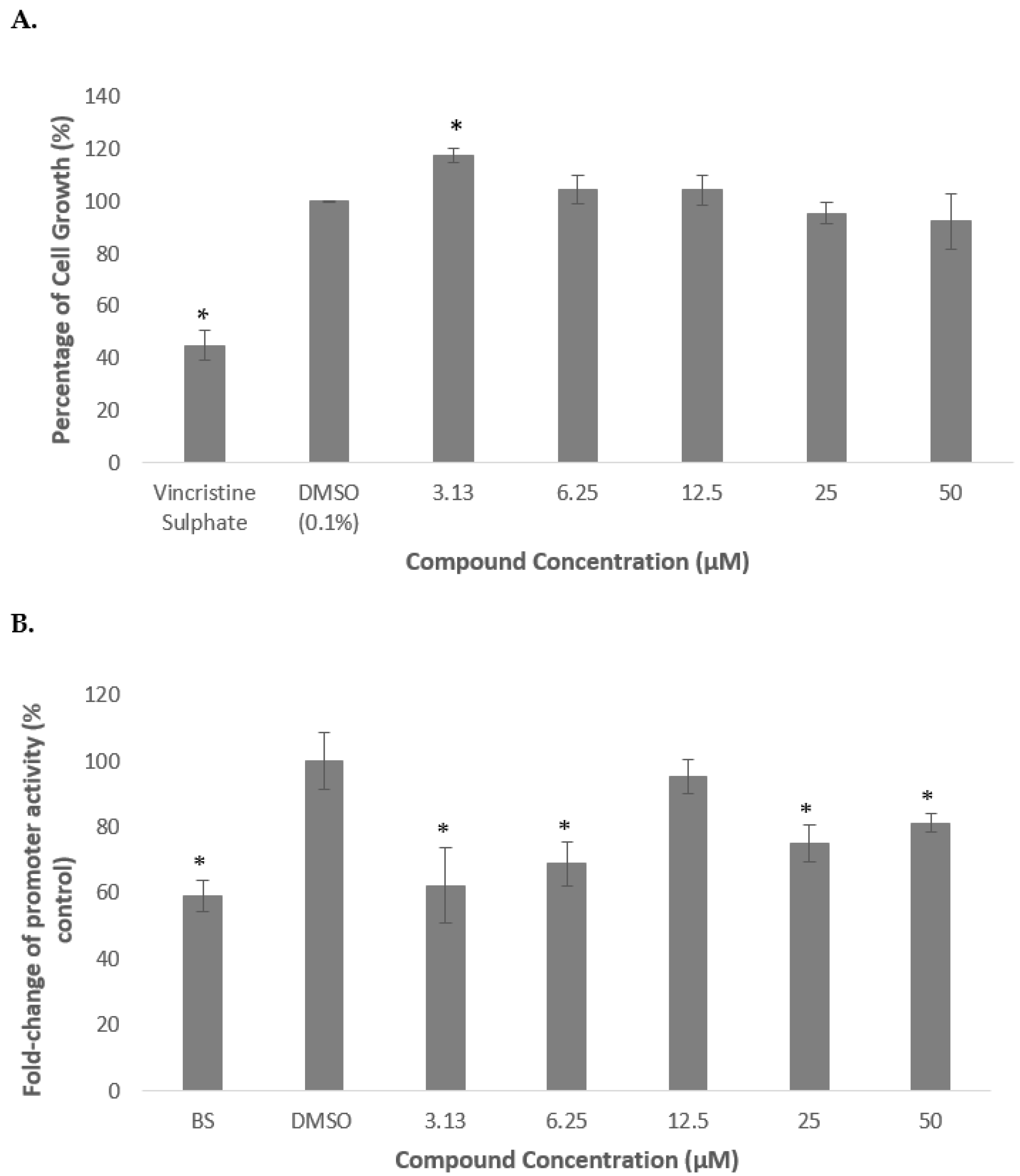
2.1. Methanolic Extract and Fractions of A. planci Do Not Produce Cytotoxicity Effects on HepG2 Cell Line
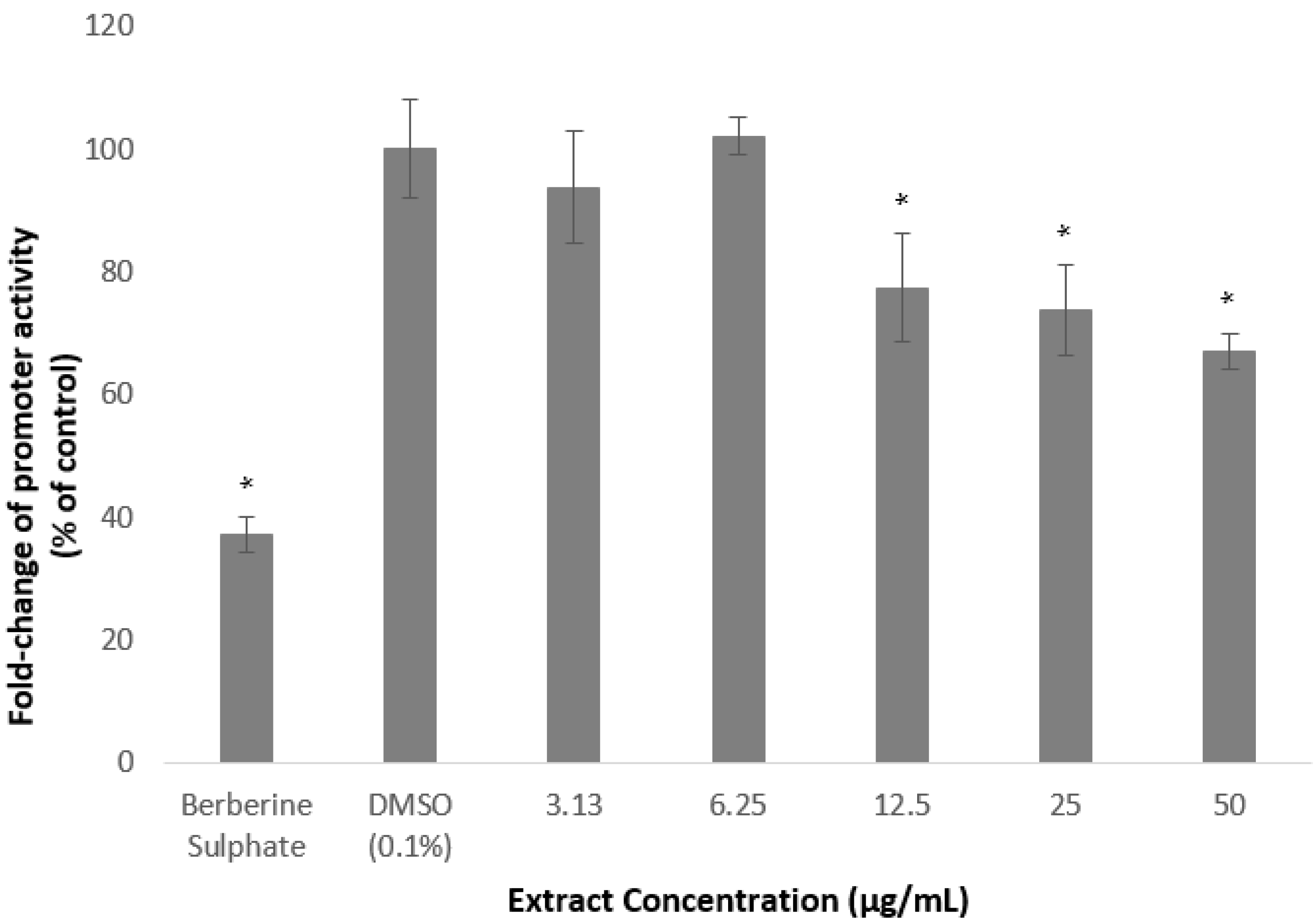
2.2. Methanolic Extract and Fractions of A. planci Reduce the PCSK9 Promoter Activity
2.3. Deoxythymidine, a Compound Isolated from Fraction 1 Reduces PCSK9 Promoter Activity
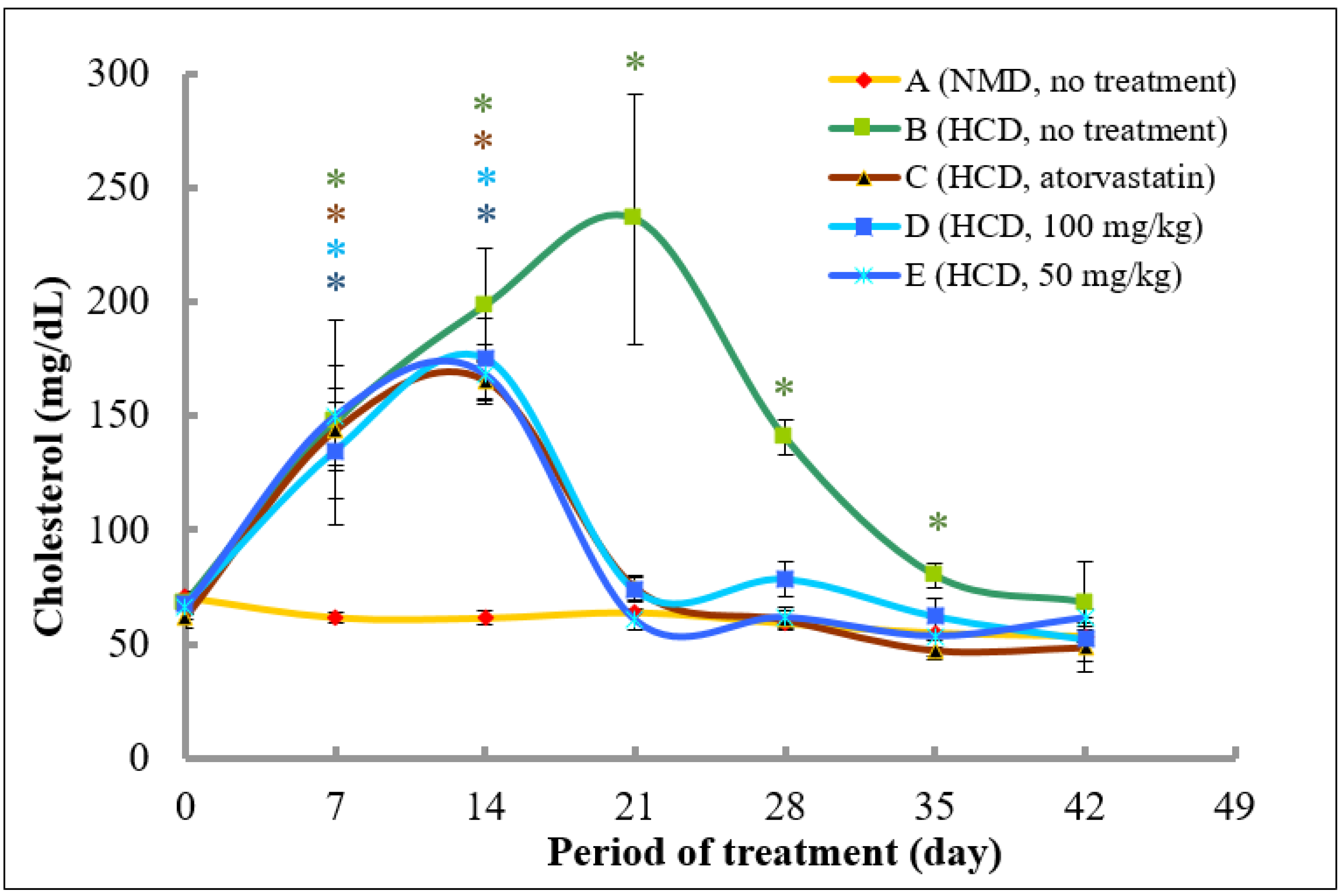
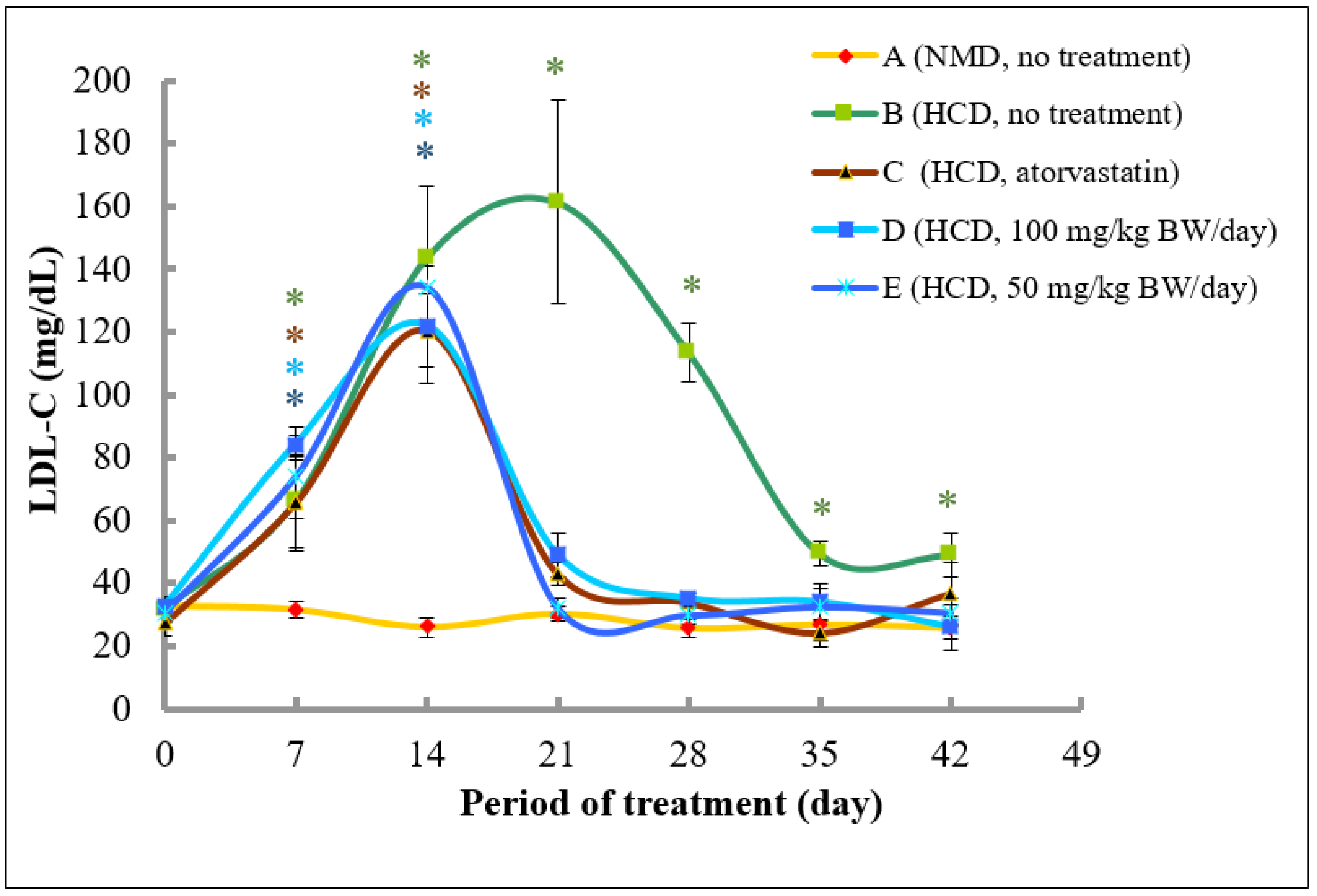
2.4. Methanol Extract of A. planci Significantly Reduces Plasma Total Cholesterols and LDL-C in Rats
2.4.1. Analysis of Plasma Total Cholesterol Levels
2.4.2. Analysis of LDL Cholesterol Levels
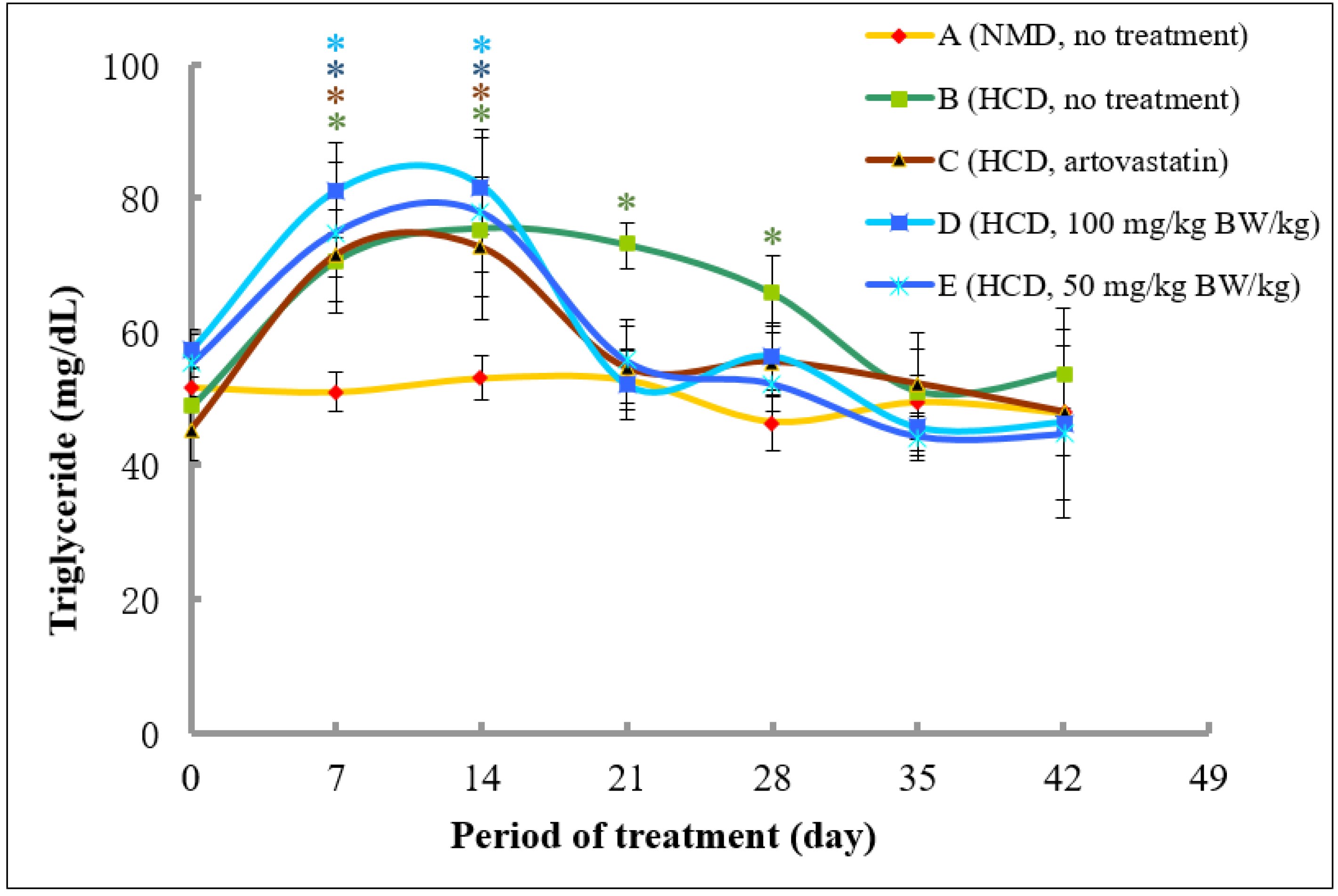
2.4.3. Analysis of Triglycerides Cholesterol Levels
2.5. Methanol Extract of A. Planci Does Not Produce Significant Changes in SGOT and SGPT Levels
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Extraction
4.2. Fractionation and Profiling
4.3. Cell Culture
4.4. Cytotoxicity Assay
4.5. Transient Transfection and Luciferase Assay
4.6. In Vivo Study
4.6.1. Total Cholesterol ASSAY
4.6.2. LDL Cholesterol Assay
4.6.3. Triglycerides Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Stone, N.J.; Robinson, J.G.; Lichtenstein, A.H.; Bairey Merz, C.N.; Blum, C.B.; Eckel, R.H.; Goldberg, A.C.; Gordon, D.; Levy, D.; Lloyd-Jones, D.M.; et al. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults. Circulation 2013, 129, S1–S45. [Google Scholar] [CrossRef]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update From the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Barquera, S.; Pedroza-Tobias, A.; Medina, C.; Hernandez-Barrera, L.; Bibbins-Domingo, K.; Lozano, R.; Moran, E.A. Global overview of the epidemiology of atherosclerotic cardiovascular disease. Arch. Med. Res. 2015, 46, 328–338. [Google Scholar] [CrossRef]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef]
- Libby, P.; Aikawa, M.; Schonbeck, U. Cholesterol and atherosclerosis. Biochim. Biophys. Acta Mol. 2000, 1529, 299–309. [Google Scholar] [CrossRef]
- Virmani, R.; Burke, A.P.; Kolodgie, F.D.; Farb, A. Vulnerable plaque: The pathology of unstable coronary lesions. J. Interv. Cardiol. 2002, 15, 439–446. [Google Scholar] [CrossRef]
- Sirtori, C.R. The pharmacology of statins. Pharmacol. Res. 2014, 88, 3–11. [Google Scholar] [CrossRef]
- Poznyak, A.V.; Zhang, D.; Orekhova, V.; Grechko, A.V.; Wetzker, R.; Orekhov, A.N. A brief overview of currently used atherosclerosis treatment approaches targeting lipid metabolism alterations. Am. J. Cardiovasc. Dis. 2020, 10, 62–71. [Google Scholar] [PubMed]
- Akyea, R.K.; Kai, J.; Qureshi, N.; Iyen, B.; Weng, S.F. Sub-optimal cholesterol response to initiation of statins and future risk of cardiovascular disease. Heart 2019, 105, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Seidah, N.G.; Benjannet, S.; Wickham, L.; Marcinkiewicz, J.; Jasmin, S.B.; Stifani, S.; Basak, A.; Prat, A.; Chretien, M. The secretory proprotein convertase neural apoptosis- regulated convertase 1 (NARC1): Liver regeneration and neuronal differentiation. Proc. Natl. Acad. Sci. USA 2003, 100, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Page, M.M.; Watts, G.F. Experimental and clinical pharmacology: PCSK9 inhibitors- mechanisms of action. Aust. Prescr. 