
Selective Inhibitors of the Inducible Nitric Oxide Synthase as Modulators of Cell Responses in LPS-Stimulated Human Monocytes

 ,
, 

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
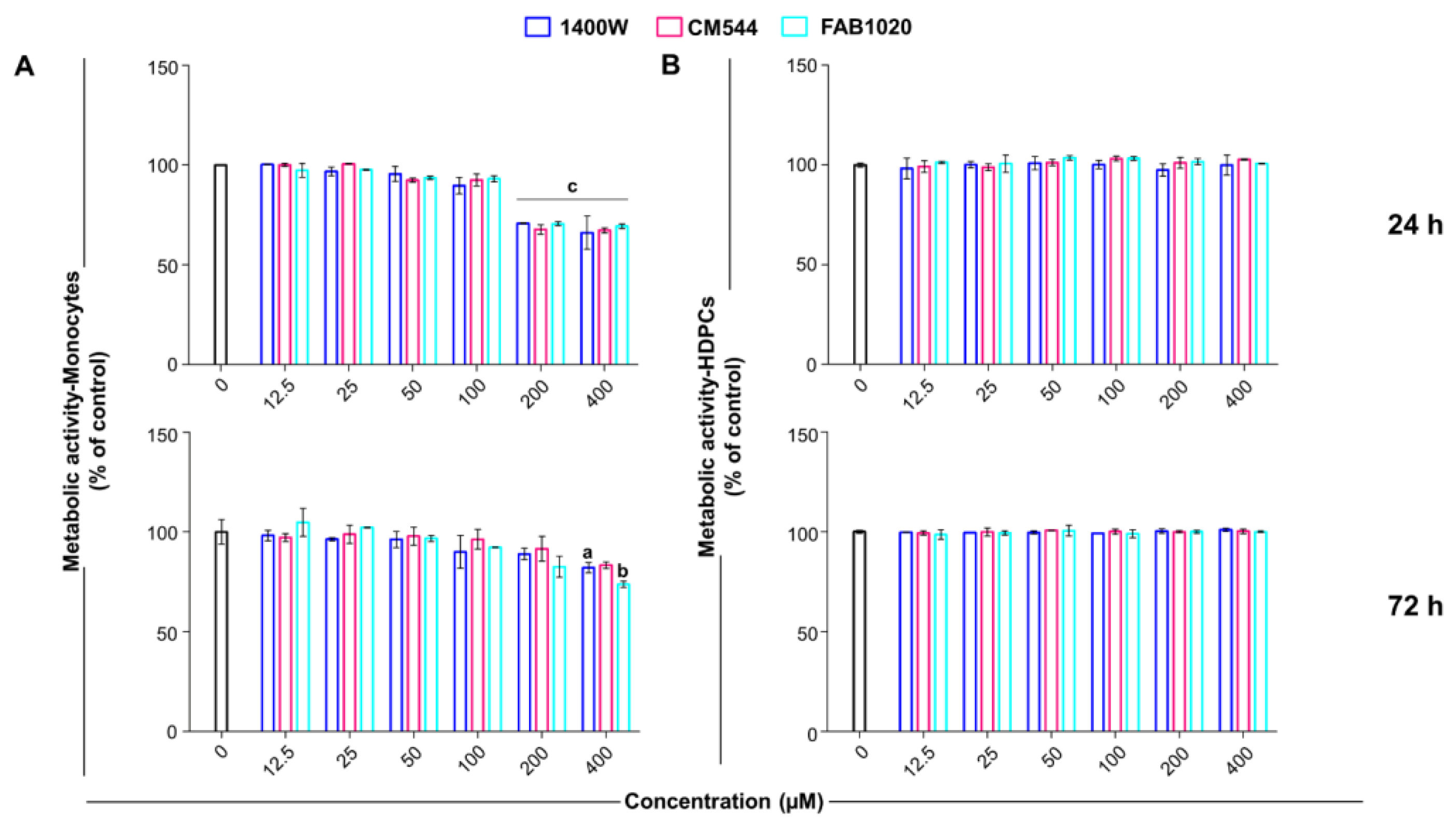
2.1. Cell Metabolic Activity and Selectivity of Compounds
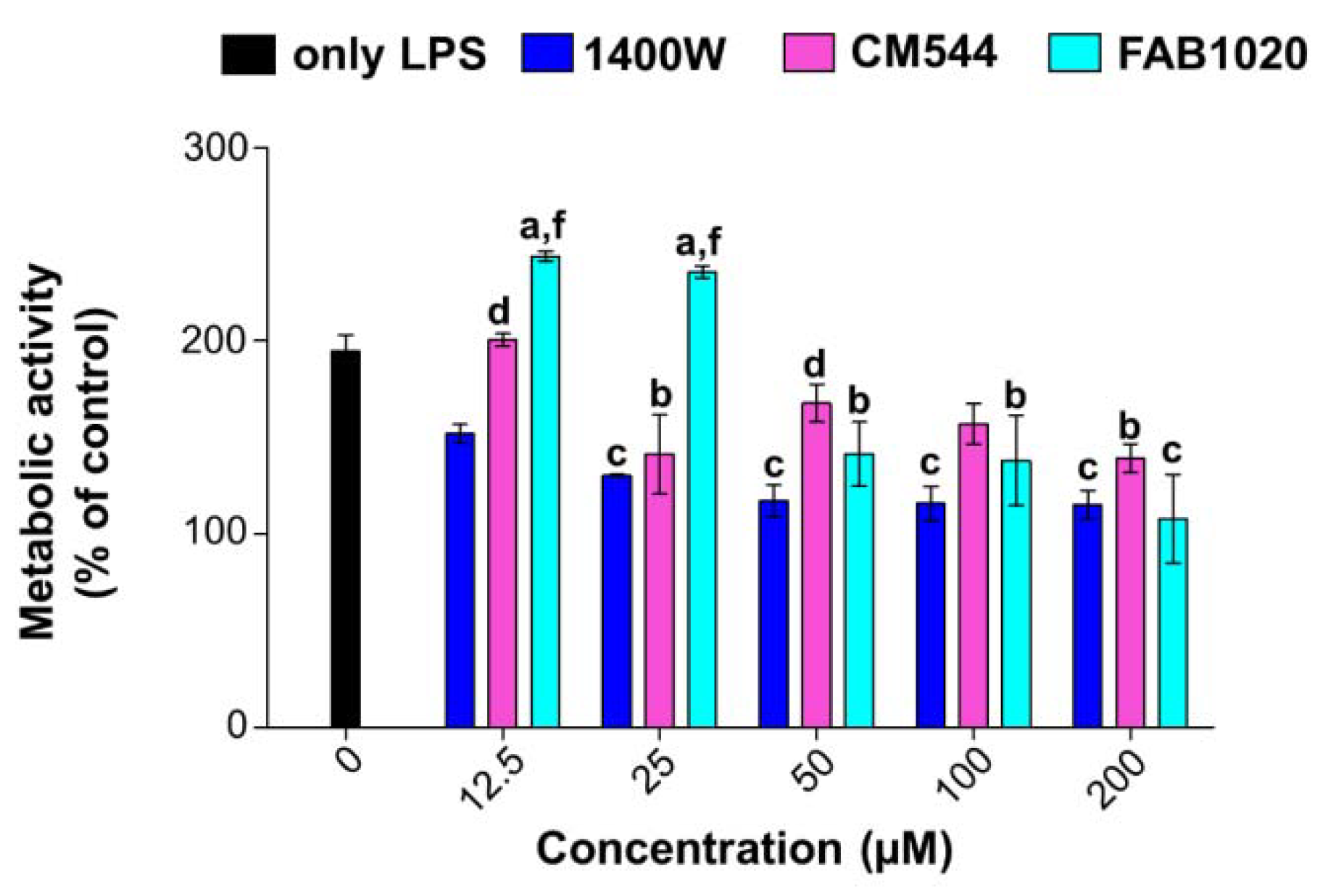
2.2. Cell Metabolic Activity in LPS-Stimulated Monocytes
2.3. Cytotoxicity Occurrence
2.4. IL-6 Secretion
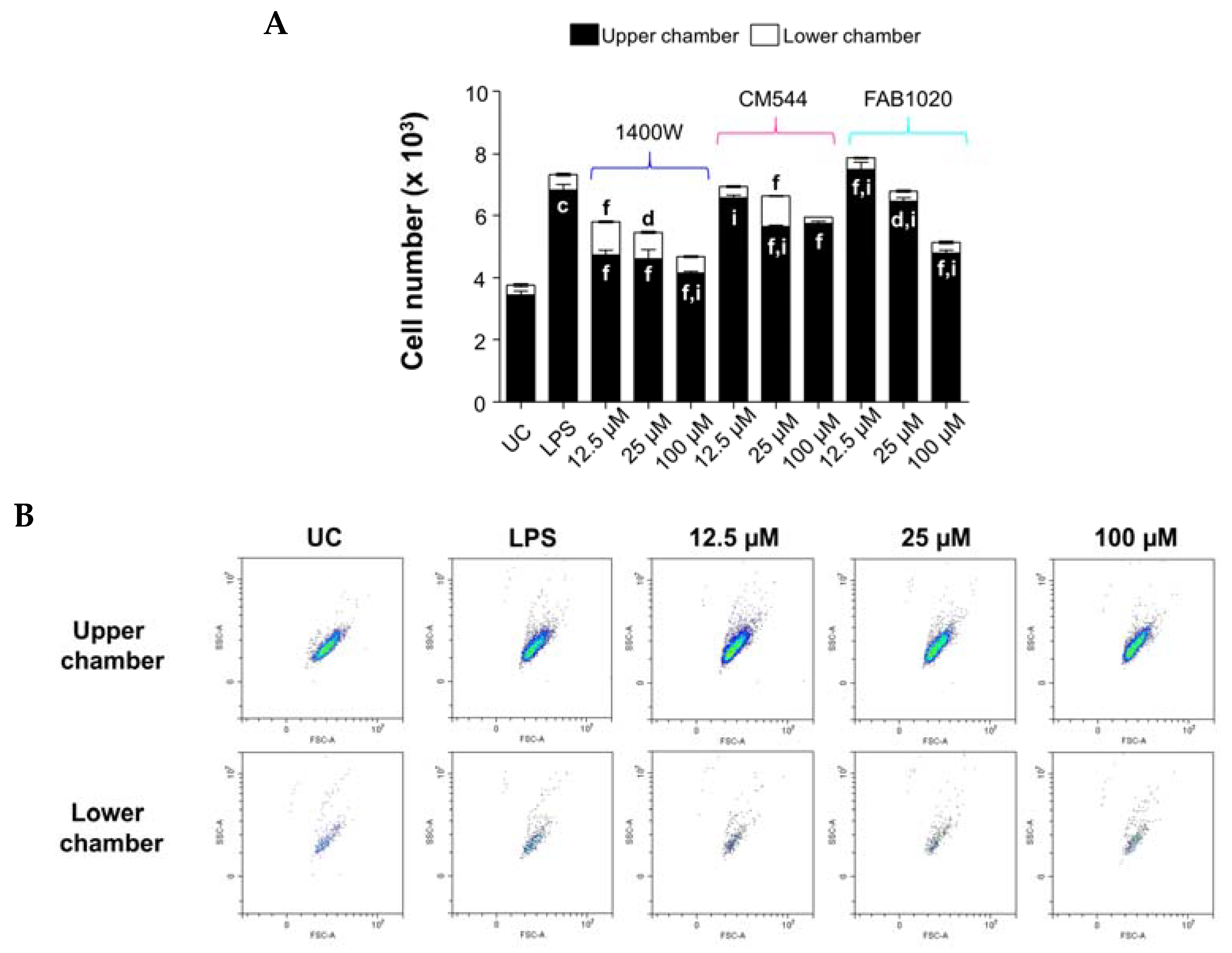
2.5. Counteraction of LPS-Stimulated Monocyte Migration in the Presence of iNOS Inhibitors
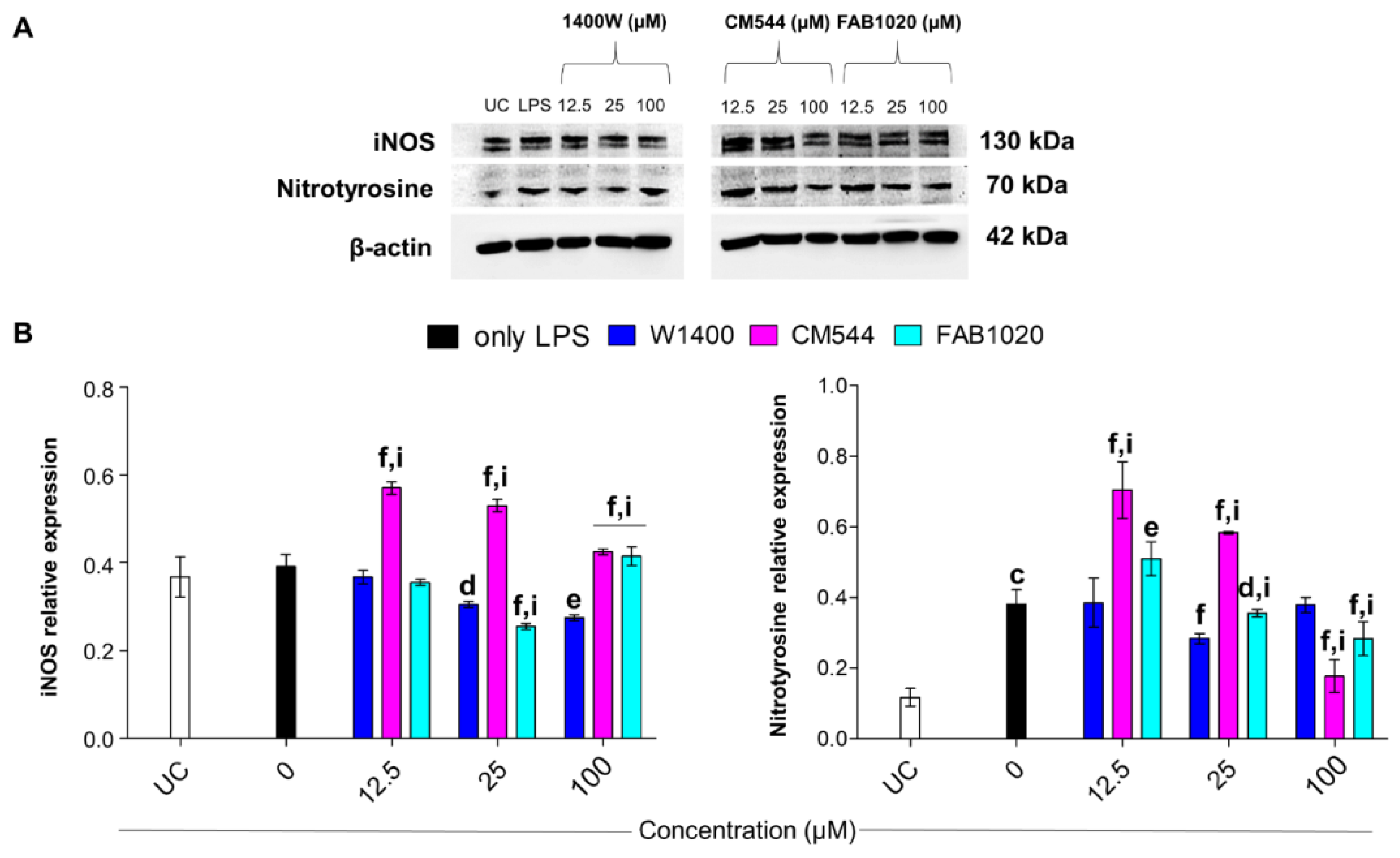
2.6. Modulation of iNOS and Protein Nitrosylation
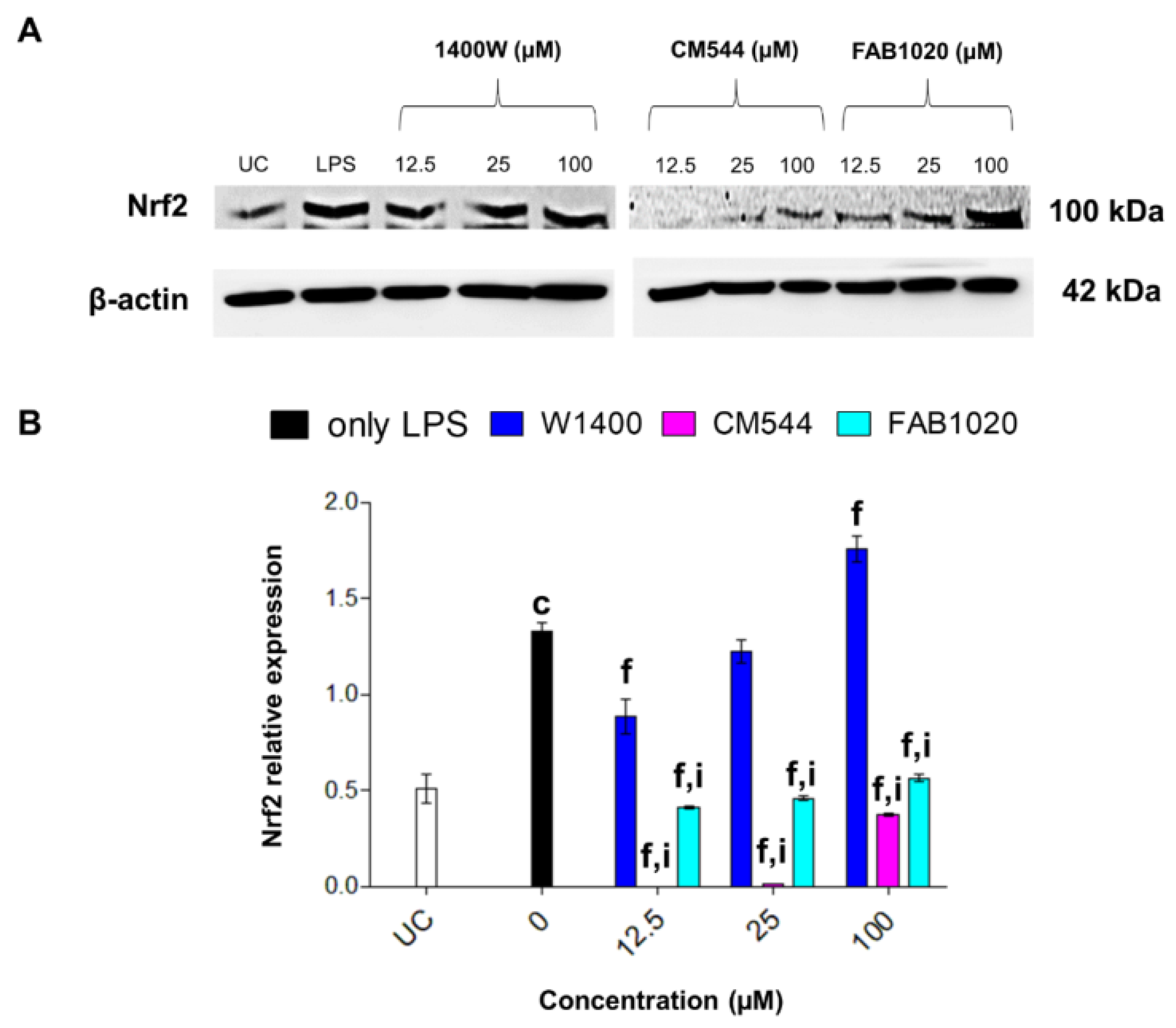
2.7. Expression of Nrf-2
3. Materials and Methods
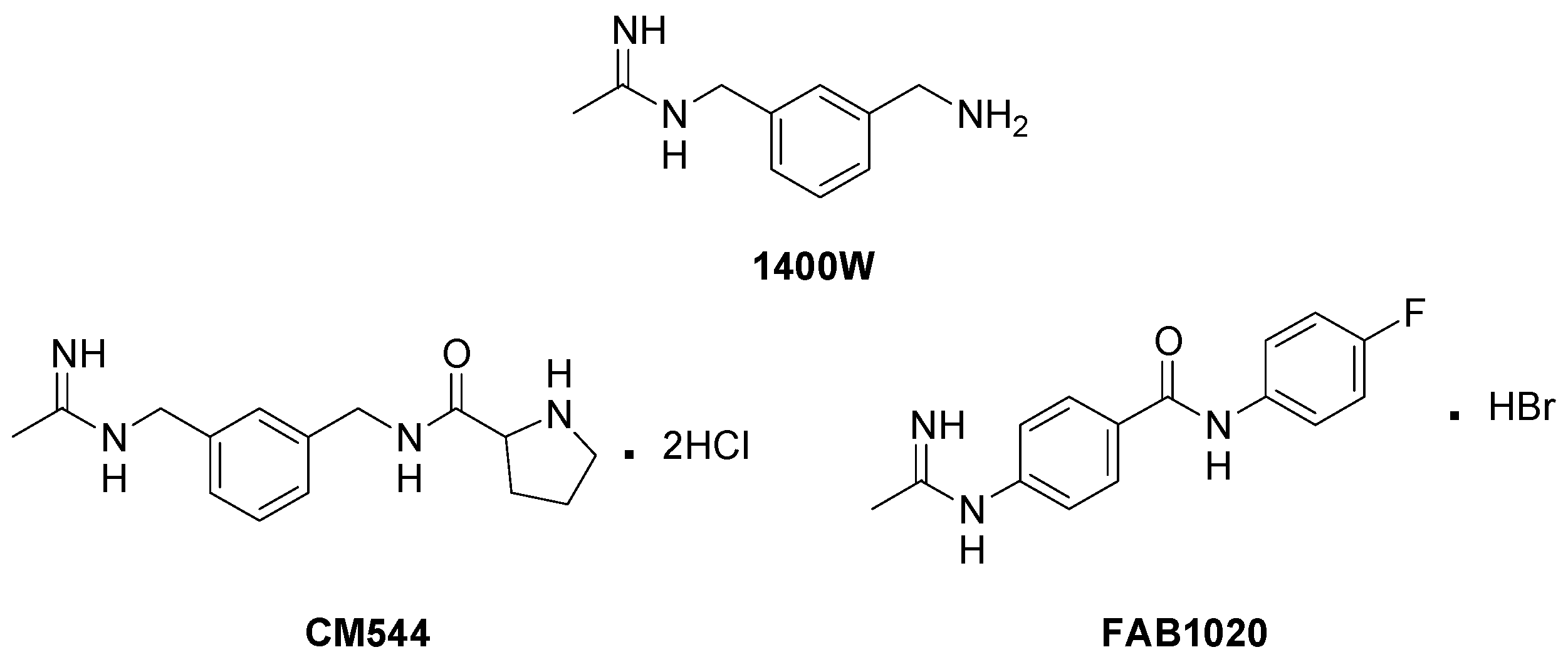
3.1. Synthesis of iNOS Inhibitors
3.2. Cell Cultures
3.3. Cell Metabolic Activity (MTS Test)
3.4. Cytotoxicity Assay
3.5. Interleukin-6 (IL-6) Release
3.6. Migration Assay
3.7. Protein Extraction and Immunoblotting
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Serbina, N.V.; Jia, T.; Hohl, T.M.; Pamer, E.G. Monocyte-mediated defense against microbial pathogens. Annu. Rev. Immunol. 2008, 26, 421–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, L.E.; Jakubzick, C.; Randolph, G.J. Regulation of the migration and survival of monocyte subsets by chemokine receptors and its relevance to atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1412–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izikson, L.; Klein, R.S.; Luster, A.D.; Weiner, H.L. Targeting monocyte recruitment in cns autoimmune disease. Clin. Immunol. 