
Stable DOPG/Glycyrrhizin Vesicles with a Wide Range of Mixing Ratios: Structure and Stability as Seen by Scattering Experiments and Cryo-TEM

 , ,
, ,  , , , and
, , , and 
Abstract
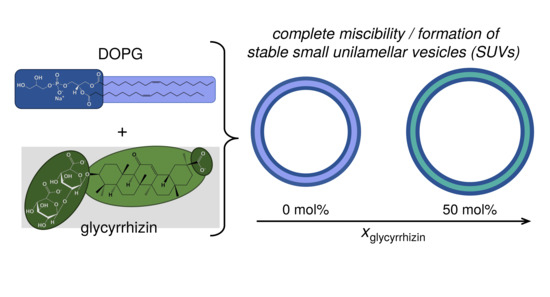
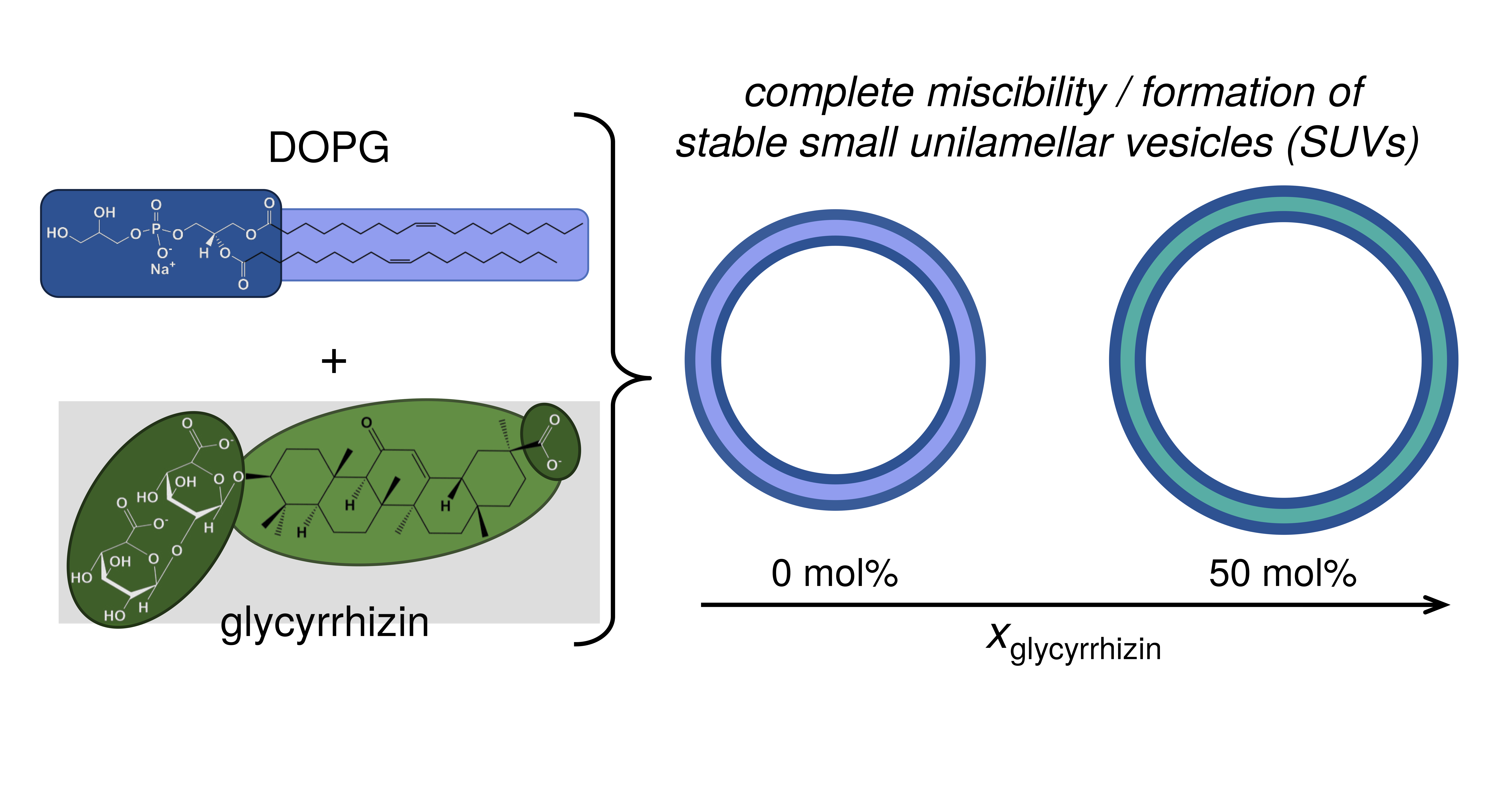
1. Introduction
2. Materials and Methods
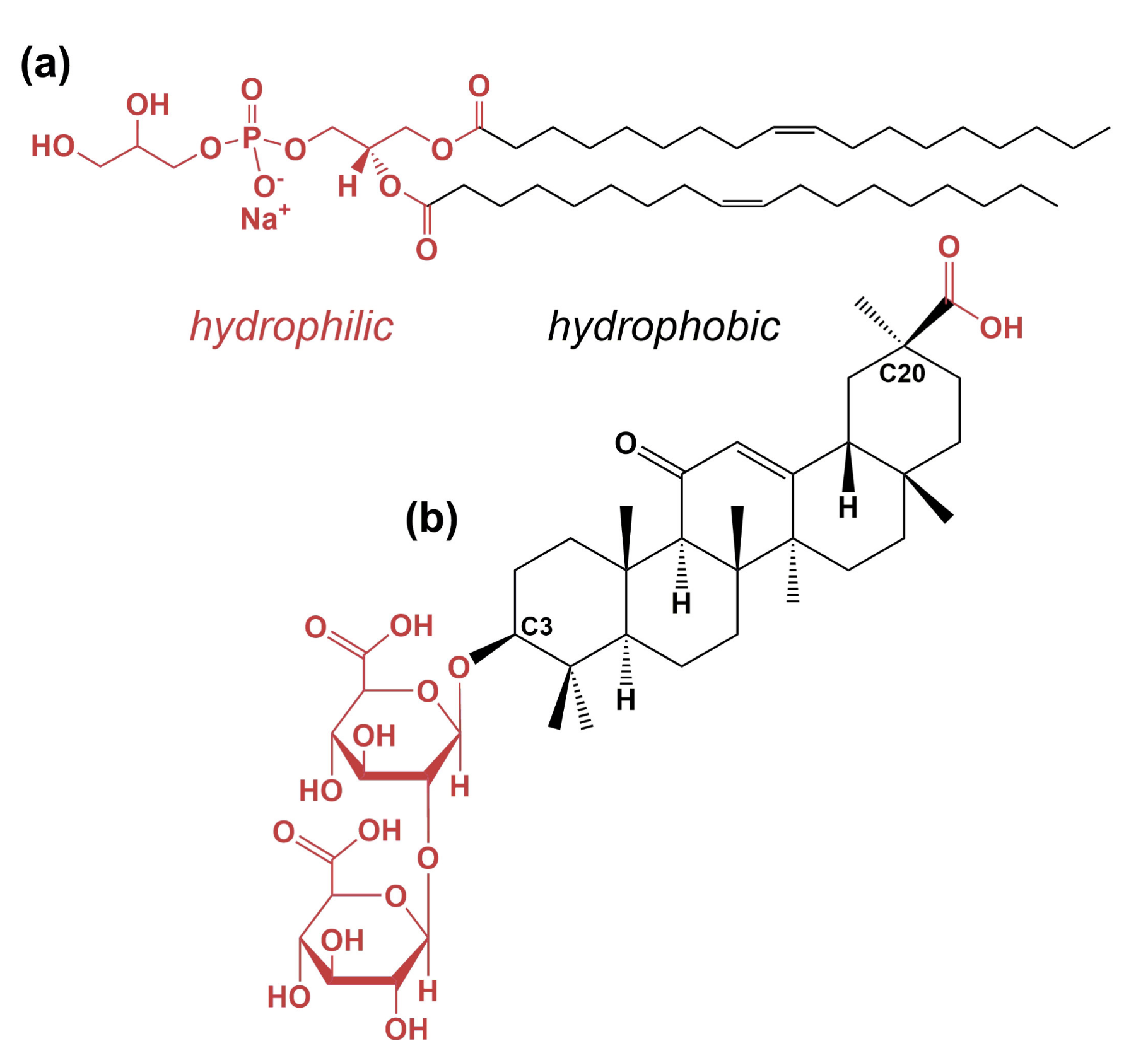
2.1. Chemicals and Sample Preparation
2.2. Cryogenic Transmission Electron Microscopy (Cryo-Tem)
2.3. Small-Angle Scattering (SAS)
2.3.1. Small-Angle Neutron Scattering (SANS)
2.3.2. Small-Angle X-ray Scattering (SAXS)
2.4. Wide-Angle X-ray Scattering (WAXS)
3. Results
3.1. General Phase Behavior and Identification of Particle Shape by Cryo-TEM
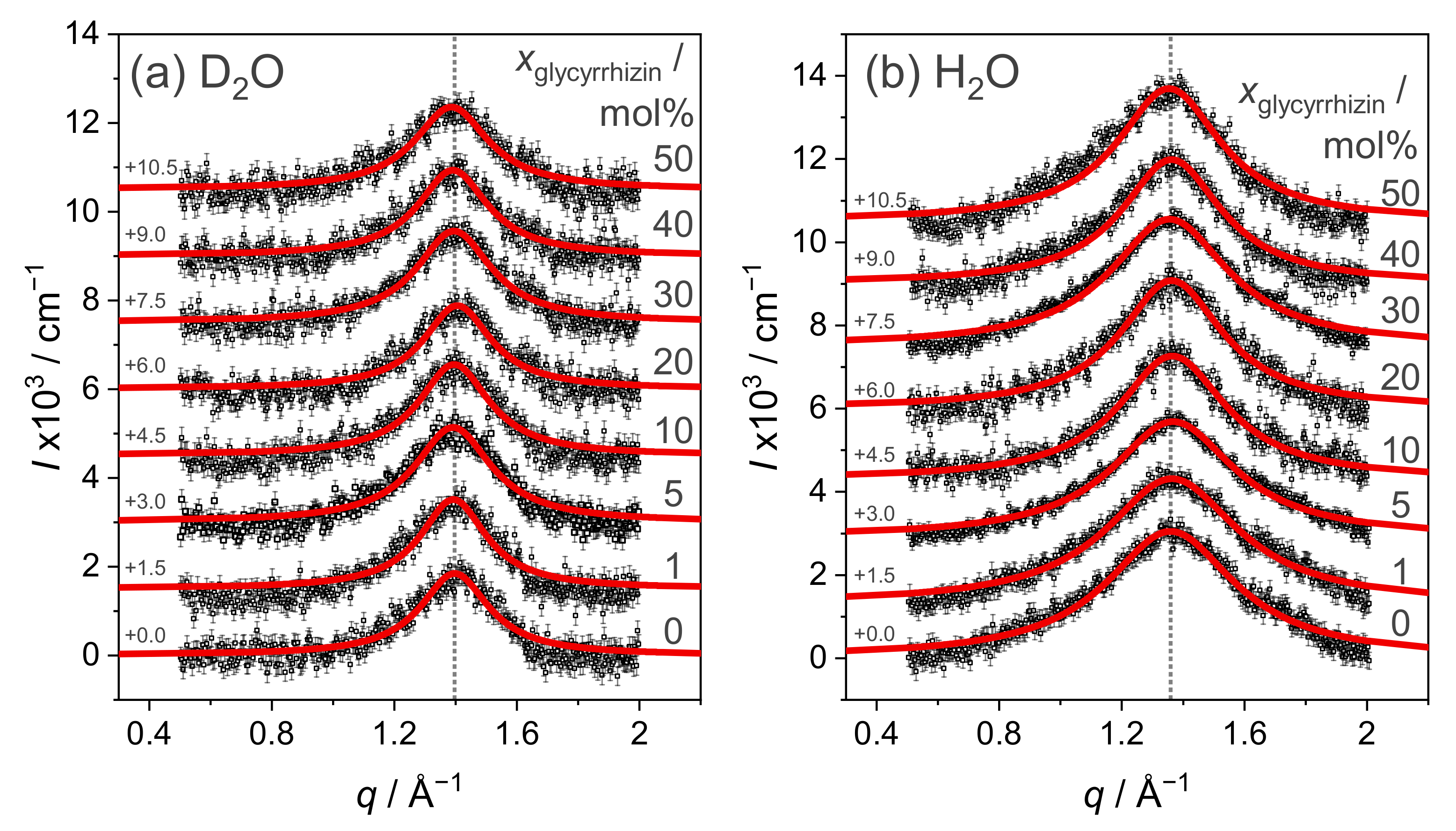
3.2. Influence of Glycyrrhizin on the Chain–Chain Correlation Distance in DOPG Membranes Resolved by WAXS
3.3. Characterization of SUV Structure by SAS
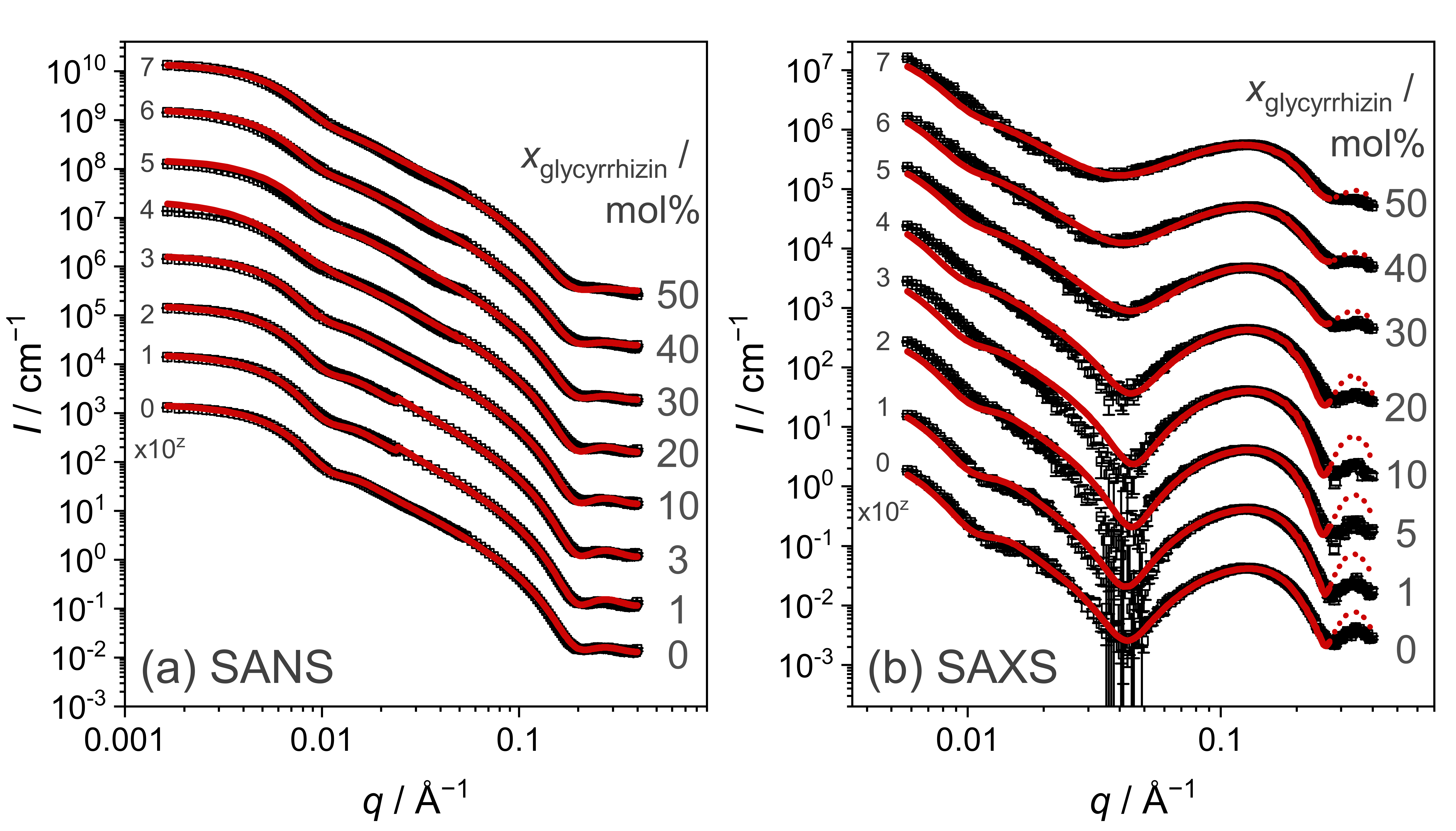
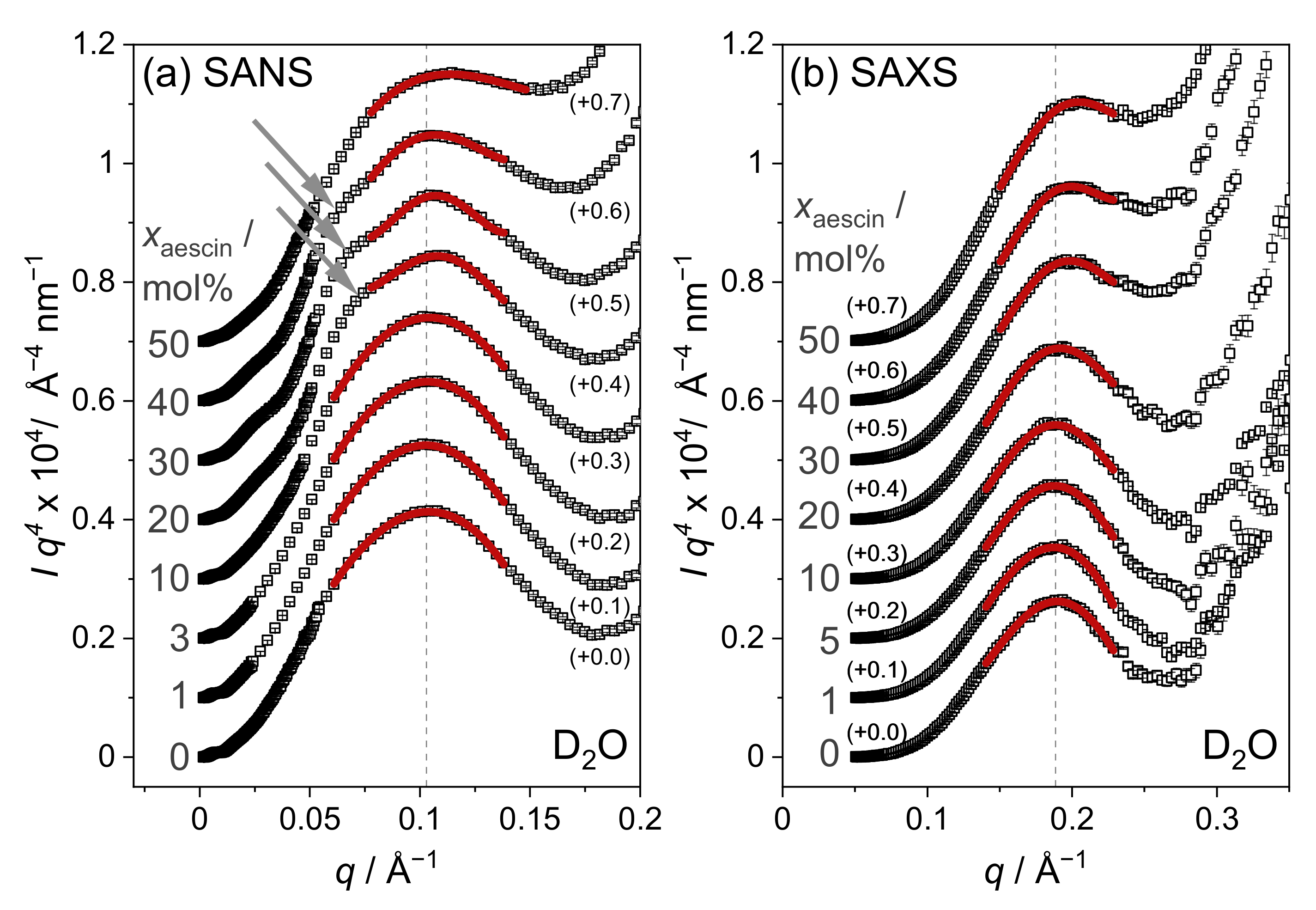
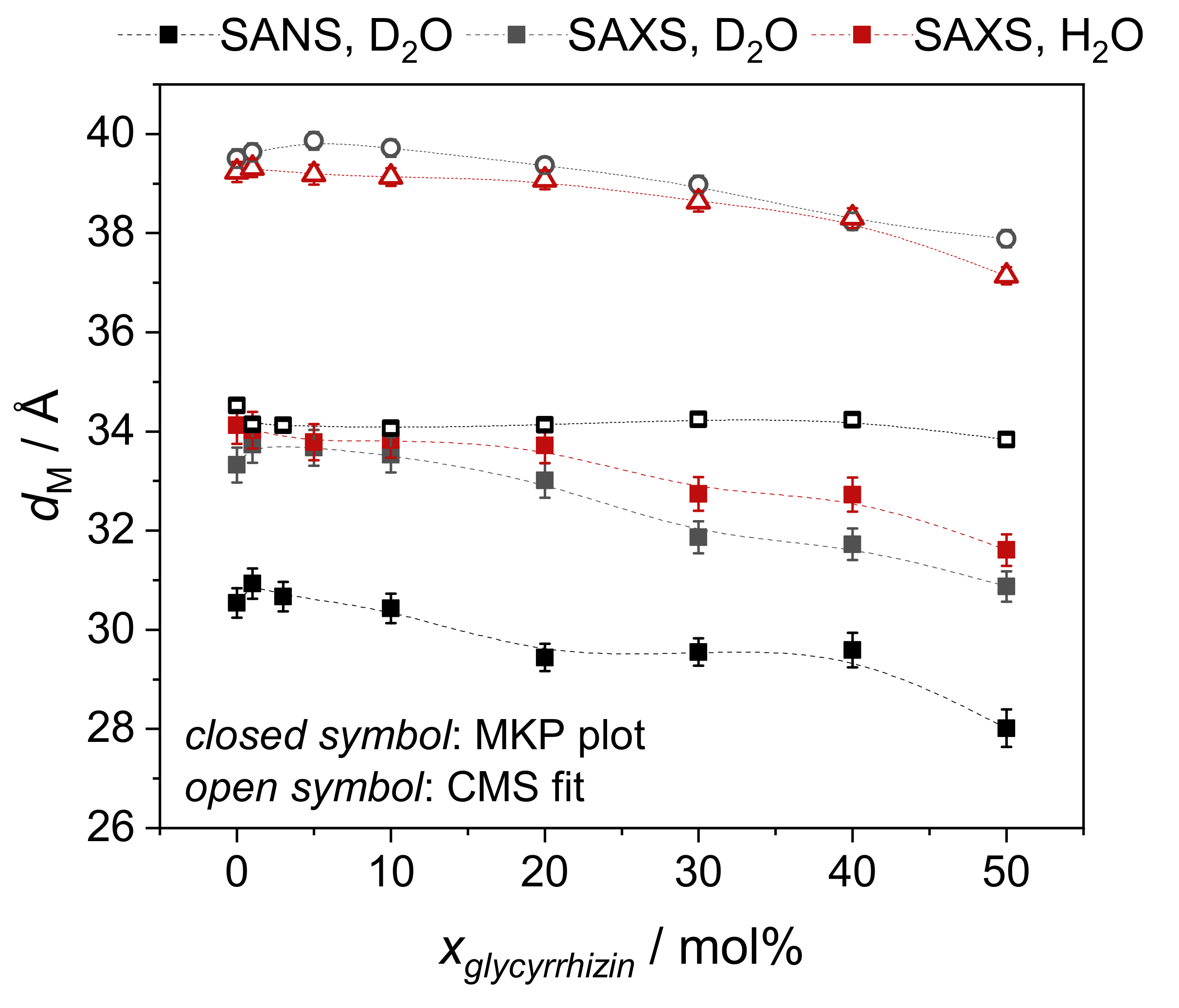
3.3.1. Model-Independent Evaluation of SAS Data
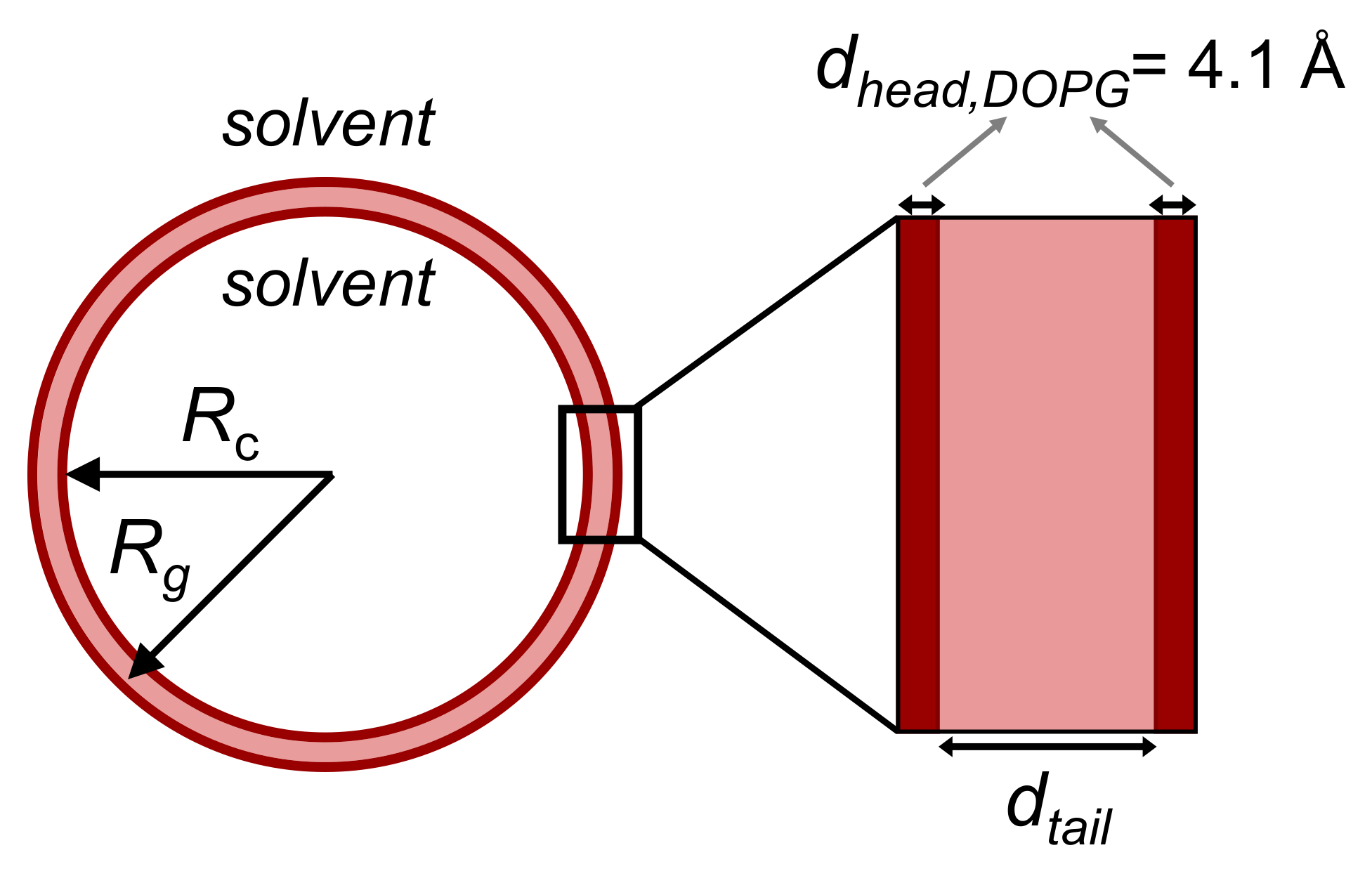
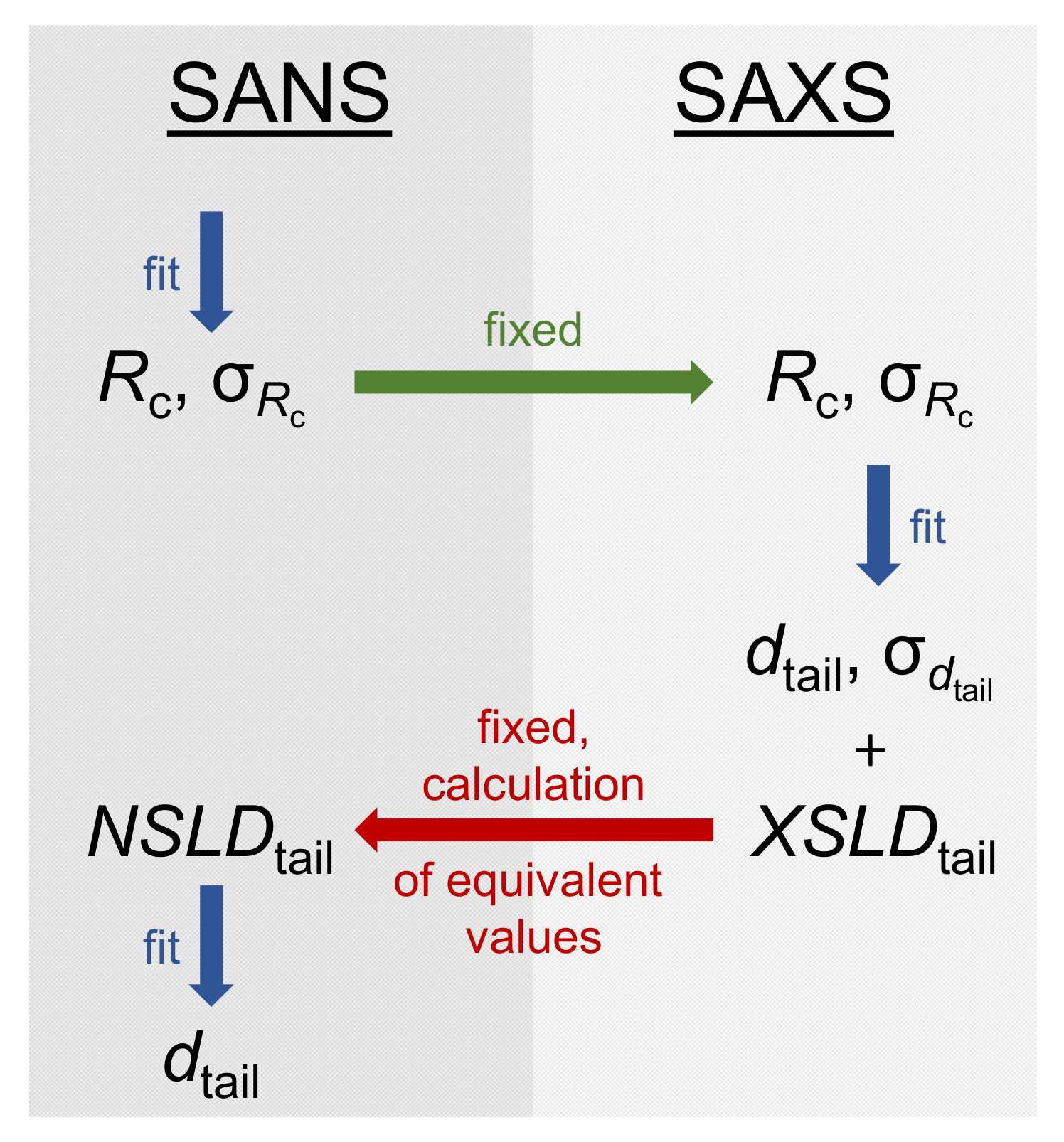
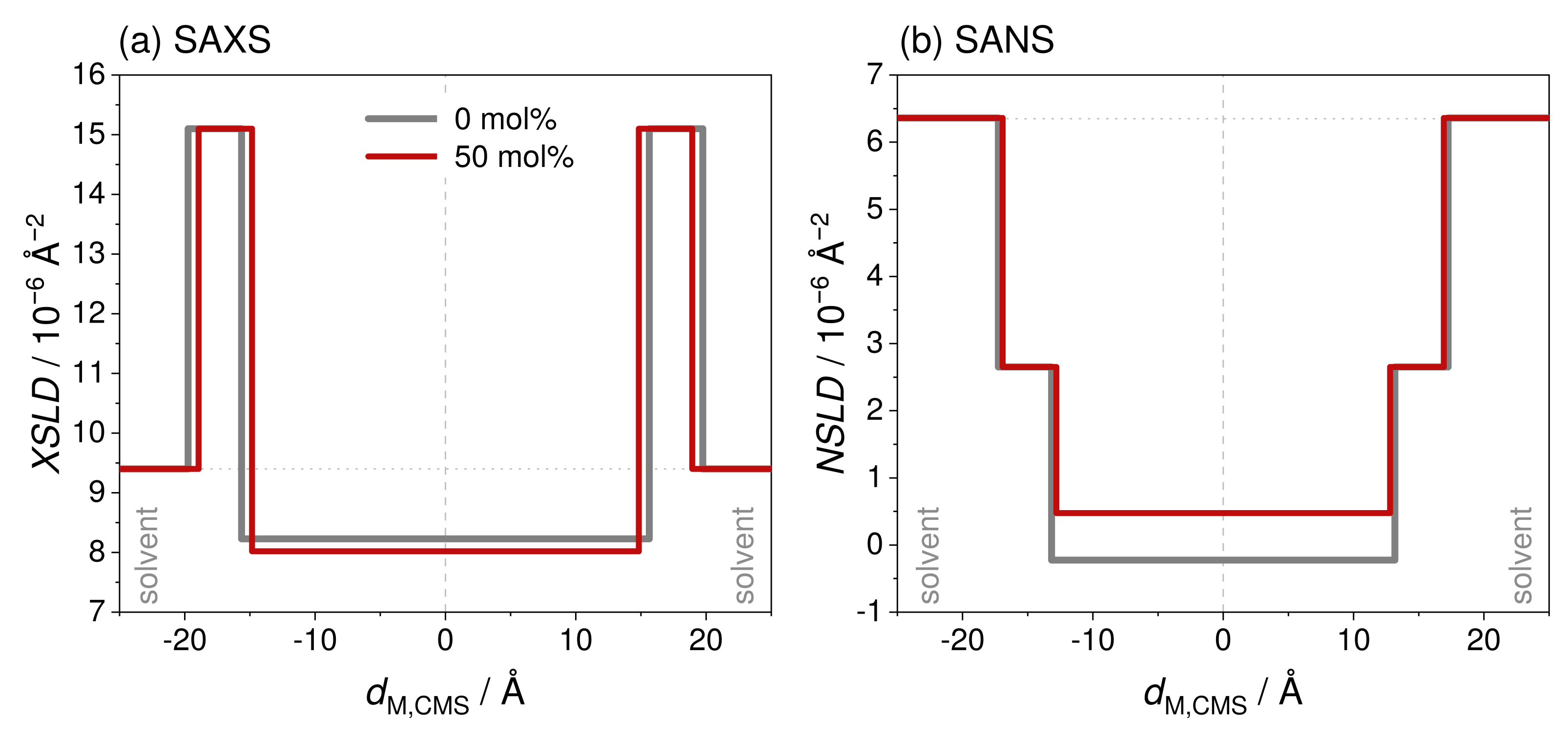
3.3.2. Model-Dependent Fitting of SAS Data
4. Conclusions and Outlook
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Güçlü-Üstündağ, O.; Mazza, G. Saponins: Properties, applications and processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef] [PubMed]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef] [PubMed]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef]
- Góral, I.; Wojciechowski, K. Surface activity and foaming properties of saponin-rich plants extracts. Adv. Colloid Interface Sci. 2020, 279, 102145. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1995. [Google Scholar] [CrossRef]
- Graebin, C.S.; Verli, H.; Guimarães, J.A. Glycyrrhizin and glycyrrhetic acid: Scaffolds to promising new pharmacologically active compounds. J. Braz. Chem. Soc. 2010, 21, 1595–1615. [Google Scholar] [CrossRef]
