
Phenolic Extract from Aralia nudicaulis L. Rhizomes Inhibits Cellular Oxidative Stresses

 ,
,
Abstract
1. Introduction
2. Results and Discussion
2.1. Extraction Yield and Purification Method
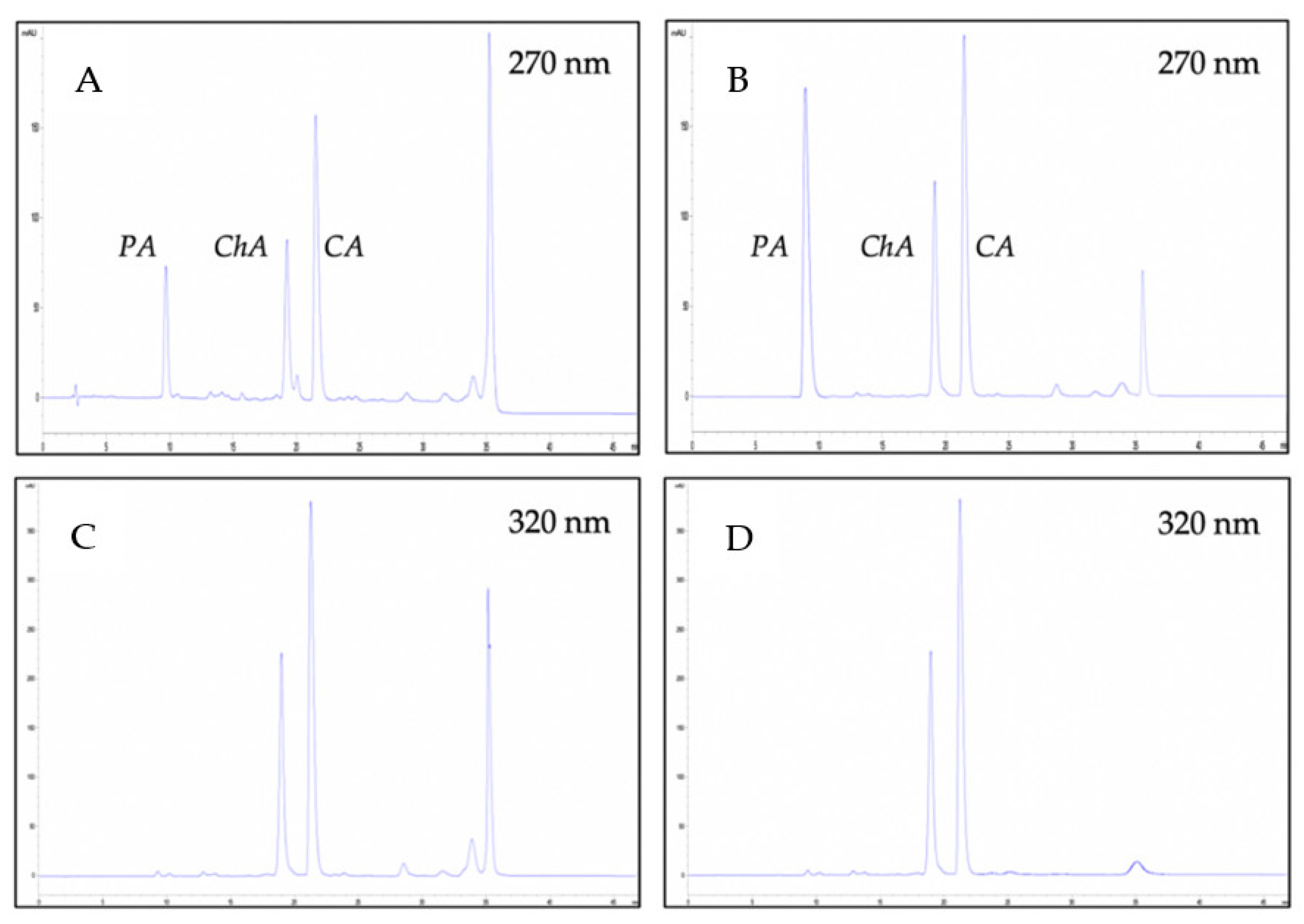
2.2. Identification and Optimization of LC-MS Conditions
2.3. Quantitative Method Validation
2.4. Quantitative Analysis
2.5. Biological Activities Screening
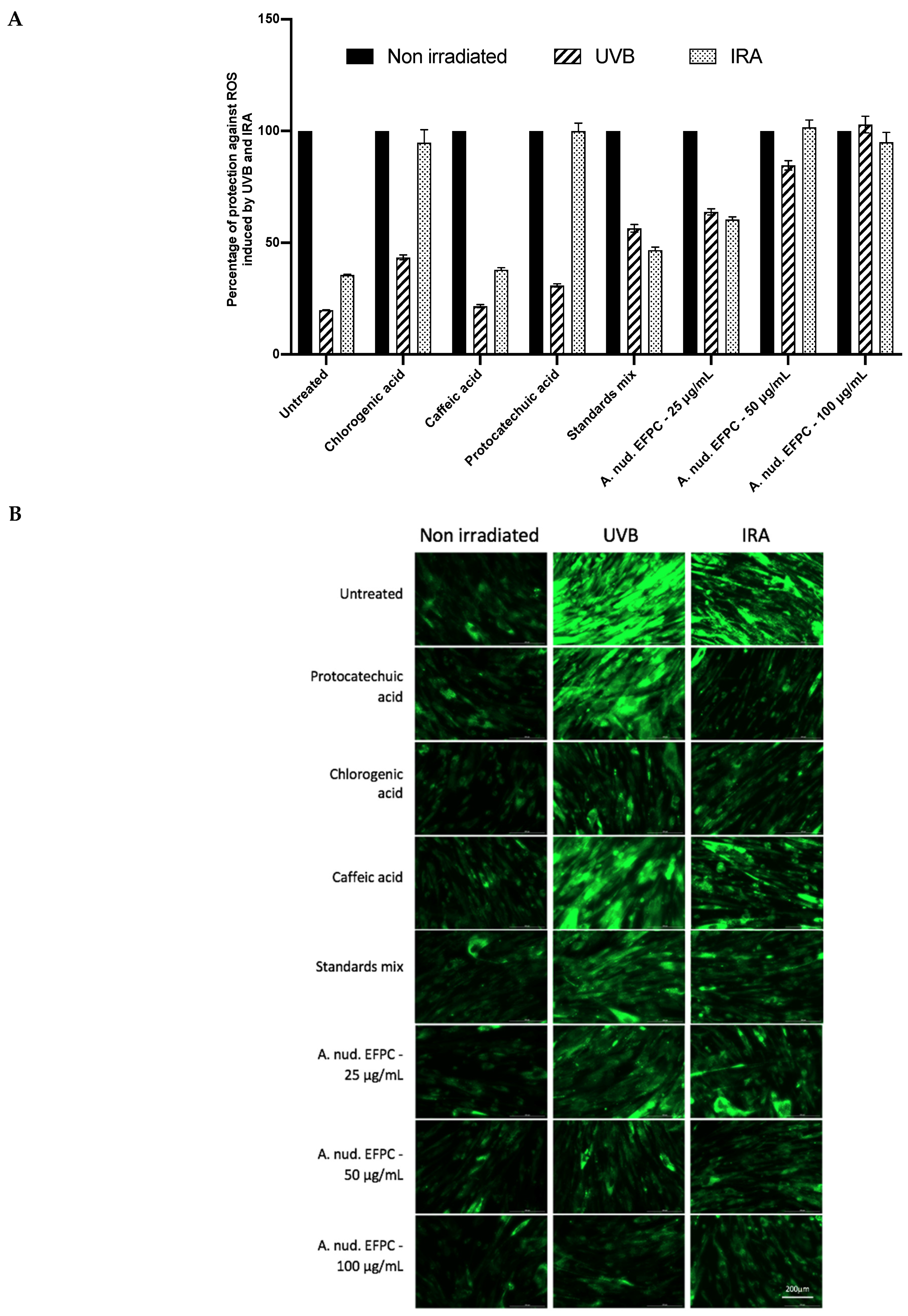
2.6. Evaluation of FEPC Protection against UV-B and IR-A Radiations
3. Materials and Methods
3.1. Crude Extract and Enriched Fraction in Phenolic Compounds
3.2. Standardization Extract and Standards Preparation
3.3. Chromatographic Instrumentation and Conditions
3.4. HPLC Method Validation
3.5. Measurement of Different Biological Activities
3.5.1. ORACFL Assay
3.5.2. Antioxidant Cell Assay Using 2′,7′-Dichlorofluorescin-Diacetate (DCFH-DA)
3.5.3. Measurement of Anti-Inflammatory Activity by Nitrite Quantification
3.5.4. Evaluation of Cytotoxicity against WS-1 Cell Lines
3.5.5. UV-B and IR-A Stress Protection Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Clemens, T.L.; Parrish, J.A.; Holick, M.F. Vitamin-D Synthesis and Metabolism after Ultraviolet Irradiation of Normal and Vitamin-D-Deficient Subjects. N. Engl. J. Med. 1982, 306, 722–725. [Google Scholar] [CrossRef]
- Lu, Y.P.; Lou, Y.R.; Yen, P.; Mitchell, D.; Huang, M.T.; Conney, A.H. Time course for early adaptive responses to ultraviolet B light in the epidermis of SKH-1 mice. Cancer Res. 1999, 59, 4591. [Google Scholar]
- Afaq, F.; Adhami, V.M.; Mukhtar, H. Photochemoprevention of Ultraviolet B Signaling and Photocarcinogenesis. Mutat. Res. 2005, 571, 153–173. [Google Scholar] [CrossRef] [PubMed]
- McLoone, P.; Simics, E.; Barton, A.; Norval, M.; Gibbs, N.K. An Action Spectrum for the Production of cis-Urocanic Acid in Human Skin In Vivo. J. Investig. Dermatol. 2005, 124, 1071–1074. [Google Scholar] [CrossRef]
- Beak, S.M.; Lee, Y.S.; Kim, J.A. NADPH Oxidase and Cyclooxygenase Mediate the Ultraviolet B-Induced Generation of Reactive Oxygen Species and Activation of Nuclear Factor-κB in HaCaT Human Keratinocytes. Biochimie 2004, 86, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Heck, D.E.; Vetrano, A.M.; Mariano, T.M.; Laskin, J.D. UVB Light Stimulates Production of Reactive Oxygen Species Unexpected Role for Catalase. J. Biol. Chem. 2003, 278, 22432–22436. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, P.; Haendeler, J.; Krutmann, J. The role of near infrared radiation in photoaging of the skin. Exp. Gerontol. 2008, 43, 629–632. [Google Scholar] [CrossRef]