2002, 103, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Mokhdomi, T.A.; Bukhari, S.; Wani, Z.; Chikan, N.A.; Shah, B.A.; Koul, A.M.; Majeed, U.; Farooq, F.; Qadri, A.; et al. Lung cancer cell-derived EDA-containing fibronectin induces an inflammatory response from monocytes and promotes metastatic tumor microenvironment. J. Cell. Biochem. 2021, 122, 562–576. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. Colony-stimulating factor in inflammation and autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Leibovich, S.J.; Polverini, P.J.; Fong, T.W.; Harlow, L.A.; Koch, A.E. Production of angiogenic activity by human monocytes requires an L-arginine/nitric oxide-synthase-dependent effector mechanism. Proc. Natl. Acad. Sci. USA 1994, 91, 4190–4194. [Google Scholar] [CrossRef] [Green Version]
- Al-Shehri, S.S. Reactive oxygen and nitrogen species and innate immune response. Biochimie 2021, 181, 52–64. [Google Scholar] [CrossRef]
- Burney, S.; Caulfield, J.L.; Niles, J.C.; Wishnok, J.S.; Tannenbaum, S.R. The chemistry of DNA damage from nitric oxide and peroxynitrite. Mutat. Res. 1999, 424, 37–49. [Google Scholar] [CrossRef]
- Grimm, E.A.; Sikora, A.G.; Ekmekcioglu, S. Molecular pathways: Inflammation-associated nitric-oxide production as a cancer-supporting redox mechanism and a potential therapeutic target. Clin. Cancer Res. 2013, 19, 5557–5563. [Google Scholar] [CrossRef] [Green Version]