2016, 39, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Glerup, S.; Schulz, R.; Laufs, U.; Schlüter, K.D. Physiological and therapeutic regulation of PCSK9 activity in cardiovascular disease. Basic Res. Cardiol. 2017, 112, 1–23. [Google Scholar] [CrossRef]
- Paton, D. PCSK9 inhibitors: Monoclonal antibodies for the treatment of hypercholesterolemia. Drugs Today 2016, 52, 183. [Google Scholar] [CrossRef] [PubMed]
- Warden, B.A.; Duell, P.B. Inclisiran: A novel agent for lowering apolipoprotein B-containing lipoproteins. J. Cardiovasc. Pharmacol. 2021, 78, e157–e174. [Google Scholar] [CrossRef]
- Dressel, A.; Schmidt, B.; Schmidt, N.; Laufs, U.; Fath, F.; Chapman, M.J.; Grammer, T.B.; März, W. Cost effectiveness of lifelong therapy with PCSK9 inhibitors for lowering cardiovascular events in patients with stable coronary artery disease: Insights from the Ludwigshafen Risk and Cardiovascular Health cohort. Vascul. Pharmacol. 2019, 120, 106566. [Google Scholar] [CrossRef] [PubMed]
- Jick, H.; Wilson, A.; Wiggins, P.; Chamberlin, D.P. Comparison of prescription drug costs in the United States and the United Kingdom, part 1: Statins. Pharmacotherapy 2012, 32, 1–6. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef]
- Dong, G.; Xu, T.; Yang, B.; Lin, X.; Zhou, X.; Yang, X.; Liu, Y. Chemical constituents and bioactivities of starfish. Chem. Biodiver. 2011, 8, 740–791. [Google Scholar] [CrossRef]
- Lee, C.C.; Hsieh, H.J.; Hsieh, C.H.; Hwang, D.F. Antioxidative and anticancer activities of various ethanolic extract fractions from crown-of-thorns starfish (Acanthaster planci). Environ. Toxicol. Pharmacol. 2014, 38, 761–773. [Google Scholar] [CrossRef]
- Andersson, L.; Bohlin, L.; Iorizzi, M.; Riccio, R.; Minale, L.; Moreno-López, W. Biological activity of saponins and saponin-like compounds from starfish and brittle-stars. Toxicon 1989, 27, 179–188. [Google Scholar] [CrossRef]
- Komori, T.; Nanri, H.; Itakura, Y.; Sakamoto, K.; Taguchi, S.; Higuchi, R.; Kawasaki, T.; Higuchi, T. Biologically active glycosides from asteroidea, III. Steroid oligoglycosides from the starfish Acanthaster planci L., 2. Structures of two newly characterized genuine sapogenins and an oligoglycoside sulfate. Liebigs Ann. Chem. 1983, 1983, 37–55. [Google Scholar] [CrossRef]
- Farid, C.G. Acanthaster planci biomass potential in medical and pharmaceutical therapeutics. Malays. J. Microsc. 2015, 11, 107–114. [Google Scholar]
- Mat-Lazim, N.H.; Asari, A.; Mohamad, F.; Ismail, N.; Ahmad, A.; Taib, M.; Mohamad, H. Potential anti-atherosclerotic compound isolated from Acanthaster planci. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 482–487. [Google Scholar]
