
Impact of Different Biochars on Microbial Community Structure in the Rhizospheric Soil of Rice Grown in Albic Soil

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Influence of Biochar on the Nutrient Content of Albic Soil
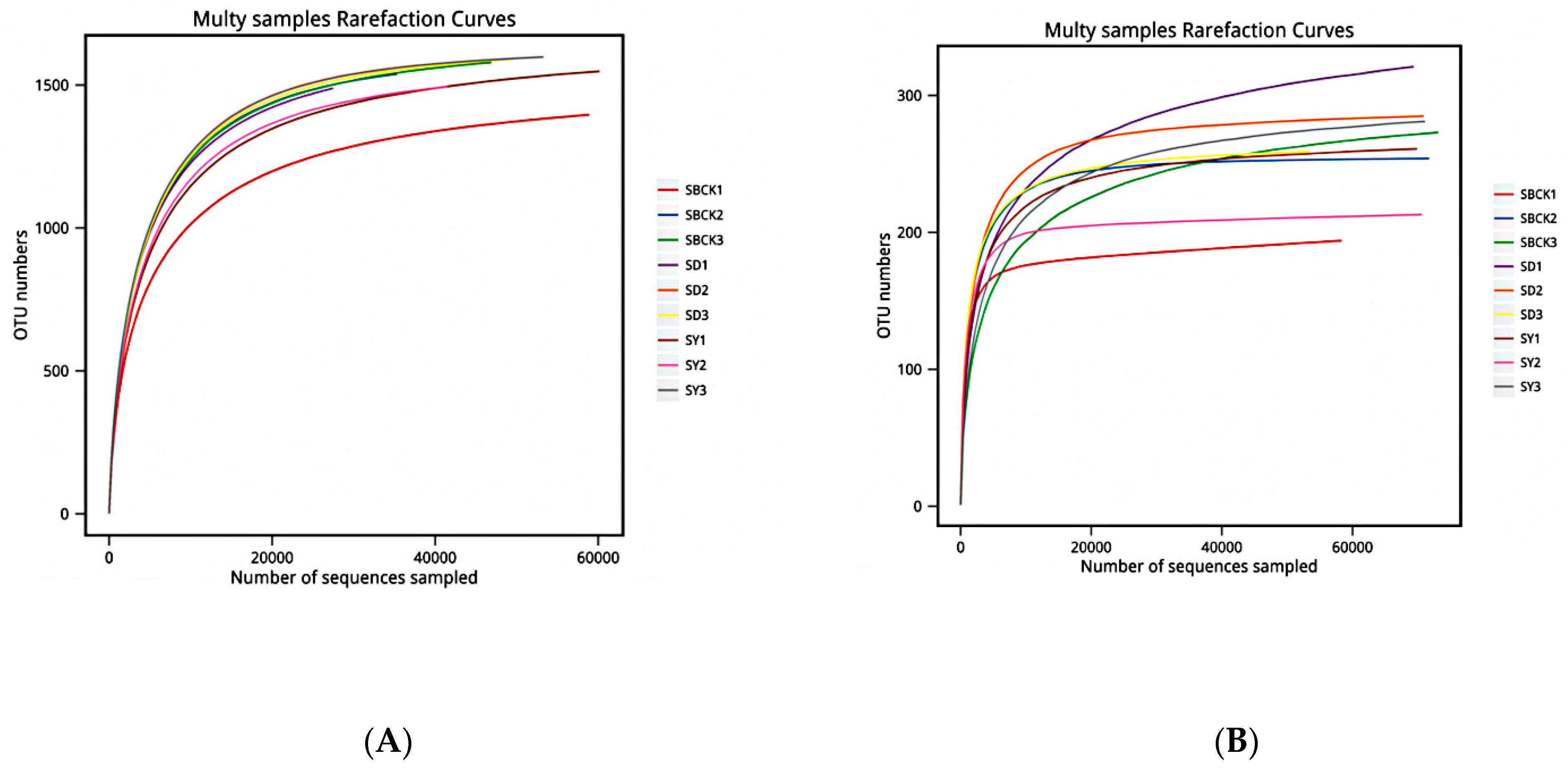
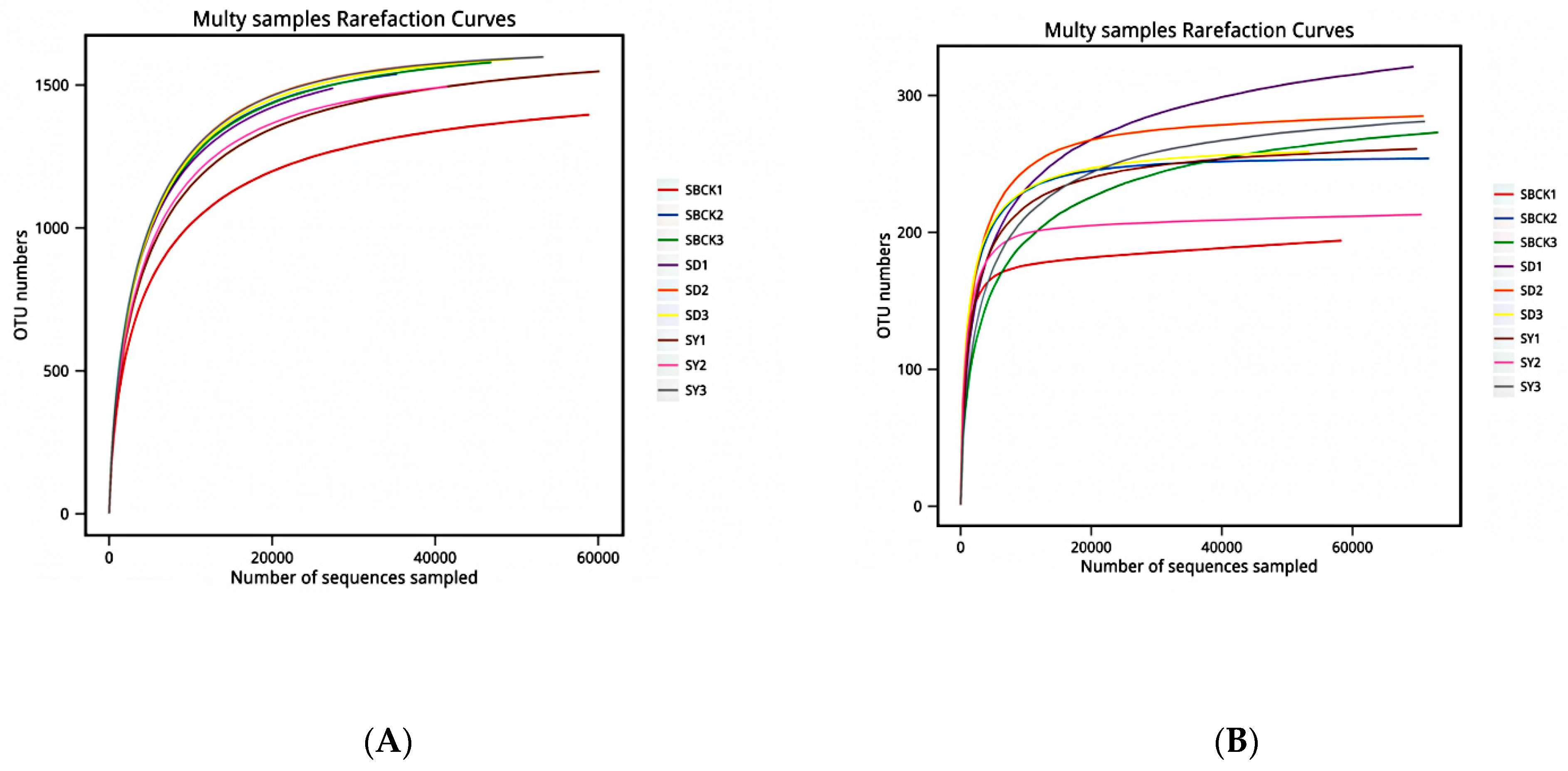
2.2. Depth Assessment of Soil Sample Sequencing
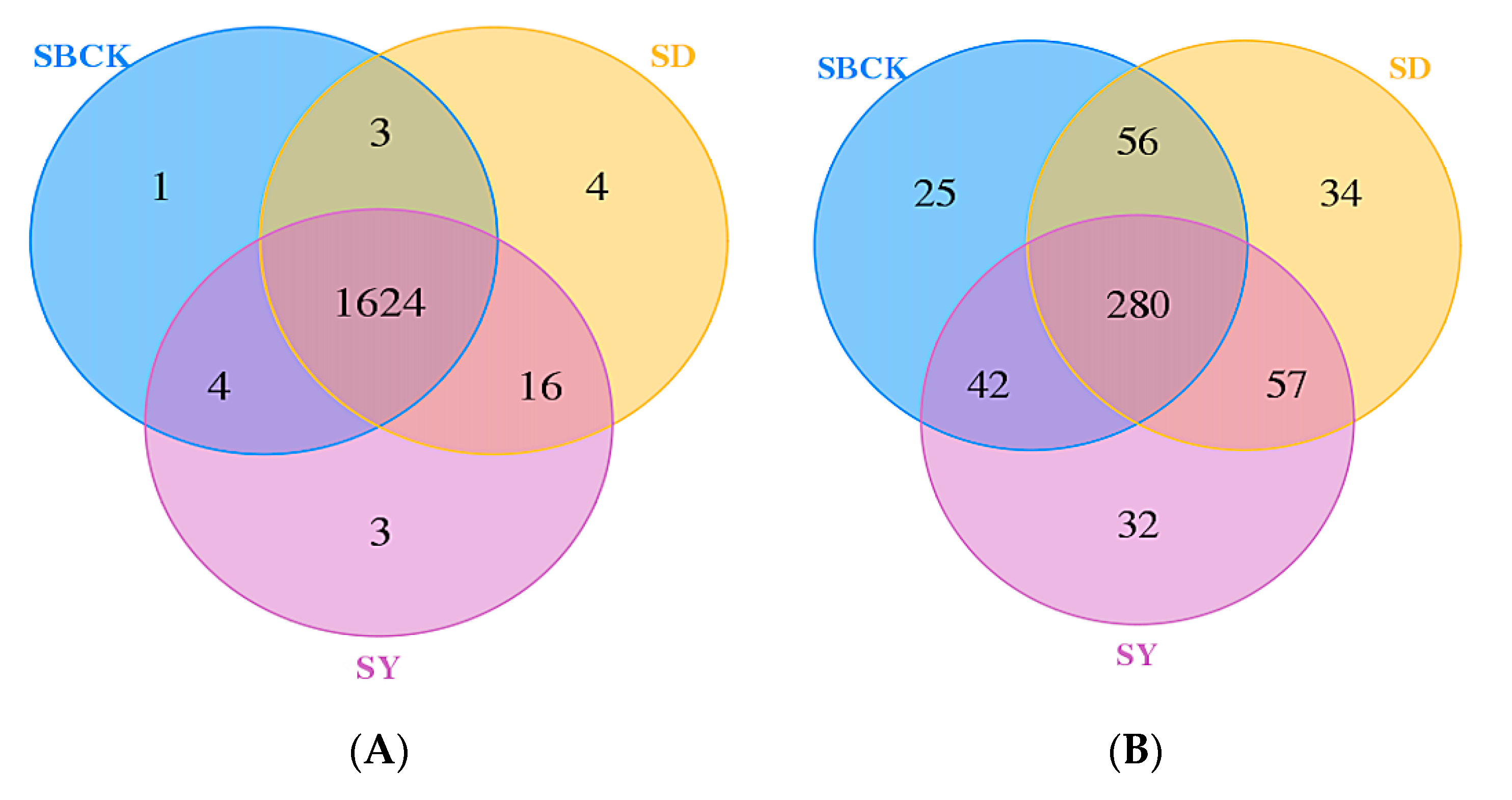
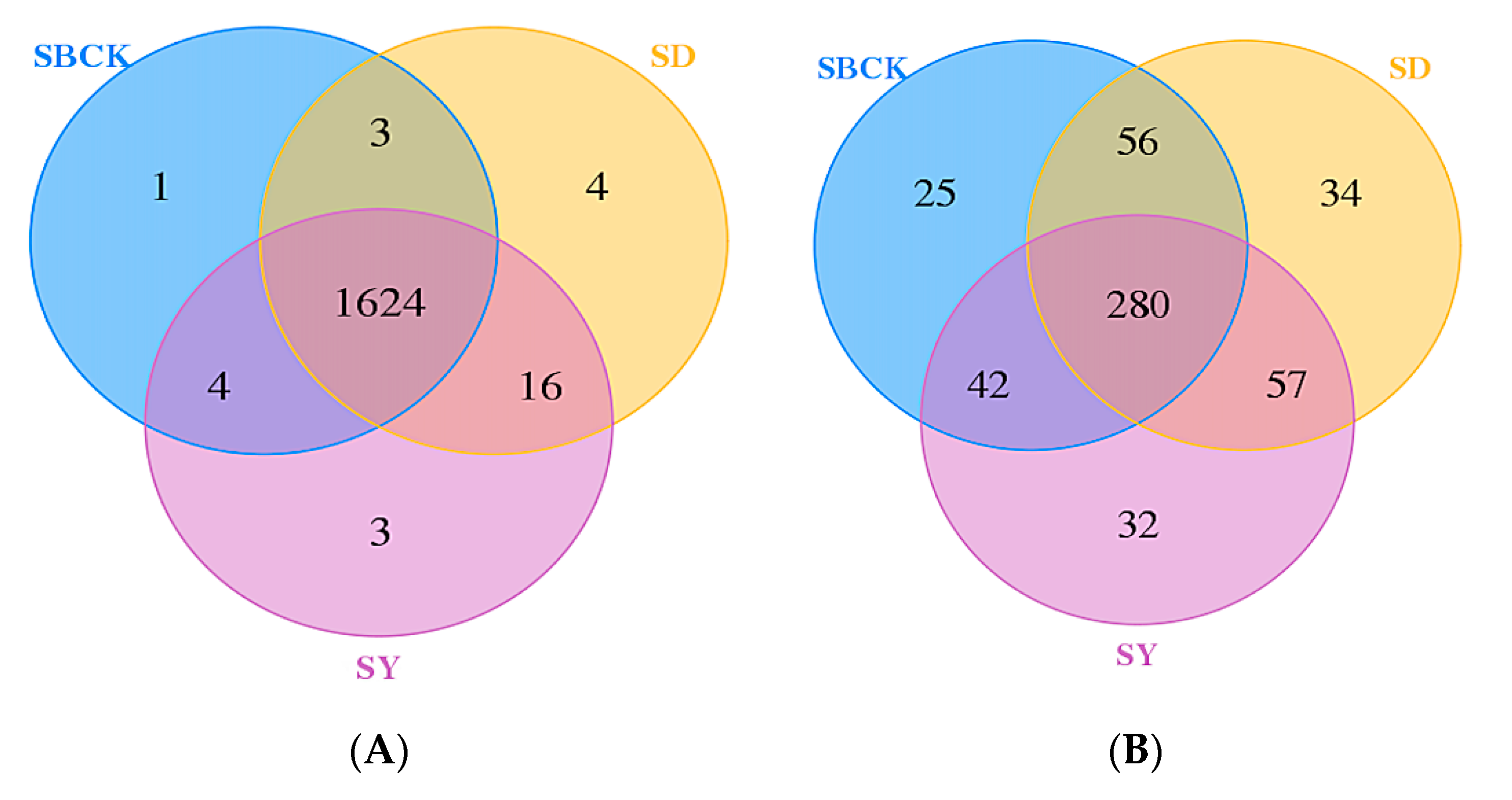
2.3. Effects of Biochar on the Abundance and Diversity of Soil Bacterial and Fungal Communities
2.4. Effects of Biochar on the Relative Abundance of Bacteria and Fungi in Root Zone Soil
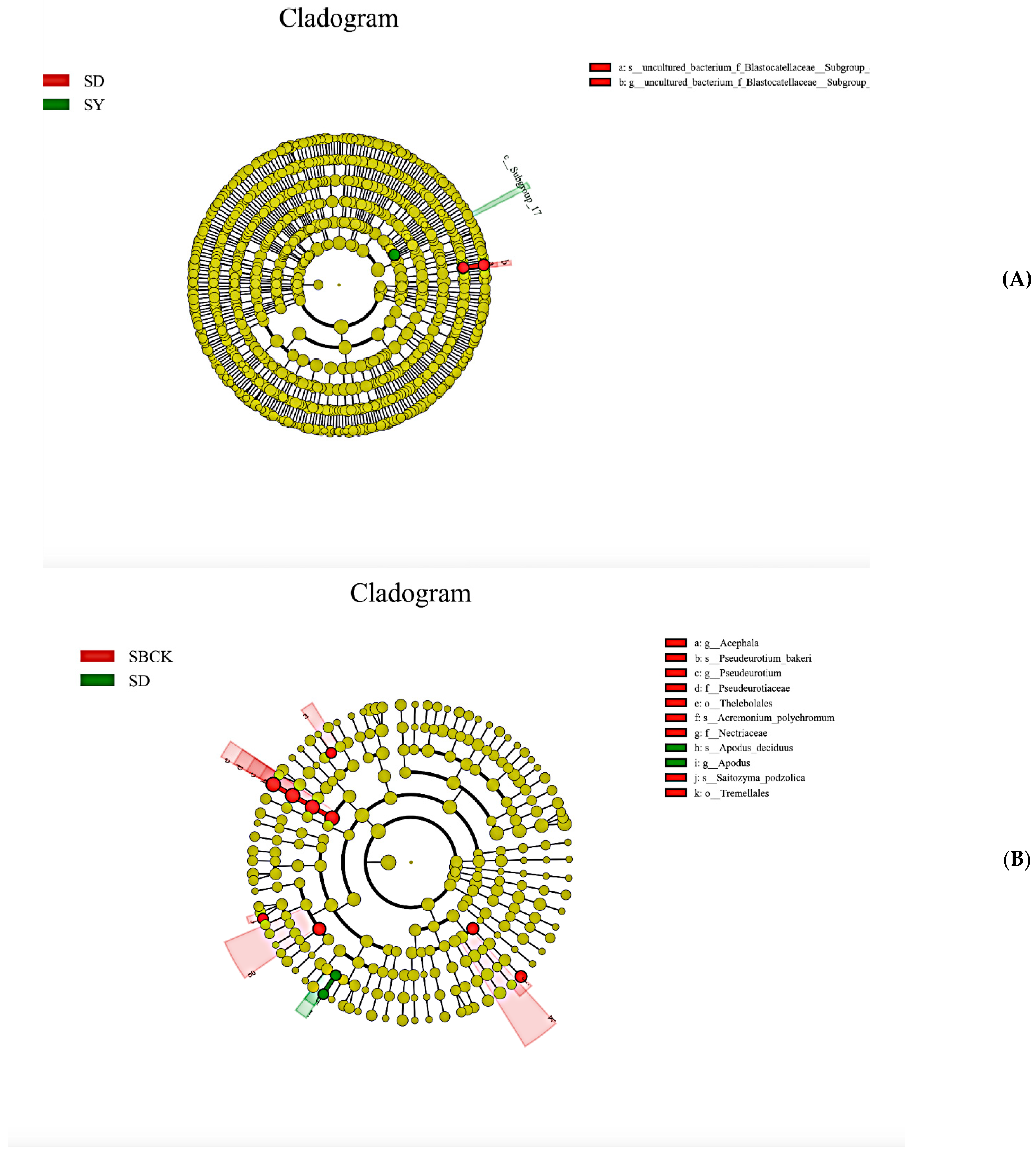
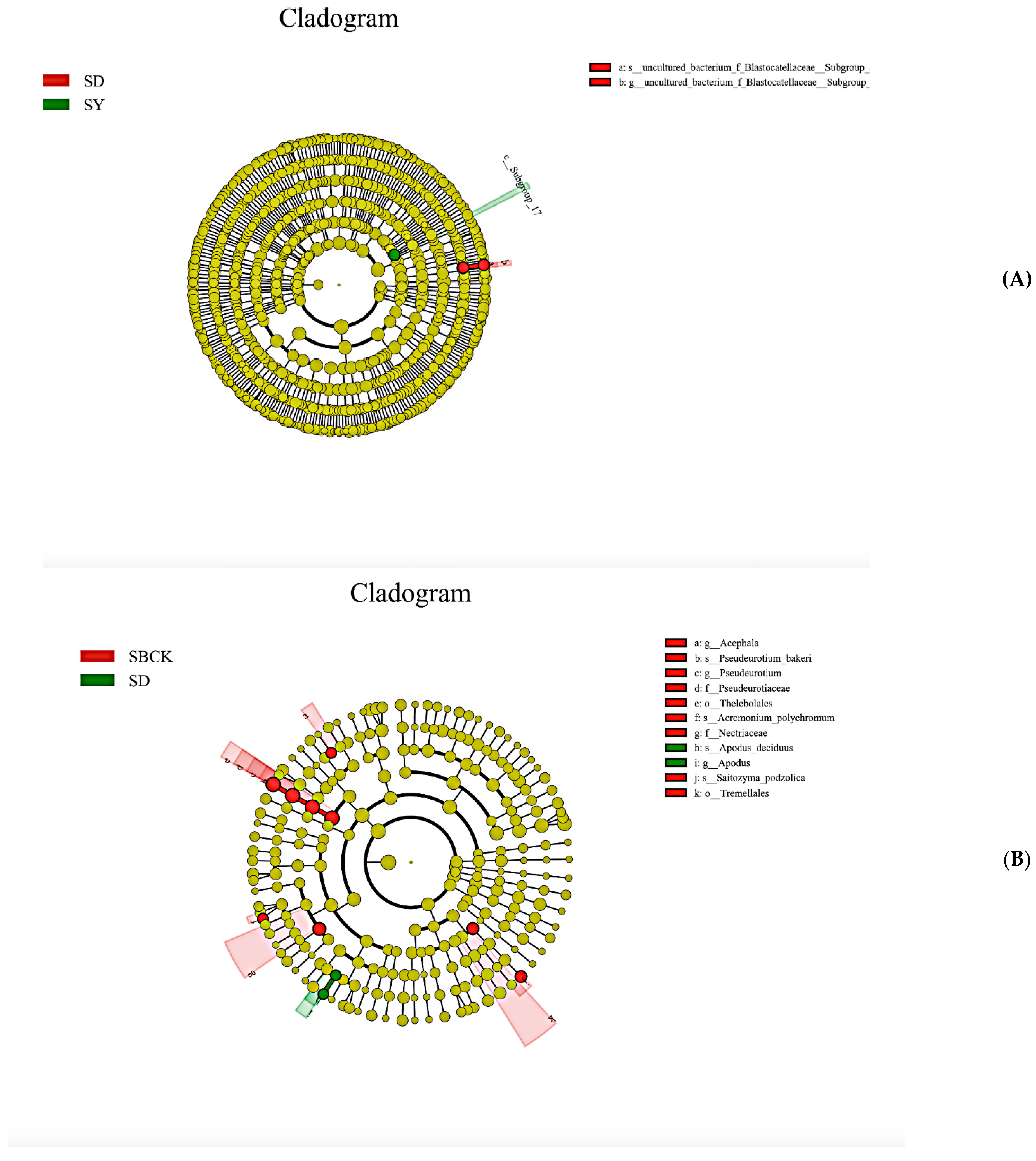
2.5. Analysis of Soil Bacteria and Fungi Groups in the Context of Biochar Treatment
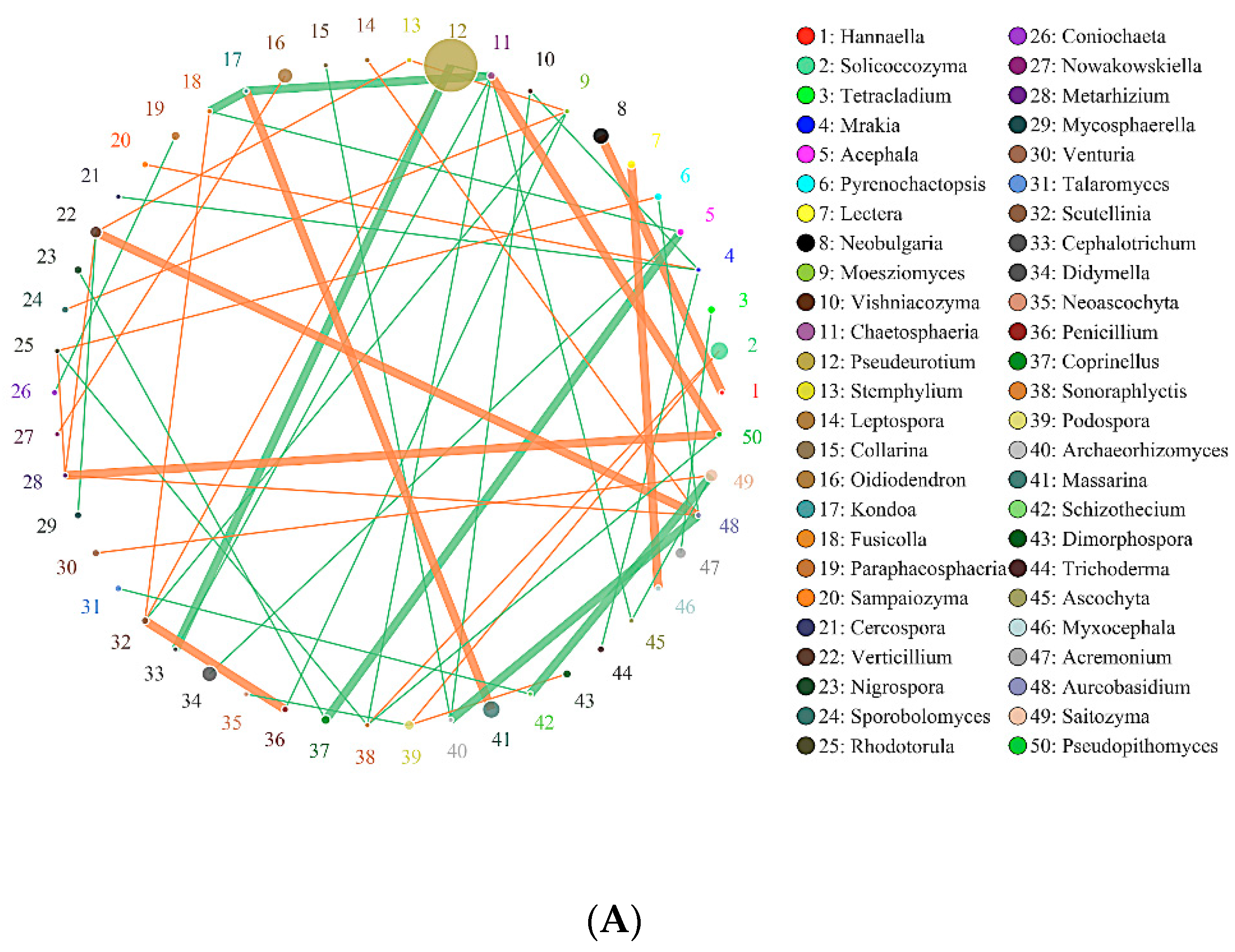
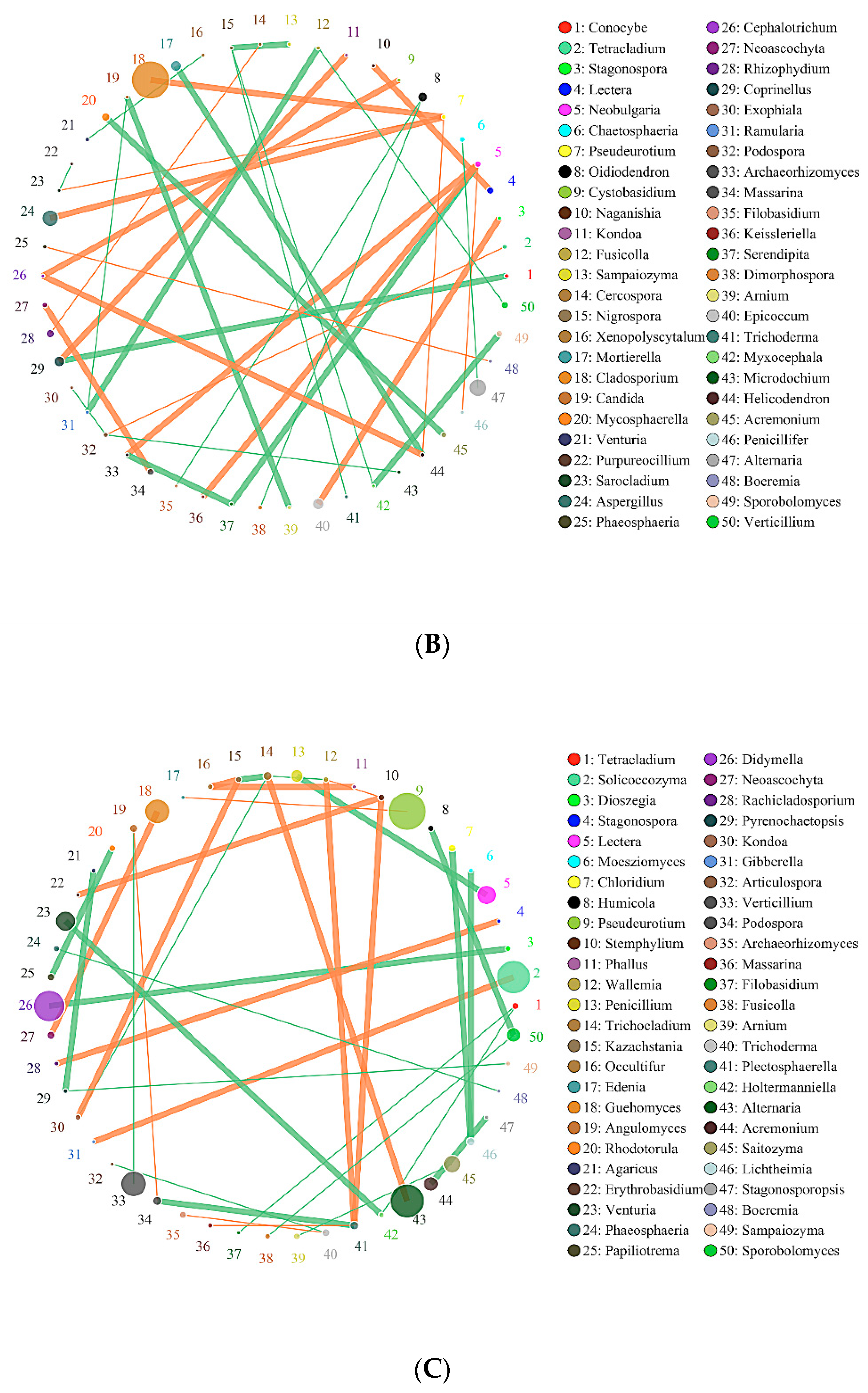
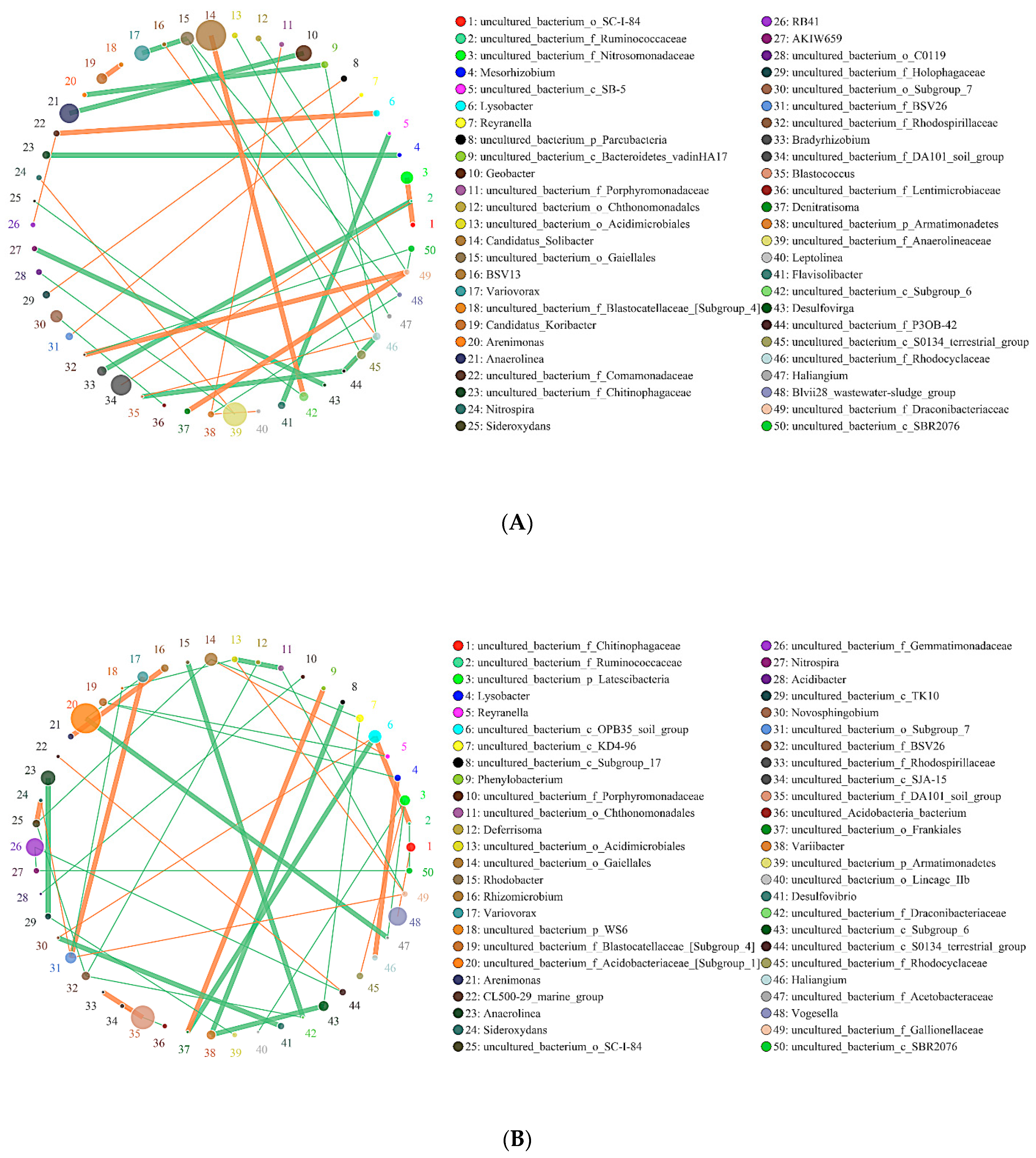
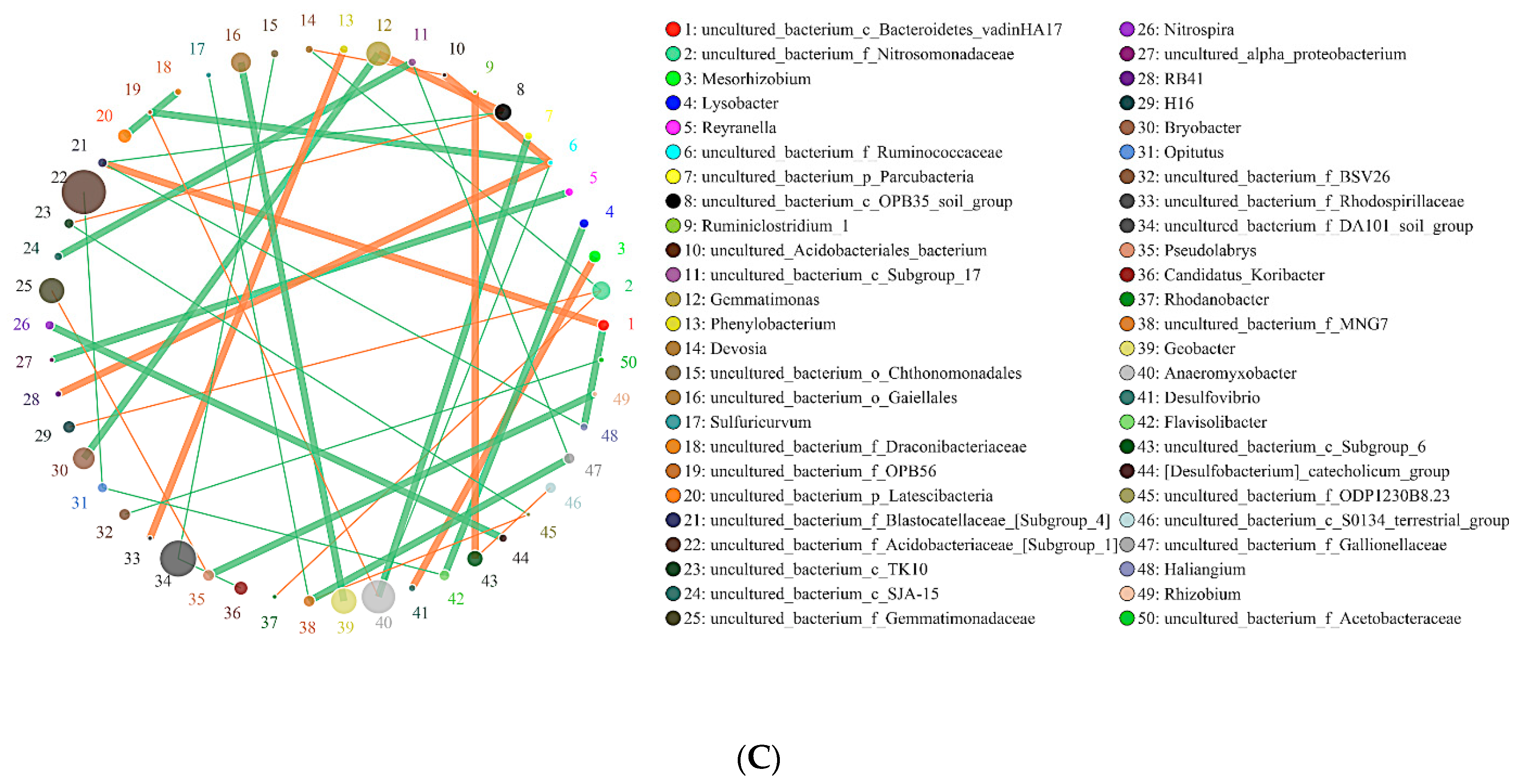
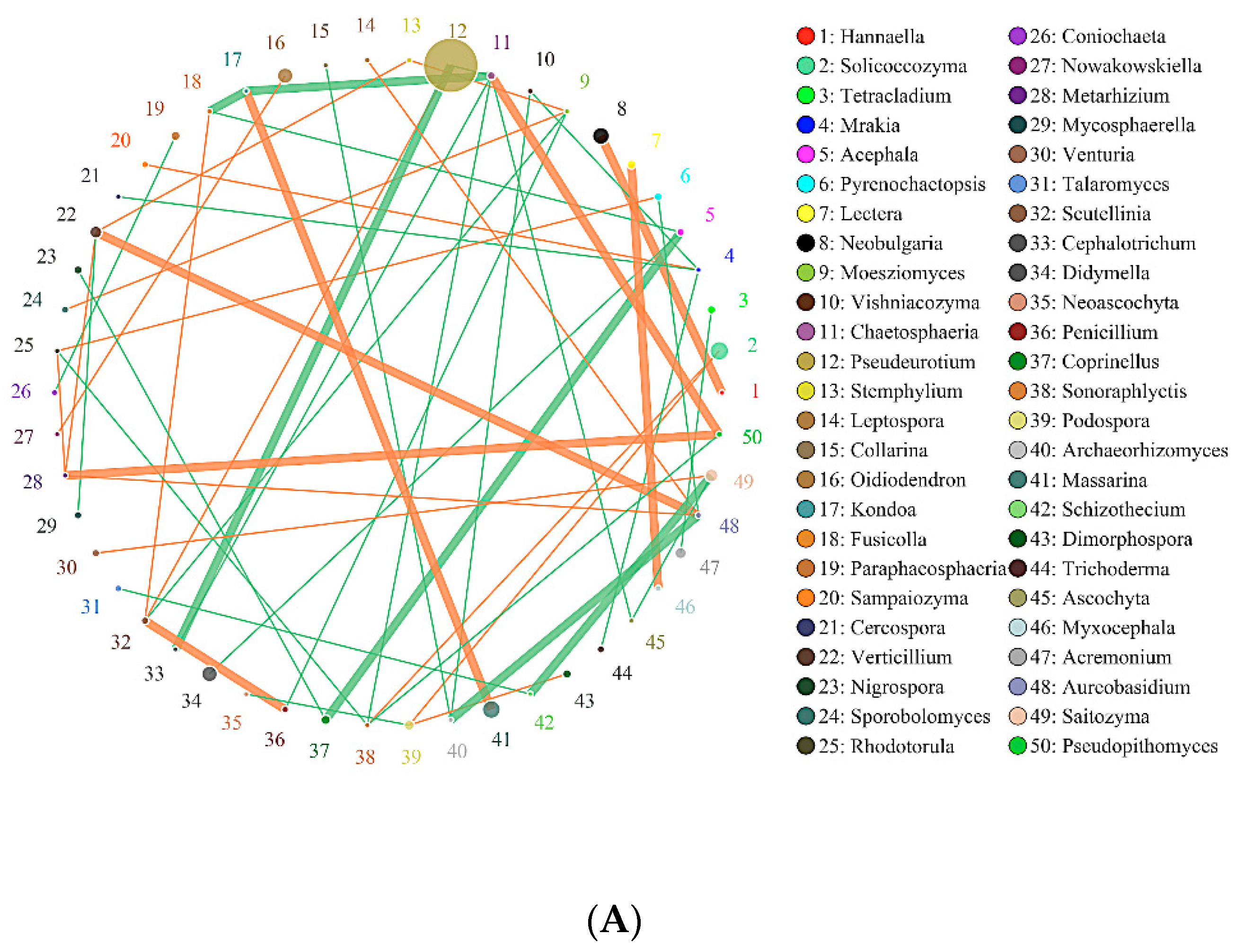
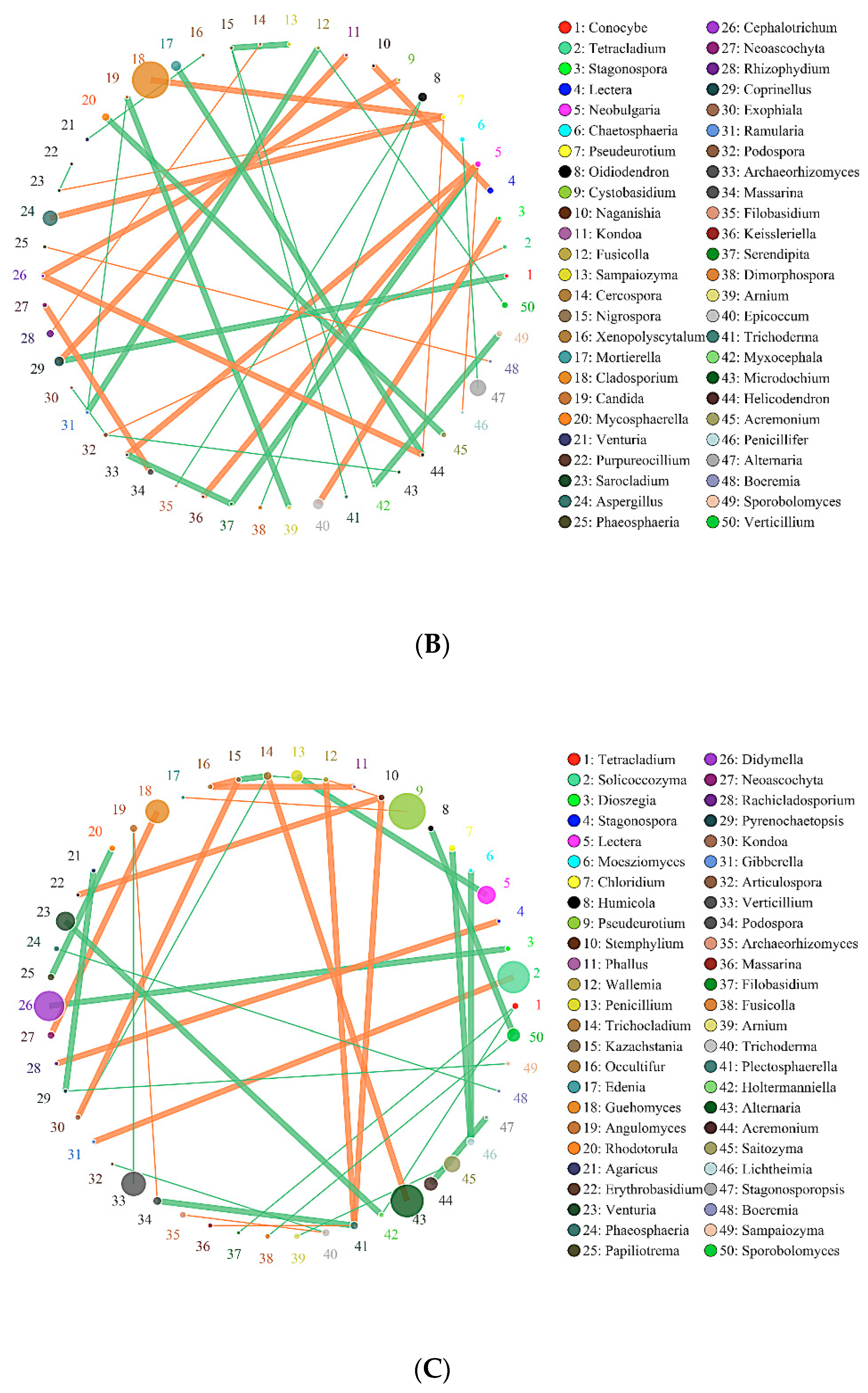
2.6. Effects of Biochar on the Interaction of Soil Bacteria and Fungi
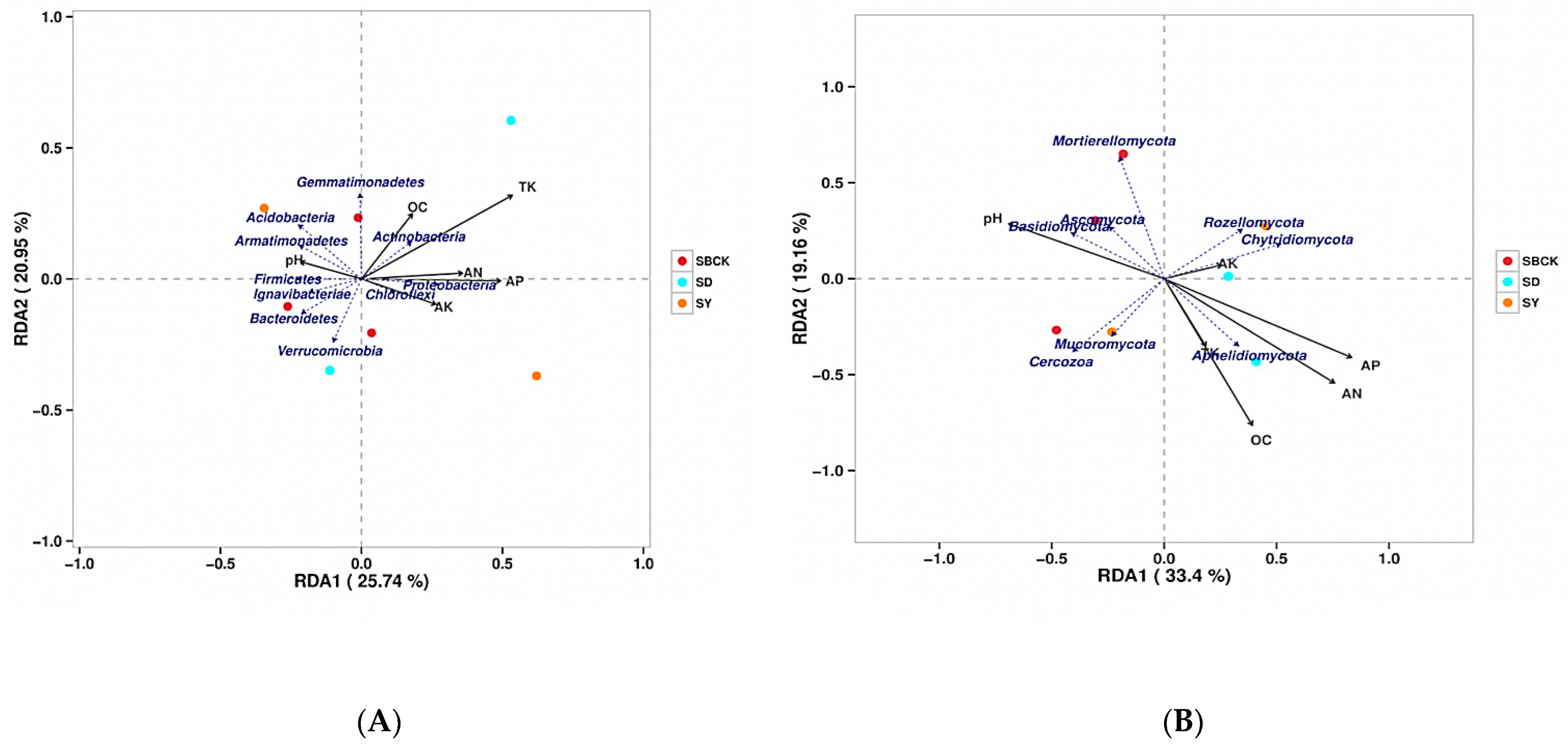
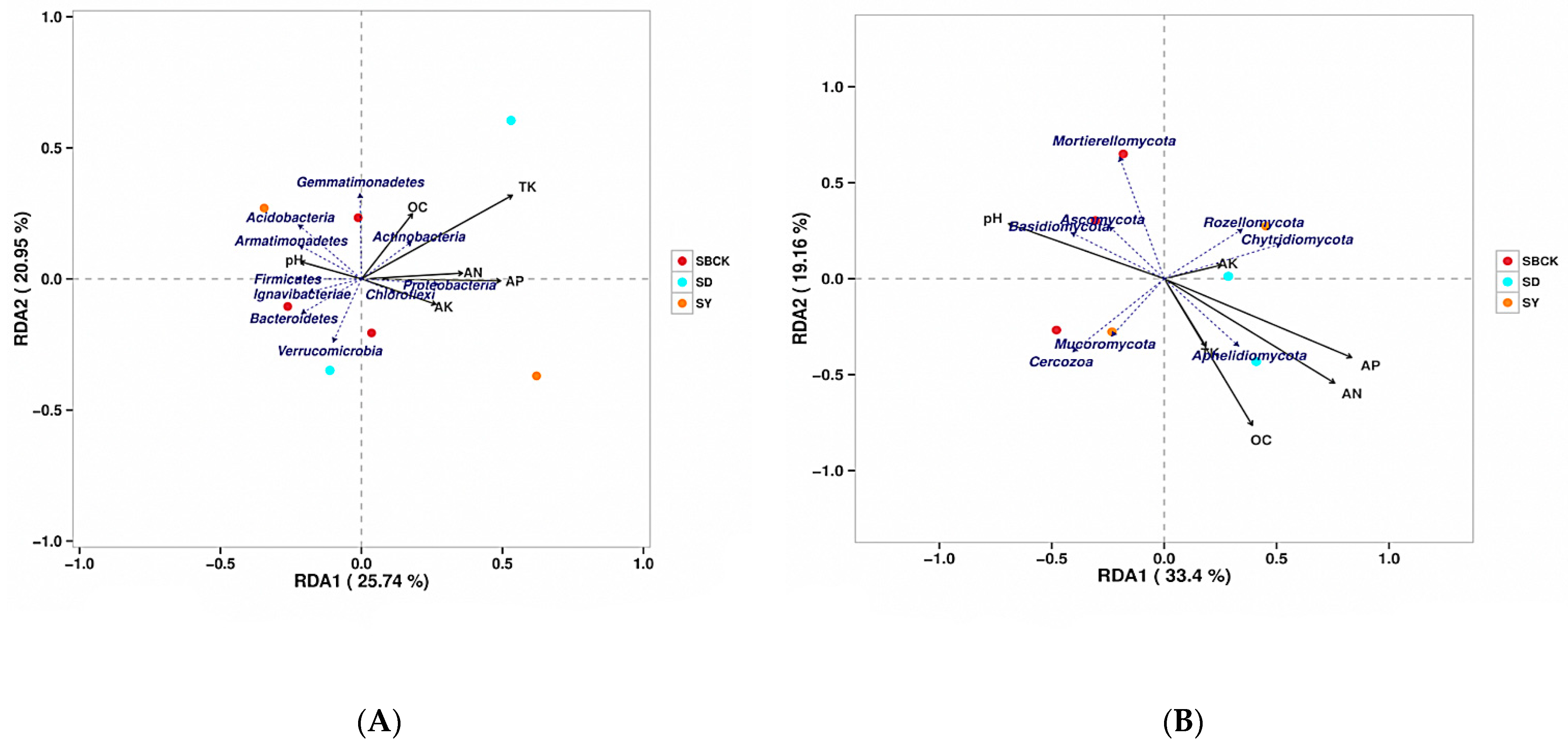
2.7. Correlation Analysis of Soil Microbial Community and Nutrients
3. Materials and Methods
3.1. Test Site
3.2. Test Materials
3.3. Experimental Design
3.4. Sample Collection
3.5. Test Methods
3.6. Data Processing and Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wang, X.H. The characteristic and influence factors of extinction depth of shallow groundwater on the high-latitude region: A case study on the Sanjiang Plain, northeast China. Environ. Sci. Pollut. Res. 2018, 25, 6695–6706. [Google Scholar] [CrossRef]
- Xiu, L.; Zhang, W.; Sun, Y.; Wu, D.; Meng, J.; Chen, W. Effects of biochar and straw returning on the key cultivation limitations of Albic soil and soybean growth over 2 years. Catena 2019, 173, 481–493. [Google Scholar] [CrossRef]
- Denef, K.; Roobroeck, D.; Manimel Wadu, M.C.W.; Lootens, P.; Boeckx, P. Microbial community composition and rhizodeposit-carbon assimilation in differently managed temperate grassland soils. Soil Biol. Biochem. 2009, 41, 144–153. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Hewedy, O.A.; Abdel Lateif, K.S.; Seleiman, M.F.; Shami, A.; Albarakaty, F.M.; El-Meihy, M.R. Phylogenetic diversity of trichoderma strains and their antagonistic potential against soil-borne pathogens under stress conditions. Biology 2020, 9, 189. [Google Scholar] [CrossRef]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Vojvodic-Vukovic, M. Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil Biol. Biochem. 2000, 32, 189–196. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Olanrewaju, O.S.; Babalola, O.O. Plant growth promoting rhizobacterial mitigation of drought stress in crop plants: Implications for sustainable agriculture. Agronomy 2019, 9, 712. [Google Scholar] [CrossRef] [Green Version]
- Sarfraz, R.; Hussain, A.; Sabir, A.; Fekih, I.B.; Ditta, A.; Xing, S.H. Role of biochar and plant growth promoting rhizobacteria to enhance soil carbon sequestration—A review. Environ. Monit. Assess. 2019, 191, 251. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M.; Mohsin, F.; Hussain, M. ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar-Ul-Hye, M.; Danish, S.; Abbas, M.; Ahmad, M.; Munir, T.M. ACC deaminase producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with biochar improve wheat productivity under drought stress. Agronomy 2019, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Li, D.; Wang, M.; Gong, T.C.; Sun, M.Y.; Yang, T.X. A scientometric review of biochar preparation research from 2006 to 2019. Biochar 2021, 3, 283–298. [Google Scholar] [CrossRef]
- Pranagal, J.; Oleszczuk, P.; Tomaszewska-Krojańska, D.; Kraska, P.; Różyło, K. Effect of biochar application on the physical properties of Haplic Podzol. Soil. Tillage Res. 2017, 174, 92–103. [Google Scholar] [CrossRef]
- Rizwan, M.S.; Imtiaz, M.; Zhu, J.; Yousaf, B.; Hu, H. Immobilization of pb and cu by organic and inorganic amendments in contaminated soil. Geoderma 2021, 385, 114803. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Battaglia, M.L.; Shami, A.; Jalal, R.S.; Alhammad, B.A.; Almutairi, K.F.; Al-Saif, A.M. Biochar and its broad impacts in soil quality and fertility, nutrient leaching and crop productivity: A review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Alotaibi, M.A.; Alhammad, B.A.; Alharbi, B.M.; Refay, Y.; Badawy, S.A. Effects of ZnO nanoparticles and biochar of rice straw and cow manure on characteristics of contaminated soil and sunflower productivity, oil quality, and heavy metals uptake. Agronomy 2020, 10, 790. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M. Integrative effects of rice-straw biochar and silicon on oil and seed quality, yield and physiological traits of Helianthus annuus L. grown under water deficit stress. Agronomy 2019, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Ullah, N.; Ditta, A.; Imtiaz, M.; Li, X.M.; Jan, A.U.; Mehmood, S.; Rizwan, M.S.; Rizwan, M. Appraisal for organic amendments and plant growth-promoting rhizobacteria to enhance crop productivity under drought stress: A review. J. Agron. Crop. Sci. 2021. [Google Scholar] [CrossRef]
- Li, M.Y.; Sun, W.J.; Wang, Y.J.; Sun, D.A.; Tan, Y.Z. Air permeability of biochar-amended clay cover. Arab. J. Geosci. 2021, 14, 732. [Google Scholar] [CrossRef]
- Dai, Z.; Xiong, X.; Zhu, H.; Xu, H.; Xu, J. Association of biochar properties with changes in soil bacterial, fungal and fauna communities and nutrient cycling processes. Biochar 2021, 3, 239–254. [Google Scholar] [CrossRef]
- Murtaza, G.; Ditta, A.; Ullah, N.; Usman, M.; Ahmed, Z. Biochar for the management of nutrient impoverished and metal contaminated soils: Preparation, applications, and prospects. J. Soil. Sci. Plant. Nut. 2021, 1–23. [Google Scholar] [CrossRef]
- Maienza, A.; Baronti, S.; Cincinelli, A.; Martellini, T.; Grisolia, A.; Miglietta, F.; Renella, G.; Stazi, S.R.; Vaccari, F.P.; Genesio, L. Biochar improves the fertility of a Mediterranean vineyard without toxic impact on the microbial community. Agron. Sustain. Dev. 2017, 37, 47. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Sun, X.; Zheng, J.F.; Liu, X.; Bian, R.; Li, L.; Cheng, K.; Zheng, J.; Pan, G. Biochar amendment changes temperature sensitivity of soil respiration and composition of microbial communities 3 years after incorporation in an organic carbon-poor dry cropland soil. Biol. Fert. Soils 2018, 54, 175–188. [Google Scholar] [CrossRef]
- Zhao, B.; Niebuhr, A.J.; Lv, Y.; Douangdalangsy, K. Effects of soybean stover-derived biochar on microbial community and structure in loess soil. Nat. Environ. Pollut. Technol. 2020, 19, 703–710. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, C.; Chen, G.; Zhou, J.L.; Chen, Z.; Li, Z.X.; Zhu, J.W.; Feng, T.; Chen, Y.Q. Response of soil microbial communities to additions of straw biochar, iron oxide, and iron oxide-modified straw biochar in an arsenic-contaminated soil. Environ. Sci. Pollut. Res. 2020, 27, 23761–23768. [Google Scholar] [CrossRef]