- Ekmekcioglu, S.; Grimm, E.A.; Roszik, J. Targeting iNOS to increase efficacy of immunotherapies. Hum. Vaccin. Immunother. 2017, 13, 1105–1108. [Google Scholar] [CrossRef] [Green Version]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible nitric oxide synthase and inflammatory diseases. Mol. Med. 2000, 6, 347–373. [Google Scholar] [CrossRef] [Green Version]
- Minhas, R.; Bansal, Y.; Bansal, G. Inducible nitric oxide synthase inhibitors: A comprehensive update. Med. Res. Rev. 2020, 40, 823–855. [Google Scholar] [CrossRef]
- Re, N.; Fantacuzzi, M.; Maccallini, C.; Paciotti, R.; Amoroso, R. Recent developments of amidine-like compounds as selective NOS inhibitors. Curr. Enzym. Inhib. 2016, 12, 30–39. [Google Scholar] [CrossRef]
- Maccallini, C.; Di Matteo, M.; Gallorini, M.; Montagnani, M.; Graziani, V.; Ammazzalorso, A.; Amoia, P.; De Filippis, B.; Di Silvestre, S.; Fantacuzzi, M.; et al. Discovery of N-{3-[(ethanimidoylamino)methyl]benzyl}-l-prolinamide dihydrochloride: A new potent and selective inhibitor of the inducible nitric oxide synthase as a promising agent for the therapy of malignant glioma. Eur. J. Med. Chem. 2018, 152, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Gallorini, M.; Maccallini, C.; Ammazzalorso, A.; Amoia, P.; De Filippis, B.; Fantacuzzi, M.; Giampietro, L.; Cataldi, A.; Amoroso, R. The selective