- Mohamad, H.; Abd Razak, M.F.; Kamaruddin, N.N.; Mohd Din, L.H.; Asari, A.; Andriani, Y.; Mustafa, S.F.Z.; Saidin, J.; Mohd Aluwi, M.F.F.; Latip, J.; et al. PCSK9 inhibitory activity of marine-derived compounds, aaptaminoids, and benzamide originated from Aaptos aaptos and Acanthaster planci as a potential treatment for atherosclerosis. J. Appl. Pharm. Sci. 2020, 10, 111–123. [Google Scholar] [CrossRef]
- Boik, J. Natural Compounds in Cancer Therapy; Oregon Medical Press: Princeton, MN, USA, 2001. [Google Scholar]
- Cao, S.; Sng, V.H.; Wu, X.; Sim, K.; Tan, B.; Pereira, J.; Goh, S. Novel cytotoxic polyprenylated xanthonoids from Garcinia gaudichaudii (Guttiferae). Tetrahedron 1998, 54, 10915–10924. [Google Scholar] [CrossRef]
- Tavori, H.; Rashid, S.; Fazio, S. On the function and homeostasis of PCSK9: Reciprocal interaction with LDLR and additional lipid effects. Atherosclerosis 2015, 238, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Michael, M.C. Lehninger Principles of Biochemistry, 3rd ed.; Worth Publisher: New York, NY, USA, 2000. [Google Scholar]
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H. Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 38, 2459–2472. [Google Scholar] [CrossRef]
- Klein-Szanto, A.J.; Bassi, D.E. Keep recycling going: New approaches to reduce LDL-C. Biochem. Pharmacol. 2019, 164, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Garg, J.; Shah, N.; Sumner, A. PCSK9 inhibitors: A new era of lipid lowering therapy. World J. Cardiol. 2017, 9, 76. [Google Scholar] [CrossRef]
- Zhang, J.Z. An Approach to Analyzing Histology Segmentations Using Shape Distributions. Master’s Thesis, Drexel University, Philadelphia, PA, USA, 2008. [Google Scholar]
- Katzmann, J.L.; Gouni-Berthold, I.; Laufs, U. PCSK9 Inhibition: Insights from clinical trials and future prospects. Front. Physiol. 2020, 11, 1391. [Google Scholar] [CrossRef]
- European Medicines Agency. EMA. 2021. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/leqvio (accessed on 4 March 2021).
- Lamb, Y.N. Inclisiran: First approval. Drugs 2021, 81, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Baum, S.J.; Cannon, C.P. PCSK9 inhibitor valuation: A science-based review of the two recent models. Clin. Cardiol. 2018, 41, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Rady, H.; Salem, S.; El-Arab, M.E. Primmorph extracts and mesohyls of marine sponges inhibit proliferation and migration of hepatocellular carcinoma cells in vitro. J. Pharm. Anal. 2019, 9, 284–291. [Google Scholar] [CrossRef]
- Cui, H.; Bashar, M.A.E.; Rady, I.; El-Naggar, H.A.; Abd El-Maoula, L.M.; Mehany, A.B.M. Antiproliferative activity, proapoptotic effect, and cell cycle arrest in human cancer cells of some marine natural product extract. Oxid. Med. Cell. Longev. 2020, 2020, 7948705. [Google Scholar] [CrossRef] [PubMed]
- Mat-Lazim, N.H. Identification of Bioactive Compounds From Acanthaster planci and Their Anti-atherosclerotic Activity. Master Thesis, Universiti Malaysia Terengganu, Kuala Terengganu, TRG, Malaysia, 2013. [Google Scholar]
- Luqman, A.B. The Effect of Methyl Benzoate and Other Fractions from Acanthaster planci in Reducing Hypercholesterolemia. Ph.D. Thesis, Universiti Malaysia Terengganu, Kuala Terengganu, TRG, Malaysia, 2019. [Google Scholar]