- Graebin, C.S. The Pharmacological Activities of Glycyrrhizinic Acid. In Sweeteners: Pharmacology, Biotechnology, and Applications; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–17. [Google Scholar] [CrossRef]
- Cosmetic Ingredient Review Expert Panel. Final Report on the Safety Assessment of Glycyrrhetinic Acid, Potassium Glycyrrhetinate, Disodium Succinoyl Glycyrrhetinate, Glyceryl Glycyrrhetinate, Glycyrrhetinyl Stearate, Stearyl Glycyrrhetinate, Glycyrrhizic Acid, Ammonium Glycyrrhizate, Dipotassium Glycyrrhizate, Disodium Glycyrrhizate, Trisodium Glycyrrhizate, Methyl Glycyrrhizate, and Potassium Glycyrrhizinate. Int. J. Toxicol. 2007, 26, 79–112. [Google Scholar] [CrossRef]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. PTR 2008, 22, 709–724. [Google Scholar] [CrossRef]
- Isbrucker, R.; Burdock, G. Risk and safety assessment on the consumption of Licorice root (Glycyrrhiza sp.), its extract and powder as a food ingredient, with emphasis on the pharmacology and toxicology of glycyrrhizin. Regul. Toxicol. Pharmacol. 2006, 46, 167–192. [Google Scholar] [CrossRef]
- Netala, V.R.; Ghosh, S.B.; Bobbu, P.; Anitha, D.; Tartte, V. Triterpenoid saponins: A review on biosynthesis, applications and mechanism of their action. Int. J. Pharm. Pharm. Sci. 2015, 7, 24–28. [Google Scholar]
- Cheok, C.Y.; Salman, H.A.K.; Sulaiman, R. Extraction and quantification of saponins: A review. Food Res. Int. 2014, 59, 16–40. [Google Scholar] [CrossRef]
- Su, X.; Wu, L.; Hu, M.; Dong, W.; Xu, M.; Zhang, P. Glycyrrhizic acid: A promising carrier material for anticancer therapy. Biomed. Pharmacother. 2017, 95, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef]