acetamidine-based iNOS inhibitor CM544 reduces glioma cell proliferation by enhancing PARP-1 cleavage in vitro. Int. J. Mol. Sci. 2019, 20, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccallini, C.; Arias, F.; Gallorini, M.; Amoia, P.; Ammazzalorso, A.; De Filippis, B.; Fantacuzzi, M.; Giampietro, L.; Cataldi, A.; Camacho, M.E.; et al. Antiglioma activity of aryl and amido-aryl acetamidine derivatives targeting iNOS: Synthesis and biological evaluation. ACS Med. Chem. Lett. 2020, 11, 1470–1475. [Google Scholar] [CrossRef] [PubMed]
- Garvey, E.P.; Oplinger, J.A.; Furfine, E.S.; Kiff, R.J.; Laszlo, F.; Whittle, B.J.; Knowles, R.G. 1400 W is a slow, tight binding, and highly selective inhibitor of inducible nitric-oxide synthase in vitro and in vivo. J. Biol. Chem. 1997, 272, 4959–4963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccallini, C.; Patruno, A.; Besker, N.; Alì, J.I.; Ammazzalorso, A.; De Filippis, B.; Franceschelli, S.; Giampietro, L.; Pesce, M.; Reale, M.; et al. Synthesis, biological evaluation, and docking studies of N-substituted acetamidines as selective inhibitors of inducible nitric oxide synthase. J. Med. Chem. 2009, 52, 1481–1485. [Google Scholar] [CrossRef]
- Fantacuzzi, M.; Maccallini, C.; Lannutti, F.; Patruno, A.; Masella, S.; Pesce, M.; Speranza, L.; Ammazzalorso, A.; De Filippis, B.; Giampietro, L.; et al. Selective inhibition of iNOS by benzyl- and dibenzyl derivatives of N-(3-aminobenzyl)acetamidine. ChemMedChem 2011, 6, 1203–1206. [Google Scholar] [CrossRef]