- Choi, H.; Hwang, J.; Nam, T.; Kim, S.H.; Min, D.; Park, S.W.; Chung, M. Welsh onion extract inhibits PCSK9 expression contributing to the maintenance of the LDLR level under lipid depletion conditions of HepG2 cells. Food Funct. 2017, 8, 4582–4591. [Google Scholar] [CrossRef]
- Song, K.H.; Kim, Y.H.; Im, A.; Kim, Y.H. Black raspberry extract enhances ldl uptake in Hepg2 cells by suppressing PCSK9 expression to upregulate LDLR expression. J. Med. Food 2018, 21, 560–567. [Google Scholar] [CrossRef]
- Chang, H.; Wu, J.; Gao, W.; Lin, H.; Chen, P.; Chen, C.; Wu, M.; Yen, J. The cholesterol- modulating effect of methanol extract of pigeon pea (Cajanus cajan (L.) Millsp.) leaves on regulating LDLR and PCSK9 expression in HepG2 cells. Molecules 2019, 24, 493. [Google Scholar] [CrossRef]
- Hwang, J.; Choi, E.; Choi, H.; Park, J.; Chung, M. The cholesterol-lowering effect of Capsella Bursa-Pastoris is mediated via SREBP2 and HNF-1α-regulated PCSK9 inhibition in obese mice and HepG2 cells. Foods 2021, 10, 408. [Google Scholar] [CrossRef]
- Mat-Lazim, N.H.; Asari, A.; Mohamad, F.; Tengku Muhammad, T.S.; Ismail, N.; Taib, M.; Ahmad, A.; Mohamad, H. Phenyl ethanone and sterols from Acanthaster planci as potential PPAR-ligand. J. Chem. Pharm. Res. 2015, 7, 121–126. [Google Scholar]
- Mitsuya, H.; Weinhold, K.J.; Furman, P.A.; St Clair, M.H.; Lehrman, S.N.; Gallo, R.C.; Bolognesi, D.; Barry, D.W.; Broder, S. 3′-Azido-3′-deoxythymidine (BWA509U): An antiviral agent that inhibits the infectivity and cytopathic effect of human T-lymphotropic virus type III/lymphadenopathy-associated virus in vitro. Proc. Natl. Acad. Sci. USA 1985, 82, 7096–7100. [Google Scholar] [CrossRef] [PubMed]
- Furman, P.A.; Fyfe, J.A.; St Clair, M.H.; Weinhold, K.; Rideout, J.L.; Freeman, G.A.; Lehrman, S.N.; Bolognesi, D.P.; Broder, S.; Mitsuya, H. Phosphorylation of 3′-azido-3′-deoxythymidine and selective interaction of the 5′-triphosphate with human immunodeficiency virus reverse transcriptase. Proc. Natl. Acad. Sci. USA 1986, 83, 8333–8337. [Google Scholar] [CrossRef]
- Mitsuya, H.; Broder, S. Strategies for antiviral therapy in AIDS. Nature 1987, 325, 773–778. [Google Scholar] [CrossRef]
- Cameron, J.; Ranheim, T.; Kulseth, M.A.; Leren, T.P.; Berge, K.E. Berberine decreases PCSK9 expression in HepG2 cells. Atherosclerosis 2008, 201, 266–273. [Google Scholar] [CrossRef]
- Tai, M.; Chen, P.; Chen, P.; Wu, M.; Ho, C.; Yen, J. Curcumin enhances cell- surface LDLR level and promotes LDL uptake through downregulation of PCSK9 gene expression in HepG2 cells. Mol. Nutr. Food Res. 2014, 58, 2133–2145. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Hu, T.; Lin, C.; Xiong, Q.; Liu, F.; Yuan, J.; Zhao, X.; Wang, R. Resveratrol downregulates PCSK9 expression and attenuates steatosis through estrogen receptor-α mediated pathway in L02 cells. Eur. J. Pharmacol. 2019, 855, 216–226. [Google Scholar] [CrossRef]
