
Inhibition of Bacterial Efflux Pumps by Crude Extracts and Essential Oil from Myristica fragrans Houtt. (Nutmeg) Seeds against Methicillin-Resistant Staphylococcus aureus

 , , , , and
, , , , and
Abstract
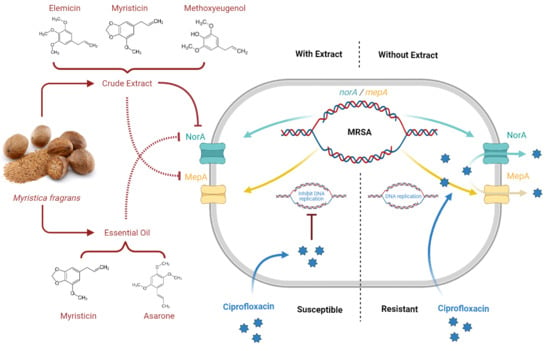
1. Introduction
2. Results
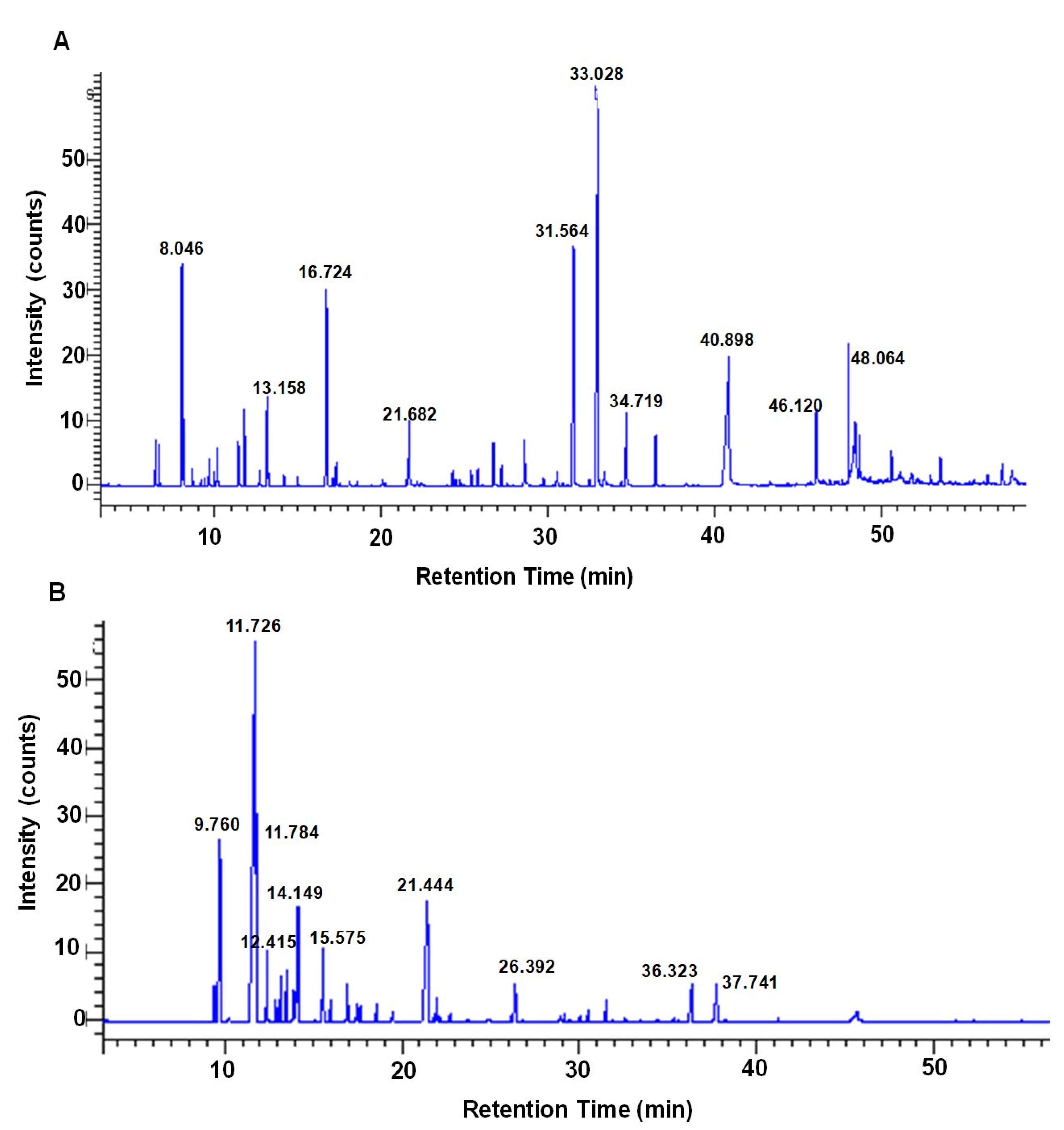
2.1. Determination of Chemical Composition of Crude Extract and Essential Oil
2.2. Determination of MRSA
2.3. Genotypic Determination of Resistant Genes and Efflux Genes in MRSA Strains
2.4. Antimicrobial Susceptibility Testing
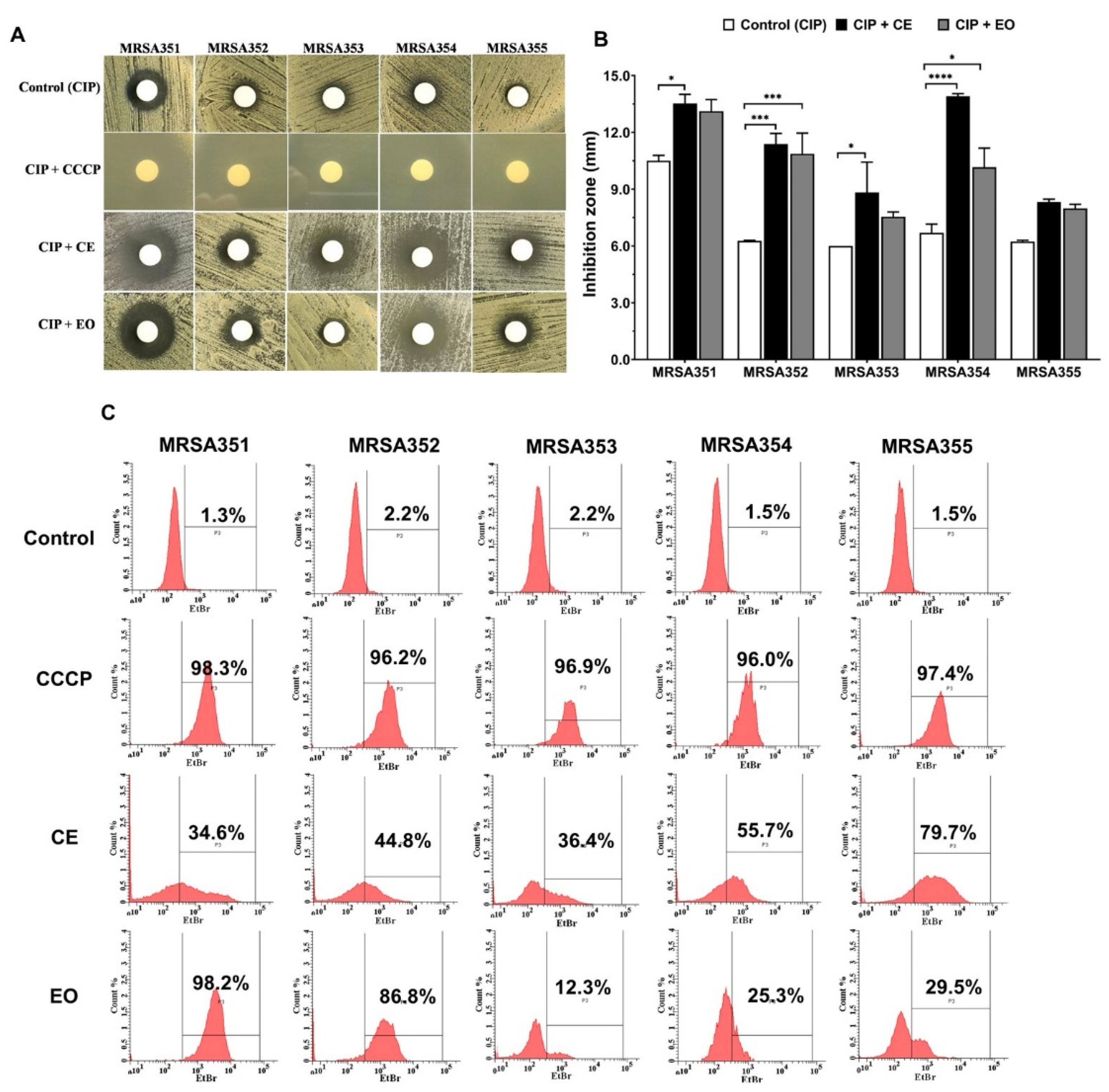
2.5. Combination Activity of CE or EO with Ciprofloxacin
2.6. Determination of Efflux Pump Inhibitors (EPIs)
3. Discussions
4. Materials and Methods
4.1. Preparation of Crude Extract and Essential Oil
4.2. Bacterial Strains
4.3. Antimicrobial Susceptibility Testing
4.4. Polymerase Chain Reaction (PCR) Method
4.5. RNA Extraction and cDNA Synthesis
4.6. Quantitative Real-Time PCR (qPCR) Method
4.7. Checkerboard Titration Assay
4.8. Examination of Inhibition Zone, CE and EO with Ciprofloxacin Combinations by Agar Disc Diffusion
4.9. Evaluation of Ethidium Bromide Accumulation in the MRSA Strains by Flow Cytometry
4.10. Inhibition of Bacterial Growth by CE and EO with Ciprofloxacin Combinations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Asgarpanah, J.; Kazemivash, N. Phytochemistry and pharmacologic properties of Myristica fragrans Hoyutt.: A review. AFRICAN J. Biotechnol. 2012, 11, 12787–12793. [Google Scholar] [CrossRef]
- Matulyte, I.; Jekabsone, A.; Jankauskaite, L.; Zavistanaviciute, P.; Sakiene, V.; Bartkiene, E.; Ruzauskas, M.; Kopustinskiene, D.M.; Santini, A.; Bernatoniene, J. The Essential Oil and Hydrolats from Myristica fragrans Seeds with Magnesium Aluminometasilicate as Excipient: Antioxidant, Antibacterial, and Anti-inflammatory Activity. Foods 2020, 9, 37. [Google Scholar] [CrossRef]
- Takikawa, A.; Abe, K.; Yamamoto, M.; Ishimaru, S.; Yasui, M.; Okubo, Y.; Yokoigawa, K. Antimicrobial activity of Nutmeg against Escherichia coli O157. J. Biosci. Bioeng. 2002, 94, 315–320. [Google Scholar] [CrossRef]
- Olajide, O.A.; Ajayi, F.F.; Ekhelar, A.I.; Awe, S.O.; Makinde, J.M.; Akinola Alada, A.R. Biological Effects of Myristica fragrans (Nutmeg) Extract. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 1999, 13, 344–345. [Google Scholar] [CrossRef]