- Grottelli, S.; Amoroso, R.; Macchioni, L.; D’Onofrio, F.; Fettucciari, K.; Bellezza, I.; Maccallini, C. Acetamidine-based iNOS inhibitors as molecular tools to counteract inflammation in BV2 microglial cells. Molecules 2020, 25, 2646. [Google Scholar] [CrossRef]
- Yasuhara, R.; Suzawa, T.; Miyamoto, Y.; Wang, X.; Takami, M.; Yamada, A.; Kamijo, R. Nitric oxide in pulp cell growth, differentiation, and mineralization. J. Dent. Res. 2007, 86, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Takashiba, S.; Van Dyke, T.E.; Amar, S.; Murayama, Y.; Soskolne, A.W.; Shapira, L. Differentiation of monocytes to macrophages primes cells for lipopolysaccharide stimulation via accumulation of cytoplasmic nuclear factor kB. Infect. Immun. 1999, 67, 5573–5578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. LPS-induced cytokine production in human monocytes and macrophages. Crit. Rev. Immunol. 2011, 31, 379–446. [Google Scholar] [CrossRef]
- Saluja, R.; Jyoti, A.; Chatterjee, M.; Habib, S.; Verma, A.; Mitra, K.; Barthwal, M.K.; Bajpai, V.K.; Dikshit, M. Molecular and biochemical characterization of nitric oxide synthase isoforms and their intracellular distribution in human peripheral blood mononuclear cells. Biochim. Biophys. Acta 2011, 1813, 1700–1707. [Google Scholar] [CrossRef] [Green Version]