- Cui, C.; Jin, J.; Guo, L.; Sun, J.; Wu, N.; Guo, Y.; Liu, G.; Dong, Q.; Li, J. Beneficial impact of epigallocatechin gallate on LDL-C through PCSK9/LDLR pathway by blocking HNF1α and activating FoxO3a. J. Transl. Med. 2020, 18, 1–13. [Google Scholar] [CrossRef]
- Segatto, M.; Trapani, L.; Marino, M.; Pallottini, V. Age- and sex-related differences in extra-hepatic low-density lipoprotein receptor. J. Cell. Physiol. 2011, 226, 2610–2616. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Hobbs, H.H.; Brown, M.S. Familial hypercholesterolemia. In The Metabolic and Molecular Bases of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S.D., Valle, D., Eds.; McGraw-Hill, Inc.: New York, NY, USA, 2001; pp. 2863–2913. [Google Scholar]
- Davis, C.G.; Goldstein, J.L.; Südhof, T.C.; Anderson, R.G.W.; Russell, D.W.; Brown, M.S. Acid-dependent ligand dissociation and recycling of LDL receptor mediated by growth factor homology region. Nature 1987, 326, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lagace, T.A.; Garuti, R.; Zhao, Z.; Mcdonald, M.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Binding of Proprotein Convertase Subtilisin/Kexin Type 9 to epidermal growth factor-like repeat A of low density lipoprotein receptor decreases receptor recycling and increases degradation. J. Biol. Chem. 2007, 282, 18602–18612. [Google Scholar] [CrossRef] [PubMed]
- Seidah, N.G.; Awan, Z.; Chrétien, M.; Mbikay, M. PCSK9: A key modulator of cardiovascular health. Circ. Res. 2014, 114, 1022–1036. [Google Scholar] [CrossRef] [PubMed]
- Andriani, Y.; Ramli, N.M.; Syamsumir, D.F.; Kassim, M.N.I.; Jaafar, J.; Azis, N.A.; Marlina, L.; Musa, N.S.; Mohamad, H. Phytochemical analysis, antioxidant, antibacterial and cytotoxicity activities of keys and cores part of Pandanus tectorius fruits. Arabian J. Chem. 2015, 12, 3555–3564. [Google Scholar] [CrossRef]
- Gilat, T.; Leikin-Frenkel, A.; Goldiner, I.; Juhel, C.; Lafont, H.; Gobbi, D.; Konikof, F.M. Prevention of diet-induced fatty liver in experimental animals by the oral administration of a fatty acid bile acid conjugate (FABAC). Hepatology 2003, 38, 436–442. [Google Scholar] [CrossRef]
- AlSharari, S.D.; Al-Rejaie, S.S.; Abuohashish, H.M.; Ahmed, M.M.; Hafez, M.M. Rutin attenuates hepatoxicity in high-cholesterol-diet-fed-rats. Oxid. Med. Cell. Longev. 2016, 2016, 5436745. [Google Scholar] [CrossRef]
- Sa’adah, N.N.; Purwani, K.I.; Nurhayati, A.P.D.; Ashuri, N.M. Analysis of lipid profile and atherogenic index in hyperlipidemic rat (Rattus norvegicus Berkenhout, 1769) that given the methanolic extract of Parijoto (Medinilla speciosa). AIP Conf. Proc. 2017, 1854, 020031-1–020031-8. [Google Scholar] [CrossRef]
- Rachmawati, N.; Wasita, B.; Kartikasari, L. Basil Leaves (Ocimum sanctum linn.) extract decreases total cholesterol levels in hypercholesterolemia Sprague Dawley rats model. IOP Conf. Ser. Mater. Sci. Eng. 2019, 546, 062020. [Google Scholar] [CrossRef]