- Schieke, S.M. Photoaging and Infrared Radiation. Novel Aspects of Molecular Mechanisms. Hautarzt 2003, 54, 822–824. [Google Scholar] [CrossRef]
- Schieke, S.M.; Schroeder, P.; Krutmann, J. Cutaneous effects of infrared radiation: From clinical observations to molecular response mechanisms. Photodermatol. Photoimmunol. Photomed. 2003, 19, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, P.; Pohl, C.; Calles, C.; Marks, C.; Wild, S.; Krutmann, J. Cellular Response to Infrared Radiation Involves Retrograde Mitochondrial Signaling. Free Radic. Biol. Med. 2007, 43, 128–135. [Google Scholar] [CrossRef]
- Krutmann, J.; Schroeder, P. Role of Mitochondria in Photoaging of Human Skin: The Defective Powerhouse Model. J. Investig. Dermatol. Symp. Proc. 2009, 14, 44–49. [Google Scholar] [CrossRef]
- De Gálvez, M.V.; Aguilera, J.; Sánchez-Roldán, C.; Herrera-Ceballos, E. Infrared radiation increases skin damage induced by other wavelengths in solar urticaria. Photodermatol. Photoimmunol. Photomed. 2016, 32, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- F’Guyer, S.; Afaq, F.; Mukhtar, H. Photochemoprevention of skin cancer by botanical agents. Photodermatol. Photoimmunol. Photomed. 2003, 19, 56–72. [Google Scholar] [CrossRef]
- Bowden, G.T. Prevention of non-melanoma skin cancer by targeting ultraviolet-B-light signalling. Nat. Rev. Cancer 2004, 4, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Sander, C.S.; Chang, H.; Salzmann, S.; Müller, C.S.; Ekanayake-Mudiyanselage, S.; Elsner, P.; Thiele, J.J. Photoaging is Associated with Protein Oxidation in Human Skin In Vivo. J. Investig. Dermatol. 2002, 118, 618–625. [Google Scholar] [CrossRef]
- Puzanowska-Tarasiewicz, H.; Kuźmicka, L.; Tarasiewicz, M. Organism defense against reactive oxygen species. Wiad. Lek. 2009, 62, 248–256. [Google Scholar] [PubMed]
- Kang, N.J.; Shin, S.H.; Lee, H.J.; Lee, K.W. Polyphenols as Small Molecular Inhibitors of Signaling Cascades in Carcinogenesis. Pharmacol. Ther. 2011, 130, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Potargowicz, E.; Szerszenowicz, E.; Staniszewska, M.; Nowak, D. Mitochondria as a Source of Reactive Oxygen Species. Postepy Hig. Med. Dosw. 2005, 59, 259–266. [Google Scholar]