- Luo, P.; Liu, D.; Li, J. Pharmacological perspective: Glycyrrhizin may be an efficacious therapeutic agent for COVID-19. Int. J. Antimicrob. Agents 2020, 55, 105995. [Google Scholar] [CrossRef] [PubMed]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of Natural Products as Potential Inhibitors of COVID-19 Main Protease (Mpro): In-Silico Evidences. Nat. Prod. Bioprospect. 2020, 10, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Rossum, T.G.V.; Vulto, A.G.; Hop, W.C.; Brouwer, J.T.; Niesters, H.G.; Schalm, S.W. Intravenous glycyrrhizin for the treatment of chronic hepatitis C: A double-blind, randomized, placebo-controlled phase I/II trial. J. Gastroenterol. Hepatol. 1999, 14, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, R.K.; Chawla, Y.K. Herbal Medicines for Liver Diseases. Dig. Dis. Sci. 2005, 50, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Osaka, S.; Kiwada, H. Targeting of liposomes surface-modified with glycyrrhizinto the liver. 1. Preparation and Biological disposition. Chem. Pharm. Bull. 1991, 39, 1004–1008. [Google Scholar] [CrossRef]
- Barone, A.; Cristiano, M.C.; Cilurzo, F.; Locatelli, M.; Iannotta, D.; Marzio, L.D.; Celia, C.; Paolino, D. Ammonium glycyrrhizate skin delivery from ultradeformable liposomes: A novel use as an anti-inflammatory agent in topical drug delivery. Colloids Surf. B Biointerfaces 2020, 193, 111152. [Google Scholar] [CrossRef]
- Matsuoka, K.; Miyajima, R.; Ishida, Y.; Karasawa, S.; Yoshimura, T. Aggregate formation of glycyrrhizic acid. Colloids Surf. A Physicochem. Eng. Asp. 2016, 500, 112–117. [Google Scholar] [CrossRef]
- Böttcher, S.; Drusch, S. Saponins—Self-assembly and behavior at aqueous interfaces. Adv. Colloid Interface Sci. 2017, 243, 105–113. [Google Scholar] [CrossRef]
- Saha, A.; Adamcik, J.; Bolisetty, S.; Handschin, S.; Mezzenga, R. Fibrillar networks of glycyrrhizic acid for hybrid nanomaterials with catalytic features. Angew. Chem. 2015, 127, 5498–5502. [Google Scholar] [CrossRef]
- Wojciechowski, K.; Orczyk, M.; Gutberlet, T.; Geue, T. Complexation of phospholipids and cholesterol by triterpenic saponins in bulk and in monolayers. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 363–373. [Google Scholar] [CrossRef]