- Tajuddin, A.; Ahmad, S.; Latif, A.; Qasmi, I.A. Aphrodisiac activity of 50% ethanolic extracts of Myristica fragrans Houtt. (nutmeg) and Syzygium aromaticum (L) Merr. & Perry. (clove) in male mice: A comparative study. BMC Complement. Altern. Med. 2003, 3, 1–5. [Google Scholar] [CrossRef]
- Gupta, A.D.; Rajpurohit, D. Antioxidant and Antimicrobial Activity of Nutmeg (Myristica fragrans). Nuts Seeds Heal. Dis. Prev. 2011, 831–839. [Google Scholar] [CrossRef]
- Singh, G.; Marimuthu, P.; De Heluani, C.S.; Catalan, C. Antimicrobial and antioxidant potentials of essential oil and acetone extract of Myristica fragrans Houtt. (aril part). J. Food Sci. 2005, 70, M141–M148. [Google Scholar] [CrossRef]
- Ansory, H.M.; Putri, P.K.K.; Hidayah, N.A.; Nilawati, A. Analisis Senyawa Minyak Atsiri Fuli Pala Secara GC-MS dan Uji Aktivitas Antibakteri Terhadap Ecschericia coli dan Staphylococcus aureus. Maj. Farm. 2018, 13, 56. [Google Scholar] [CrossRef]
- Nurjanah, S.; Putri, I.L.; Sugiarti, D.P. Antibacterial Activity of Nutmeg Oil. KnE Life Sci. 2017, 2, 563. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; El-Saadony, M.T.; Alagawany, M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Elwan, H.A.M.; Elnesr, S.S.; Abd El-Hack, M.E. Using essential oils to overcome bacterial biofilm formation and their antimicrobial resistance. Saudi J. Biol. Sci. 2021. [Google Scholar] [CrossRef]
- Seukep, A.J.; Kuete, V.; Nahar, L.; Sarker, S.D.; Guo, M. Plant-derived secondary metabolites as the main source of efflux pump inhibitors and methods for identification. J. Pharm. Anal. 2020, 10, 277–290. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Javdan, S.; Narimani, T.; Shahini Shams Abadi, M.; Gholipour, A. Agr typing of Staphylococcus aureus species isolated from clinical samples in training hospitals of Isfahan and Shahrekord. BMC Res. Notes 2019, 12, 1–6. [Google Scholar] [CrossRef]
- Lowy, F.D. Antimicrobial resistance: The example of Staphylococcus aureus. J. Clin. Invest. 2003, 111, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Luan, W.; Liu, X.; Wang, X.; An, Y.; Wang, Y.; Wang, C.; Shen, K.; Xu, H.; Li, S.; Liu, M.; et al. Inhibition of Drug Resistance of Staphylococcus aureus by Efflux Pump Inhibitor and Autolysis Inducer to Strengthen the Antibacterial Activity of β-lactam Drugs. Pol. J. Microbiol. 2019, 68, 477–491. [Google Scholar] [CrossRef]