- Ulmer, A.J.; Flad, H.; Rietschel, T.; Mattern, T. Induction of proliferation and cytokine production in human T lymphocytes by lipopolysaccharide (LPS). Toxicology 2000, 152, 37–45. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Masuda, K.; Kishimoto, T. Regulation of IL-6 in immunity and diseases. Adv. Exp. Med. Biol. 2016, 941, 79–88. [Google Scholar] [PubMed]
- Schweikl, H.; Birke, M.; Gallorini, M.; Petzel, C.; Bolay, C.; Waha, C.; Hiller, K.A.; Buchalla, W. HEMA-induced oxidative stress inhibits NF-κB nuclear translocation and TNF release from LTA- and LPS-stimulated immunocompetent cells. Dent. Mater. 2021, 37, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.J.; Liu, J.X.; Xie, Y.; Luo, P.; Liu, Z.Q.; Liu, L.; Zhou, H. Suppression of macrophage migration by down-regulating Src/FAK/P130Cas activation contributed to the anti-inflammatory activity of sinomenine. Pharmacol. Res. 2021, 167, 105513. [Google Scholar] [CrossRef]
- Ivan, D.C.; Walthert, S.; Locatelli, G. Central nervous system barriers impact distribution and expression of iNOS and arginase-1 in infiltrating macrophages during neuroinflammation. Front. Immunol. 2021, 12, 666961. [Google Scholar] [CrossRef]
- Dasgupta, S.; Gomez, J.J.; Singh, I.; Khan, M. S-Nitrosylation in regulation of inflammation and cell damage. Curr. Drug Targets 2018, 19, 1831–1838. [Google Scholar] [CrossRef]
- Schweikl, H.; Gallorini, M.; Pöschl, G.; Urmann, V.; Petzel, C.; Bolay, C.; Hiller, K.A.; Cataldi, A.; Buchalla, W. Functions of transcription factors NF-κB and Nrf2 in the inhibition of LPS-stimulated cytokine release by the resin monomer HEMA. Dent. Mater. 2018, 34, 1661–1678. [Google Scholar] [CrossRef] [PubMed]