- Wojciechowski, K.; Orczyk, M.; Trapp, M.; Gutberlet, T. Effect of triterpene and steroid saponins on lecithin bilayers. Colloids Surf. A Physicochem. Eng. Asp. 2016, 510, 150–158. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Apanasenko, I.E.; Kim, A.V.; Shelepova, E.A.; Khalikov, S.S.; Polyakov, N.E. Spectroscopic and molecular dynamics characterization of glycyrrhizin membrane-modifying activity. Colloids Surf. B Biointerfaces 2016, 147, 459–466. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Polyakov, N.E.; Korneev, D.V.; Zaitsev, B.N. Influence of glycyrrhizin on permeability and elasticity of cell membrane: Perspectives for drugs delivery. Drug Deliv. 2016, 23, 858–865. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Apanasenko, I.E.; Shilov, A.G.; Khalikov, S.S.; Polyakov, N.E. Effect of natural polysaccharides and oligosaccharides on the permeability of cell membranes. Russ. Chem. Bull. 2017, 66, 129–135. [Google Scholar] [CrossRef]
- Shelepova, E.A.; Kim, A.V.; Voloshin, V.P.; Medvedev, N.N. Intermolecular Voids in Lipid Bilayers in the Presence of Glycyrrhizic Acid. J. Phys. Chem. B 2018, 122, 9938–9946. [Google Scholar] [CrossRef] [PubMed]
- Dargel, C.; Hannappel, Y.; Hellweg, T. Heating-Induced DMPC/Glycyrrhizin Bicelle-to-Vesicle Transition: A X-ray Contrast Variation Study. Biophys. J. 2020, 118, 2411–2425. [Google Scholar] [CrossRef]
- Geisler, R.; Pedersen, M.C.; Hannappel, Y.; Schweins, R.; Prévost, S.; Dattani, R.; Arleth, L.; Hellweg, T. Aescin-induced conversion of gel-phase lipid membranes into bicelle-like lipid nanoparticles. Langmuir 2019, 35, 16244–16255. [Google Scholar] [CrossRef] [PubMed]
- Geisler, R.; Pedersen, M.C.; Preisig, N.; Hannappel, Y.; Prévost, S.; Dattani, R.; Arleth, L.; Hellweg, T. Aescin—A natural soap for the formation of lipid nanodiscs with tunable size. Soft Matter 2021, 17, 1888–1900. [Google Scholar] [CrossRef] [PubMed]