- Jaganath, D.; Jorakate, P.; Makprasert, S.; Sangwichian, O.; Akarachotpong, T.; Thamthitiwat, S.; Khemla, S.; DeFries, T.; Baggett, H.C.; Whistler, T.; et al. Staphylococcus aureus bacteremia incidence and methicillin resistance in Rural Thailand, 2006–2014. Am. J. Trop. Med. Hyg. 2018, 99, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Apisarnthanarak, A.; Ratz, D.; Khawcharoenporn, T.; Patel, P.K.; Weber, D.J.; Saint, S.; Greene, M.T. National survey of practices to prevent methicillin-resistant staphylococcus aureus and multidrug-resistant acinetobacter baumannii in Thailand. Clin. Infect. Dis. 2017, 64, S161–S166. [Google Scholar] [CrossRef]
- Ashraf, M.; Mustafa, B.-E.; Rehman, S.-U.; Khalid Bashir, M.; Adnan Ashraf, M. Emergence of Antimicrobial Resistance, Causes, Molecular Mechanisms, and Prevention Strategies: A Bovine Perspective. Bov. Sci. A Key Sustain. Dev. 2019. [Google Scholar] [CrossRef]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef]
- Schindler, B.D.; Jacinto, P.; Kaatz, G.W. Inhibition of drug efflux pumps in Staphylococcus aureus: Current status of potentiating existing antibiotics. Future Microbiol. 2013, 8, 491–507. [Google Scholar] [CrossRef]
- Zimmermann, S.; Klinger-Strobel, M.; Bohnert, J.A.; Wendler, S.; Rödel, J.; Pletz, M.W.; Löffler, B.; Tuchscherr, L. Clinically Approved Drugs Inhibit the Staphylococcus aureus Multidrug NorA Efflux Pump and Reduce Biofilm Formation. Front. Microbiol. 2019, 10, 2762. [Google Scholar] [CrossRef]
- German, N.; Wei, P.; Kaatz, G.W.; Kerns, R.J. Synthesis and evaluation of fluoroquinolone derivatives as substrate-based inhibitors of bacterial efflux pumps. Eur. J. Med. Chem. 2008, 43, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Sonavane, G.S.; Sarveiya, V.P.; Kasture, V.S.; Kasture, S.B. Anxiogenic activity of Myristica fragrans seeds. Pharmacol. Biochem. Behav. 2002, 71, 239–244. [Google Scholar] [CrossRef]
- Gupta, A.D.; Bansal, V.K.; Babu, V.; Maithil, N. Chemistry, antioxidant and antimicrobial potential of nutmeg (Myristica fragrans Houtt). J. Genet. Eng. Biotechnol. 2013, 11, 25–31. [Google Scholar] [CrossRef]
- Dzotam, J.K.; Simo, I.K.; Bitchagno, G.; Celik, I.; Sandjo, L.P.; Tane, P.; Kuete, V. In vitro antibacterial and antibiotic modifying activity of crude extract, fractions and 3’,4’,7-trihydroxyflavone from Myristica fragrans Houtt against MDR Gram-negative enteric bacteria. BMC Complement. Altern. Med. 2018, 18, 1–9. [Google Scholar] [CrossRef]