- Maccallini, C.; Di Matteo, M.; Ammazzalorso, A.; D’Angelo, A.; De Filippis, B.; Di Silvestre, S.; Fantacuzzi, M.; Giampietro, L.; Pandolfi, A.; Amoroso, R. Reversed-phase high-performance liquid chromatography method with fluorescence detection to screen nitric oxide synthases inhibitors. J. Sep. Sci. 2014, 37, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Sancilio, S.; Gallorini, M.; Di Nisio, C.; Marsich, E.; Di Pietro, R.; Schweikl, H.; Cataldi, A. Alginate/hydroxyapatite-based nanocomposite scaffolds for bone tissue engineering improve dental pulp biomineralization and differentiation. Stem Cells Int. 2018, 2018, 9643721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cataldi, A.; Gallorini, M.; Di Giulio, M.; Guarnieri, S.; Mariggiò, M.A.; Traini, T.; Di Pietro, R.; Cellini, L.; Marsich, E.; Sancilio, S. Adhesion of human gingival fibroblasts/Streptococcus mitis co-culture on the nanocomposite system Chitlac-nAg. J. Mater. Sci. Mater. Med. 2016, 27, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancilio, S.; Marsich, E.; Schweikl, H.; Cataldi, A.; Gallorini, M. Redox control of IL-6-mediated dental pulp stem-cell differentiation on alginate/hydroxyapatite biocomposites for bone ingrowth. Nanomaterials 2019, 9, 1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zara, S.; De Colli, M.; di