- Pascher, I.; Sundell, S.; Harlos, K.; Eibl, H. Conformation and packing properties of membrane lipids: The crystal structure of sodium dimyristoylphosphatidylglycerol. Biochim. Biophys. Acta (BBA) Biomembr. 1987, 896, 77–88. [Google Scholar] [CrossRef]
- Unitt, M.D.; Harwood, J.L. Sidedness studies of thylakoid phosphatidylglycerol in higher plants. Biochem. J. 1985, 228, 707–711. [Google Scholar] [CrossRef]
- Van Meer, G.; de Kroon, A.I.P.M. Lipid map of the mammalian cell. J. Cell Sci. 2011, 124, 5–8. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112. [Google Scholar] [CrossRef]
- Frentzen, M. Phosphatidylglycerol and sulfoquinovosyldiacylglycerol: Anionic membrane lipids and phosphate regulation. Curr. Opin. Plant Biol. 2004, 7, 270–276. [Google Scholar] [CrossRef]
- Páli, T.; Garab, G.; Horváth, L.I.; Kóta, Z. Functional significance of the lipid-protein interface in photosynthetic membranes. Cell. Mol. Life Sci. CMLS 2003, 60, 1591–1606. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. The essential role of phosphatidylglycerol in photosynthesis. Photosynth. Res. 2007, 92, 205–215. [Google Scholar] [CrossRef]
- Claessens, M.M.A.E.; van Oort, B.F.; Leermakers, F.A.M.; Hoekstra, F.A.; Stuart, M.A.C. Charged lipid vesicles: Effects of salts on bending rigidity, stability, and size. Biophys. J. 2004, 87, 3882–3893. [Google Scholar] [CrossRef] [PubMed]
- Esseling-Ozdoba, A.; Vos, J.W.; van Lammeren, A.A.M.; Emons, A.M.C. Synthetic Lipid (DOPG) Vesicles Accumulate in the Cell Plate Region But Do Not Fuse. Plant Physiol. 2008, 147, 1699–1709. [Google Scholar] [CrossRef] [PubMed]
- Clergeaud, G.; Genç, R.; Ortiz, M.; O’Sullivan, C.K. Liposomal nanoreactors for the synthesis of monodisperse palladium nanoparticles using glycerol. Langmuir 2013, 29, 15405–15413. [Google Scholar] [CrossRef] [PubMed]