- Roy, R.; Núez-Delgado, A.; Sultana, S.; Wang, J.; Mmunirf, A.; Battaglia, M.; Sarker, T.; Seleiman, M.F.; Barmon, M.; Zhang, R.Q. Additions of optimum water, spent mushroom compost and wood biochar to improve the growth performance of althaea rosea in drought-prone coal-mined spoils. J. Environ. Manag. 2021, 295, 113076. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K. Biochemical cycling of nitrogen and phosphorus in biochar amended soils. Soil Biol. Biochem. 2016, 103, 1–15. [Google Scholar] [CrossRef]
- Luo, X.X.; Liu, G.C.; Xia, Y.; Chen, L.; Jiang, Z.X.; Zheng, H.; Wang, Z.Y. Use of biocharcompost to improve properties and productivity of the degraded coastal soil in the Yellow River Delta, China. J. Soils Sediments 2017, 17, 780–789. [Google Scholar] [CrossRef]
- Sun, H.J.; Lu, H.Y.; Chu, L.; Shao, H.B.; Shi, W.M. Biochar applied with appropriate rates can reduce N leaching, keep N retention and not increase NH3 volatilization in a coastal saline soil. Sci. Total. Environ. 2017, 575, 820–825. [Google Scholar] [CrossRef]
- Agegnehu, G.; Srivastava, A.K.; Bird, M.I. The role of biochar and biochar-compost in improving soil quality and crop performance: A review. Appl. Soil Ecol. 2017, 119, 156–170. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zeng, G.; Zhou, L.; Zheng, B. Biochar to improve soil fertility. A review. Agron. Sustain. Dev. 2016, 36, 35–52. [Google Scholar] [CrossRef] [Green Version]
- El-Naggar, A.; Lee, S.S.; Rinklebe, J.; Farooq, M.; Song, H.; Sarmah, A.K.; Ok, Y.S. Biochar application to low fertility soils: A review of current status, and future prospects. Geoderma 2019, 337, 536–554. [Google Scholar] [CrossRef]
- Cui, Q.; Xia, J.; Yang, H.; Liu, J.; Shao, P. Biochar and effective microorganisms promote Sesbania cannabina growth and soil quality in the coastal saline-alkali soil of the Yellow River Delta, China. Sci. Total. Environ. 2021, 756, 143801. [Google Scholar] [CrossRef]
- Li, S.L.; Wang, S.; Fan, M.C.; Wu, Y.; Shangguan, Z.P. Interactions between biochar and nitrogen impact soil carbon mineralization and the microbial community. Soil. Tillage Res. 2020, 196, 104437. [Google Scholar] [CrossRef]
- Lucheta, A.R.; Cannavan, F.D.S.; Roesch, L.; Tsai, S.M.; Kuramae, E.E. Fungal community assembly in the amazonian dark Earth. Microb. Ecol. 2016, 71, 962–973. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Cao, L.X.; Zhang, R.D. Bacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soil. World J. Microb. Biotechnol. 2014, 30, 1085–1092. [Google Scholar] [CrossRef]