- Pangestika, I.; Oksal, E.; Tengku Muhammad, T.S.; Amir, H.; Syamsumir, D.F.; Wahid, M.E.A.; Andriani, Y. Inhibitory effects of tangeretin and trans-ethyl caffeate on the HMG-COA reductase activity: Potential agents for reducing cholesterol levels. Saudi J. Biol. Sci. 2020, 27, 1947–1960. [Google Scholar] [CrossRef] [PubMed]
- Nurhidajah; Astuti, R.; Nurrahman. Black rice potential in HDL and LDL profile in Sprague Dawley rat with high cholesterol diet. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012019. [Google Scholar] [CrossRef]
- Johnston, T.P.; Nguyen, L.B.; Chu, W.A.; Shefer, S. Potency of select statin drugs in a new mouse model of hyperlipidemia and atherosclerosis. Int. J. Pharm. 2001, 229, 75–86. [Google Scholar] [CrossRef]
- Eguchi, K.; Fujiwara, Y.; Hayashida, A.; Horlad, H.; Kato, H.; Rotinsulu, H.; Losung, F.; Mangindaan, R.E.; De Voogd, N.J.; Takeya, M. Manzamine A, a marine-derived alkaloid, inhibits accumulation of cholesterol ester in macrophages and suppresses hyperlipidemia and atherosclerosis in vivo. Bioorg. Med. Chem. 2013, 21, 3831–3838. [Google Scholar] [CrossRef]
- Yeo, A.; Lee, J.; Tae, I.; Park, S.; Cho, Y.; Lee, B.; Shin, H.; Kim, S.; Yoo, Y. Anti-hyperlipidemic effect of polyphenol extract (SeapolynolTM) and dieckol isolated from Ecklonia cava in in vivo and in vitro Models. Prev. Nutr. Food Sci. 2012, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yoon, N.Y.; Kim, H.R.; Chung, H.Y.; Choi, J.S. Anti-hyperlipidemic effect of an edible brown algae, Ecklonia stolonifera, and its constituents on poloxamer 407-induced hyperlipidemic and cholesterol-fed rats. Arch. Pharm. Res. 2008, 31, 1564–1571. [Google Scholar] [CrossRef]
- Catapano, A.L.; Papadopoulos, N. The safety of therapeutic monoclonal antibodies: Implications for cardiovascular disease and targeting the PCSK9 pathway. Atherosclerosis 2013, 228, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Deepa, P.R.; Varalakshmi, P. Atheroprotective effect of exogenous heparin derivative treatment on the aortic disturbances and lipoprotein oxidation in hypercholesterolemic diet fed rats. Clin. Chim. Acta. 2005, 355, 119–130. [Google Scholar] [CrossRef]
- Mathews, C.K.; Holde, K.E.V.; Appling, D.R.; Antony-cahil, S.J. Biochemistry, 4th ed.; Pearson Canada: Toronto, ON, Canada, 2013. [Google Scholar]
- Akpanabiatu, M.I.; Umoh, I.B.; Udosen, E.O.; Udoh, A.E.; Edet, E.E. Rat serum electrolytes, lipid profile and cardiovascular activity on Nauclea latifolia leaf extract administration. Indian J. Clin. Biochem. 2005, 20, 29–34. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Hegele, R.A.; Koenig, W. Cholesterol-lowering agents. Circ. Res. 2019, 124, 364–385. [Google Scholar] [CrossRef] [PubMed]
- Maron, D.J.; Lu, G.P.; Cai, N.S.; Wu, Z.G.; Li, Y.H.; Chen, H.; Zhu, J.Q.; Jin, X.J.; Wouters, B.C.; Zhao, J. Cholesterol-lowering effect of a theaflavin-enriched green tea extract: A randomized controlled trial. Arch. Int. Med. 2003, 163, 448–1453. [Google Scholar] [CrossRef]
- Rajendran, R.; Hemalatha, S.; Akasakalai, K.; Madhu-Khrisna, C.H.; Sohil, B.V.; Sundaram, R.M. Hepatoprotective activity of Mimosa pudica leaves against carbon tetrachloride induced toxicity. J. Nat. Prod. 2009, 2, 116–122. [Google Scholar]