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng Pharmacology–Multiple Constituents and Multiple Actions. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Rege, N.N.; Thatte, U.M.; Dahanukar, S.A. Adaptogenic Properties of Six Rasayana Herbs Used in Ayurvedic Medicine. Phytother. Res. 1999, 13, 275–291. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Helenurm, K. Floral Sex Ratios and Life History in Aralia Nudicaulis (Araliaceae). Evolution 1981, 35, 752–762. [Google Scholar] [CrossRef]
- Lacey, L. Micmac Medicines Remedies and Recollections; Nimbus Pub.: Hallifax, UK, 1993. [Google Scholar]
- Moerman, D.E. Native American Medicinal Plants: An Ethnobotanical Dictionary; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Wang, J.; Li, Q.; Ivanochko, G.; Huang, Y. Anticancer Effect of Extracts from a North American Medicinal Plant–Wild Sarsaparilla. Anticancer. Res. 2006, 26, 2157. [Google Scholar]
- Webster, D.; Lee, T.D.; Moore, J.; Manning, T.; Kunimoto, D.; Leblanc, D.; Johnson, J.A.; Gray, C.A. Antimycobacterial screening of traditional medicinal plants using the microplate resazurin assay. Can. J. Microbiol. 2010, 56, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.G.; Li, Q.Z.; Ivanochko, G.; Wang, R. Novel selective cytotoxicity of wild sarsaparilla rhizome extract. J. Pharm. Pharmacol. 2006, 58, 1399–1403. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; O’Neill, T.; Webster, D.; Johnson, J.A.; Gray, C.A. Anti-mycobacterial diynes from the Canadian medicinal plant Aralia nudicaulis. J. Ethnopharmacol. 2012, 140, 141–144. [Google Scholar] [CrossRef]
- Hyun, S.K.; Jung, H.A.; Min, B.S.; Jung, J.H. Isolation of Phenolics, Nucleosides, Saccharides and an Alkaloid from the Root of Aralia Cordata. Nat. Prod. Sci. 2010, 16, 20–25. [Google Scholar]
- Kim, M.H.; Kim, J.M.; Yoon, K.Y. Effects of blanching on antioxidant activity and total phenolic content according to type of medicinal plants. Food Sci. Biotechnol. 2013, 22, 817–823. [Google Scholar] [CrossRef]
- International Conference on Harmonization. Validation of Analytical Procedures: Text and Methodology Q2 (R1). In Proceedings of the International Conference on Harmonization of Technical Requirements for Registration of Pharmaceuticals for Human Use: ICH Harmonised Tripartite Guideline, Chicago, IL, USA, 7–10 November 2005. [Google Scholar]