- Gradelski, E.; Kolek, B.; Bonner, D.P.; Valera, L.; Minassian, B.; Fung-Tomc, J. Activity of gatifloxacin and ciprofloxacin in combination with other antimicrobial agents. Int. J. Antimicrob. Agents 2001, 17, 103–107. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Cantrell, C.L.; Jeliazkova, E.A.; Astatkie, T.; Zheljazkov, V.D. Utilization of nutmeg (Myristica fragrans Houtt.) seed hydrodistillation time to produce essential oil fractions with varied compositions and pharmacological effects. Molecules 2020, 25, 565. [Google Scholar] [CrossRef]
- Kapoor, I.P.S.; Singh, B.; Singh, G.; De Heluani, C.S.; De Lampasona, M.P.; Catalan, C.A.N. Chemical composition and antioxidant activity of essential oil and oleoresins of nutmeg (Myristica fragrans Houtt.) fruits. Int. J. Food Prop. 2013, 16, 1059–1070. [Google Scholar] [CrossRef]
- Shafiei, Z.; Shuhairi, N.N.; Md Fazly Shah Yap, N.; Harry Sibungkil, C.A.; Latip, J. Antibacterial activity of Myristica fragrans against oral pathogens. Evid.-Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Muhamad Ansory, H.; Khania Kusuma Putri, P.; Hidayah, A.; Nilawati, A. The Analysis of Nutmeg Mace Essential Oil Compound Using GC-MS and Antibacterial Activity Test toward Ecschericia coli and Staphylococcus aureus. Maj. Farm. 2020, 16, 1–8. [Google Scholar] [CrossRef]
- Narasimhan, B.; Dhake, A.S. Antibacterial principles from Myristica fragrans seeds. J. Med. Food 2006, 9, 395–399. [Google Scholar] [CrossRef]
- Izah, S.C.; Zige, D.V.; Alagoa, K.J.; Uhunmwangho, E.J.; Iyamu, A.O. Antibacterial Efficacy of Aqueous Extract of Myristica fragrans (Common Nutmeg). EC Pharmacol. Toxicol. 2018, 6, 291–295. [Google Scholar]
- Muchtaridi; Subarnas, A.; Apriyantono, A.; Mustarichie, R. Identification of compounds in the essential oil of nutmeg seeds (Myristica fragrans Houtt.) that inhibit locomotor activity in mice. Int. J. Mol. Sci. 2010, 11, 4771–4781. [Google Scholar] [CrossRef]
- Morsy, N.F.S. A comparative study of nutmeg (Myristica fragrans Houtt.) oleoresins obtained by conventional and green extraction techniques. J. Food Sci. Technol. 2016, 53, 3770–3777. [Google Scholar] [CrossRef] [PubMed]
- Felicetti, T.; Cannalire, R.; Pietrella, D.; Latacz, G.; Lubelska, A.; Manfroni, G.; Barreca, M.L.; Massari, S.; Tabarrini, O.; Kieć-Kononowicz, K.; et al. 2-Phenylquinoline S. aureus NorA Efflux Pump Inhibitors: Evaluation of the Importance of Methoxy Group Introduction. J. Med. Chem. 2018, 61, 7827–7848. [Google Scholar] [CrossRef]
- Hassoun, A.; Linden, P.K.; Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations-a review of recent developments in MRSA management and treatment. Crit. Care 2017, 21, 211. [Google Scholar] [CrossRef] [PubMed]