Giacomo, V.; Zizzari, V.L.; Di Nisio, C.; Di Tore, U.; Salini, V.; Gallorini, M.; Tetè, S.; Cataldi, A. Zoledronic acid at subtoxic dose extends osteoblastic stage span of primary human osteoblasts. Clin. Oral Investig. 2015, 19, 601–611. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallorini, M.; Rapino, M.; Schweikl, H.; Cataldi, A.; Amoroso, R.; Maccallini, C. Selective Inhibitors of the Inducible Nitric Oxide Synthase as Modulators of Cell Responses in LPS-Stimulated Human Monocytes. Molecules 2021, 26, 4419. https://doi.org/10.3390/molecules26154419
Gallorini M, Rapino M, Schweikl H, Cataldi A, Amoroso R, Maccallini C. Selective Inhibitors of the Inducible Nitric Oxide Synthase as Modulators of Cell Responses in LPS-Stimulated Human Monocytes. Molecules. 2021; 26(15):4419. https://doi.org/10.3390/molecules26154419
Chicago/Turabian StyleGallorini, Marialucia, Monica Rapino, Helmut Schweikl, Amelia Cataldi, Rosa Amoroso, and Cristina Maccallini. 2021. "Selective Inhibitors of the Inducible Nitric Oxide Synthase as Modulators of Cell Responses in LPS-Stimulated Human Monocytes" Molecules 26, no. 15: 4419. https://doi.org/10.3390/molecules26154419
APA StyleGallorini, M., Rapino, M., Schweikl, H., Cataldi, A., Amoroso, R., & Maccallini, C. (2021). Selective Inhibitors of the Inducible Nitric Oxide Synthase as Modulators of Cell Responses in LPS-Stimulated Human Monocytes. Molecules, 26(15), 4419. https://doi.org/10.3390/molecules26154419