- Shikov, A.N.; Pozharitskaya, O.; Makarov, V.G. Aralia elata var. mandshurica (Rupr. & Maxim.) J.Wen: An overview of pharmacological studies. Phytomedicine 2016, 23, 1409–1421. [Google Scholar]
- De Freitas, M.M.; Fontes, P.R.; Souza, P.M.; Fagg, C.W.; Guerra, E.N.S.; Nóbrega, Y.K.D.M.; Silveira, D.; Fonseca-Bazzo, Y.; Simeoni, L.A.; Homem-De-Mello, M.; et al. Extracts of Morus Nigra L. Leaves Standardized in Chlorogenic Acid, Rutin and Isoquercitrin: Tyrosinase Inhibition and Cytotoxicity. PLoS ONE 2016, 11, e0163130. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (Resazurin) Fluorescent Dye for the Assessment of Mammalian Cell Cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Girard-Lalancette, K.; Pichette, A.; Legault, J. Sensitive Cell-Based Assay Using Dcfh Oxidation for the Determination of Pro- and Antioxidant Properties of Compounds and Mixtures: Analysis of Fruit and Vegetable Juices. Food Chem. 2009, 115, 720–726. [Google Scholar] [CrossRef]
- Kim, S.-H.; Park, S.-Y.; Park, Y.-L.; Myung, D.-S.; Rew, J.-S.; Joo, Y.-E. Chlorogenic acid suppresses lipopolysaccharide-induced nitric oxide and interleukin-1β expression by inhibiting JAK2/STAT3 activation in RAW264.7 cells. Mol. Med. Rep. 2017, 16, 9224–9232. [Google Scholar] [CrossRef]
- Jiang, F.; Dusting, G.J. Natural phenolic compounds as cardiovascular therapeutics: Potential role of their antiinflammatory effects. Curr. Vasc. Pharmacol. 2003, 1, 135–156. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Bryant, C.; Lawson, M.F.; Chilvers, E.R.; Plevin, R. Dissociation of lipopolysaccharide-mediated induction of nitric oxide synthase and inhibition of DNA synthesis in RAW 264.7 macrophages and rat aortic smooth muscle cells. Br. J. Pharmacol. 1997, 120, 1439–1444. [Google Scholar] [CrossRef][Green Version]
- Prakash, V. Terpenoids as cytotoxic compounds: A perspective. Pharmacogn. Rev. 2018, 12, 166–176. [Google Scholar] [CrossRef]
- Miguel-Chávez, R. Phenolic Antioxidant Capacity: A Review of the State of the Art; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; Heredia, J.B.; Ambriz-Perez, D.L.; Leyva-López, N. Phenolic compounds: Natural alternative in inflammation treatment. A Review. Cogent Food Agric. 2016, 2, 1131412. [Google Scholar]
- Kim, J.S.; Kang, S.S.; Choi, J.S.; Lee, M.W.; Lee, T.S. Antioxident Components from Aralia Continentalis. Proc. Korean Soc. Appl. Pharmacol. 1998, 29, 13–17. [Google Scholar]