- Sundaramoorthy, N.S.; Mitra, K.; Ganesh, J.S.; Makala, H.; Lotha, R.; Bhanuvalli, S.R.; Ulaganathan, V.; Tiru, V.; Sivasubramanian, A.; Nagarajan, S. Ferulic acid derivative inhibits NorA efflux and in combination with ciprofloxacin curtails growth of MRSA in vitro and in vivo. Microb. Pathog. 2018, 124, 54–62. [Google Scholar] [CrossRef]
- Hassanzadeh, S.; Mashhadi, R.; Yousefi, M.; Askari, E.; Saniei, M.; Pourmand, M.R. Frequency of efflux pump genes mediating ciprofloxacin and antiseptic resistance in methicillin-resistant Staphylococcus aureus isolates. Microb. Pathog. 2017, 111, 71–74. [Google Scholar] [CrossRef]
- Lekshmi, M.; Ammini, P.; Adjei, J.; Sanford, L.M.; Shrestha, U.; Kumar, S.; Varela, M.F. Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus. AIMS Microbiol. 2018, 4, 1–18. [Google Scholar] [CrossRef]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug Efflux Pumps in Staphylococcus aureus: An Update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef]
- Wielders, C.L.C.; Fluit, A.C.; Brisse, S.; Verhoef, J.; Schmitz, F.J. mecA gene is widely disseminated in Staphylococcus aureus population. J. Clin. Microbiol. 2002, 40, 3970–3975. [Google Scholar] [CrossRef]
- Felicetti, T.; Cannalire, R.; Nizi, M.G.; Tabarrini, O.; Massari, S.; Barreca, M.L.; Manfroni, G.; Schindler, B.D.; Cecchetti, V.; Kaatz, G.W.; et al. Studies on 2-phenylquinoline Staphylococcus aureus NorA efflux pump inhibitors: New insights on the C-6 position. Eur. J. Med. Chem. 2018, 155, 428–433. [Google Scholar] [CrossRef]
- De Azevedo Maia, G.L.; Dos Santos Falcão-Silva, V.; Aquino, P.G.V.; De Araújo-Júnior, J.X.; Tavares, J.F.; Da Silva, M.S.; Rodrigues, L.C.; De Siqueira-Júnior, J.P.; Barbosa-Filho, J.M. Flavonoids from Praxelis clematidea R.M. King and Robinson modulate bacterial drug resistance. Molecules 2011, 16, 4828–4835. [Google Scholar] [CrossRef]
- Holler, J.G.; Slotved, H.C.; Molgaard, P.; Olsen, C.E.; Christensen, S.B. Chalcone inhibitors of the NorA efflux pump in Staphylococcus aureus whole cells and enriched everted membrane vesicles. Bioorg. Med. Chem. 2012, 20, 4514–4521. [Google Scholar] [CrossRef]
- Brincat, J.P.; Broccatelli, F.; Sabatini, S.; Frosini, M.; Neri, A.; Kaatz, G.W.; Cruciani, G.; Carosati, E. Ligand promiscuity between the efflux pumps human p-glycoprotein and s. aureus NorA. ACS Med. Chem. Lett. 2012, 3, 248–251. [Google Scholar] [CrossRef]