- Karwani, G.; Sosodia, S. Hepatoprotective activity of Mimosa pudica Linn. in carbon tetra chloride induced hepatotoxicity in rats. J. Herb. Med. Toxicol. 2011, 5, 27–32. [Google Scholar]
- Ravikumar, S.; Gnanadesigan, M. Hepatoprotective and antioxidant activity of a mangrove plant Lumnitzera racemosa. Asian Pac. J. Trop. Biomed. 2011, 1, 348–352. [Google Scholar] [CrossRef]
- Jeong, W.I.; Jeong, D.H.; Do, S.H.; Kim, Y.K.; Park, H.Y.; Kwon, O.D.; Kim, T.H.; Jeong, K.S. Mild hepatic fibrosis in cholesterol and sodium cholate diet-feed rats. J. Vet. Med. Sci. 2005, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Barltrop, J.A.; Owen, T.C.; Cory, A.H.; Cory, J.G. 5-(3-carboxymethoxyphenyl)-2-(4,5-dimethylthiazolyl)-3-(4-sulfophenyl)tetrazolium, inner salt (MTS) and related analogs of 3-(4,5-dimethylthiazolyl)-2,5-diphenyltetrazolium bromide (MTT) reducing to purple water-soluble formazans as cell-viability indicators. Bioorg. Med. Chem. Lett. 1991, 1, 611–614. [Google Scholar] [CrossRef]
- Nissen, S.E.; Wolski, K. Effects of Rosiglitazone on the risk of myocardial infarction and death from cardiovascular diseases. N. Engl. J. Med. 2007, 356, 2457–2472. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Lee, H.S.; Kim, K.S.; Kim, Y.K.; Yoon, D.; Park, S.W. Sterol-dependent regulation of Proprotein Convertase Subtilisin/Kexin Type 9 expression by Sterol-Regulatory Element Binding Protein-2. J. Lipid Res. 2008, 49, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Xu, R.; Sun, J.; Tang, Y.; Li, J. Enhanced circulating PCSK9 concentration by berberine through SREBP-2 pathway in high fat diet-fed rats. J. Transl. Med. 2014, 12, 103. [Google Scholar] [CrossRef]
- Al-Najjar, B.O.; Wahab, H.A.; Tengku Muhammad, T.S.; Shu-Chien, A.C.; Ahmad Noruddin, N.A.; Taha, M.O. Discovery of new nanomolar Peroxisome Proliferator-activated Receptor γ Activators via elaborate ligand-based modelling. Eur. J. Med. Chem. 2011, 46, 2513–2529. [Google Scholar] [CrossRef]
- Duan, Y.; Chen, Y.; Hu, W.; Li, X.; Yang, X.; Zhou, X.; Yin, Z.; Kong, D.; Yao, Z.; Hajjar, D.P.; et al. Peroxisome Proliferator-Activated Receptor γ activation by ligands and dephosphorylation induces Proprotein Convertase Subtilisin Kexin Type 9 and low density lipoprotein receptor expression. J. Biol. Chem. 2012, 287, 23667–23677. [Google Scholar] [CrossRef]
- Andriani, Y.; Chaudhry, G.; Oksal, E.; Pangestika, I.; Ramli, N.M.; Mohamad, H.; Elvia, R.; Amir, H.; Effendy, A.W.M.; Sung, Y.Y.; et al. Antihypercholesterolemic and antiatherosclerotic potencies of Pandanus tectorius fruits via increasing Scavenger Receptor-B1 genes expression and inhibition of 3-hydroxy-3-methylglutaryl coenzyme: A reductase activity. J. Adv. Pharm. Technol. Res. 2020, 11, 30–35. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | The Level of SGOT (U/I) during the Treatment Period Measured Every 7 Days | ||||||
|---|---|---|---|---|---|---|---|
| Day 0 | Day 7 | Day 14 | Day 21 | Day 28 | Day 35 | Day 42 | |