- Himeno, H.; Shimokawa, N.; Komura, S.; Andelman, D.; Hamada, T.; Takagi, M. Charge-induced phase separation in lipid membranes. Soft Matter 2014, 10, 7959–7967. [Google Scholar] [CrossRef]
- Findlay, E.J.; Barton, P.G. Phase behavior of synthetic phosphatidylglycerols and binary mixtures with phosphatidylcholines in the presence and absence of calcium ions. Biochemistry 1978, 17, 2400–2405. [Google Scholar] [CrossRef] [PubMed]
- Kučerka, N.; Pencer, J.; Sachs, J.N.; Nagle, J.F.; Katsaras, J. Curvature Effect on the Structure of Phospholipid Bilayers. Langmuir 2007, 23, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Rai, D.K.; Sharma, V.K.; Anunciado, D.; O’Neill, H.; Mamontov, E.; Urban, V.; Heller, W.T.; Qian, S. Neutron Scattering Studies of the Interplay of Amyloid β Peptide (1–40) and An Anionic Lipid 1, 2-dimyristoyl-sn-glycero-3-phosphoglycerol. Sci. Rep. 2016, 6, 30983. [Google Scholar] [CrossRef]
- Elmore, D.E. Molecular dynamics simulation of a phosphatidylglycerol membrane. FEBS Lett. 2006, 580, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Lewis, R.N.A.H.; McElhaney, R.N. Calorimetric and Spectroscopic Studies of the Thermotropic Phase Behavior of the n-Saturated 1,2-Diacylphosphatidylglycerols. Biophys. J. 1997, 72, 779–793. [Google Scholar] [CrossRef]
- Rubinson, K.A. Practical corrections for p(H,D) measurements in mixed H2O/D2O biological buffers. Anal. Methods 2017, 9, 2744–2750. [Google Scholar] [CrossRef]
- Kiselev, M.A.; Lombardo, D. Structural characterization in mixed lipid membrane systems by neutron and X-ray scattering. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 3700–3717. [Google Scholar] [CrossRef]
- Kiselev, M.A.; Zemlyanaya, E.V.; Aswal, V.K.; Neubert, R.H.H. What can we learn about the lipid vesicle structure from the small-angle neutron scattering experiment? Eur. Biophys. J. 2006, 35, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Kučerka, N.; Nieh, M.P.; Katsaras, J. Small-Angle Scattering from Homogenous and Heterogeneous Lipid Bilayers. In Advances in Planar Lipid Bilayers and Liposomes; Academic Press: Oxford, UK, 2010. [Google Scholar] [CrossRef]