- de Sousa Andrade, L.M.; de Oliveira, A.B.M.; Leal, A.L.A.B.; de Alcântara Oliveira, F.A.; Portela, A.L.; de Sousa Lima Neto, J.; de Siqueira-Júnior, J.P.; Kaatz, G.W.; da Rocha, C.Q.; Barreto, H.M. Antimicrobial activity and inhibition of the NorA efflux pump of Staphylococcus aureus by extract and isolated compounds from Arrabidaea brachypoda. Microb. Pathog. 2020, 140, 103935. [Google Scholar] [CrossRef]
- Thileepan, T.; Thevanesam, V.; Kathirgamanathar, S. Antimicrobial Activity of Seeds and Leaves of Myristica fragrans against Multi-resistant Microorganisms. J. Agric. Sci. Technol. A 2017, 7, 302–308. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Al-Mariri, A.; Safi, M. In vitro antibacterial activity of several plant extracts and oils against some gram-negative bacteria. Iran. J. Med. Sci. 2014, 39, 36–43. [Google Scholar]
- Lahmar, I.; Belghith, H.; Ben Abdallah, F.; Belghith, K. Nutritional Composition and Phytochemical, Antioxidative, and Antifungal Activities of Pergularia tomentosa L. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Chaiittianan, R.; Chayopas, P.; Rattanathongkom, A.; Tippayawat, P.; Sutthanut, K. Anti-obesity potential of corn silks: Relationships of phytochemicals and antioxidation, anti-pre-adipocyte proliferation, anti-adipogenesis, and lipolysis induction. J. Funct. Foods 2016, 23, 497–510. [Google Scholar] [CrossRef]
- Matulyte, I.; Marksa, M.; Ivanauskas, L.; Kalveniene, Z.; Lazauskas, R.; Bernatoniene, J. GC-MS analysis of the composition of the extracts and essential Oil from Myristica fragrans Seeds Using Magnesium Aluminometasilicate as Excipient. Molecules 2019, 24, 1062. [Google Scholar] [CrossRef] [PubMed]
- Sutthamee, P.; Thaiderm, A.; Wongthong, S.; Srisrattakarn, A.; Wonglakorn, L.; Puang-Ngern, P.; Tavichakorntrakool, R.; Chanawong, A.; Lulitanond, A. SCCmec types of methicillin-resistant Staphylococcus aureus isolated in 2010, 2015 and 2016 from patients of Srinagarind Hospital, Khon Kaen Province. Arch. Allied Heal. Sci. 2019, 31, 140–149. [Google Scholar]
- Yu, J.L.; Grinius, L.; Hooper, D.C. NorA functions as a multidrug efflux protein in both cytoplasmic membrane vesicles and reconstituted proteoliposomes. J. Bacteriol. 2002, 184, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xie, K.; Zou, D.; Meng, M.; Xie, M. Inhibitory effects of silybin on the efflux pump of methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2018, 18, 827–833. [Google Scholar] [CrossRef]