- Liu, G.L. Effects of Transgenic Soybeans on the Rhizospheric Soil Microbial Communities and Their Diversity Indices Measurement. Ph.D. Thesis, Nanjing University, Nanjing, China, 2012. (In Chinese). [Google Scholar]
- Muhammad, N.; Dai, Z.; Xiao, K.; Meng, J.; Brookes, P.C.; Liu, X.; Wang, H.; Wu, J.; Xu, J. Changes in microbial community structure due to biochars generated from different feedstocks and their relationships with soil chemical properties. Geoderma 2014, 226–227, 270–278. [Google Scholar] [CrossRef]
- Yao, Q.; Liu, J.; Yu, Z.; Li, Y.; Jin, J.; Liu, X.; Wang, G. Changes of bacterial community compositions after three years of biochar application in a black soil of northeast China. Appl. Soil Ecol. 2017, 113, 11–21. [Google Scholar] [CrossRef]
- Ji, F.Y.; Ming, H.X.; Li, H.B.; Zan, S.J.; Wang, J.N.; Su, J.; Guo, L.M.; Chang, Y.K.; Shi, Y.; Guan, C.J.; et al. Diversity of CO2 fixation gene in the surface waters of northern South China Sea in the Calvin cycle. Acta Sci. Circumst. 2016, 36, 4037–4043. (In Chinese) [Google Scholar]
- Liu, Q.; Wei, X.M.; Wu, X.H.; Yuan, H.C.; Wang, J.R.; Li, Y.Y.; Ge, T.D.; Wu, J.S. Characteristic of abundances and diversity of carbon dioxide fixation microbes in paddy soils. Environ. Sci. 2017, 38, 760–768. (In Chinese) [Google Scholar]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low- and high-molecularweight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Hall, E.K.; Wanek, W.; Szukics, U.; Hämmerle, I.; Ellersdorfer, G.; Böck, S.; Strauss, J.; Sterflinger, K.; Richter, A.; et al. The effect of resource quantity and resource stoichiometry on microbial carbon-use-efficiency. FEMS Microbiol. Ecol. 2010, 73, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Hug, L.A.; Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Frischkorn, K.R.; Williams, K.H.; Tringe, S.G.; Banfield, J.F. Community genomic analyses constrain the distribution of metabolic traits across the Chloroflexi phylum and indicate roles in sediment carbon cycling. Microbiome 2013, 1, 22. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Khan, S.; Li, Y.; Zheng, N.; Yao, H. Influence of biochars on the accessibility of organochlorine pesticides and microbial community in contaminated soils. Sci. Total. Environ. 2019, 647, 551–560. [Google Scholar] [CrossRef]