- Pan, J.; Heberle, F.A.; Tristram-Nagle, S.; Szymanski, M.; Koepfinger, M.; Katsaras, J.; Kučerka, N. Molecular structures of fluid phase phosphatidylglycerol bilayers as determined by small angle neutron and X-ray scattering. Biochim. Biophys. Acta (BBA) Biomembr. 2012, 1818, 2135–2148. [Google Scholar] [CrossRef]
- Kučerka, N.; Kiselev, M.A.; Balgavý, P. Determination of bilayer thickness and lipid surface area in unilamellar dimyristoylphosphatidylcholine vesicles from small-angle neutron scattering curves: A comparison of evaluation methods. Eur. Biophys. J. 2004, 33, 328–334. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Geisler, P.; Hertle, Y.; Radulescu, A.; Pasini, S.; Perez, J.; Moleiro, L.H.; Hellweg, T. DMPC vesicle structure and dynamics in the presence of low amounts of the saponin aescin. Phys. Chem. Chem. Phys. 2018, 20, 9070–9083. [Google Scholar] [CrossRef]
- Boggara, M.B.; Krishnamoorti, R. Small-angle neutron scattering studies of phospholipid-NSAID adducts. Langmuir 2009, 26, 5734–5745. [Google Scholar] [CrossRef] [PubMed]
- Pencer, J.; Hallett, F.R. Small-angle neutron scattering from large unilamellar vesicles: An improved method for membrane thickness determination. Phys. Rev. E 2000, 61, 3003–3008. [Google Scholar] [CrossRef] [PubMed]
- Sreij, R.; Dargel, C.; Hannappel, Y.; Jestin, J.; Prévost, S.; Dattani, R.; Wrede, O.; Hellweg, T. Temperature dependent self-organization of DMPC membranes promoted by intermediate amounts of the saponin aescin. Biochim. Biophys. Acta (BBA) Biomembr. 2019, 1861, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Doucet, M.; Cho, J.H.; Alina, G.; Bakker, J.; Bouwman, W.; Butler, P.; Campbell, K.; Gonzales, M.; Heenan, R.; Jackson, A.; et al. SasView version 4.2.2. 2019. Available online: www.sasview.org/2019-05-20-release-4.2.2/ (accessed on 16 August 2021).