| A | 102.0 ± 19.8 | 90.1 ± 10.2 | 91.7 ± 3.8 | 96.3 ± 12.4 | 106.7 ± 15.9 | 94.3 ± 15.9 | 97.8 ± 10.2 |
| B | 103.7 ± 9.8 | 101.6 ± 15.7 | 99.3 ± 17.5 | 100.8 ± 7.9 | 102.1 ± 18.3 | 99.0 ± 5.4 | 114.3 ± 10.2 |
| C | 97.5 ± 13.2 | 104.0 ± 14.7 | 96.5 ± 16.5 | 102.5 ± 11.1 | 106.6 ± 11.1 | 108.7 ± 11.4 | 104.0 ± 24.5 |
| D | 105 ± 2.4 | 98.0 ± 10.1 | 96.8 ± 15.0 | 90.0 ± 5.0 | 91.3 ± 3.6 | 86.3 ± 2.0 | 89.3 ± 5.2 |
| E | 92.5 ± 11.5 | 86.6 ± 4.2 | 89.6 ± 6.0 | 95.4 ± 13.3 | 100.0 ± 17.2 | 97.0 ± 8.3 | 91.0 ± 2.2 |
| Group | The Level of SGPT (U/I) during the Treatment Period Measured Every 7 Days | ||||||
|---|---|---|---|---|---|---|---|
| Day 0 | Day 7 | Day 14 | Day 21 | Day 28 | Day 35 | Day 42 | |
| A | 42.6 ± 7.8 | 46.1 ± 8.0 | 46.9 ± 7.2 | 48.2 ± 3.3 | 44.7 ± 6.3 | 46.5 ± 7.6 | 43.8 ± 11.9 |
| B | 42.4 ± 4.7 | 38.6 ± 7.1 | 48.7 ± 6.2 | 48.9 ± 7.2 | 45.2 ± 6.9 | 42.7 ± 8.4 | 41.5 ± 17.1 |
| C | 39.5 ± 4.4 | 38.3 ± 5.8 | 44.0 ± 3.3 | 44.5 ± 7.2 | 43.3 ± 2.4 | 38.0 ± 2.9 | 37.7 ± 4.6 |
| D | 40.6 ± 7.1 | 44.2 ± 4.8 | 41.0 ± 6.3 | 40.3 ± 5.0 | 43.8 ± 4.5 | 49.0 ± 7.1 | 45.0 ± 8.5 |
| E | 42.1 ± 6.7 | 45.8 ± 5.6 | 43.0 ± 4.1 | 42.5 ± 7.0 | 43.8 ± 6.2 | 34.7 ± 8.1 | 42.3 ± 7.6 |
| Group | Number of Rats | Diet | Treatment |
|---|---|---|---|
| A | 8 | Normal diet for 42 days | No treatment |
| B | 8 | High fat diet for 14 days then continued with normal diet for an additional 28 days | No treatment |
| C | 8 | High cholesterol diet for 14 days then continued with normal diet for an additional 28 days | Atorvastatin (positive control) from Day 15 to 28 |
| D | 8 | High cholesterol diet for 14 days then continued with normal diet for an additional 28 days | A. planci methanolic extract dose 1 (100 mg/kg) from Day 15 to 28 |
| E | 8 | High cholesterol diet for 14 days then continued with normal diet for an additional 28 days | A. planci methanolic extract dose 2 (50 mg/kg) from Day 15 to 28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamaruddin, N.N.; Hajri, N.A.; Andriani, Y.; Abdul Manan, A.F.; Tengku Muhammad, T.S.; Mohamad, H. Acanthaster planci Inhibits PCSK9 and Lowers Cholesterol Levels in Rats. Molecules 2021, 26, 5094. https://doi.org/10.3390/molecules26165094
Kamaruddin NN, Hajri NA, Andriani Y, Abdul Manan AF, Tengku Muhammad TS, Mohamad H. Acanthaster planci Inhibits PCSK9 and Lowers Cholesterol Levels in Rats. Molecules. 2021; 26(16):5094. https://doi.org/10.3390/molecules26165094
Chicago/Turabian StyleKamaruddin, Nurjannatul Naim, Nor Azwin Hajri, Yosie Andriani, Aina Farahiyah Abdul Manan, Tengku Sifzizul Tengku Muhammad, and Habsah Mohamad. 2021. "Acanthaster planci Inhibits PCSK9 and Lowers Cholesterol Levels in Rats" Molecules 26, no. 16: 5094. https://doi.org/10.3390/molecules26165094
APA StyleKamaruddin, N. N., Hajri, N. A., Andriani, Y., Abdul Manan, A. F., Tengku Muhammad, T. S., & Mohamad, H. (2021). Acanthaster planci Inhibits PCSK9 and Lowers Cholesterol Levels in Rats. Molecules, 26(16), 5094. https://doi.org/10.3390/molecules26165094