- Lingpeng, P. Effect of Aralia Echinocaulis Hand. Mazz Flavonoids on Osteoporosis Induced by Prednisone in Rats. Shizhen Guoyi Guoyao 2011, 22. [Google Scholar] [CrossRef]
- Han, B.H.; Woo, E.R.; Park, M.H.; Han, Y.N. Studies on the Anti-Inflammatory Activity Ofaralia Continentalis (I). Arch. Pharmacal Res. 1983, 6, 17. [Google Scholar] [CrossRef]
- Lee, J.H.; Ha, Y.W.; Jeong, C.S.; Kim, Y.S.; Park, Y. Isolation and tandem mass fragmentations of an anti-inflammatory compound from Aralia elata. Arch. Pharmacal Res. 2009, 32, 831–840. [Google Scholar] [CrossRef]
- Yang, J.; Kwak, C. Inhibitory Effect of Aralia Elata Ethanol Extract against Skin Damage in UVB-Exposed Human Keratinocytes and Human Dermal Fibroblasts. J. Nutr. Health 2016, 49, 429–436. [Google Scholar] [CrossRef]
- Cha, J.W.; Piao, M.J.; Kim, K.C.; Yao, C.W.; Zheng, J.; Kim, S.M.; Hyun, C.L.; Ahn, Y.S.; Hyun, J.W. The Polyphenol Chlorogenic Acid Attenuates UVB-mediated Oxidative Stress in Human HaCaT Keratinocytes. Biomol. Ther. 2014, 22, 136–142. [Google Scholar] [CrossRef]
- Balupillai, A.; Prasad, R.N.; Ramasamy, K.; Muthusamy, G.; Shanmugham, M.; Govindasamy, K.; Gunaseelan, S. Caffeic Acid Inhibits UVB-Induced Inflammation and Photocarcinogenesis through Activation of Peroxisome Proliferator-Activated Receptor-γ in Mouse Skin. Photochem. Photobiol. 2015, 91, 1458–1468. [Google Scholar] [CrossRef]
- Tian, S.; Nakamura, K.; Cui, T.; Kayahara, H. High-performance liquid chromatographic determination of phenolic compounds in rice. J. Chromatogr. A 2005, 1063, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.B.; Hibbs, J.B.; Taintor, R.R.; Krahenbuhl, J.L. Microbiostatic effect of murine-activated macrophages for Toxoplasma gondii. Role for synthesis of inorganic nitrogen oxides from l-arginine. J. Immunol. 1990, 144, 2725–2729. [Google Scholar]
- Lion, Q. Identification Et Quantification De Composés Biologiquement Actifs Des Rhizomes D’Aralia nudicaulis L. Master Thesis, Université du Québec à Chicoutimi, Chicoutimi, QC, Canada, 2019. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytes | Regression Equation | Linear Range (mg/mL) | Correlation Coefficient (R2) | LOD (mg/mL) | LOQ (mg/mL) |
|---|---|---|---|---|---|
| Protocatechuic acid | y = (13,871 ± 99.732)x − (22.992 ± 10.331) | 0.030–0.500 | 0.999 | 0.1 | 0.029 |
| Chlorogenic acid | y = (14,364 ± 103.94)x − (20.592 ± 11.008) | 0.050–0.500 | 0.999 | 0.014 | 0.043 |
| Caffeic acid | y = (26,397 ± 463.86)x − (7.5724 ± 23.462) | 0.030–0.500 | 0.999 | 0.009 | 0.027 |