- Dabul, A.N.G.; Avaca-Crusca, J.S.; Van Tyne, D.; Gilmore, M.S.; Camargo, I.L.B.C. Resistance in In Vitro Selected Tigecycline-Resistant Methicillin-Resistant Staphylococcus aureus Sequence Type 5 is Driven by Mutations in mepR and mepA Genes. Microb. Drug Resist. 2018, 24, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Turkey, A.M.; Barzani, K.K.; Suleiman, A.A.J.; Abed, J.J. Molecular assessment of accessory gene regulator (Agr) quorum sensing system in biofilm forming Staphylococcus aureus and study of the effect of silver nanoparticles on agr system. Iran. J. Microbiol. 2018, 10, 14–21. [Google Scholar] [PubMed]
- Gaur, R.; Gupta, V.K.; Pal, A.; Darokar, M.P.; Bhakuni, R.S.; Kumar, B. In vitro and in vivo synergistic interaction of substituted chalcone derivatives with norfloxacin against methicillin resistant Staphylococcus aureus. RSC Adv. 2015, 5, 5830–5845. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutmeg (Seed Powder) | TPC a (µg) | TFC b (µg) |
|---|---|---|
| Crude extract | 397 ± 0.70 | 21 ± 0.08 |
| Essential oil | 964 ± 0.98 | 180 ± 0.26 |
| Peak Area (%) | Molecular Formula | Peak Name | RT a (min) | MW b (g/mol) |
|---|---|---|---|---|
| Crude extract | ||||
| 22.663 | C12H16O3 | Elemicin | 33.028 | 22.66 |
| 15.234 | C14H28O2 | Tetradecanoic acid | 40.898 | 228.37 |
| 11.183 | C11H12O3 | Myristicin | 31.564 | 192.21 |
| 6.880 | C10H18O | 4-terpineol | 16.724 | 154.25 |
| 5.771 | C10H16 | Sabinene | 8.046 | 136.23 |
| 2.774 | C11H14O3 | Methoxyeugenol | 34.719 | 194.22 |
| 2.740 | C10H18O | cis-4-Thujanol | 13.158 | 154.25 |
| 2.603 | C4H6O | Decanoic acid, 2-oxo-, methyl ester | 48.064 | 200.27 |
| 2.283 | C10H10O2 | Safrene | 21.682 | 162.18 |
| 2.283 | C16H32O | n-Hexadecanoic acid | 46.120 | 256.42 |
| Essential oil | ||||
| 36.907 | C10H16 | Sabinene | 11.726 | 136.23 |
| 11.544 | C10H18O | 4-terpineol | 21.444 | 154.25 |
| 9.414 | C10H16 | α-pinene | 9.760 | 136.23 |
| 6.135 | C10H16 | β-phellandrene | 14.149 | 136.23 |
| 4.445 | C10H16 | β-pinene | 11.784 | 136.2 |
| 3.312 | C10H16 | γ-terpinene | 15.575 | 136.23 |
| 2.624 | C10H16 | β-myrcene | 12.415 | 136.23 |
| 2.551 | C11H12O3 | Myristicin | 36.323 | 192.21 |
| 2.549 | C12H16O3 | Cis-asarone | 37.741 | 208.25 |
| 1.884 | C10H10O2 | 1,3-Benzodioxole, 5-(2-propenyl)- | 26.392 | 162.18 |
| Microorganism | Cefoxitin | Ciprofloxacin | ||
|---|---|---|---|---|