- Rennie, A.R.; Hellsing, M.S.; Wood, K.; Gilbert, E.P.; Porcar, L.; Schweins, R.; Dewhurst, C.D.; Lindner, P.; Heenan, R.K.; Rogers, S.E.; et al. Learning about SANS instruments and data reduction from round robin measurements on samples of polystyrene latex. J. Appl. Crystallogr. 2013, 46, 1289–1297. [Google Scholar] [CrossRef]
- Viguier, G.; Girardot, R.; Perez, J. Foxtrot 3.3.4. 2016. Available online: www.synchrotron-soleil.fr/en/beamlines/swing (accessed on 16 August 2021).
- Zhang, F.; Ilavsky, J.; Long, G.G.; Quintana, J.P.G.; Allen, A.J.; Jemian, P.R. Glassy Carbon as an Absolute Intensity Calibration Standard for Small-Angle Scattering. Metall. Mater. Trans. A 2010, 41, 1151–1158. [Google Scholar] [CrossRef]
- Sztucki, M. SAXSutilities—On-Line Processing and Analysis of SAXS Data. 2018. Available online: www.sztucki.de/ (accessed on 16 August 2021).
- Zantl, R.; Artzner, F.; Rapp, G.; Rädler, J. Thermotropic structural changes of saturated-cationic-lipid–DNA complexes. EPL Europhys. Lett. 1999, 45, 90. [Google Scholar] [CrossRef][Green Version]
- Pedersen, J.S.; Egelhaaf, S.U.; Schurtenberger, P. Formation of polymerlike mixed micelles and vesicles in lecithin-bile salt solutions: A small-angle neutron-scattering study. J. Phys. Chem. 1995, 99, 1299–1305. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Moleiro, L.H.; Monroy, F.; Hellweg, T. Aescin Incorporation and Nanodomain Formation in DMPC Model Membranes. Langmuir 2017, 33, 12351–12361. [Google Scholar] [CrossRef]
- Geisler, R.; Prévost, S.; Dattani, R.; Hellweg, T. Effect of Cholesterol and Ibuprofen on DMPC-β-Aescin Bicelles: A Temperature-Dependent Wide-Angle X-ray Scattering Study. Crystals 2020, 10, 401. [Google Scholar] [CrossRef]
- Caracciolo, G.; Pozzi, D.; Caminiti, R.; Marchini, C.; Montani, M.; Amici, A.; Amenitsch, H. Transfection efficiency boost by designer multicomponent lipoplexes. Biochim. Biophys. Acta (BBA) Biomembr. 2007, 1768, 2280–2292. [Google Scholar] [CrossRef] [PubMed]
- Selyutina, O.Y.; Polyakov, N.E. Glycyrrhizic acid as a multifunctional drug carrier–from physicochemical properties to biomedical applications: A modern insight on the ancient drug. Int. J. Pharm. 2019, 559, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Sreij, R.; Prévost, S.; Dargel, C.; Dattani, R.; Hertle, Y.; Wrede, O.; Hellweg, T. Interaction of the saponin aescin with ibuprofen in DMPC model membranes. Mol. Pharm. 2018, 15, 4446–4461. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Schweins, R.; Prévost, S.; Dattani, R.; Hellweg, T. Aescin-Cholesterol Complexes in DMPC Model Membranes: A DSC and Temperature-Dependent Scattering Study. Sci. Rep. 2019, 9, 5542. [Google Scholar] [CrossRef]
- Kučerka, N.; Holland, B.W.; Gray, C.G.; Tomberli, B.; Katsaras, J. Scattering Density Profile Model of POPG Bilayers As Determined by Molecular Dynamics Simulations and Small-Angle Neutron and X-ray Scattering Experiments. J. Phys. Chem. B 2012, 116, 232–239. [Google Scholar] [CrossRef] [PubMed]
- ACD/Labs: ChemSketch; Freeware Version; Advanced Chemistry Development, Inc., Toronto, ON, Canada. 2015. Available online: www.acdlabs.com/resources/freeware/chemsketch/ (accessed on 16 August 2021).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| /mol% | /Å | /Å |
|---|---|---|
| 0 | 4.50 ± 0.05 | 4.62 ± 0.04 |
| 1 | 4.51 ± 0.05 | 4.61 ± 0.03 |
| 5 | 4.52 ± 0.04 | 4.60 ± 0.03 |
| 10 | 4.50 ± 0.04 | 4.60 ± 0.03 |
| 20 | 4.47 ± 0.05 | 4.61 ± 0.04 |
| 30 | 4.50 ± 0.04 | 4.64 ± 0.03 |
| 40 | 4.52 ± 0.05 | 4.66 ± 0.03 |
| 50 | 4.53 ± 0.05 | 4.64 ± 0.03 |
| /mol% | /Å | /% |
|---|---|---|
| 0 | 207 ± 2 | 37 |
| 1 | 208 ± 3 | 39 |
| 3 | 203 ± 3 | 42 |
| 10 | 202 ± 3 | 45 |
| 20 | 205 ± 3 | 48 |
| 30 | 176 ± 3 | 55 |
| 40 | 172 ± 3 | 61 |
| 50 | 156 ± 2 | 64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dargel, C.; Gräbitz-Bräuer, F.; Geisler, R.; Fandrich, P.; Hannappel, Y.; Porcar, L.; Hellweg, T. Stable DOPG/Glycyrrhizin Vesicles with a Wide Range of Mixing Ratios: Structure and Stability as Seen by Scattering Experiments and Cryo-TEM. Molecules 2021, 26, 4959. https://doi.org/10.3390/molecules26164959
Dargel C, Gräbitz-Bräuer F, Geisler R, Fandrich P, Hannappel Y, Porcar L, Hellweg T. Stable DOPG/Glycyrrhizin Vesicles with a Wide Range of Mixing Ratios: Structure and Stability as Seen by Scattering Experiments and Cryo-TEM. Molecules. 2021; 26(16):4959. https://doi.org/10.3390/molecules26164959
Chicago/Turabian StyleDargel, Carina, Friederike Gräbitz-Bräuer, Ramsia Geisler, Pascal Fandrich, Yvonne Hannappel, Lionel Porcar, and Thomas Hellweg. 2021. "Stable DOPG/Glycyrrhizin Vesicles with a Wide Range of Mixing Ratios: Structure and Stability as Seen by Scattering Experiments and Cryo-TEM" Molecules 26, no. 16: 4959. https://doi.org/10.3390/molecules26164959
APA StyleDargel, C., Gräbitz-Bräuer, F., Geisler, R., Fandrich, P., Hannappel, Y., Porcar, L., & Hellweg, T. (2021). Stable DOPG/Glycyrrhizin Vesicles with a Wide Range of Mixing Ratios: Structure and Stability as Seen by Scattering Experiments and Cryo-TEM. Molecules, 26(16), 4959. https://doi.org/10.3390/molecules26164959