| Compound | Intra-Day (RSD, n = 3) % | Inter-Day (RSD, n = 3) % | ||||
|---|---|---|---|---|---|---|
| 0.01 mg/mL | 0.05 mg/mL | 0.5 mg/mL | 0.01 mg/mL | 0.05 mg/mL | 0.5 mg/mL | |
| Protocatechuic acid | 0.40 | 0.41 | 0.24 | 0.70 | 0.62 | 0.20 |
| Chlorogenic acid | 0.81 | 1.4 | 1.3 | 0.60 | 0.81 | 0.81 |
| Caffeic acid | 0.64 | 1.1 | 2.1 | 0.43 | 1.3 | 0.54 |
| Methanolic Extract (μg/mg; w/w%) | Ethyl Acetate Extract (μg/mg; w/w%) | FEPC (μg/mg; w/w%) | |
|---|---|---|---|
| Protocatechuic acid | 4.1 ± 0.6 (0.4%) | 23.0 ± 0.7 (2.3%) | 48 ± 1 (4.8%) |
| Chlorogenic acid | 24.0 ± 0.3 (2.4%) | 25 ± 2 (2.5%) | 156 ± 1 (15.6%) |
| Caffeic acid | 2.80 ± 0.09 (0.28%) | 18.0 ± 0.2 (1.8%) | 153 ± 2 (15.3%) |
| Cytotoxicity IC50 (μg/mL) | ORAC (μmol Trolox/mg) | Antioxidant IC50 (μg/mL) | Anti-Inflammatory IC50 (μg/mL) | |
|---|---|---|---|---|
| Methanolic extract | 20 ± 1 | 0.09 ± 0.01 | 145 ± 48 | >160 |
| Ethyl acetate extract | 30 ± 5 | 5.7 ± 0.7 | 0.38 ± 0.04 | >160 |
| FEPC | 87 ± 6 | 12 ± 1 | 0.31 ± 0.03 | 35 ± 9 |
| Protocatechuic acid | Nd | 12 ± 1 | 0.13 ± 0.02 | 193 ± 58 |
| Chlorogenic acid | Nd | 11 ± 1 | 0.12 ± 0.01 | 89 ± 1 |
| Caffeic acid | Nd | 18 ± 1 | 0.13 ± 0.01 | 27 ± 14 |
| Mix of standards | Nd | 12 ± 2 | 0.46 ± 0.03 | >160 |
| Quercetin | Nd | 23 ± 3 | 0.27 ± 0.02 | Nd |
| l-NAME 250 μM | Nd | Nd | Nd | 75 ± 9% |
| l-NAME 1 mM | Nd | Nd | Nd | 89 ± 10% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lion, Q.; Pichette, A.; Mihoub, M.; Mshvildadze, V.; Legault, J. Phenolic Extract from Aralia nudicaulis L. Rhizomes Inhibits Cellular Oxidative Stresses. Molecules 2021, 26, 4458. https://doi.org/10.3390/molecules26154458
Lion Q, Pichette A, Mihoub M, Mshvildadze V, Legault J. Phenolic Extract from Aralia nudicaulis L. Rhizomes Inhibits Cellular Oxidative Stresses. Molecules. 2021; 26(15):4458. https://doi.org/10.3390/molecules26154458
Chicago/Turabian StyleLion, Quentin, Andre Pichette, Mouadh Mihoub, Vakhtang Mshvildadze, and Jean Legault. 2021. "Phenolic Extract from Aralia nudicaulis L. Rhizomes Inhibits Cellular Oxidative Stresses" Molecules 26, no. 15: 4458. https://doi.org/10.3390/molecules26154458
APA StyleLion, Q., Pichette, A., Mihoub, M., Mshvildadze, V., & Legault, J. (2021). Phenolic Extract from Aralia nudicaulis L. Rhizomes Inhibits Cellular Oxidative Stresses. Molecules, 26(15), 4458. https://doi.org/10.3390/molecules26154458