| Inhibition Zone (mm) | Interpretation * | Inhibition Zone (mm) | Interpretation * | |
| MRSA351 | 6 | aR | 8 | R |
| MRSA352 | 13 | R | 6 | R |
| MRSA353 | 6 | R | 6 | R |
| MRSA354 | 6 | R | 6 | R |
| MRSA355 | 6 | R | 6 | R |
| Microorganism | CE (µg/mL) | EO (% v/v) | ||
|---|---|---|---|---|
| MIC a | MBC b | MIC | MBC | |
| MRSA351 | 312 | 625 | 0.098 | 0.195 |
| MRSA352 | 78 | 156 | 0.098 | 0.195 |
| MRSA353 | 625 | 1250 | 0.098 | 0.195 |
| MRSA354 | 1250 | 2500 | 0.098 | 0.195 |
| MRSA355 | 312 | 625 | 0.195 | 0.391 |
| Microorganism | ∑FIC of CE | Interpretation * | ∑FIC of EO | Interpretation |
|---|---|---|---|---|
| MRSA351 | 3.40 | A/I a | 0.98 | A/I |
| MRSA352 | 1.41 | A/I | 2.62 | A/I |
| MRSA353 | 1.37 | A/I | 1.05 | A/I |
| MRSA354 | 0.88 | A/I | 1.23 | A/I |
| MRSA355 | 1.34 | A/I | 1.90 | A/I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oo, T.; Saiboonjan, B.; Srijampa, S.; Srisrattakarn, A.; Sutthanut, K.; Tavichakorntrakool, R.; Chanawong, A.; Lulitanond, A.; Tippayawat, P. Inhibition of Bacterial Efflux Pumps by Crude Extracts and Essential Oil from Myristica fragrans Houtt. (Nutmeg) Seeds against Methicillin-Resistant Staphylococcus aureus. Molecules 2021, 26, 4662. https://doi.org/10.3390/molecules26154662
Oo T, Saiboonjan B, Srijampa S, Srisrattakarn A, Sutthanut K, Tavichakorntrakool R, Chanawong A, Lulitanond A, Tippayawat P. Inhibition of Bacterial Efflux Pumps by Crude Extracts and Essential Oil from Myristica fragrans Houtt. (Nutmeg) Seeds against Methicillin-Resistant Staphylococcus aureus. Molecules. 2021; 26(15):4662. https://doi.org/10.3390/molecules26154662
Chicago/Turabian StyleOo, Thidar, Bhanubong Saiboonjan, Sukanya Srijampa, Arpasiri Srisrattakarn, Khaetthareeya Sutthanut, Ratree Tavichakorntrakool, Aroonwadee Chanawong, Aroonlug Lulitanond, and Patcharaporn Tippayawat. 2021. "Inhibition of Bacterial Efflux Pumps by Crude Extracts and Essential Oil from Myristica fragrans Houtt. (Nutmeg) Seeds against Methicillin-Resistant Staphylococcus aureus" Molecules 26, no. 15: 4662. https://doi.org/10.3390/molecules26154662
APA StyleOo, T., Saiboonjan, B., Srijampa, S., Srisrattakarn, A., Sutthanut, K., Tavichakorntrakool, R., Chanawong, A., Lulitanond, A., & Tippayawat, P. (2021). Inhibition of Bacterial Efflux Pumps by Crude Extracts and Essential Oil from Myristica fragrans Houtt. (Nutmeg) Seeds against Methicillin-Resistant Staphylococcus aureus. Molecules, 26(15), 4662. https://doi.org/10.3390/molecules26154662