- Yao, Q.; Liu, J.; Yu, Z.J.; Li, Y.; Jin, J.; Liu, X.; Wang, G. Three years of biochar amendment alters soil physiochemical properties and fungal community composition in a black soil of northeast China. Soil Biol. Biochem. 2017, 110, 56–67. [Google Scholar] [CrossRef]
- Liu, H.; Pan, F.J.; Han, X.Z.; Song, F.B.; Zhang, Z.M.; Yan, J.; Xu, Y.L. A comprehensive analysis of the response of the fungal community structure to long-term continuous cropping in three typical upland crops. J. Integr. Agric. 2020, 19, 866–880. [Google Scholar] [CrossRef]
- Dai, Z.M.; Enders, A.; Rodrigues, J.; Hanley, K.; Brookes, P.; Xu, J.M.; Lehmann, J. Soil fungal taxonomic and functional community composition as affected by biochar properties. Soil Biol. Biochem. 2018, 126, 159–167. [Google Scholar] [CrossRef]
- Wang, Y. Effects of Biochar Combined with Plant Growth Promoting Rhizobacteria (PGPR) on Crop Growth and Soil Physical, Chemical and Biological Properties; Shandong Agricultural University: Qingdao, China, 2019. [Google Scholar]
- Ullah, N.; Ditta, A.; Khalid, A.; Mehmood, S.; Rizwan, M.S.; Ashraf, M.; Mubeen, F.; Imtiaz, M.; Iqbal, M.M. Integrated effect of algal biochar and plant growth promoting rhizobacteria on physiology and growth of maize under water deficit irrigations. J. Soil Sci. Plant Nutr. 2020, 20, 346–356. [Google Scholar] [CrossRef]
- Chen, S.; Rotaru, A.E.; Shrestha, P.M.; Malvankar, N.S.; Liu, F.; Fan, W.; Nevin, K.P.; Lovley, D.R. Promoting interspecies electron transfer with biochar. Sci. Rep. 2014, 4, 5019. [Google Scholar] [CrossRef] [Green Version]
- Hale, L.; Luth, M.; Kenney, R.; Crowley, D. Evaluation of pinewood biochar as a carrier of bacterial strain Enterobacter cloacae UW5 for soil inoculation. Appl. Soil Ecol. 2014, 84, 192–199. [Google Scholar] [CrossRef]
- Liu, S.D.; Han, G.Y.; Zhu, X.P.; Wu, X.N. Effects of cotton stalk carbon regulation on fungal community structure and function in alkaline Cadmium contaminated rice rhizosphere soil. Environ. Sci. 2020, 41, 3846–3854. (In Chinese) [Google Scholar]
- Li, S.; Liang, C.; Shangguan, Z.P. Effects of apple branch biochar on soil C mineralization and nutrient cycling under two levels of N. Sci. Total. Environ. 2017, 607–608, 109–111. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.; Chu, H.; Jin, J. Soil carbon content drives the biogeographical distribution of fungal communities in the black soil zone of Northeast China. Soil Biol. Biochem. 2015, 83, 29–39. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Kennedy, J.F.; Jiang, M.; Wu, X. Chitosan induces resistance to tuber rot in stored potato caused by alternaria tenuissima. Int. J. Biol. Macromol. 2019, 140, 13204. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Ma, L.N. Study on Soil Microbial Response Characteristics of Soda Saline-Sodic Soil Improved by Rice Cultivation in Northeast China; University of Chinese Academy of Sciences: Haerbin, China, 2020. [Google Scholar]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Li, Y.G.; Zhou, C.R.; Ning, L.F.; Yang, H.Q. Effects of carbonized apple branches on soil bacterial and fungal diversity in apple root zone. Sci. Agric. Sin. 2016, 49, 3413–3424. [Google Scholar]
- Ding, Y.L.; Liu, J.; Wang, Y.Y. Effects of biochar on microbial ecology in agriculture soil: A review. Chin. J. Appl. Ecol. 2013, 24, 3311–3317. (In Chinese) [Google Scholar]
- Guo, Y.; Sun, B.H.; Wang, Y.; Wei, J.; Gao, M.X.; Zhang, S.L.; Yang, X.Y. PLFA fingerprint characteristics of an anthropogenic loess soil under long-term different fertilizations. Sci. Agric. Sin. 2017, 50, 94–103. (In Chinese) [Google Scholar]
- Yue, L.Y.; Zheng, J.Q.; Han, S.J.; Yang, J.H.; Geng, S.C.; Chen, Z.J.; Zhang, X.; Gu, Y. Soil chemical properties and microbial community structure at different succession stages of temperate forest in Changbai Mountains. Chin. J. Ecol. 2015, 34, 2590–2597. (In Chinese) [Google Scholar]
- Li, C.Y.; Wang, Y.; Brookes, P.; Dang, T.H.; Wang, W.Z. Effect of soil pH on soil microbial carbon phosphorus ratio. Sci. Agric. Sin. 2013, 46, 2709–2716. (In Chinese) [Google Scholar]
- Xu, Y.C.; Shen, Q.R.; Ran, W. Effects of zero-tillage and application of manure on soil microbial biomass C, N and P after sixteen years of cropping. Acta Pedol. Sin. 2002, 39, 83–90. (In Chinese) [Google Scholar]
- Yang, L.; Jiang, M.; Zhu, W.; Han, L.; Qin, L. Soil bacterial communities with an indicative function response to nutrients in wetlands of Northeastern China that have undergone natural restoration. Ecol. Indic. 2019, 101, 562–571. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; Chinese Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
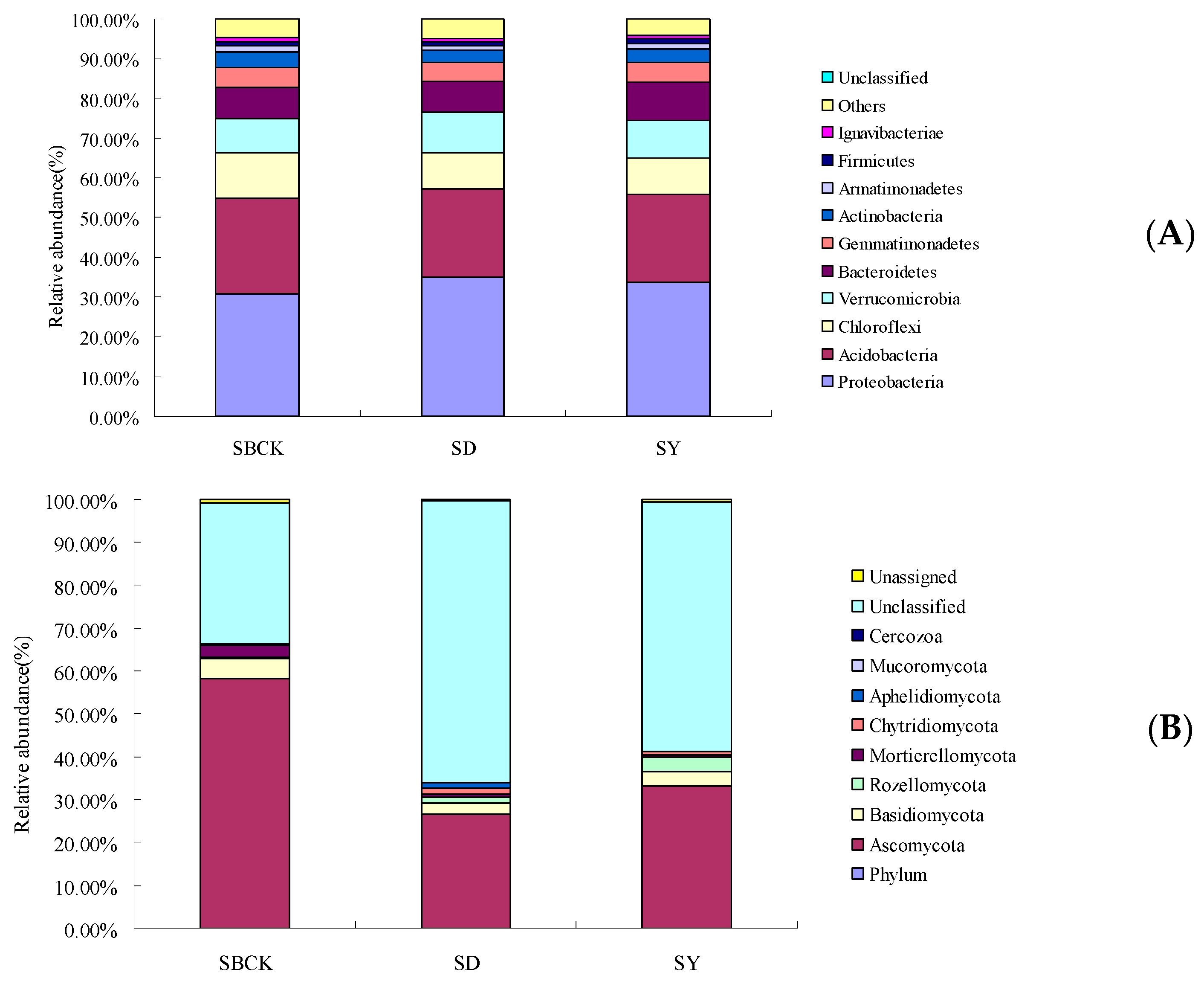




{kind=link}
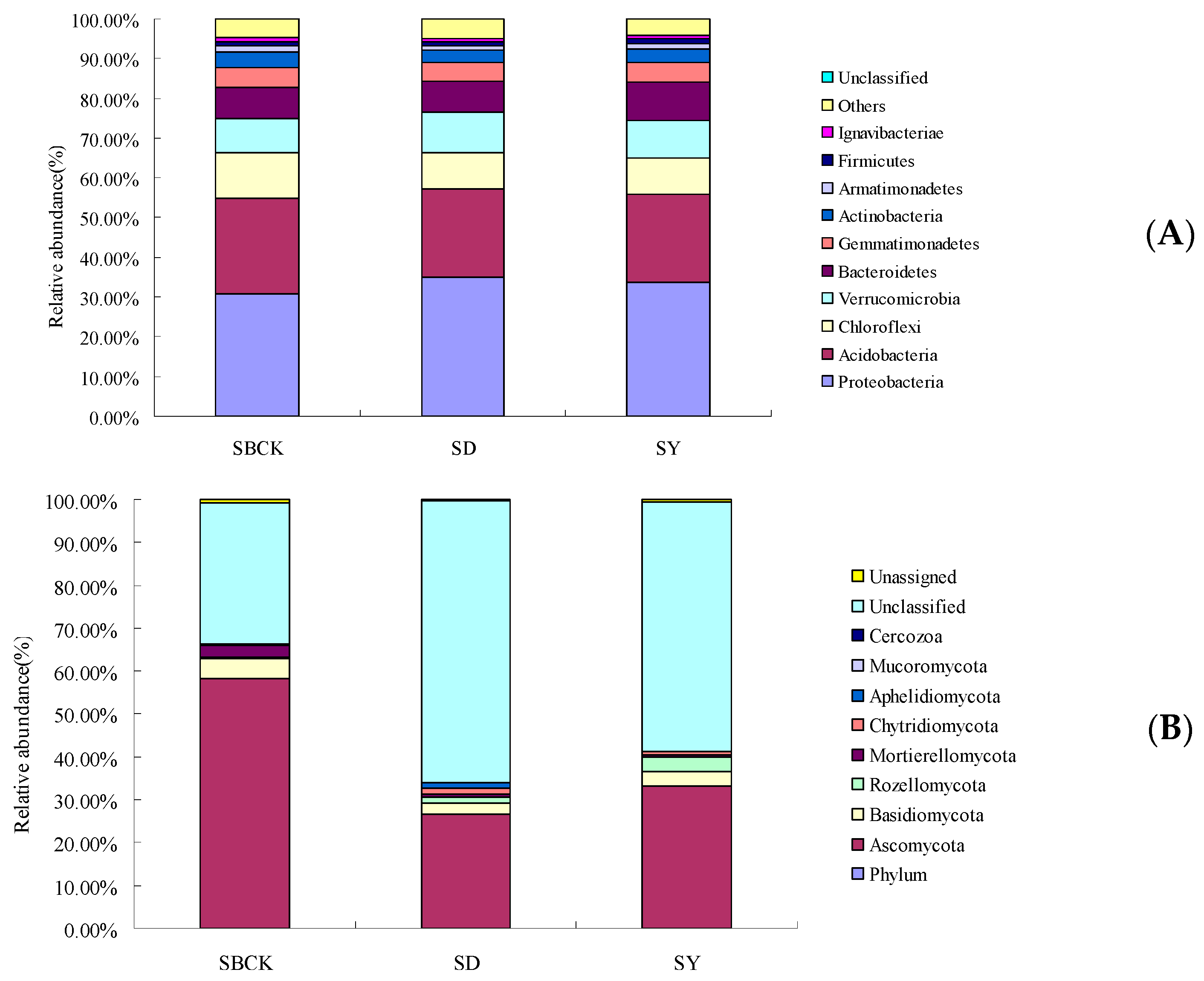{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Organic Matter (OC) (g/kg) | Alkali-Hydrolyzed Nitrogen (AN) (mg/kg) | Available Phosphorus (AP) (mg/kg) | Available
Potassium (AK) (mg/kg) | Total Nitrogen (TN) (g/kg) | Total Phosphorus (TP) (g/kg) | Total Potassium (TK) (g/kg) | |
|---|---|---|---|---|---|---|---|---|
| SBCK | 6.80 a | 38.27 a | 147.67 c | 57.20 b | 121.00 b | 1.85 a | 1.04 a | 17.33 a |
| SD | 6.30 a | 38.67 a | 166.67 a | 62.47 a | 126.00 a | 1.94 a | 1.08 a | 17.40 a |
| SY | 6.60 a | 38.77 a | 158.67 b | 62.13 a | 104.67 c | 1.93 a | 1.13 a | 18.27 a |
| Treatment | ACE Index | Chao Index | Simpson Index | Shannon Index | ACE Index | Chao Index | Simpson Index | Shannon Index |
|---|---|---|---|---|---|---|---|---|
| Bacterial (V3 + V4) | Fungal (ITS1) | |||||||
| SBCK | 1555.28 a | 1569.04 a | 0.01 a | 6.02 a | 278.18 ab | 269.35 ab | 0.12 ab | 3.34 a,b |
| SD | 1603.53 a | 1619.97 a | 0.01 a | 6.05 a | 300.04 a | 307.49 a | 0.08 b | 3.47 a |
| SY | 1587.25 a | 1604.48 a | 0.01 a | 6.00 a | 262.53 b | 262.74 b | 0.14 a | 3.11 b |
| Treatment | Bacterial (V3 + V4) OTU Numbers | Coverage | Fungal (ITS1) OTU Numbers | Coverage |
|---|---|---|---|---|
| SBCK | 1504.33 a | 0.9973 a | 240.33 b | 0.9998 a |
| SD | 1549.67 a | 0.9961 a | 288.33 a | 0.9997 a |
| SY | 1546.67 a | 0.9979 a | 251.67 a,b | 0.9998 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, D.; Li, H.; Wang, H.; Guo, X.; Wang, Z.; Lv, Y.; Ding, G.; Jin, L.; Lan, Y. Impact of Different Biochars on Microbial Community Structure in the Rhizospheric Soil of Rice Grown in Albic Soil. Molecules 2021, 26, 4783. https://doi.org/10.3390/molecules26164783
Yin D, Li H, Wang H, Guo X, Wang Z, Lv Y, Ding G, Jin L, Lan Y. Impact of Different Biochars on Microbial Community Structure in the Rhizospheric Soil of Rice Grown in Albic Soil. Molecules. 2021; 26(16):4783. https://doi.org/10.3390/molecules26164783
Chicago/Turabian StyleYin, Dawei, Hongyu Li, Haize Wang, Xiaohong Guo, Zhihui Wang, Yandong Lv, Guohua Ding, Liang Jin, and Yu Lan. 2021. "Impact of Different Biochars on Microbial Community Structure in the Rhizospheric Soil of Rice Grown in Albic Soil" Molecules 26, no. 16: 4783. https://doi.org/10.3390/molecules26164783
APA StyleYin, D., Li, H., Wang, H., Guo, X., Wang, Z., Lv, Y., Ding, G., Jin, L., & Lan, Y. (2021). Impact of Different Biochars on Microbial Community Structure in the Rhizospheric Soil of Rice Grown in Albic Soil. Molecules, 26(16), 4783. https://doi.org/10.3390/molecules26164783